
Red Blood Cell Adenylate Energetics Is Related to Endothelial and Microvascular Function in Long COVID
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Peripheral Blood Sampling and Morphology
2.3. Erythrocyte Nucleotide Measurements
2.4. Serum Amino Acid Measurements
2.5. Serum hs-CRP Measurement
2.6. Microvascular Function Measurements
2.7. Statistical Analysis
3. Results
3.1. Long COVID Patients Demonstrate Decreased Peripheral Tissue Oxygenation with Changes in Endothelial and Microvascular Function Parameters
3.2. Adenine Nucleotides’ Concentration in the Erythrocytes of Long COVID Patients Is at Similar Levels as in the Healthy Controls
3.3. Adenosine Triphosphate (ATP) Concentration in the Erythrocytes of Long COVID Patients Correlates with Markers of Endothelial and Microcirculatory Function
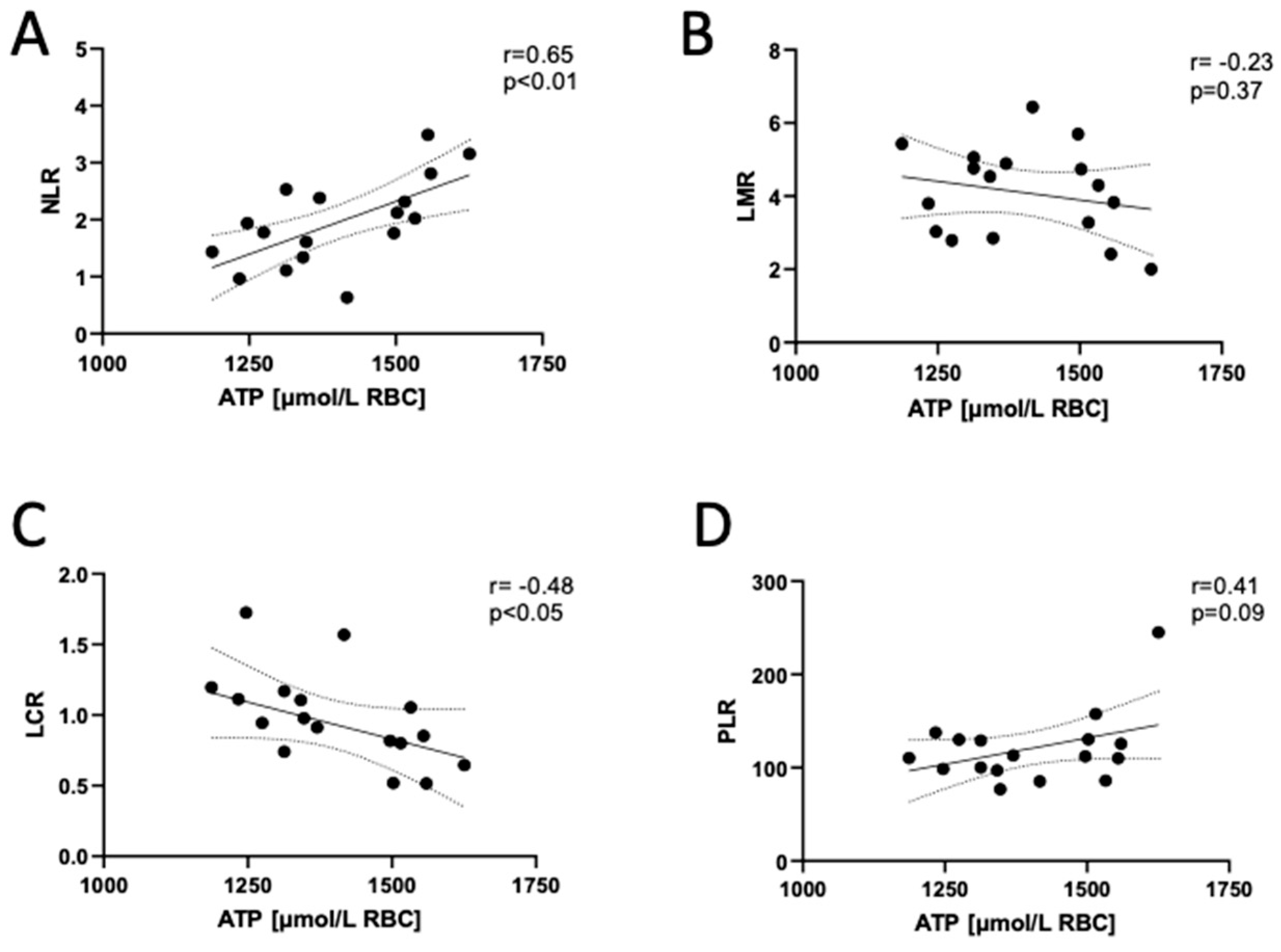
3.4. Adenosine Triphosphate (ATP) Concentration in the Erythrocytes of Long COVID Patients Correlates with Markers of Systemic Inflammation Reactivation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Touma, M. COVID-19: Molecular Diagnostics Overview. J. Mol. Med. 2020, 98, 947. [Google Scholar] [CrossRef]
- Staffolani, S.; Iencinella, V.; Cimatti, M.; Tavio, M. Long COVID-19 Syndrome as a Fourth Phase of SARS-CoV-2 Infection. Infez. Med. 2022, 30, 22–29. [Google Scholar] [CrossRef]
- Østergaard, L. SARS CoV-2 Related Microvascular Damage and Symptoms during and after COVID-19: Consequences of Capillary Transit-Time Changes, Tissue Hypoxia and Inflammation. Physiol. Rep. 2021, 9, e14726. [Google Scholar] [CrossRef]
- Böning, D.; Kuebler, W.M.; Bloch, W. The Oxygen Dissociation Curve of Blood in COVID-19. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321, L349–L357. [Google Scholar] [CrossRef] [PubMed]
- Gérard, D.; Ben Brahim, S.; Lesesve, J.F.; Perrin, J. Are Mushroom-Shaped Erythrocytes an Indicator of COVID-19? Br. J. Haematol. 2021, 192, 230. [Google Scholar] [CrossRef] [PubMed]
- Gagiannis, D.; Umathum, V.G.; Bloch, W.; Rother, C.; Stahl, M.; Witte, H.M.; Djudjaj, S.; Boor, P.; Steinestel, K. Antemortem vs Postmortem Histopathologic and Ultrastructural Findings in Paired Transbronchial Biopsy Specimens and Lung Autopsy Samples from Three Patients with Confirmed SARS-CoV-2. Am. J. Clin. Pathol. 2022, 157, 54–63. [Google Scholar] [CrossRef]
