
Molecular and Cellular Involvement in CIPN




{kind=link}
{kind=link}
Abstract
1. Introduction
2. Underlying Mechanisms of CIPN
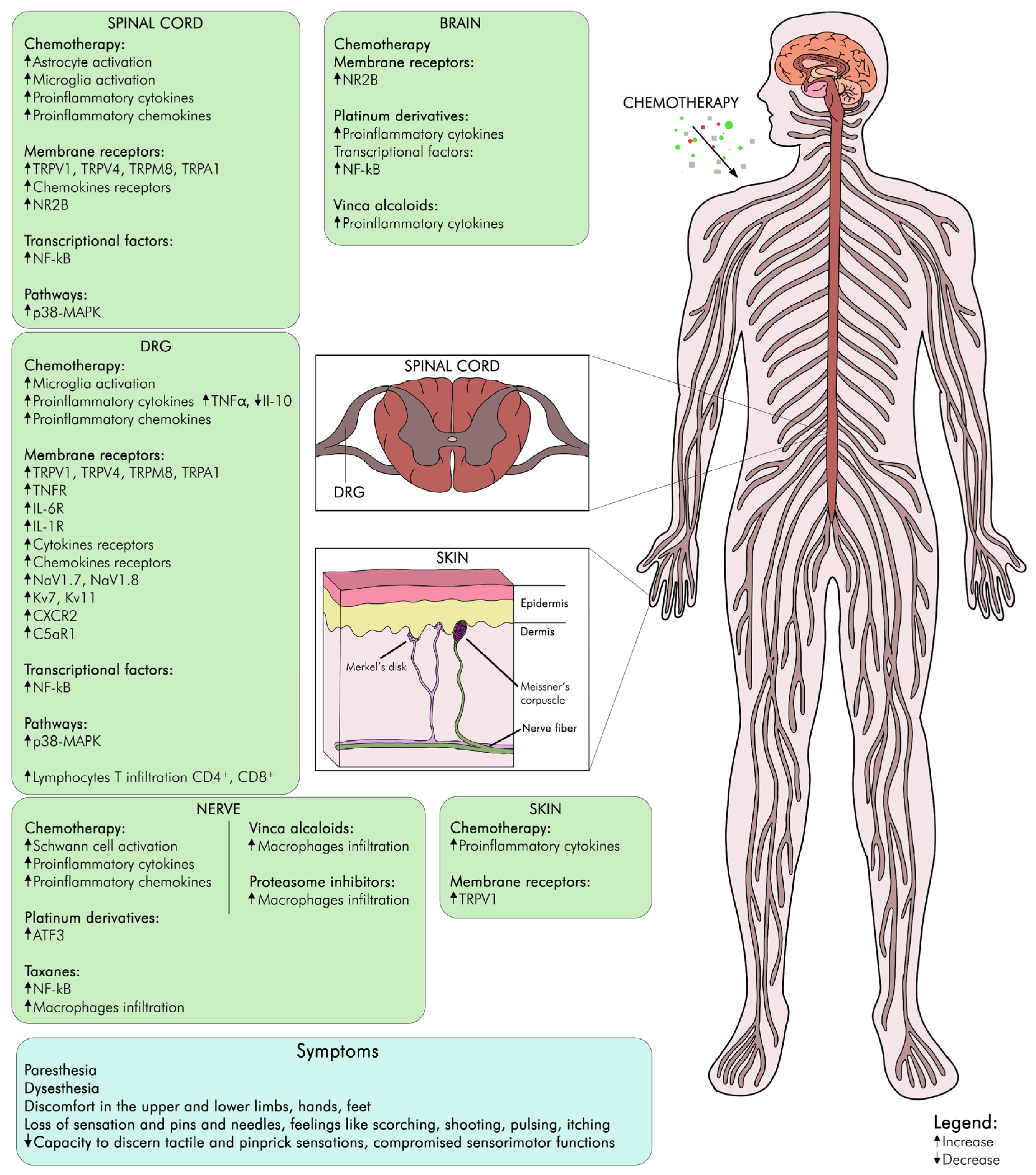
2.1. Neurophysiological Mechanisms of CIPN
2.2. Modifications in Transient Receptor Potential Channels in CIPN
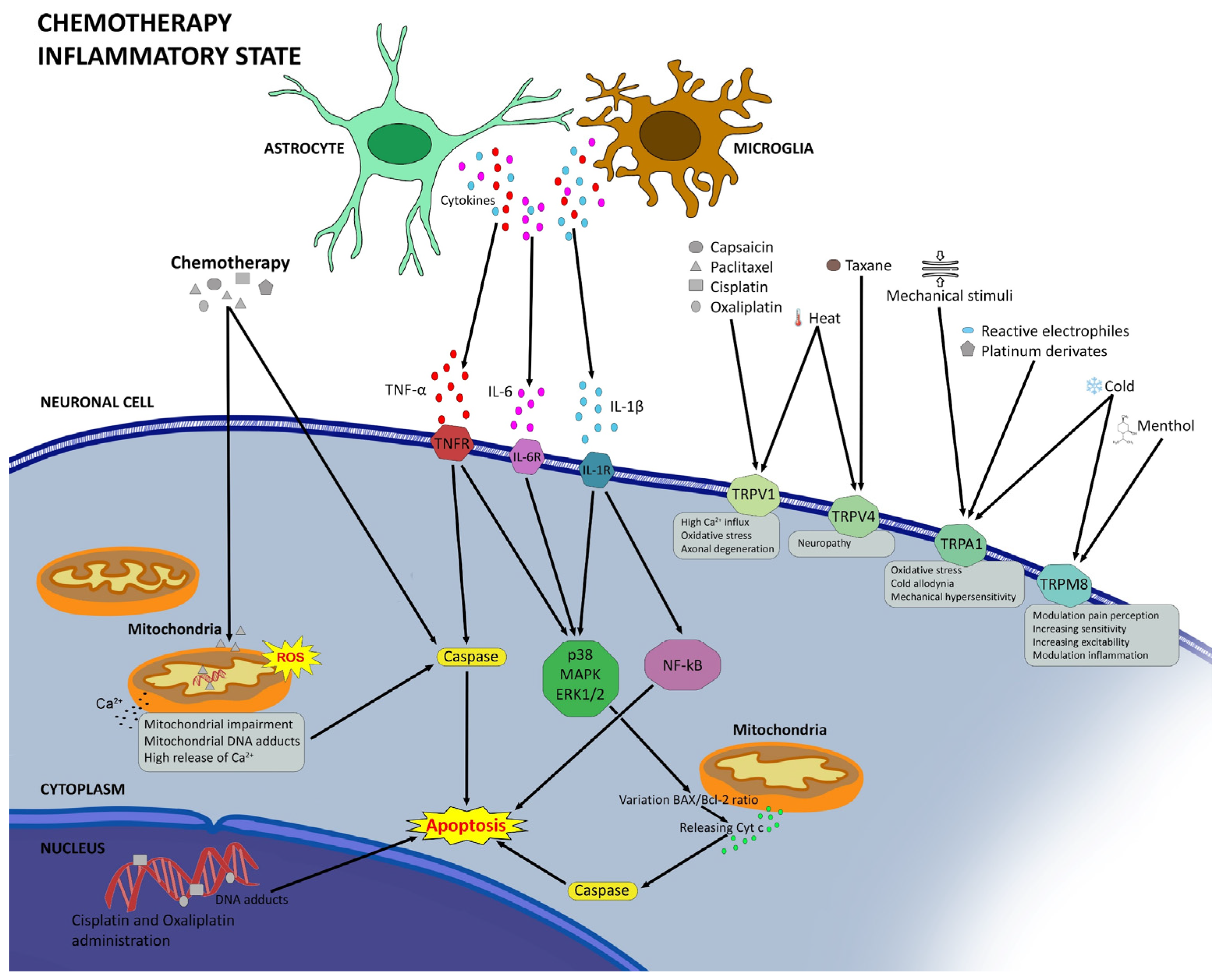
2.3. Neuroinflammation in CIPN
2.4. Oxidative Stress and Mitochondrial Dysfunction in CIPN
2.5. Degeneration of Intraepidermal Nerve Fibers in CIPN
2.6. Genetic Evidence of CIPN
3. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cho, Y.; Ruddy, K.J.; Lavoie Smith, E.M. Evaluation of Chemotherapy-Induced Peripheral Neuropathy. In Diagnosis, Management and Emerging Strategies for Chemotherapy-Induced Neuropathy: A MASCC Book; Lustberg, M., Loprinzi, C., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 53–93. [Google Scholar] [CrossRef]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-Induced Peripheral Neuropathy: A Current Review. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.; Cavaletti, G. Clinical and biochemical markers in CIPN: A reappraisal. Rev. Neurol. 2021, 177, 890–907. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; Palumbo, P.; D’Angelo, M.; Moorthy, N.K.; Antonosante, A.; Catanesi, M.; Lombardi, F.; Iannotta, D.; Cinque, B.; Benedetti, E.; et al. Probiotic DSF counteracts chemotherapy induced neuropathic pain. Oncotarget 2018, 9, 27998–28008. [Google Scholar] [CrossRef]
