

Acyl-Carnitines Exert Positive Effects on Mitochondrial Activity under Oxidative Stress in Mouse Oocytes: A Potential Mechanism Underlying Carnitine Efficacy on PCOS
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Oocyte Collection and Treatments
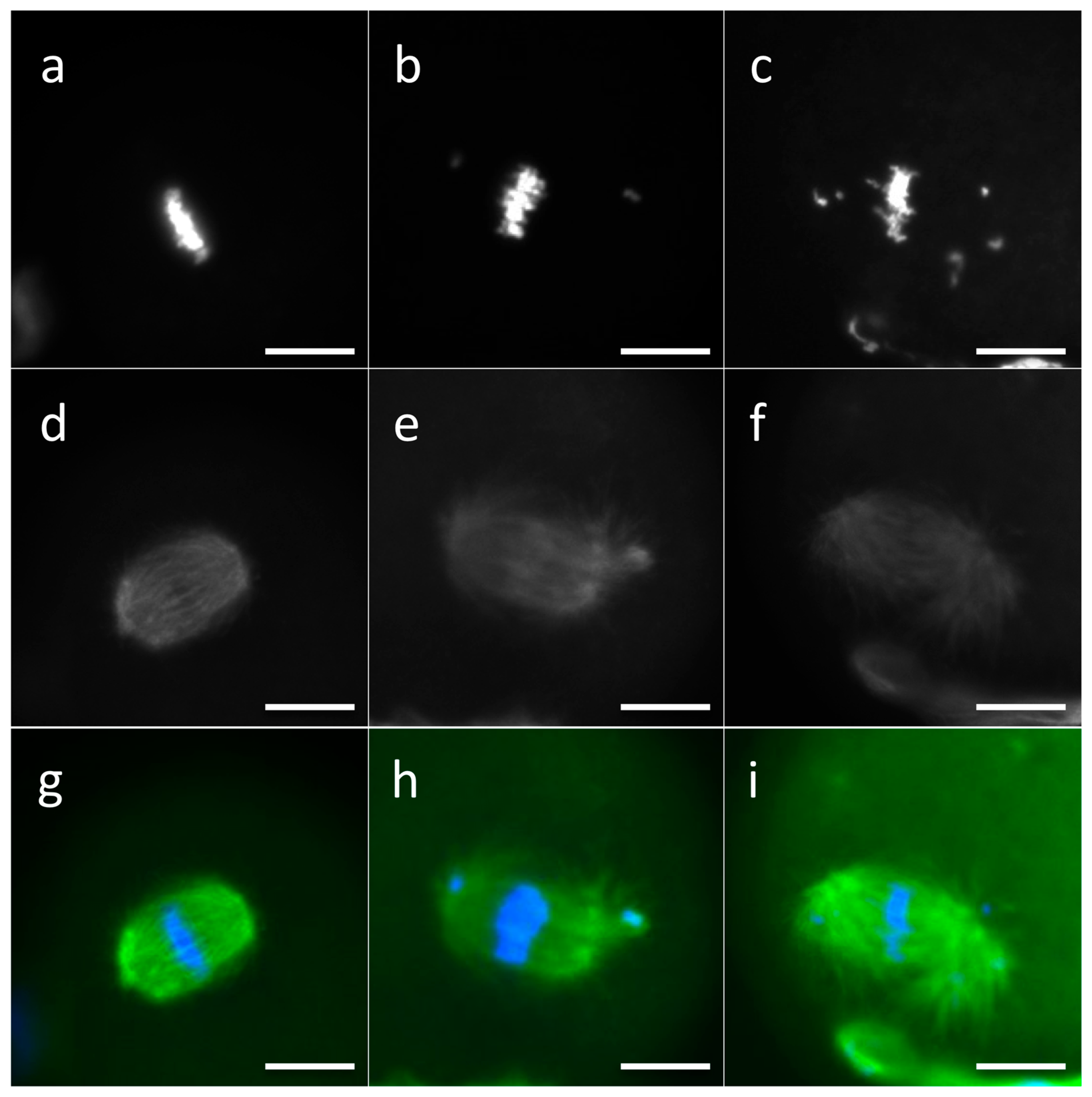
2.3. Analysis of DNA Distribution and Spindle Configuration of In Vitro Matured MII Oocytes
2.4. RNA Extraction and Real-Time Reverse Transcriptase-Polymerase Chain Reaction Analysis
2.5. Application of Seahorse XFp to Measure Oxygen Consumption in Oocytes
2.6. Analysis of ATP Production
2.7. Statistical Analysis
3. Results
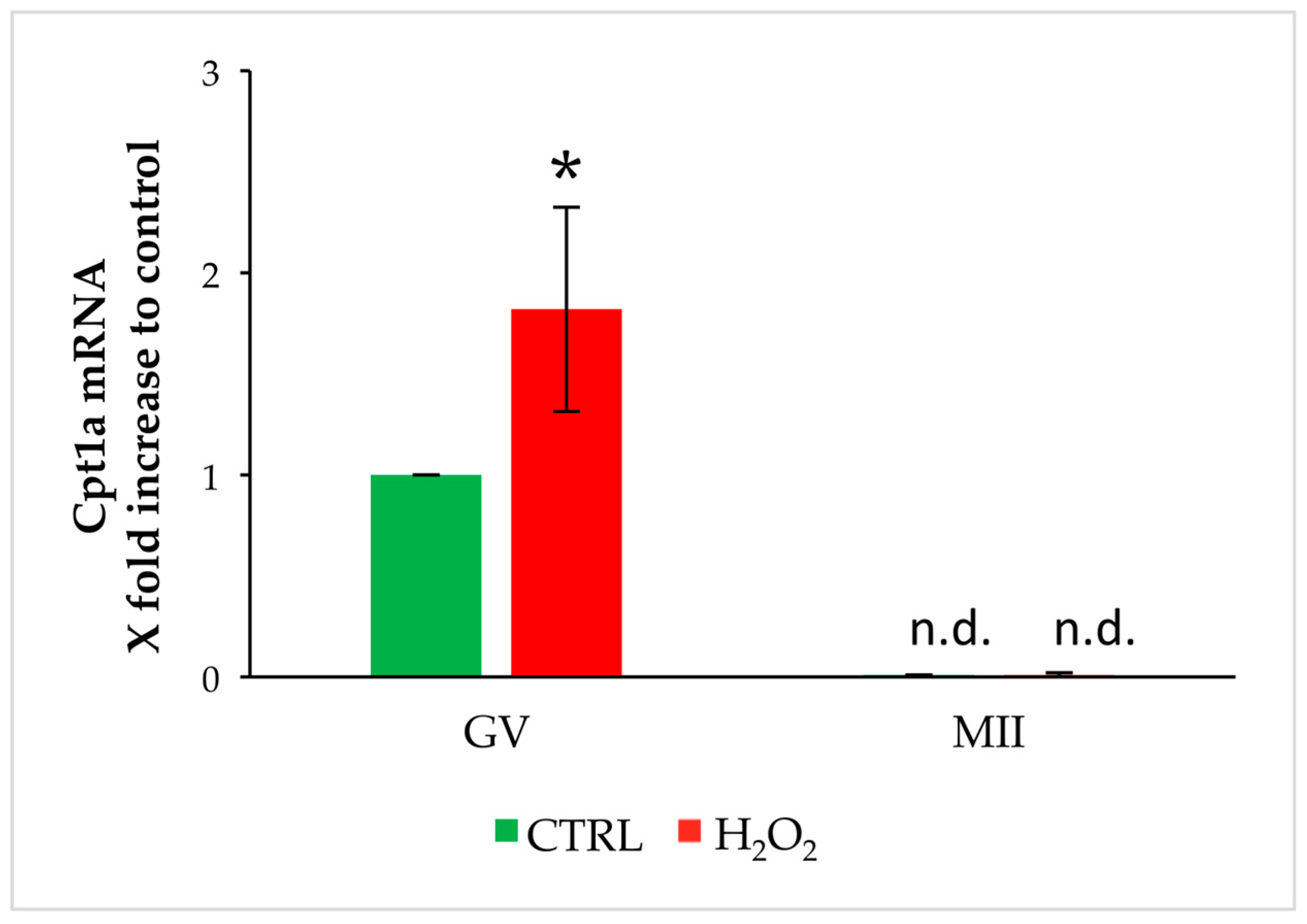
3.1. Analysis of Cpt1a and Cpt1c Expression in Oocytes Exposed to Oxidative Stress
3.2. Effects of L-Carnitine and Acetyl-L-Carnitine on In Vitro Maturation and MII Configuration of Oocytes Exposed to Oxidative Stress
3.3. Effects of Propionyl-L-Carnitine on In Vitro Maturation and MII Configuration of Oocytes Exposed to Oxidative Stress
3.4. Effects of Medium Supplementation with a Mixture of L-Carnitine, Acetyl-L-Carnitine, and Propionyl-L-Carnitine on In Vitro Maturation and MII Configuration of Oocytes Exposed to Oxidative Stress
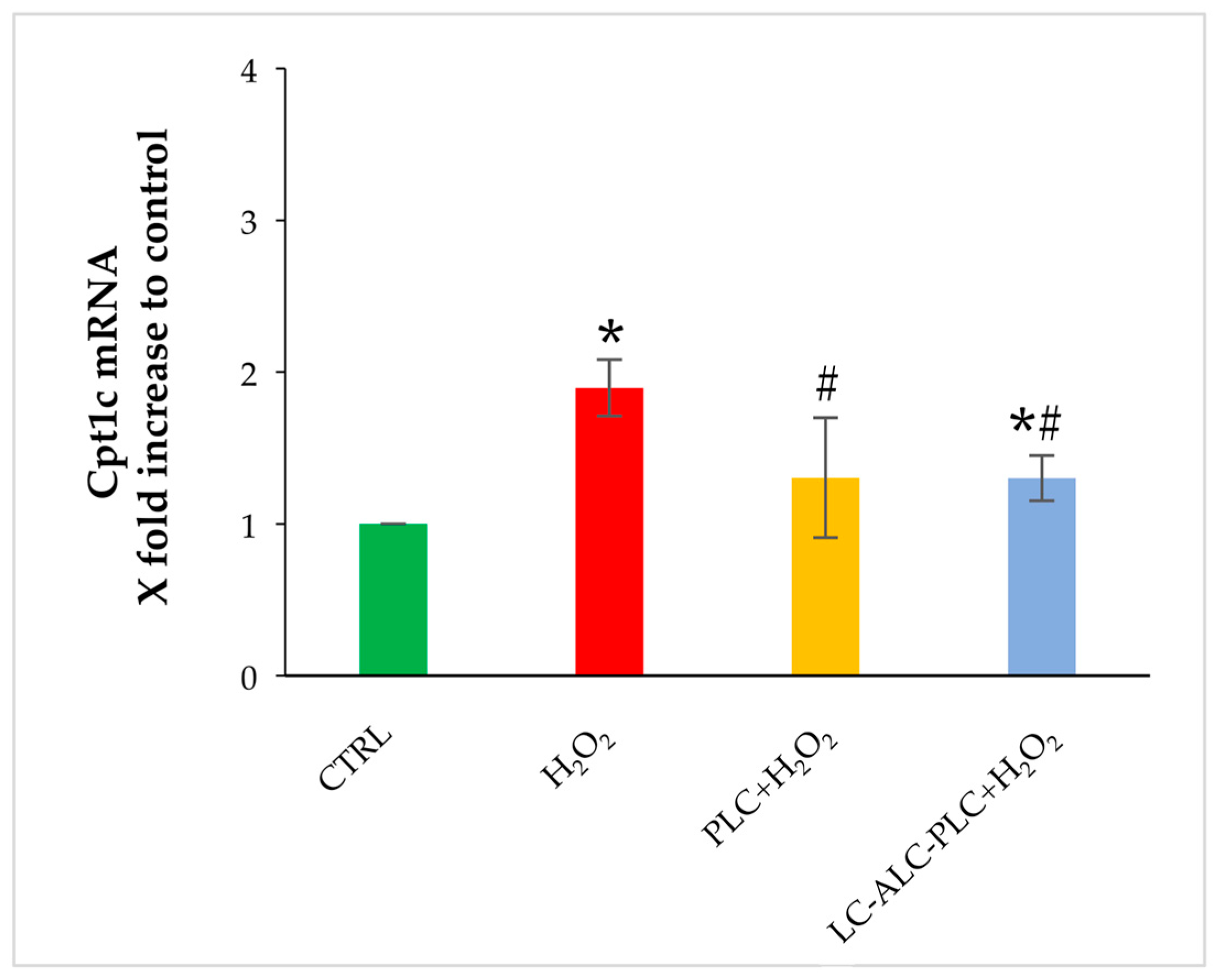
3.5. Effects of Carnitines on Cpt1a and Cpt1c Gene Expression in Oocytes Exposed to Oxidative Stress
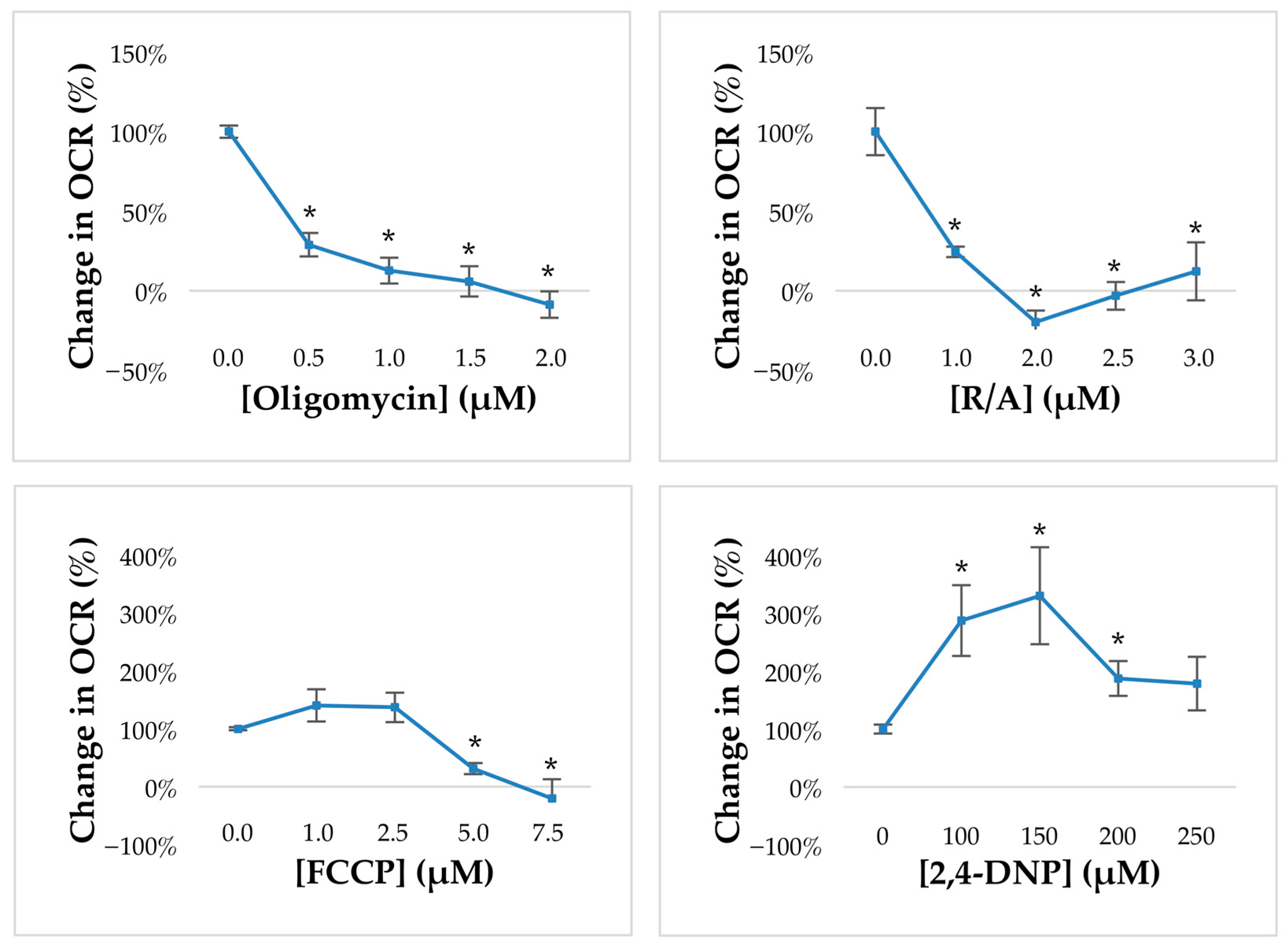
3.6. Establishment of Mitochondrial Inhibitors for Measurements of Oxygen Consumption Rate
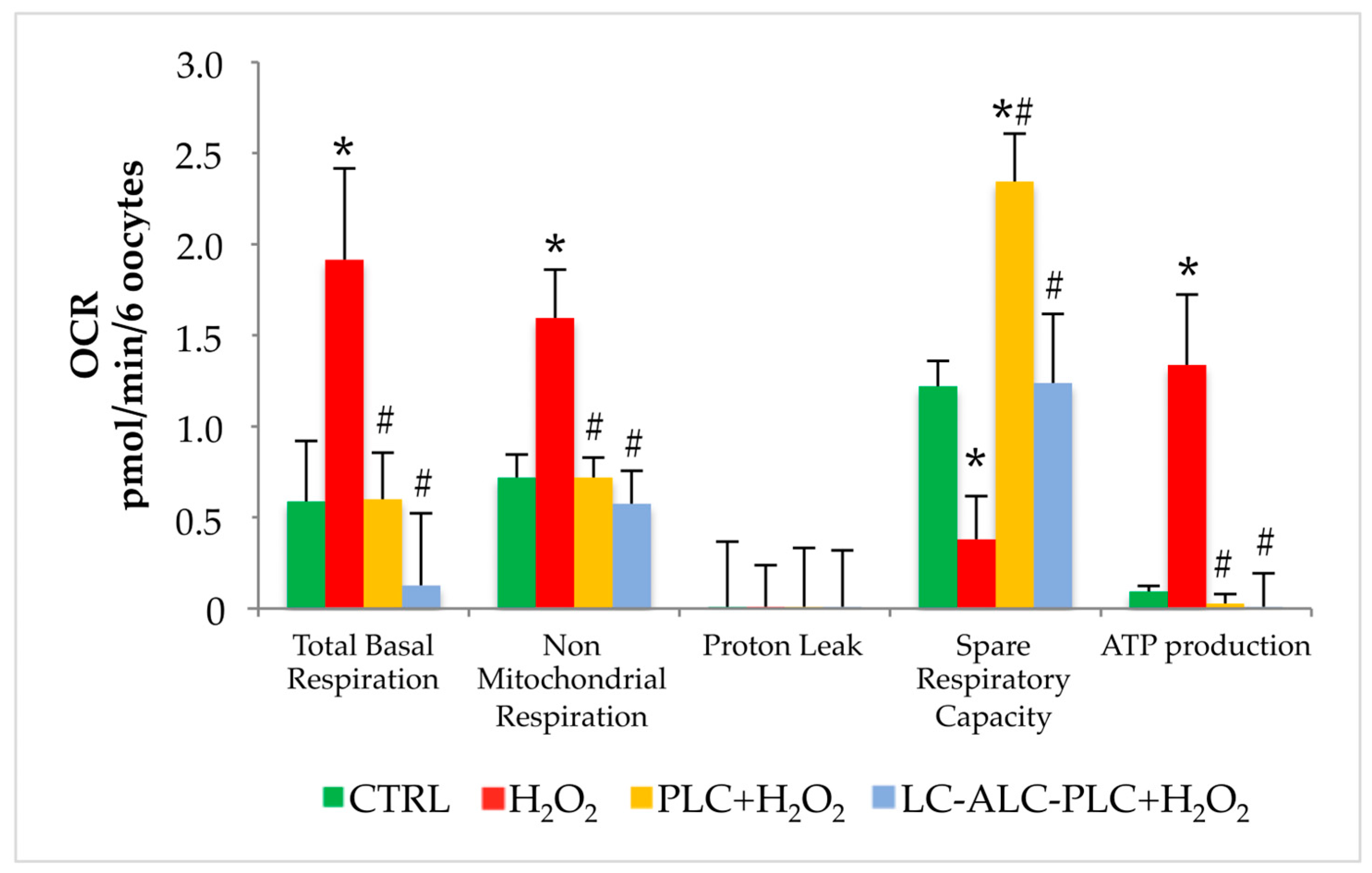
3.7. Effect of PLC or Mixture of L-Carnitine, Acetyl-L-Carnitine, and Propionyl-L-Carnitine on Mitochondrial Activity of GV Oocytes Exposed to Oxidative Stress
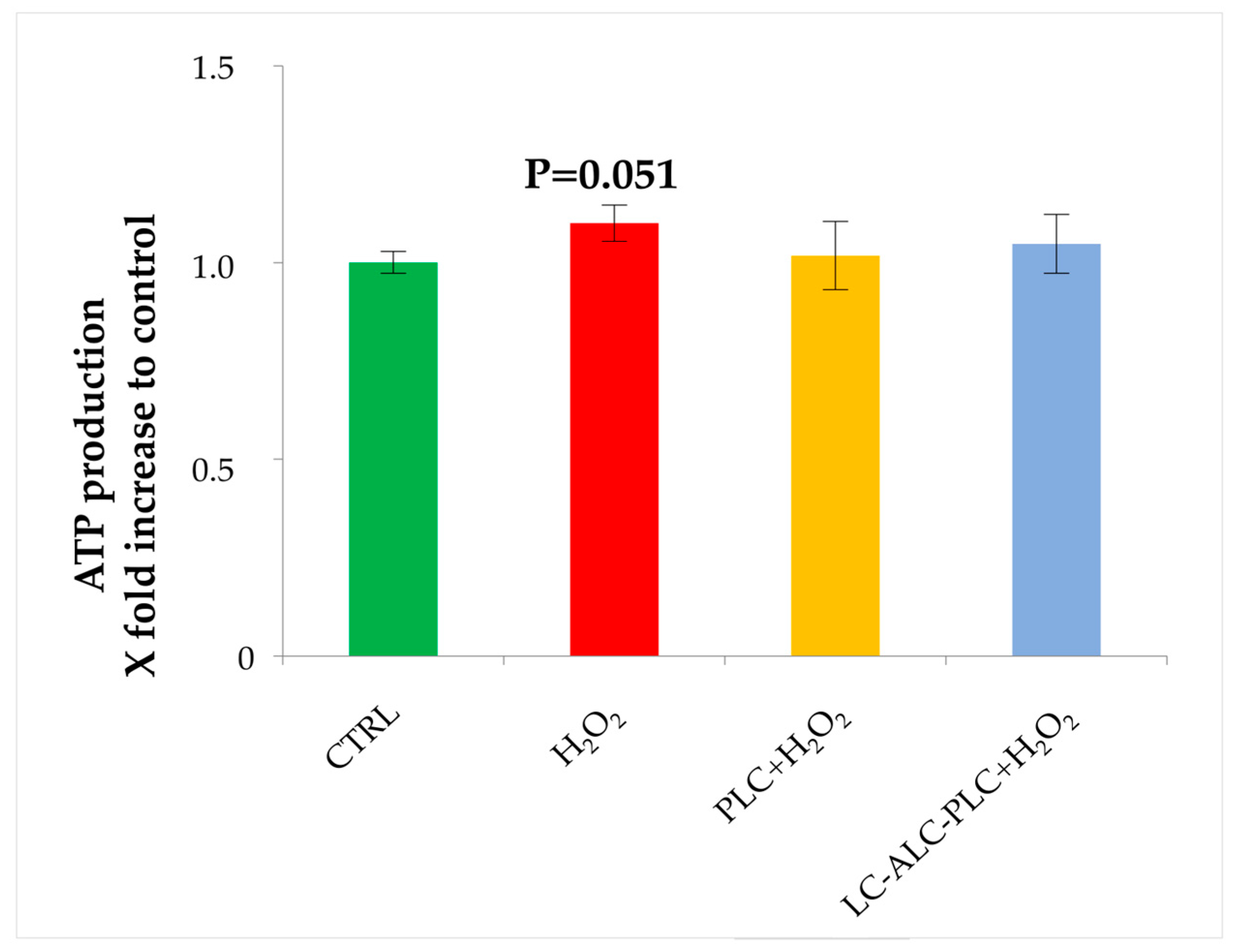
3.8. Analysis of ATP Production
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reuter, S.E.; Evans, A.M. Carnitine and Acylcarnitines. Clin. Pharmacokinet. 2012, 51, 553–572. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine Transport and Fatty Acid Oxidation. Biochim. Biophys. Acta 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Dahash, B.A.; Sankararaman, S. Carnitine Deficiency. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- McCann, M.R.; George De la Rosa, M.V.; Rosania, G.R.; Stringer, K.A. L-Carnitine and Acylcarnitines: Mitochondrial Biomarkers for Precision Medicine. Metabolites 2021, 11, 51. [Google Scholar] [CrossRef]
- Dunning, K.R.; Akison, L.K.; Russell, D.L.; Norman, R.J.; Robker, R.L. Increased Beta-Oxidation and Improved Oocyte Developmental Competence in Response to l-Carnitine during Ovarian in Vitro Follicle Development in Mice. Biol. Reprod. 2011, 85, 548–555. [Google Scholar] [CrossRef]
- Zare, Z.; Abouhamzeh, B.; Masteri Farahani, R.; Salehi, M.; Mohammadi, M. Supplementation of L-Carnitine during in Vitro Maturation of Mouse Oocytes Affects Expression of Genes Involved in Oocyte and Embryo Competence: An Experimental Study. Int. J. Reprod. Biomed. 2017, 15, 779–786. [Google Scholar] [CrossRef]