- Renoux, C.; Fort, R.; Nader, E.; Boisson, C.; Joly, P.; Stauffer, E.; Robert, M.; Girard, S.; Cibiel, A.; Gauthier, A.; et al. Impact of COVID-19 on Red Blood Cell Rheology. Br. J. Haematol. 2021, 192, e108–e111. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.; Ibershoff, L.; Zacher, J.; Bros, J.; Tomschi, F.; Diebold, K.F.; Predel, H.G.; Bloch, W. Even Patients with Mild COVID-19 Symptoms after SARS-CoV-2 Infection Show Prolonged Altered Red Blood Cell Morphology and Rheological Parameters. J. Cell. Mol. Med. 2022, 26, 3022–3030. [Google Scholar] [CrossRef]
- Dudzinska, W.; Hlynczak, A.J.; Skotnicka, E.; Suska, M. The Purine Metabolism of Human Erythrocytes. Biochemistry 2006, 71, 467–475. [Google Scholar] [CrossRef]
- Van Wijk, R.; Van Solinge, W.W. The Energy-Less Red Blood Cell Is Lost: Erythrocyte Enzyme Abnormalities of Glycolysis. Blood 2005, 106, 4034–4042. [Google Scholar] [CrossRef]
- McMahon, T.J.; Darrow, C.C.; Hoehn, B.A.; Zhu, H. Generation and Export of Red Blood Cell ATP in Health and Disease. Front. Physiol. 2021, 12, 754638. [Google Scholar] [CrossRef] [PubMed]
- Racine, M.L.; Dinenno, F.A. Reduced Deformability Contributes to Impaired Deoxygenation-Induced ATP Release from Red Blood Cells of Older Adult Humans. J. Physiol. 2019, 597, 4503–4519. [Google Scholar] [CrossRef] [PubMed]
- Dudzinska, W.; Suska, M.; Lubkowska, A.; Jakubowska, K.; Olszewska, M.; Safranow, K.; Chlubek, D. Comparison of Human Erythrocyte Purine Nucleotide Metabolism and Blood Purine and Pyrimidine Degradation Product Concentrations before and after Acute Exercise in Trained and Sedentary Subjects. J. Physiol. Sci. 2018, 68, 293–305. [Google Scholar] [CrossRef]
- Kosenko, E.A.; Tikhonova, L.A.; Montoliu, C.; Barreto, G.E.; Aliev, G.; Kaminsky, Y.G. Metabolic Abnormalities of Erythrocytes as a Risk Factor for Alzheimer’s Disease. Front. Neurosci. 2018, 11, 728. [Google Scholar] [CrossRef]
- Papandreou, C.; Li, J.; Liang, L.; Bulló, M.; Zheng, Y.; Ruiz-Canela, M.; Yu, E.; Guasch-Ferré, M.; Razquin, C.; Clish, C.; et al. Metabolites Related to Purine Catabolism and Risk of Type 2 Diabetes Incidence; Modifying Effects of the TCF7L2-Rs7903146 Polymorphism. Sci. Rep. 2019, 9, 2892. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Stefanoni, D.; Dzieciatkowska, M.; Issaian, A.; Nemkov, T.; Hill, R.C.; Francis, R.O.; Hudson, K.E.; Buehler, P.W.; Zimring, J.C.; et al. Evidence of Structural Protein Damage and Membrane Lipid Remodeling in Red Blood Cells from COVID-19 Patients. J. Proteome Res. 2020, 19, 4455–4469. [Google Scholar] [CrossRef]