- Tay, N.; Laakso, E.-L.; Schweitzer, D.; Endersby, R.; Vetter, I.; Starobova, H. Chemotherapy-induced peripheral neuropathy in children and adolescent cancer patients. Front. Mol. Biosci. 2022, 9, 1015746. [Google Scholar] [CrossRef]
- Grisold, W.; Cavaletti, G.; Windebank, A.J. Peripheral neuropathies from chemotherapeutics and targeted agents: Diagnosis, treatment, and prevention. Neuro. Oncol. 2012, 14 (Suppl. S4), iv45–iv54. [Google Scholar] [CrossRef]
- Trendowski, M.R.; Lusk, C.M.; Ruterbusch, J.J.; Seaton, R.; Simon, M.S.; Greenwald, M.K.; Harper, F.W.K.; Beebe-Dimmer, J.L.; Schwartz, A.G. Chemotherapy-induced peripheral neuropathy in African American cancer survivors: Risk factors and quality of life outcomes. Cancer Med. 2021, 10, 8151–8161. [Google Scholar] [CrossRef] [PubMed]
- Progress in Cancer Survival, Mortality, and Incidence in Seven High-Income Countries 1995–2014 (ICBP SURVMARK-2): A Population-Based Study—The Lancet Oncology. Available online: https://www.thelancet.com/journals/lanonc/article/PIIS1470-2045(19)30456-5/fulltext (accessed on 1 February 2024).
- Cancer Statistics, 2019—Siegel—2019—CA: A Cancer Journal for Clinicians—Wiley Online Library. Available online: https://acsjournals.onlinelibrary.wiley.com/doi/full/10.3322/caac.21551 (accessed on 1 February 2024).