- Gentile, L.; Monti, M.; Sebastiano, V.; Merico, V.; Nicolai, R.; Calvani, M.; Garagna, S.; Redi, C.A.; Zuccotti, M. Single-Cell Quantitative RT-PCR Analysis of Cpt1b and Cpt2 Gene Expression in Mouse Antral Oocytes and in Preimplantation Embryos. Cytogenet. Genome Res. 2004, 105, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Di Emidio, G.; Rea, F.; Placidi, M.; Rossi, G.; Cocciolone, D.; Virmani, A.; Macchiarelli, G.; Palmerini, M.G.; D’Alessandro, A.M.; Artini, P.G.; et al. Regulatory Functions of L-Carnitine, Acetyl, and Propionyl L-Carnitine in a PCOS Mouse Model: Focus on Antioxidant/Antiglycative Molecular Pathways in the Ovarian Microenvironment. Antioxidants 2020, 9, 867. [Google Scholar] [CrossRef]
- Trikudanathan, S. Polycystic Ovarian Syndrome. Med. Clin. N. Am. 2015, 99, 221–235. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.E.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic Ovary Syndrome. Nat. Rev. Dis. Primer 2016, 2, 16057. [Google Scholar] [CrossRef] [PubMed]
- Sirmans, S.; Pate, K. Epidemiology, Diagnosis, and Management of Polycystic Ovary Syndrome. Clin. Epidemiol. 2014, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fenkci, S.M.; Fenkci, V.; Oztekin, O.; Rota, S.; Karagenc, N. Serum Total L-Carnitine Levels in Non-Obese Women with Polycystic Ovary Syndrome. Hum. Reprod. 2008, 23, 1602–1606. [Google Scholar] [CrossRef] [PubMed]
- Papalou, O.; Victor, V.M.; Diamanti-Kandarakis, E. Oxidative Stress in Polycystic Ovary Syndrome. Curr. Pharm. Des. 2016, 22, 2709–2722. [Google Scholar] [CrossRef] [PubMed]
- Barthelmess, E.K.; Naz, R.K. Polycystic Ovary Syndrome: Current Status and Future Perspective. Front. Biosci. Elite Ed. 2014, 6, 104–119. [Google Scholar]
- Liao, D.; Liu, X.; Yuan, X.; Feng, P.; Ouyang, Z.; Liu, Y.; Li, C. Clinical Evidence of the Effects of Carnitine Supplementation on Body Weight, Glycemic Control and Serum Lipids in Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Gynecol. Endocrinol. 2022, 38, 110–115. [Google Scholar] [CrossRef]
- Mohd Shukri, M.F.; Norhayati, M.N.; Badrin, S.; Abdul Kadir, A. Effects of L-Carnitine Supplementation for Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. PeerJ 2022, 10, e13992. [Google Scholar] [CrossRef]
- Gong, Y.; Jiang, T.; He, H.; Wang, Y.; Wu, G.-L.; Shi, Y.; Cai, Q.; Xiong, C.-L.; Shen, R.; Li, J. Effects of Carnitine on Glucose and Lipid Metabolic Profiles and Fertility Outcomes in Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Clin. Endocrinol. 2023, 98, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Celik, F.; Kose, M.; Yilmazer, M.; Köken, G.N.; Arioz, D.T.; Kanat Pektas, M. Plasma L-Carnitine Levels of Obese and Non-Obese Polycystic Ovary Syndrome Patients. J. Obstet. Gynaecol. 2017, 37, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Kordus, R.J.; Hossain, A.; Malter, H.E.; LaVoie, H.A. Mitochondrial Metabolic Substrate Utilization in Granulosa Cells Reflects Body Mass Index and Total Follicle Stimulating Hormone Dosage in in Vitro Fertilization Patients. J. Assist. Reprod. Genet. 2020, 37, 2743–2756. [Google Scholar] [CrossRef]