- Kutryb-Zajac, B.; Kawecka, A.; Caratis, F.; Urbanowicz, K.; Braczko, A.; Furihata, T.; Karaszewski, B.; Smolenski, R.T.; Rutkowska, A. The Impaired Distribution of Adenosine Deaminase Isoenzymes in Multiple Sclerosis Plasma and Cerebrospinal Fluid. Front. Mol. Neurosci. 2022, 15, 998023. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejewska, A.; Kawecka, A.; Braczko, A.; Romanowska-Kocejko, M.; Stawarska, K.; Deptuła, M.; Zawrzykraj, M.; Franczak, M.; Krol, O.; Harasim, G.; et al. Changes in Adenosine Deaminase Activity and Endothelial Dysfunction after Mild Coronavirus Disease-2019. Int. J. Mol. Sci. 2023, 24, 13140. [Google Scholar] [CrossRef]
- Olkowicz, M.; Debski, J.; Jablonska, P.; Dadlez, M.; Smolenski, R.T. Application of a New Procedure for Liquid Chromatography/Mass Spectrometry Profiling of Plasma Amino Acid-Related Metabolites and Untargeted Shotgun Proteomics to Identify Mechanisms and Biomarkers of Calcific Aortic Stenosis. J. Chromatogr. A 2017, 1517, 66–78. [Google Scholar] [CrossRef]
- Tarnawska, M.; Dorniak, K.; Kaszubowski, M.; Dudziak, M.; Hellmann, M. A Pilot Study with Flow Mediated Skin Fluorescence: A Novel Device to Assess Microvascular Endothelial Function in Coronary Artery Disease. Cardiol. J. 2018, 25, 120–127. [Google Scholar] [CrossRef]
- Romanowska-Kocejko, M.; Dudziak, M.; Hellmann, M. Nicotinamide Adenine Dinucleotide Fluorescence to Assess Microvascular Disturbances in Post-COVID-19 Patients. Cardiol. J. 2022, 29, 154–156. [Google Scholar] [CrossRef]
- Katarzynska, J.; Zielinski, J.; Marcinek, A.; Gebicki, J. New Approach to Non-Invasive Assessment of Vascular Circulation Based on the Response to Transient Ischemia. Vasc. Health Risk Manag. 2022, 18, 113–116. [Google Scholar] [CrossRef]
- Li, C.Y. Can Glycine Mitigate COVID-19 Associated Tissue Damage and Cytokine Storm? Radiat. Res. 2020, 194, 199. [Google Scholar] [CrossRef]
- Toori, K.U.; Qureshi, M.A.; Chaudhry, A.; Safdar, M.F. Neutrophil to Lymphocyte Ratio (NLR) in COVID-19: A Cheap Prognostic Marker in a Resource Constraint Setting. Pak. J. Med. Sci. 2021, 37, 1435. [Google Scholar] [CrossRef]
- Fisher, L.; Goka, B.; Pater, J.; Chang, F.-L. Neutrophil-to-Lymphocyte Ratio (NLR) to Monitor Neuroinflammation Status During Long COVID. Proc. IMPRS 2023, 6, 1. [Google Scholar] [CrossRef]