- Henley, S.J.; Singh, S.D.; King, J.; Wilson, R.J.; O’Neil, M.E.; Ryerson, A.B. Invasive Cancer Incidence and Survival—United States, 2012. Morb. Mortal. Wkly. Rep. 2015, 64, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Argyriou, A.A.; Bruna, J.; Marmiroli, P.; Cavaletti, G. Chemotherapy-induced peripheral neurotoxicity (CIPN): An update. Crit. Rev. Oncol. Hematol. 2012, 82, 51–77. [Google Scholar] [CrossRef]
- Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/pii/S0304395914004436 (accessed on 1 February 2024).
- Colvin, L.A. Chemotherapy-induced peripheral neuropathy (CIPN): Where are we now? Pain 2019, 160, S1–S10. [Google Scholar] [CrossRef]
- Alberti, P.; Bernasconi, D.P.; Cornblath, D.R.; Merkies, I.S.; Park, S.B.; Velasco, R.; Bruna, J.; Psimaras, D.; Koeppen, S.; Pace, A.; et al. Prospective Evaluation of Health Care Provider and Patient Assessments in Chemotherapy-Induced Peripheral Neurotoxicity. Neurology 2021, 97, e660–e672. [Google Scholar] [CrossRef]
- Molassiotis, A.; Cheng, H.L.; Lopez, V.; Au, J.S.K.; Chan, A.; Bandla, A.; Leung, K.T.; Li, Y.C.; Wong, K.H.; Suen, L.K.P.; et al. Are we mis-estimating chemotherapy-induced peripheral neuropathy? Analysis of assessment methodologies from a prospective, multinational, longitudinal cohort study of patients receiving neurotoxic chemotherapy. BMC Cancer 2019, 19, 132. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kwok, J.; Asher, R.; Lee, C.; Beale, P.; Selle, F.; Friedlander, M. Clinical and genetic predictors of paclitaxel neurotoxicity based on patient- versus clinician-reported incidence and severity of neurotoxicity in the ICON7 trial. Ann. Oncol. 2017, 28, 2733–2740. [Google Scholar] [CrossRef] [PubMed]
- Yeo, F.; Ng, C.C.; Loh, K.W.J.; Molassiotis, A.; Cheng, H.L.; Au, J.S.K.; Leung, K.T.; Li, Y.C.; Wong, K.-H.; Suen, L.; et al. Minimal clinically important difference of the EORTC QLQ-CIPN20 for worsening peripheral neuropathy in patients receiving neurotoxic chemotherapy. Support. Care Cancer 2019, 27, 4753–4762. [Google Scholar] [CrossRef]
- Kaiser, K.; Lyleroehr, M.; Shaunfield, S.; Lacson, L.; Corona, M.; Kircher, S.; Nittve, M.; Cella, D. Neuropathy experienced by colorectal cancer patients receiving oxaliplatin: A qualitative study to validate the Functional Assessment of Cancer Therapy/Gynecologic Oncology Group-Neurotoxicity scale. World J. Gastrointest. Oncol. 2020, 12, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Soveri, L.M.; Lamminmäki, A.; Hänninen, U.A.; Karhunen, M.; Bono, P.; Osterlund, P. Long-term neuropathy and quality of life in colorectal cancer patients treated with oxaliplatin containing adjuvant chemotherapy. Acta Oncol. 2019, 58, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Thawani, S.P.; Tanji, K.; De Sousa, E.A.; Weimer, L.H.; Brannagan, T.H. Bortezomib-associated demyelinating neuropathy—clinical and pathologic features. J. Clin. Neuromuscul. Dis. 2015, 16, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef]
- Desforges, A.D.; Hebert, C.M.; Spence, A.L.; Reid, B.; Dhaibar, H.A.; Cruz-Topete, D.; Cornett, E.M.; Kaye, A.D.; Urits, I.; Viswanath, O. Treatment and diagnosis of chemotherapy-induced peripheral neuropathy: An update. Biomed. Pharmacother. 2022, 147, 112671. [Google Scholar] [CrossRef]
- Min, H.-Y.; Lee, H.-Y. Molecular targeted therapy for anticancer treatment. Exp. Mol. Med. 2022, 54, 1670. [Google Scholar] [CrossRef]
- Zahavi, D.; Weiner, L. Monoclonal Antibodies in Cancer Therapy. Antibodies 2020, 9, 34. [Google Scholar] [CrossRef]
- Omran, M.; Belcher, E.K.; Mohile, N.A.; Kesler, S.R.; Janelsins, M.C.; Hohmann, A.G.; Kleckner, I.R. Review of the Role of the Brain in Chemotherapy-Induced Peripheral Neuropathy. Front. Mol. Biosci. 2021, 8, 693133. [Google Scholar] [CrossRef] [PubMed]
- Isom, L.L.; Patino, G.A.; Yuan, Y.; Lopez-Santiago, L. Voltage-Gated Ion Channels. In Neuroscience in the 21st Century: From Basic to Clinical; Pfaff, D.W., Volkow, N.D., Rubenstein, J., Eds.; Springer: New York, NY, USA, 2020; pp. 1–28. [Google Scholar] [CrossRef]
- Verma, P.; Eaton, M.; Kienle, A.; Flockerzi, D.; Yang, Y.; Ramkrishna, D. Examining Sodium and Potassium Channel Conductances Involved in Hyperexcitability of Chemotherapy-Induced Peripheral Neuropathy: A Mathematical and Cell Culture-Based Study. Front. Comput. Neurosci. 2020, 14, 564980. [Google Scholar] [CrossRef] [PubMed]