- Chen, X.; Lu, T.; Wang, X.; Sun, X.; Zhang, J.; Zhou, K.; Ji, X.; Sun, R.; Wang, X.; Chen, M.; et al. Metabolic Alterations Associated with Polycystic Ovary Syndrome: A UPLC Q-Exactive Based Metabolomic Study. Clin. Chim. Acta 2020, 502, 280–286. [Google Scholar] [CrossRef]
- Rajska, A.; Buszewska-Forajta, M.; Rachoń, D.; Markuszewski, M.J. Metabolomic Insight into Polycystic Ovary Syndrome—An Overview. Int. J. Mol. Sci. 2020, 21, 4853. [Google Scholar] [CrossRef]
- Kalhori, Z.; Mehranjani, M.S.; Azadbakht, M.; Shariatzadeh, M.A.; Kalhori, Z.; Mehranjani, M.S.; Azadbakht, M.; Shariatzadeh, M.A. L-Carnitine Improves Endocrine Function and Folliculogenesis by Reducing Inflammation, Oxidative Stress and Apoptosis in Mice Following Induction of Polycystic Ovary Syndrome. Reprod. Fertil. Dev. 2019, 31, 282–293. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant and Antiradical Activities of L-Carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef]
- Kelek, S.E.; Afşar, E.; Akçay, G.; Danışman, B.; Aslan, M. Effect of Chronic L-Carnitine Supplementation on Carnitine Levels, Oxidative Stress and Apoptotic Markers in Peripheral Organs of Adult Wistar Rats. Food Chem. Toxicol. 2019, 134, 110851. [Google Scholar] [CrossRef]
- González, F. Inflammation in Polycystic Ovary Syndrome: Underpinning of Insulin Resistance and Ovarian Dysfunction. Steroids 2012, 77, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Vilmann, L.S.; Thisted, E.; Baker, J.L.; Holm, J.-C. Development of Obesity and Polycystic Ovary Syndrome in Adolescents. Horm. Res. Paediatr. 2012, 78, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.A.; Kyaw-Tun, T.; Young, I.S.; Phelan, N.A.; Gibney, J.; McEneny, J. Oxidative Stress and Inflammation in Lean and Obese Subjects with Polycystic Ovary Syndrome. J. Reprod. Med. 2013, 58, 107–114. [Google Scholar] [PubMed]
- Desai, V.; Prasad, N.R.; Manohar, S.M.; Sachan, A.; Narasimha, S.R.P.V.L.; Bitla, A.R.R. Oxidative Stress in Non-Obese Women with Polycystic Ovarian Syndrome. J. Clin. Diagn. Res. 2014, 8, CC01–CC03. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhu, M.; Xu, W. Roles of Oxidative Stress in Polycystic Ovary Syndrome and Cancers. Oxid. Med. Cell. Longev. 2016, 2016, 8589318. [Google Scholar] [CrossRef] [PubMed]
- Fatima, Q.; Amin, S.; Kawa, I.A.; Jeelani, H.; Manzoor, S.; Rizvi, S.M.; Rashid, F. Evaluation of Antioxidant Defense Markers in Relation to Hormonal and Insulin Parameters in Women with Polycystic Ovary Syndrome (PCOS): A Case-Control Study. Diabetes Metab. Syndr. 2019, 13, 1957–1961. [Google Scholar] [CrossRef]
- Sandhu, J.K.; Waqar, A.; Jain, A.; Joseph, C.; Srivastava, K.; Ochuba, O.; Alkayyali, T.; Ruo, S.W.; Poudel, S. Oxidative Stress in Polycystic Ovarian Syndrome and the Effect of Antioxidant N-Acetylcysteine on Ovulation and Pregnancy Rate. Cureus 2021, 13, e17887. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; He, B. Clinical and Biochemical Potential of Antioxidants in Treating Polycystic Ovary Syndrome. Int. J. Womens Health 2022, 14, 467–479. [Google Scholar] [CrossRef]