- Jensen, F.B. The Dual Roles of Red Blood Cells in Tissue Oxygen Delivery: Oxygen Carriers and Regulators of Local Blood Flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef]
- Płoszczyca, K.; Czuba, M.; Chalimoniuk, M.; Gajda, R.; Baranowski, M. Red Blood Cell 2,3-Diphosphoglycerate Decreases in Response to a 30 Km Time Trial Under Hypoxia in Cyclists. Front. Physiol. 2021, 12, 670977. [Google Scholar] [CrossRef]
- McMahon, T.J. Red Blood Cell Deformability, Vasoactive Mediators, and Adhesion. Front. Physiol. 2019, 10, 1417. [Google Scholar] [CrossRef] [PubMed]
- Grygorczyk, R.; Orlov, S.N. Effects of Hypoxia on Erythrocyte Membrane Properties—Implications for Intravascular Hemolysis and Purinergic Control of Blood Flow. Front. Physiol. 2017, 8, 1110. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, E.; Tikhonova, L.; Alilova, G.; Montoliu, C. Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 5739. [Google Scholar] [CrossRef] [PubMed]
- Marcinek, A.; Katarzynska, J.; Sieron, L.; Skokowski, R.; Zielinski, J.; Gebicki, J. Non-Invasive Assessment of Vascular Circulation Based on Flow Mediated Skin Fluorescence (FMSF). Biology 2023, 12, 385. [Google Scholar] [CrossRef]
- Gebicki, J.; Katarzynska, J.; Marcinek, A. Can Efficient Stimulation of Myogenic Microcirculatory Oscillations by Transient Ischemia Predict Low Incidence of COVID-19 Infection? Respir. Physiol. Neurobiol. 2021, 286, 103618. [Google Scholar] [CrossRef] [PubMed]
- Chudzik, M.; Cender, A.; Mordaka, R.; Zielinski, J.; Katarzynska, J.; Marcinek, A.; Gebicki, J. Chronic Fatigue Associated with Post-COVID Syndrome versus Transient Fatigue Caused by High-Intensity Exercise: Are They Comparable in Terms of Vascular Effects? Vasc. Health Risk Manag. 2022, 18, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ilyas, I.; Weng, J. Endothelial Dysfunction in COVID-19: An Overview of Evidence, Biomarkers, Mechanisms and Potential Therapies. Acta Pharmacol. Sin. 2023, 44, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Maccio, U.; Zinkernagel, A.S.; Shambat, S.M.; Zeng, X.; Cathomas, G.; Ruschitzka, F.; Schuepbach, R.A.; Moch, H.; Varga, Z. SARS-CoV-2 Leads to a Small Vessel Endotheliitis in the Heart. EBioMedicine 2021, 63, 103182. [Google Scholar] [CrossRef] [PubMed]