- Urru, M.; Muzzi, M.; Coppi, E.; Ranieri, G.; Buonvicino, D.; Camaioni, E.; Coppini, R.; Pugliese, A.M.; Tanaka, B.; Estacion, M.; et al. Dexpramipexole blocks Nav1.8 sodium channels and provides analgesia in multiple nociceptive and neuropathic pain models. Pain 2020, 161, 831. [Google Scholar] [CrossRef]
- Kang, L.; Tian, Y.; Xu, S.; Chen, H. Oxaliplatin-induced peripheral neuropathy: Clinical features, mechanisms, prevention and treatment. J. Neurol. 2021, 268, 3269–3282. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, A.G.; Dimitrova, N.A. A possible link of oxaliplatin-induced neuropathy with potassium channel deficit. Muscle Nerve 2012, 45, 403–411. [Google Scholar] [CrossRef]
- Becker, G.; Atuati, S.F.; Oliveira, S.M. G Protein-Coupled Receptors and Ion Channels Involvement in Cisplatin-Induced Peripheral Neuropathy: A Review of Preclinical Studies. Cancers 2024, 16, 580. [Google Scholar] [CrossRef] [PubMed]
- de Castro Fonseca, M.; Marazzi-Diniz, P.H.S.; Leite, M.F.; Ehrlich, B.E. Calcium signaling in chemotherapy-induced neuropathy. Cell Calcium 2023, 113, 102762. [Google Scholar] [CrossRef]
- Bae, E.H.; Greenwald, M.K.; Schwartz, A.G. Chemotherapy-Induced Peripheral Neuropathy: Mechanisms and Therapeutic Avenues. Neurotherapeutics 2021, 18, 2384–2396. [Google Scholar] [CrossRef]
- Li, Y.; Tatsui, C.E.; Rhines, L.D.; North, R.Y.; Harrison, D.S.; Cassidy, R.M.; Johansson, C.A.; Kosturakis, A.K.; Edwards, D.D.; Zhang, H.; et al. Dorsal root ganglion neurons become hyperexcitable and increase expression of voltage-gated T-type calcium channels (Cav3.2) in paclitaxel-induced peripheral neuropathy. Pain 2017, 158, 417–429. [Google Scholar] [CrossRef]
- Grothey, A. Clinical management of oxaliplatin-associated neurotoxicity. Clin. Color. Cancer 2005, 5 (Suppl. S1), S38–S46. [Google Scholar] [CrossRef]
- Alles, S.R.A.; Smith, P.A. Peripheral Voltage-Gated Cation Channels in Neuropathic Pain and Their Potential as Therapeutic Targets. Front. Pain Res. 2021, 2, 750583. [Google Scholar] [CrossRef] [PubMed]
- Labau, J.I.R.; Alsaloum, M.; Estacion, M.; Tanaka, B.; Dib-Hajj, F.B.; Lauria, G.; Smeets, H.J.M.; Faber, C.G.; Dib-Hajj, S.; Waxman, S.G. Lacosamide Inhibition of NaV1.7 Channels Depends on its Interaction with the Voltage Sensor Domain and the Channel Pore. Front. Pharmacol. 2021, 12, 791740. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. Pharmacological Inhibition of Voltage-gated Ca2+ Channels for Chronic Pain Relief. Curr. Neuropharmacol. 2013, 11, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Nodera, H.; Spieker, A.; Sung, M.; Rutkove, S. Neuroprotective effects of Kv7 channel agonist, retigabine, for cisplatin-induced peripheral neuropathy. Neurosci. Lett. 2011, 505, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M.; Braidy, N. Thermo-Sensitive TRP Channels: Novel Targets for Treating Chemotherapy-Induced Peripheral Pain. Front. Physiol. 2017, 8, 1040. [Google Scholar] [CrossRef] [PubMed]
- Brandolini, L.; D’angelo, M.; Novelli, R.; Castelli, V.; Giorgio, C.; Sirico, A.; Cocchiaro, P.; D’egidio, F.; Benedetti, E.; Cristiano, C.; et al. Paclitaxel binds and activates C5aR1: A new potential therapeutic target for the prevention of chemotherapy-induced peripheral neuropathy and hypersensitivity reactions. Cell Death Dis. 2022, 13, 500. [Google Scholar] [CrossRef]
- Chukyo, A.; Chiba, T.; Kambe, T.; Yamamoto, K.; Kawakami, K.; Taguchi, K.; Abe, K. Oxaliplatin-induced changes in expression of transient receptor potential channels in the dorsal root ganglion as a neuropathic mechanism for cold hypersensitivity. Neuropeptides 2018, 67, 95–101. [Google Scholar] [CrossRef]
- Cristiano, C.; Giorgio, C.; Cocchiaro, P.; Boccella, S.; Cesta, M.; Castelli, V.; Liguori, F.; Cuozzo, M.; Brandolini, L.; Russo, R.; et al. Inhibition of C5aR1 as a promising approach to treat taxane-induced neuropathy. Cytokine 2023, 171, 156370. [Google Scholar] [CrossRef]
- avia-Pelaez, J.M.; Lemes, J.B.P.; Gonzalez, L.; Delay, L.; Capettini, L.d.S.A.; Lu, J.W.; Dos Santos, G.G.; Gregus, A.M.; Dougherty, P.M.; Yaksh, T.L.; et al. AIBP regulates TRPV1 activation in chemotherapy-induced peripheral neuropathy by controlling lipid raft dynamics and proximity to TLR4 in dorsal root ganglion neurons. Pain 2023, 164, e274–e285. [Google Scholar] [CrossRef]