- Zhang, J.; Bao, Y.; Zhou, X.; Zheng, L. Polycystic Ovary Syndrome and Mitochondrial Dysfunction. Reprod. Biol. Endocrinol. 2019, 17, 67. [Google Scholar] [CrossRef]
- Shukla, P.; Mukherjee, S. Mitochondrial Dysfunction: An Emerging Link in the Pathophysiology of Polycystic Ovary Syndrome. Mitochondrion 2020, 52, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Malamouli, M.; Levinger, I.; McAinch, A.J.; Trewin, A.J.; Rodgers, R.J.; Moreno-Asso, A. The Mitochondrial Profile in Women with Polycystic Ovary Syndrome: Impact of Exercise. J. Mol. Endocrinol. 2022, 68, R11–R23. [Google Scholar] [CrossRef]
- Murri, M.; Luque-Ramírez, M.; Insenser, M.; Ojeda-Ojeda, M.; Escobar-Morreale, H.F. Circulating Markers of Oxidative Stress and Polycystic Ovary Syndrome (PCOS): A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2013, 19, 268–288. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Naha, M.; Malhotra, N.; Lata, K.; Vanamail, P.; Tiwari, A. Comparison of Gonadotropin-Releasing Hormone Agonist with GnRH Antagonist in Polycystic Ovary Syndrome Patients Undergoing in Vitro Fertilization Cycle: Retrospective Analysis from a Tertiary Center and Review of Literature. J. Hum. Reprod. Sci. 2014, 7, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Calonge, R.; Cortés, S.; Gutierrez Gonzalez, L.M.; Kireev, R.; Vara, E.; Ortega, L.; Caballero, P.; Rancan, L.; Tresguerres, J. Oxidative Stress in Follicular Fluid of Young Women with Low Response Compared with Fertile Oocyte Donors. Reprod. Biomed. Online 2016, 32, 446–456. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, Z.; Zhao, S.; Cheng, L.; Man, Y.; Gao, X.; Zhao, H. Oxidative Stress Markers in the Follicular Fluid of Patients with Polycystic Ovary Syndrome Correlate with a Decrease in Embryo Quality. J. Assist. Reprod. Genet. 2021, 38, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Reader, K.L.; Cox, N.R.; Stanton, J.-A.L.; Juengel, J.L. Effects of Acetyl-L-Carnitine on Lamb Oocyte Blastocyst Rate, Ultrastructure, and Mitochondrial DNA Copy Number. Theriogenology 2015, 83, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-Y.; Yang, X.-G.; Lu, S.-S.; Liang, X.-W.; Lu, Y.-Q.; Zhang, M.; Lu, K.-H. Treatment with Acetyl-l-Carnitine during in Vitro Maturation of Buffalo Oocytes Improves Oocyte Quality and Subsequent Embryonic Development. Theriogenology 2018, 118, 80–89. [Google Scholar] [CrossRef]
- Carrillo-González, D.F.; Maldonado-Estrada, J.G. L-Carnitine Supplementation in Culture Media Improves the Pregnancy Rate of in Vitro Produced Embryos with Sexed Semen from Bos Taurus Indicus Cows. Trop. Anim. Health Prod. 2020, 52, 2559–2565. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Wu, J.; Gong, S.; Ma, Y.; Zi, X. Regulation of Proliferation, Apoptosis, Hormone Secretion and Gene Expression by Acetyl-L-Carnitine in Yak (Bos Grunniens) Granulosa Cells. Theriogenology 2023, 203, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Triggiani, M.; Oriente, A.; Golino, P.; Gentile, M.; Battaglia, C.; Brevetti, G.; Marone, G. Inhibition of Platelet-Activating Factor Synthesis in Human Neutrophils and Platelets by Propionyl-l-Carnitine. Biochem. Pharmacol. 