- Fedorowski, A.; Fanciulli, A.; Raj, S.R.; Sheldon, R.; Shibao, C.A.; Sutton, R. Cardiovascular Autonomic Dysfunction in Post-COVID-19 Syndrome: A Major Health-Care Burden. Nat. Rev. Cardiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.J.; Kuck, L.; Michael, J.S. Beyond Oxygen Transport: Active Role of Erythrocytes in the Regulation of Blood Flow. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H866–H872. [Google Scholar] [CrossRef]
- Kirby, B.S.; Sparks, M.A.; Lazarowski, E.R.; Lopez Domowicz, D.A.; Zhu, H.; McMahon, T.J. Pannexin 1 Channels Control the Hemodynamic Response to Hypoxia by Regulating O2-Sensitive Extracellular ATP in Blood. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1055–H1065. [Google Scholar] [CrossRef]
- Da Silva, C.G.; Specht, A.; Wegiel, B.; Ferran, C.; Kaczmarek, E. Mechanism of Purinergic Activation of Endothelial Nitric Oxide Synthase in Endothelial Cells. Circulation 2009, 119, 871–879. [Google Scholar] [CrossRef]
- Cosic, I.; Cosic, D.; Loncarevic, I. RRM Prediction of Erythrocyte Band3 Protein as Alternative Receptor for SARS-CoV-2 Virus. Appl. Sci. 2020, 10, 4053. [Google Scholar] [CrossRef]
- Misiti, F. SARS-CoV-2 Infection and Red Blood Cells: Implications for Long Term Symptoms during Exercise. Sports Med. Health Sci. 2021, 3, 181–182. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control (n = 20) | Long COVID (n = 19) |
|---|---|---|
| Age (years) | 40 ± 3 | 38 ± 2 |
| Sex (F/M) | 14/6 | 12/7 |
| BMI (kg/m2) | 23.3 ± 0.9 | 23.5 ± 0.8 |
| Long COVID symptoms | ||
| Fatigue | Na | 16 (84%) |
| Tachycardia | Na | 3 (16%) |
| Chest pain | Na | 6 (32%) |
| Dyspnea | Na | 1 (5%) |
| Headache | Na | 1 (5%) |
| Comorbidities | ||
| Hypothyroidism | 5 (25%) | 5 (26%) |
| Asthma | 0 (0%) | 1 (5%) |
| Atopic dermatitis | 0 (0%) | 1 (5%) |
| Irritable bowel syndrome | 1 (5%) | 1 (5%) |
| Depression | 0 (0%) | 1 (5%) |
| Hyperlipidemia | 0 (0%) | 1 (5%) |
| Diabetes | 0 (0%) | 1 (5%) |
| Thrombosis | 0 (0%) | 1 5%) |
| Drugs taken | ||
| Levothyroxine | 5 (25%) | 5 (26%) |
| Antidepressants | 0 (0%) | 2 (11%) |
| Metformin | 0 (0%) | 1 (5%) |
| Acetylsalicylic acid | 0 (0%) | 1 (5%) |
| Drugs prescribed | ||
| Beta-adrenolytics | 0 (0%) | 6 (32%) |
| ACE-inhibitors | 0 (0%) | 3 (16%) |
| Statins | 0 (0%) | 1 (5%) |
| Parameter | Control | Long COVID | p Value |
|---|---|---|---|
| Microcirculatory function parameters | |||
| IR index [%] | 14.5 ± 1.74 | 6.06 ± 1.09 | <0.001 |