- Nassini, R.; Gees, M.; Harrison, S.; De Siena, G.; Materazzi, S.; Moretto, N.; Failli, P.; Preti, D.; Marchetti, N.; Cavazzini, A.; et al. Oxaliplatin elicits mechanical and cold allodynia in rodents via TRPA1 receptor stimulation. Pain 2011, 152, 1621–1631. [Google Scholar] [CrossRef]
- Ochoa, S.V.; Casas, Z.; Albarracín, S.L.; Sutachan, J.J.; Torres, Y.P. Therapeutic potential of TRPM8 channels in cancer treatment. Front. Pharmacol. 2023, 14, 1098448. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, G.; Monza, L.; Cavaletti, G.; Rigolio, R.; Meregalli, C. Neuroinflammatory Process Involved in Different Preclinical Models of Chemotherapy-Induced Peripheral Neuropathy. Front. Immunol. 2021, 11, 626687. [Google Scholar] [CrossRef] [PubMed]
- Brandolini, L.; d’Angelo, M.; Antonosante, A.; Cimini, A.; Allegretti, M. Chemokine Signaling in Chemotherapy-Induced Neuropathic Pain. Int. J. Mol. Sci. 2019, 20, 2904. [Google Scholar] [CrossRef]
- Lees, J.G.; Makker, P.G.; Tonkin, R.S.; Abdulla, M.; Park, S.B.; Goldstein, D.; Moalem-Taylor, G. Immune-mediated processes implicated in chemotherapy-induced peripheral neuropathy. Eur. J. Cancer 2017, 73, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Makker, P.G.S.; Duffy, S.S.; Lees, J.G.; Perera, C.J.; Tonkin, R.S.; Butovsky, O.; Park, S.B.; Goldstein, D.; Moalem-Taylor, G. Characterisation of Immune and Neuroinflammatory Changes Associated with Chemotherapy-Induced Peripheral Neuropathy. PLoS ONE 2017, 12, e0170814. [Google Scholar] [CrossRef]
- Bráz, J.M.; Basbaum, A.I. Differential ATF3 expression in dorsal root ganglion neurons reveals the profile of primary afferents engaged by diverse noxious chemical stimuli. Pain 2010, 150, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.-W.; Cheng, K.-I.; Chen, J.-Y.; Cheng, Y.-C.; Chang, Y.-C.; Yeh, J.-L.; Hsu, J.-H.; Dai, Z.-K.; Wu, B.-N. Loganin prevents chronic constriction injury-provoked neuropathic pain by reducing TNF-α/IL-1β-mediated NF-κB activation and Schwann cell demyelination. Phytomedicine 2020, 67, 153166. [Google Scholar] [CrossRef]
- Li, G.-Z.; Hu, Y.-H.; Li, D.-Y.; Zhang, Y.; Guo, H.-L.; Li, Y.-M.; Chen, F.; Xu, J. Vincristine-induced peripheral neuropathy: A mini-review. NeuroToxicology 2020, 81, 161–171. [Google Scholar] [CrossRef]
- Silva, R.L.; Lopes, A.H.; Guimarães, R.M.; Cunha, T.M. CXCL1/CXCR2 signaling in pathological pain: Role in peripheral and central sensitization. Neurobiol. Dis. 2017, 105, 109–116. [Google Scholar] [CrossRef]
- Singh, G.; Singh, A.; Singh, P.; Bhatti, R. Bergapten Ameliorates Vincristine-Induced Peripheral Neuropathy by Inhibition of Inflammatory Cytokines and NFκB Signaling. ACS Chem. Neurosci. 2019, 10, 3008–3017. [Google Scholar] [CrossRef]
- Liu, L.-R.; Liu, J.-C.; Bao, J.-S.; Bai, Q.-Q.; Wang, G.-Q. Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front. Immunol. 2020, 11, 1024. [Google Scholar] [CrossRef]
- Klein, I.; Boenert, J.; Lange, F.; Christensen, B.; Wassermann, M.K.; Wiesen, M.H.J.; Olschewski, D.N.; Rabenstein, M.; Müller, C.; Lehmann, H.C.; et al. Glia from the central and peripheral nervous system are differentially affected by paclitaxel chemotherapy via modulating their neuroinflammatory and neuroregenerative properties. Front. Pharmacol. 2022, 13, 1038285. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Y.; Xia, R.; Wei, Y.; Wei, X. Role of the CCL2-CCR2 signalling axis in cancer: Mechanisms and therapeutic targeting. Cell Prolif. 2021, 54, e13115. [Google Scholar] [CrossRef] [PubMed]
- Brandolini, L.; Castelli, V.; Aramini, A.; Giorgio, C.; Bianchini, G.; Russo, R.; De Caro, C.; D’angelo, M.; Catanesi, M.; Benedetti, E.; et al. DF2726A, a new IL-8 signalling inhibitor, is able to counteract chemotherapy-induced neuropathic pain. Sci. Rep. 2019, 9, 11729. [Google Scholar] [CrossRef] [PubMed]
- Laura, B.; Elisabetta, B.; Adelchi, R.P.; Roberto, R.; Loredana, C.; Andrea, A.; Michele, D.; Vanessa, C.; Antonio, G.; Marcello, A.; et al. CXCR1/2 pathways in paclitaxel-induced neuropathic pain. Oncotarget 2017, 8, 23188–23201. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.H.; Bennett, G.J. Effects of mitochondrial poisons on the neuropathic pain produced by the chemotherapeutic agents, paclitaxel and oxaliplatin. Pain 2012, 153, 704–709. [Google Scholar] [CrossRef]