1999, 58, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Mota, S.I.; Pita, I.; Águas, R.; Tagorti, S.; Virmani, A.; Pereira, F.C.; Rego, A.C. Mechanistic Perspectives on Differential Mitochondrial-Based Neuroprotective Effects of Several Carnitine Forms in Alzheimer’s Disease in Vitro Model. Arch. Toxicol. 2021, 95, 2769–2784. [Google Scholar] [CrossRef]
- Tatone, C.; Heizenrieder, T.; Di Emidio, G.; Treffon, P.; Amicarelli, F.; Seidel, T.; Eichenlaub-Ritter, U. Evidence That Carbonyl Stress by Methylglyoxal Exposure Induces DNA Damage and Spindle Aberrations, Affects Mitochondrial Integrity in Mammalian Oocytes and Contributes to Oocyte Ageing. Hum. Reprod. 2011, 26, 1843–1859. [Google Scholar] [CrossRef]
- Gupta, A.; Chaube, S.K. Cilostamide and Rolipram Prevent Spontaneous Meiotic Resumption from Diplotene Arrest in Rat Oocytes Cultured in Vitro. Eur. J. Pharmacol. 2020, 878, 173115. [Google Scholar] [CrossRef]
- Di Emidio, G.; Falone, S.; Vitti, M.; D’Alessandro, A.M.; Vento, M.; Di Pietro, C.; Amicarelli, F.; Tatone, C. SIRT1 Signalling Protects Mouse Oocytes against Oxidative Stress and Is Deregulated during Aging. Hum. Reprod. 2014, 29, 2006–2017. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Muller, B.; Lewis, N.; Adeniyi, T.; Leese, H.J.; Brison, D.R.; Sturmey, R.G. Application of Extracellular Flux Analysis for Determining Mitochondrial Function in Mammalian Oocytes and Early Embryos. Sci. Rep. 2019, 9, 16778. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.C.Y.; Dunning, K.R. Non-Invasive Assessment of Oocyte Developmental Competence. Reprod. Fertil. Dev. 2022, 35, 39–50. [Google Scholar] [CrossRef]
- Capece, D.; Verzella, D.; Begalli, F.; Bennett, J.; D’Andrea, D.; Vecchiotti, D.; Zazzeroni, F.; Franzoso, G. Extracellular Flux Analysis to Investigate the Impact of NF-ΚB on Mitochondrial Respiration in Colorectal Carcinoma (CRC). Methods Mol. Biol. 2021, 2366, 293–303. [Google Scholar] [CrossRef]
- Placidi, M.; Di Emidio, G.; Virmani, A.; D’Alfonso, A.; Artini, P.G.; D’Alessandro, A.M.; Tatone, C. Carnitines as Mitochondrial Modulators of Oocyte and Embryo Bioenergetics. Antioxidants 2022, 11, 745. [Google Scholar] [CrossRef]
- Palmerini, M.G.; Macchiarelli, G.; Cocciolone, D.; Mascitti, I.A.; Placidi, M.; Vergara, T.; Di Emidio, G.; Tatone, C. Modulating Morphological and Redox/Glycative Alterations in the PCOS Uterus: Effects of Carnitines in PCOS Mice. Biomedicines 2023, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Whigham, L.D.; Butz, D.E.; Dashti, H.; Tonelli, M.; Johnson, L.K.; Cook, M.E.; Porter, W.P.; Eghbalnia, H.R.; Markley, J.L.; Lindheim, S.R.; et al. Metabolic Evidence of Diminished Lipid Oxidation in Women with Polycystic Ovary Syndrome. Curr. Metabolomics 2014, 2, 269–278. [Google Scholar] [CrossRef]