| IR max [%] | 19.5 ± 2.12 | 9.30 ± 1.32 | <0.001 |
| HR index [%] | 15.1 ± 1.15 | 13.3 ± 0.73 | 0.20 |
| HR max [%] | 22.3 ± 2.29 | 20.3 ± 0.72 | 0.42 |
| RHR [%] | 38.7 ± 2.24 | 30.8 ± 1.32 | <0.01 |
| Log (HS) | 114 ± 38.7 | 49.6 ± 9.85 | <0.05 |
| Serum inflammatory parameters | |||
| hs-CRP [mg/L] | 2.1 ± 0.30 | 2.4 ± 0.31 | 0.489 |
| Serum amino acid compounds | |||
| Arginine [μmol/L] | 112 ± 6.92 | 126 ± 5.27 | 0.12 |
| Citrulline [μmol/L] | 15.5 ± 0.93 | 17.2 ± 0.76 | 0.17 |
| SDMA [μmol/L] | 0.83 ± 0.04 | 0.86 ± 0.05 | 0.64 |
| Arginine/ADMA | 162 ± 8.00 | 151 ± 8.08 | 0.34 |
| Glycine [μmol/L] | 314 ± 37.4 | 219 ± 12.9 | <0.05 |
| Peripheral blood cell count | |||
| RBCs [T/L] | 4.86 ± 0.10 | 4.70 ± 0.09 | 0.26 |
| Hct [%] | 42.4 ± 0.74 | 42.2 ± 0.75 | 0.25 |
| Hgb [g/dL] | 14.3 ± 0.29 | 14.2 ± 0.29 | 0.79 |
| MCV [fL] | 88.7 ± 0.91 | 87.7 ± 0.95 | 0.45 |
| MCH [pg] | 30.5 ± 0.32 | 30.2 ± 0.46 | 0.60 |
| MCHC [g/dL] | 34.3 ± 0.32 | 34.4 ± 0.31 | 0.81 |
| RDW [%] | 12.9 ± 0.18 | 12.8 ± 0.20 | 0.85 |
| WBC [G/L] | 6.22 ± 1.81 | 6.28 ± 0.47 | 0.96 |
| NEU [G/L] | 3.52 ± 1.22 | 3.65 ± 0.37 | 0.92 |
| NEU [%] | 56.6 ± 2.11 | 56.5 ± 2.21 | 0.98 |
| LYMPH [G/L] | 2.31 ± 0.23 | 1.94 ± 0.11 | 0.16 |
| LYMPH [%] | 37.1 ± 2.43 | 32.3 ± 2.03 | 0.14 |
| MONO [G/L] | 0.48 ± 0.05 | 0.51 ± 0.04 | 0.64 |
| MONO [%] | 7.72 ± 0.19 | 8.22 ± 0.48 | 0.33 |
| EOS [G/L] | 0.12 ± 0.03 | 0.15 ± 0.02 | 0.42 |
| EOS [%] | 1.93 ± 0.21 | 2.35 ± 0.26 | 0.21 |
| BAS [G/L] | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.99 |
| BAS [%] | 0.64 ± 0.06 | 0.56 ± 0.05 | 0.31 |
| PLT [G/L] | 216 ± 12.0 | 222 ± 11.0 | 0.72 |
| PDW [fL] | 10.9 ± 0.51 | 13.2 ± 0.41 | <0.01 |
| PCT [%] | 0.23 ± 0.01 | 0.24 ± 0.01 | 0.48 |
| Large PLT [%] | 26.7 ± 2.31 | 32.3 ± 1.44 | <0.05 |
| NLR | 1.52 ± 0.12 | 1.93 ± 0.18 | 0.06 |
| LMR | 4.81 ± 0.32 | 4.11 ± 0.29 | 0.11 |
| LCR | 1.10 ± 0.09 | 0.94 ± 0.08 | 0.19 |
| PLR | 93.5 ± 10.4 | 120 ± 8.80 | 0.06 |
| Parameter | ATP [μmol/L RBC] | ADP [μmol/L RBC] | AMP [μmol/L RBC] | TAN [μmol/L RBC] | ||||
|---|---|---|---|---|---|---|---|---|
| r | p Value | r | p Value | r | p Value | r | p Value | |
| RBC [T/L] | −0.28 | 0.26 | 0.12 | 0.63 | 0.08 | 0.75 | −0.23 | 0.36 |
| Hct [%] | −0.27 | 0.28 | 0.14 | 0.57 | 0.04 | 0.87 | −0.21 | 0.39 |
| Hgb [g/dL] | −0.21 | 0.42 | 0.13 | 0.61 | 0.07 | 0.79 | −0.16 | 0.53 |
| MCV [fL] | 0.08 | 0.74 | 0.04 | 0.87 | −0.04 | 0.87 | 0.09 | 0.73 |