- Doyle, T.M.; Salvemini, D. Mini-Review: Mitochondrial Dysfunction and Chemotherapy-induced Neuropathic Pain. Neurosci. Lett. 2021, 760, 136087. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xiao, W.H.; Bennett, G.J. Mitotoxicity and bortezomib-induced chronic painful peripheral neuropathy. Exp. Neurol. 2012, 238, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Canta, A.; Pozzi, E.; Carozzi, V.A. Mitochondrial Dysfunction in Chemotherapy-Induced Peripheral Neuropathy (CIPN). Toxics 2015, 3, 198–223. [Google Scholar] [CrossRef]
- Marullo, R.; Werner, E.; Degtyareva, N.; Moore, B.; Altavilla, G.; Ramalingam, S.S.; Doetsch, P.W. Cisplatin induces a mitochondrial-ROS response that contributes to cytotoxicity depending on mitochondrial redox status and bioenergetic functions. PLoS ONE 2013, 8, e81162. [Google Scholar] [CrossRef]
- Malacrida, A.; Semperboni, S.; Di Domizio, A.; Palmioli, A.; Broggi, L.; Airoldi, C.; Meregalli, C.; Cavaletti, G.; Nicolini, G. Tubulin binding potentially clears up Bortezomib and Carfilzomib differential neurotoxic effect. Sci. Rep. 2021, 11, 10523. [Google Scholar] [CrossRef] [PubMed]
- Broyl, A.; Corthals, S.L.; Jongen, J.L.; van der Holt, B.; Kuiper, R.; de Knegt, Y.; van Duin, M.; el Jarari, L.; Bertsch, U.; Lokhorst, H.M.; et al. Mechanisms of peripheral neuropathy associated with bortezomib and vincristine in patients with newly diagnosed multiple myeloma: A prospective analysis of data from the HOVON-65/GMMG-HD4 trial. Lancet Oncol. 2010, 11, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Akhilesh Uniyal, A.; Mehta, A.; Tiwari, V. Combination chemotherapy in rodents: A model for chemotherapy-induced neuropathic pain and pharmacological screening. Metab. Brain Dis. 2024, 39, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Zhang, Y.P.; Gwak, Y.S.; Abdi, S. Phenyl N-tert-butylnitrone, a free radical scavenger, reduces mechanical allodynia in chemotherapy-induced neuropathic pain in rats. Anesthesiology 2010, 112, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Fidanboylu, M.; Griffiths, L.A.; Flatters, S.J.L. Global inhibition of reactive oxygen species (ROS) inhibits paclitaxel-induced painful peripheral neuropathy. PLoS ONE 2011, 6, e25212. [Google Scholar] [CrossRef]
- Doyle, T.; Chen, Z.; Muscoli, C.; Bryant, L.; Esposito, E.; Cuzzocrea, S.; Dagostino, C.; Ryerse, J.; Rausaria, S.; Kamadulski, A.; et al. Targeting the Overproduction of Peroxynitrite for the Prevention and Reversal of Paclitaxel-Induced Neuropathic Pain. J. Neurosci. 2012, 32, 6149–6160. [Google Scholar] [CrossRef]
- Silva Santos Ribeiro, P.; Willemen, H.L.D.M.; Eijkelkamp, N. Mitochondria and sensory processing in inflammatory and neuropathic pain. Front. Pain Res. 2022, 3, 1013577. [Google Scholar] [CrossRef]
- Shin, Y.K.; Jang, S.Y.; Lee, H.K.; Jung, J.; Suh, D.J.; Seo, S.; Park, H.T. Pathological adaptive responses of Schwann cells to endoplasmic reticulum stress in bortezomib-induced peripheral neuropathy. Glia 2010, 58, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Malacrida, A.; Meregalli, C.; Rodriguez-Menendez, V.; Nicolini, G. Chemotherapy-Induced Peripheral Neuropathy and Changes in Cytoskeleton. Int. J. Mol. Sci. 2019, 20, 2287. [Google Scholar] [CrossRef]
- LaPointe, N.E.; Morfini, G.; Brady, S.T.; Feinstein, S.C.; Wilson, L.; Jordan, M.A. Effects of eribulin, vincristine, paclitaxel and ixabepilone on fast axonal transport and kinesin-1 driven microtubule gliding: Implications for chemotherapy-induced peripheral neuropathy. Neurotoxicology 2013, 37, 231–239. [Google Scholar] [CrossRef]
- Staff, N.P.; Podratz, J.L.; Grassner, L.; Bader, M.; Paz, J.; Knight, A.M.; Loprinzi, C.L.; Trushina, E.; Windebank, A.J. Bortezomib alters microtubule polymerization and axonal transport in rat dorsal root ganglion neurons. Neurotoxicology 2013, 39, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Schellingerhout, D.; LeRoux, L.G.; Hobbs, B.P.; Bredow, S. Impairment of retrograde neuronal transport in oxaliplatin-induced neuropathy demonstrated by molecular imaging. PLoS ONE 2012, 7, e45776. [Google Scholar] [CrossRef]
- Čermák, V.; Dostál, V.; Jelínek, M.; Libusová, L.; Kovář, J.; Rösel, D.; Brábek, J. Microtubule-targeting agents and their impact on cancer treatment. Eur. J. Cell Biol. 2020, 99, 151075. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Magiera, M.M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 307–326. [Google Scholar] [CrossRef]