- Cree-Green, M.; Carreau, A.-M.; Rahat, H.; Garcia-Reyes, Y.; Bergman, B.C.; Pyle, L.; Nadeau, K.J. Amino Acid and Fatty Acid Metabolomic Profile during Fasting and Hyperinsulinemia in Girls with Polycystic Ovarian Syndrome. Am. J. Physiol.—Endocrinol. Metab. 2019, 316, E707–E718. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, J.; Li, M.; Shang, J.; Bai, X.; Zhang, H.; Wang, Y.; Chen, H.; Song, X. Serum Fatty Acid Profiles Associated with Metabolic Risk in Women with Polycystic Ovary Syndrome. Front. Endocrinol. 2023, 14, 1077590. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Yin, T.-L.; Yang, J.; Xiong, C.-L. Follicular Metabolic Changes and Effects on Oocyte Quality in Polycystic Ovary Syndrome Patients. Oncotarget 2017, 8, 80472–80480. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, I.R.; Joshi, M. CPT1A-Mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Liang, K. Mitochondrial CPT1A: Insights into Structure, Function, and Basis for Drug Development. Front. Pharmacol. 2023, 14, 1160440. [Google Scholar] [CrossRef]
- Dunning, K.R.; Anastasi, M.R.; Zhang, V.J.; Russell, D.L.; Robker, R.L. Regulation of Fatty Acid Oxidation in Mouse Cumulus-Oocyte Complexes during Maturation and Modulation by PPAR Agonists. PLoS ONE 2014, 9, e87327. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.-M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef]
- Liu, T.; Qu, J.; Tian, M.; Yang, R.; Song, X.; Li, R.; Yan, J.; Qiao, J. Lipid Metabolic Process Involved in Oocyte Maturation During Folliculogenesis. Front. Cell Dev. Biol. 2022, 10, 806890. [Google Scholar] [CrossRef]
- Price, N.T.; van der Leij, F.R.; Jackson, V.N.; Corstorphine, C.G.; Thomson, R.; Sorensen, A.; Zammit, V.A. A Novel Brain-Expressed Protein Related to Carnitine Palmitoyltransferase I. Genomics 2002, 80, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Wolfgang, M.J.; Kurama, T.; Dai, Y.; Suwa, A.; Asaumi, M.; Matsumoto, S.; Cha, S.H.; Shimokawa, T.; Lane, M.D. The Brain-Specific Carnitine Palmitoyltransferase-1c Regulates Energy Homeostasis. Proc. Natl. Acad. Sci. USA 2006, 103, 7282–7287. [Google Scholar] [CrossRef]
- Sierra, A.Y.; Gratacós, E.; Carrasco, P.; Clotet, J.; Ureña, J.; Serra, D.; Asins, G.; Hegardt, F.G.; Casals, N. CPT1c Is Localized in Endoplasmic Reticulum of Neurons and Has Carnitine Palmitoyltransferase Activity. J. Biol. Chem. 2008, 283, 6878–6885. [Google Scholar] [CrossRef]
- Roa-Mansergas, X.; Fadó, R.; Atari, M.; Mir, J.F.; Muley, H.; Serra, D.; Casals, N. CPT1C Promotes Human Mesenchymal Stem Cells Survival under Glucose Deprivation through the Modulation of Autophagy. Sci. Rep. 2018, 8, 6997. [Google Scholar] [CrossRef] [PubMed]
- Zaugg, K.; Yao, Y.; Reilly, P.T.; Kannan, K.; Kiarash, R.; Mason, J.; Huang, P.; Sawyer, S.K.; Fuerth, B.; Faubert, B.; et al. Carnitine Palmitoyltransferase 1C Promotes Cell Survival and Tumor Growth under Conditions of Metabolic Stress. Genes Dev. 2011, 25, 1041–1051. [Google Scholar] [CrossRef]