| MCH [pg] | 0.23 | 0.37 | 0.18 | 0.47 | 0.03 | 0.90 | 0.23 | 0.37 |
| MCHC [g/dL] | 0.14 | 0.59 | −0.01 | 0.99 | 0.07 | 0.77 | 0.13 | 0.61 |
| RDW [%] | −0.34 | 0.17 | −0.17 | 0.50 | −0.17 | 0.49 | −0.37 | 0.13 |
| WBC [G/L] | 0.26 | 0.30 | −0.10 | 0.70 | −0.16 | 0.54 | 0.22 | 0.37 |
| NEU [G/L] | 0.39 | 0.11 | −0.03 | 0.91 | −0.12 | 0.64 | 0.37 | 0.13 |
| NEU [%] | 0.55 | <0.05 | 0.07 | 0.78 | −0.06 | 0.80 | 0.55 | <0.05 |
| LYMPH [G/L] | −0.24 | 0.34 | −0.17 | 0.50 | −0.11 | 0.66 | −0.27 | 0.27 |
| LYMPH [%] | −0.52 | <0.05 | 0.01 | 0.97 | −0.13 | 0.60 | −0.50 | <0.05 |
| MONO [G/L] | 0.01 | 0.98 | −0.42 | 0.08 | −0.40 | 0.10 | −0.11 | 0.66 |
| MONO [%] | −0.28 | 0.26 | −0.40 | 0.10 | −0.32 | 0.20 | −0.38 | 0.12 |
| EOS [G/L] | 0.12 | 0.63 | 0.06 | 0.83 | 0.02 | 0.95 | 0.13 | 0.60 |
| EOS [%] | −0.04 | 0.89 | 0.07 | 0.78 | 0.09 | 0.71 | −0.02 | 0.95 |
| BAS [G/L] | 0.01 | 0.97 | −0.15 | 0.54 | −0.14 | 0.59 | −0.04 | 0.89 |
| BAS [%] | −0.27 | 0.28 | −0.02 | 0.94 | 0.05 | 0.84 | −0.27 | 0.28 |
| PLT [G/L] | 0.07 | 0.79 | 0.08 | 0.75 | 0.03 | 0.92 | 0.09 | 0.73 |
| PDW [fL] | 0.33 | 0.18 | −0.21 | 0.40 | −0.28 | 0.26 | 0.26 | 0.30 |
| PCT [%] | 0.18 | 0.48 | 0.01 | 0.97 | −0.06 | 0.80 | 0.17 | 0.49 |
| Large PLT [%] | 0.32 | 0.20 | −0.25 | 0.31 | −0.32 | 0.20 | 0.24 | 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanowska-Kocejko, M.; Jędrzejewska, A.; Braczko, A.; Stawarska, K.; Król, O.; Frańczak, M.; Harasim, G.; Smoleński, R.T.; Hellmann, M.; Kutryb-Zając, B. Red Blood Cell Adenylate Energetics Is Related to Endothelial and Microvascular Function in Long COVID. Biomedicines 2024, 12, 554. https://doi.org/10.3390/biomedicines12030554
Romanowska-Kocejko M, Jędrzejewska A, Braczko A, Stawarska K, Król O, Frańczak M, Harasim G, Smoleński RT, Hellmann M, Kutryb-Zając B. Red Blood Cell Adenylate Energetics Is Related to Endothelial and Microvascular Function in Long COVID. Biomedicines. 2024; 12(3):554. https://doi.org/10.3390/biomedicines12030554
Chicago/Turabian StyleRomanowska-Kocejko, Marzena, Agata Jędrzejewska, Alicja Braczko, Klaudia Stawarska, Oliwia Król, Marika Frańczak, Gabriela Harasim, Ryszard T. Smoleński, Marcin Hellmann, and Barbara Kutryb-Zając. 2024. "Red Blood Cell Adenylate Energetics Is Related to Endothelial and Microvascular Function in Long COVID" Biomedicines 12, no. 3: 554. https://doi.org/10.3390/biomedicines12030554