- Wordeman, L.; Vicente, J.J. Microtubule Targeting Agents in Disease: Classic Drugs, Novel Roles. Cancers 2021, 13, 5650. [Google Scholar] [CrossRef] [PubMed]
- Amjad, M.T.; Chidharla, A.; Kasi, A. Cancer Chemotherapy. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- O’Dowd, P.D.; Sutcliffe, D.F.; Griffith, D.M. Oxaliplatin and its derivatives—An overview. Coord. Chem. Rev. 2023, 497, 215439. [Google Scholar] [CrossRef]
- Benvenuto, M.; Ciuffa, S.; Focaccetti, C.; Sbardella, D.; Fazi, S.; Scimeca, M.; Tundo, G.R.; Barillari, G.; Segni, M.; Bonanno, E.; et al. Proteasome inhibition by bortezomib parallels a reduction in head and neck cancer cells growth, and an increase in tumor-infiltrating immune cells. Sci. Rep. 2021, 11, 19051. [Google Scholar] [CrossRef]
- Zhang, D.; Kanakkanthara, A. Beyond the Paclitaxel and Vinca Alkaloids: Next Generation of Plant-Derived Microtubule-Targeting Agents with Potential Anticancer Activity. Cancers 2020, 12, 1721. [Google Scholar] [CrossRef] [PubMed]
- Zafari, N.; Velayati, M.; Maftooh, M.; Khazaei, M.; Nassiri, M.; Hassanian, S.M.; Ghayour-Mobarhan, M.; Ferns, G.A.; Avan, A. Mechanism-based Pharmacological Management of Chemotherapy-induced Neuropathic Pain from Preclinical Studies to Clinical Prospective: Platinum-based Drugs, Taxanes, and Vinca Alkaloids. Curr. Pharm. Des. 2023, 29, 1245–1265. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Kavelaars, A.; Dougherty, P.M.; Heijnen, C.J. Beyond symptomatic relief for chemotherapy-induced peripheral neuropathy: Targeting the source. Cancer 2018, 124, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.; Hu, H.; Zheng, Y.; Wu, B.; Sun, W. PGC1α: An emerging therapeutic target for chemotherapy-induced peripheral neuropathy. Ther. Adv. Neurol. Disord. 2023, 16, 17562864231163361. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, H.; Wang, J.; Liu, Y.; Xu, Y.; Xu, H.; Feng, Y.; Ge, W. The Therapeutic Potential of Antioxidants in Chemotherapy-Induced Peripheral Neuropathy: Evidence from Preclinical and Clinical Studies. Neurotherapeutics 2023, 20, 339–358. [Google Scholar] [CrossRef]
- Cuozzo, M.; Castelli, V.; Avagliano, C.; Cimini, A.; D’angelo, M.; Cristiano, C.; Russo, R. Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain. Biomedicines 2021, 9, 346. [Google Scholar] [CrossRef]
- García-Mesa, Y.; García-Piqueras, J.; Cobo, R.; Martín-Cruces, J.; Suazo, I.; García-Suárez, O.; Feito, J.; Vega, J.A. Sensory innervation of the human male prepuce: Meissner’s corpuscles predominate. J. Anat. 2021, 239, 892–902. [Google Scholar] [CrossRef]
- Cobo, R.; García-Piqueras, J.; Cobo, J.; Vega, J.A. The Human Cutaneous Sensory Corpuscles: An Update. J. Clin. Med. 2021, 10, 227. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.A.; Cata, J.P.; Zhang, H.; Driver, L.C.; Wendelschafer-Crabb, G.; Kennedy, W.R.; Dougherty, P.M. Follow-up psychophysical studies in bortezomib-related chemoneuropathy patients. J. Pain 2011, 12, 1017–1024. [Google Scholar] [CrossRef]
- Boyette-Davis, J.; Xin, W.; Zhang, H.; Dougherty, P.M. Intraepidermal nerve fiber loss corresponds to the development of taxol-induced hyperalgesia and can be prevented by treatment with minocycline. Pain 2011, 152, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.; Dougherty, P.M. Protection against oxaliplatin-induced mechanical hyperalgesia and intraepidermal nerve fiber loss by minocycline. Exp. Neurol. 2011, 229, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, H.C.; Chen, W.; Borzan, J.; Mankowski, J.L.; Höke, A. Mitochondrial dysfunction in distal axons contributes to human immunodeficiency virus sensory neuropathy. Ann. Neurol. 2011, 69, 100–110. [Google Scholar] [CrossRef]
- Oaklander, A.L. The density of remaining nerve endings in human skin with and without postherpetic neuralgia after shingles. Pain 2001, 92, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Boyette-Davis, J.A.; Kosturakis, A.K.; Li, Y.; Yoon, S.-Y.; Walters, E.T.; Dougherty, P.M. Induction of monocyte chemoattractant protein-1 (MCP-1) and its receptor CCR2 in primary sensory neurons contributes to paclitaxel-induced peripheral neuropathy. J. Pain 2013, 14, 1031–1044. [Google Scholar] [CrossRef]
- Timmins, H. Chemotherapy-Induced Peripheral Neuropathy: Assessment, Phenotypes and Risk Factors. Ph.D. Thesis, The University of Sydney, Sydney, Australia, 2021. [Google Scholar]