- Joshi, M.; Kim, J.; D’Alessandro, A.; Monk, E.; Bruce, K.; Elajaili, H.; Nozik-Grayck, E.; Goodspeed, A.; Costello, J.C.; Schlaepfer, I.R. CPT1A Over-Expression Increases Reactive Oxygen Species in the Mitochondria and Promotes Antioxidant Defenses in Prostate Cancer. Cancers 2020, 12, 3431. [Google Scholar] [CrossRef]
- Jiang, N.; Xing, B.; Peng, R.; Shang, J.; Wu, B.; Xiao, P.; Lin, S.; Xu, X.; Lu, H. Inhibition of Cpt1a Alleviates Oxidative Stress-Induced Chondrocyte Senescence via Regulating Mitochondrial Dysfunction and Activating Mitophagy. Mech. Ageing Dev. 2022, 205, 111688. [Google Scholar] [CrossRef]
- Arduini, A.; Dottori, S.; Sciarroni, A.F.; Corsico, N.; Morabito, E.; Arrigoni-Martelli, E.; Calvani, M. Effect of Propionyl-L-Carnitine Treatment on Membrane Phospholipid Fatty Acid Turnover in Diabetic Rat Erythrocytes. Mol. Cell. Biochem. 1995, 152, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Mingorance, C.; Rodriguez-Rodriguez, R.; Justo, M.L.; Herrera, M.D.; de Sotomayor, M.A. Pharmacological Effects and Clinical Applications of Propionyl-L-Carnitine. Nutr. Rev. 2011, 69, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Hoppel, C. The Role of Carnitine in Normal and Altered Fatty Acid Metabolism. Am. J. Kidney Dis. 2003, 41, S4–S12. [Google Scholar] [CrossRef]
- Guan, L.; Chen, Y.; Wang, Y.; Zhang, H.; Fan, S.; Gao, Y.; Jiao, T.; Fu, K.; Sun, J.; Yu, A.; et al. Effects of Carnitine Palmitoyltransferases on Cancer Cellular Senescence. J. Cell. Physiol. 2019, 234, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Fadó, R.; Zagmutt, S.; Herrero, L.; Muley, H.; Rodríguez-Rodríguez, R.; Bi, H.; Serra, D.; Casals, N. To Be or Not to Be a Fat Burner, That Is the Question for Cpt1c in Cancer Cells. Cell Death Dis. 2023, 14, 57. [Google Scholar] [CrossRef]
- Lewis, N.; Hinrichs, K.; Leese, H.J.; Argo, C.M.; Brison, D.R.; Sturmey, R. Energy Metabolism of the Equine Cumulus Oocyte Complex during in Vitro Maturation. Sci. Rep. 2020, 10, 3493. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.G.; Benavides, G.A.; Lancaster, J.R.; Ballinger, S.; Dell’Italia, L.; Zhang, J.; Darley-Usmar, V.M. Integration of Cellular Bioenergetics with Mitochondrial Quality Control and Autophagy. Biol. Chem. 2012, 393, 1485–1512. [Google Scholar] [CrossRef]
- Paczkowski, M.; Schoolcraft, W.B.; Krisher, R.L. Fatty Acid Metabolism during Maturation Affects Glucose Uptake and Is Essential to Oocyte Competence. Reproduction 2014, 148, 429–439. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Placidi, M.; Rossi, G.; Ruggieri, S.; Taccaliti, C.; D’Alfonso, A.; Amicarelli, F.; Guido, M. AGEs-Related Dysfunctions in PCOS: Evidence from Animal and Clinical Research. J. Endocrinol. 2021, 251, R1–R9. [Google Scholar] [CrossRef]
- Sudhakaran, G.; Babu, S.R.; Mahendra, H.; Arockiaraj, J. Updated Experimental Cellular Models to Study Polycystic Ovarian Syndrome. Life Sci. 2023, 322, 121672. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | n | Degeneration (Mean% ± SEM) | GV (Mean% ± SEM) | GVBD (Mean% ± SEM) | MII (Mean% ± SEM) |
|---|---|---|---|---|---|
| CONTROL | 192 | 2.2% ± 1.3 | 12.0% ± 4.4 | 13.8% ± 4.2 | 72.0% ± 4.2 |