- Rodwin, R.L.; Siddiq, N.Z.; Ehrlich, B.E.; Lustberg, M.B. Biomarkers of Chemotherapy-Induced Peripheral Neuropathy: Current Status and Future Directions. Front. Pain Res. 2022, 3, 864910. [Google Scholar] [CrossRef] [PubMed]
- Stoehlmacher, J.; Park, D.J.; Zhang, W.; Groshen, S.; Tsao-Wei, D.D.; Yu, M.C.; Lenz, H.-J. Association between glutathione S-transferase P1, T1, and M1 genetic polymorphism and survival of patients with metastatic colorectal cancer. J. Natl. Cancer Inst. 2002, 94, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tzeng, C.; Chen, P.; Lin, J.; Lin, T.; Chen, W.; Jiang, J.; Wang, H.; Wang, W. Influence of GSTP1 I105V polymorphism on cumulative neuropathy and outcome of FOLFOX-4 treatment in Asian patients with colorectal carcinoma. Cancer Sci. 2010, 101, 530–535. [Google Scholar] [CrossRef]
- Kanai, M.; Yoshioka, A.; Tanaka, S.; Nagayama, S.; Matsumoto, S.; Nishimura, T.; Niimi, M.; Teramukai, S.; Takahashi, R.; Mori, Y.; et al. Associations between glutathione S-transferase pi Ile105Val and glyoxylate aminotransferase Pro11Leu and Ile340Met polymorphisms and early-onset oxaliplatin-induced neuropathy. Cancer Epidemiol. 2010, 34, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; Lee, J.; Park, J.O.; Park, Y.S.; Lim, H.Y.; Kang, W.K.; Kim, J.; Lee, S.; Park, S.H. Polymorphic markers associated with severe oxaliplatin-induced, chronic peripheral neuropathy in colon cancer patients. Cancer 2012, 118, 2828–2836. [Google Scholar] [CrossRef] [PubMed]
- Beutler, A.S.; Kulkarni, A.A.; Kanwar, R.; Klein, C.J.; Therneau, T.M.; Qin, R.; Banck, M.S.; Boora, G.K.; Ruddy, K.J.; Wu, Y.; et al. Sequencing of Charcot-Marie-Tooth disease genes in a toxic polyneuropathy. Ann. Neurol. 2014, 76, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Adjei, A.A.; Lopez, C.L.; Schaid, D.J.; Sloan, J.A.; Le-Rademacher, J.G.; Loprinzi, C.L.; Norman, A.D.; Olson, J.E.; Couch, F.J.; Beutler, A.S.; et al. Genetic Predictors of Chemotherapy-Induced Peripheral Neuropathy from Paclitaxel, Carboplatin and Oxaliplatin: NCCTG/Alliance N08C1, N08CA and N08CB Study. Cancers 2021, 13, 1084. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudpour, S.H.; Bandapalli, O.R.; da Silva Filho, M.I.; Campo, C.; Hemminki, K.; Goldschmidt, H.; Merz, M. Chemotherapy-Induced Peripheral Neuropathy: Evidence from Genome-Wide Association Studies and Replication Within Multiple Myeloma Patients. BMC Cancer 2018, 18, 820. [Google Scholar] [CrossRef]
- Zhi, W.I.; Baser, R.E.; Kwon, A.; Chen, C.; Li, S.Q.; Piulson, L.; Seluzicki, C.; Panageas, K.S.; Harte, S.E.; Mao, J.J.; et al. Characterization of Chemotherapy-Induced Peripheral Neuropathy Using Patient-Reported Outcomes and Quantitative Sensory Testing. Breast Cancer Res. Treat. 2021, 186, 761–768. [Google Scholar] [CrossRef]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of Chemotherapy-Induced Peripheral Neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef]
- Behrouzi, A.; Xia, H.; Thompson, E.L.; Kelley, M.R.; Fehrenbacher, J.C. Oxidative DNA Damage and Cisplatin Neurotoxicity Is Exacerbated by Inhibition of OGG1 Glycosylase Activity and APE1 Endonuclease Activity in Sensory Neurons. Int. J. Mol. Sci. 2022, 23, 1909. [Google Scholar] [CrossRef]
- Di Maio, G.; Villano, I.; Ilardi, C.R.; Messina, A.; Monda, V.; Iodice, A.C.; Porro, C.; Panaro, M.A.; Chieffi, S.; Messina, G.; et al. Mechanisms of Transmission and Processing of Pain: A Narrative Review. Int. J. Environ. Res. Public Health 2023, 20, 3064. [Google Scholar] [CrossRef] [PubMed]
- Tarasiuk, O.; Molteni, L.; Malacrida, A.; Nicolini, G. The Role of NMNAT2/SARM1 in Neuropathy Development. Biology 2024, 13, 61. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kacem, H.; Cimini, A.; d’Angelo, M.; Castelli, V. Molecular and Cellular Involvement in CIPN. Biomedicines 2024, 12, 751. https://doi.org/10.3390/biomedicines12040751
Kacem H, Cimini A, d’Angelo M, Castelli V. Molecular and Cellular Involvement in CIPN. Biomedicines. 2024; 12(4):751. https://doi.org/10.3390/biomedicines12040751
Chicago/Turabian StyleKacem, Housem, Annamaria Cimini, Michele d’Angelo, and Vanessa Castelli. 2024. "Molecular and Cellular Involvement in CIPN" Biomedicines 12, no. 4: 751. https://doi.org/10.3390/biomedicines12040751
APA StyleKacem, H., Cimini, A., d’Angelo, M., & Castelli, V. (2024). Molecular and Cellular Involvement in CIPN. Biomedicines, 12(4), 751. https://doi.org/10.3390/biomedicines12040751