| H2O2 100 µM | 145 | 1.2% ± 0.7 | 15.9% ± 7.7 | 44.1% ± 2.9 * | 36.7% ± 7.8 * |
| H2O2 100 µM + (ALC 0.04 mg/mL − LC 0.08 mg/mL) | 46 | 0.0% ± 0.0 | 16.3% ± 8.7 | 26.7% ± 11.7 # | 56.9% ± 3.1 |
| H2O2 100 µM + (ALC 0.2 mg/mL − LC 0.4 mg/mL) | 53 | 2.0% ± 2.0 | 9.4% ± 1.4 | 9.4% ± 1.36 # | 73.1% ± 12.8 # |
| H2O2 100 µM + (ALC 0.8 mg/mL − LC 1.6 mg/mL) | 47 | 0.0% ± 0.0 | 12.8% ± 0.8 | 12.8% ± 0.8 # | 72.1% ± 3.9 # |
| H2O2 100 µM + PLC 0.2 mg/mL | 89 | 0.0% ± 0.0 | 18.8% ± 4.0 | 12.5% ± 12.5 # | 69.3% ± 9.3 # |
| H2O2 100 µM + PLC 1.0 mg/mL | 42 | 0.0% ± 0.0 | 4.8% ± 1.2 | 4.8% ± 4.0 # | 90.5% ± 5.2 # |
| H2O2 100 µM + PLC 2.0 mg/mL | 48 | 4.2% ± 1.2 | 1.1% ± 1.1 | 33.3% ± 3.8 | 62.5% ± 6.1 # |
| H2O2 100 µM + (PLC 0.2 mg/mL − ALC 0.2 mg/mL − LC 0.4 mg/mL) | 68 | 0.0% ± 0.0 | 10.8% ± 3.4 | 17.5% ± 6.4 # | 71.7% ± 9.7 # |
| Experimental Group | n | Normal (Mean% ± SEM) | Slightly Aberrant (Mean% ± SEM) | Aberrant (Mean% ± SEM) |
|---|---|---|---|---|
| CONTROL | 66 | 55.7% ± 13.0 | 32.5% ± 11.5 | 11.8% ± 1.4 |
| H2O2 100 µM | 52 | 14.8% ± 5.9 * | 51.5%± 6.1 | 33.6% ± 2.0 * |
| H2O2 100 µM + (ALC 0.04 mg/mL − LC 0.08 mg/mL) | 26 | 23.5% ± 9.5 | 7.0% ± 7.0 # | 69.5% ± 2.5 #,* |
| H2O2 100 µM + (ALC 0.2 mg/mL − LC 0.4 mg/mL) | 38 | 22.5% ± 2.5 | 23.3%± 3.3 # | 54.2% ± 0.8 #* |
| H2O2 100 µM + (ALC 0.8 mg/mL − LC 1.6 mg/mL) | 33 | 9.4% ± 3.9 * | 27.2% ± 0.6 # | 63.3% ± 3.3 #,* |
| H2O2 100 µM + PLC 0.2 mg/mL | 38 | 32.7% ± 3.9 # | 51.7% ± 21.2 | 15.5% ± 18.0 |
| H2O2 100 µM + PLC 1.0 mg/mL | 19 | 42.1% ± 7.6 # | 47.4% ± 1.0 | 10.5% ± 8.6 |
| H2O2 100 µM + PLC 2.0 mg/mL | 21 | 49.0% ± 11.1# | 18.2% ± 1.2 # | 40.5% ± 9.6 * |
| H2O2 100 µM + (PLC 0.2 mg/mL − ALC 0.2 mg/mL − LC 0.4 mg/mL) | 36 | 90.9% ± 9.1 # | 9.1% ± 9.1 # | 0.0% ± 0.0 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Placidi, M.; Vergara, T.; Casoli, G.; Flati, I.; Capece, D.; Artini, P.G.; Virmani, A.; Zanatta, S.; D’Alessandro, A.M.; Tatone, C.; et al. Acyl-Carnitines Exert Positive Effects on Mitochondrial Activity under Oxidative Stress in Mouse Oocytes: A Potential Mechanism Underlying Carnitine Efficacy on PCOS. Biomedicines 2023, 11, 2474. https://doi.org/10.3390/biomedicines11092474
Placidi M, Vergara T, Casoli G, Flati I, Capece D, Artini PG, Virmani A, Zanatta S, D’Alessandro AM, Tatone C, et al. Acyl-Carnitines Exert Positive Effects on Mitochondrial Activity under Oxidative Stress in Mouse Oocytes: A Potential Mechanism Underlying Carnitine Efficacy on PCOS. Biomedicines. 2023; 11(9):2474. https://doi.org/10.3390/biomedicines11092474
Chicago/Turabian StylePlacidi, Martina, Teresa Vergara, Giovanni Casoli, Irene Flati, Daria Capece, Paolo Giovanni Artini, Ashraf Virmani, Samuele Zanatta, Anna Maria D’Alessandro, Carla Tatone, and et al. 2023. "Acyl-Carnitines Exert Positive Effects on Mitochondrial Activity under Oxidative Stress in Mouse Oocytes: A Potential Mechanism Underlying Carnitine Efficacy on PCOS" Biomedicines 11, no. 9: 2474. https://doi.org/10.3390/biomedicines11092474