

Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction?

Abstract
:1. Introduction
2. Lower Urinary Tract Symptoms (LUTS) and Overactive Bladder (OAB) Syndrome
3. Association of Metabolic Syndrome and Diabetes with Urinary Bladder Dysfunction
4. Bladder Dysfunction in Type 1 and Type 2 Diabetes in Patients and Animals
5. Methylglyoxal–Advanced Glycation End Products (AGEs)–RAGE Signaling Pathway
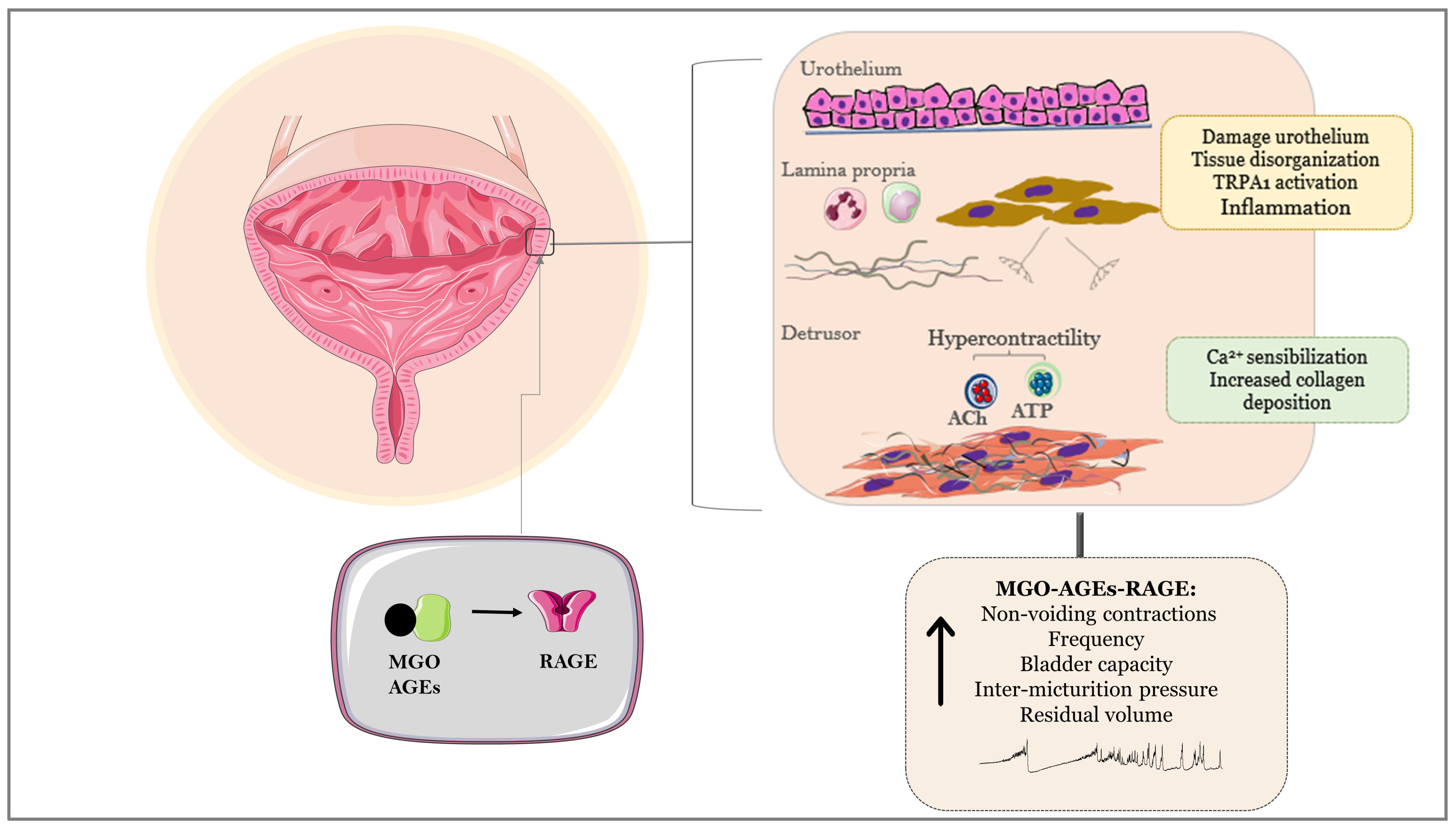
6. MGO–AGEs–RAGE Axis as a Key Player of Bladder Dysfunction in Animals and Humans
7. Drugs Presenting Potential to Downregulate AGEs Formation and Oxidative Stress in Bladder Tissues
7.1. Polyphenols: Resveratrol and Epigallocatechin-3-Gallate
7.2. Metformin
8. Concluding Remarks and Future Therapeutics
Author Contributions
Funding
Conflicts of Interest
References
- Rabbani, N.; Xue, M.; Thornalley, P.J. Dicarbonyl stress, protein glycation and the unfolded protein response. Glycoconj. J. 2021, 38, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Stratmann, B. Dicarbonyl Stress in Diabetic Vascular Disease. Int. J. Mol. Sci. 2022, 23, 6186. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeong, M.S.; Jang, S.B. Molecular characteristics of RAGE and advances in small-molecule inhibitors. Int. J. Mol. Sci. 2021, 22, 6904. [Google Scholar] [CrossRef]
- Abrams, P.; Cardozo, L.; Fall, M.; Griffiths, D.; Rosier, P.; Ulmsten, U.; Van Kerrebroeck, P.; Victor, A.; Wein, A.; Standardisation Sub-Committee of the International Continence Society. The standardisation of terminology in lower urinary tract function: Report from the standardisation sub-committee of the International Continence Society. Urology 2003, 61, 37–49. [Google Scholar] [CrossRef] [PubMed]
- D’Ancona, C.; Haylen, B.; Oelke, M.; Abranches-Monteiro, L.; Arnold, E.; Goldman, H.; Hamid, R.; Homma, Y.; Marcelissen, T.; Rademakers, K.; et al. Standardisation Steering Committee ICS and the ICS Working Group on Terminology for Male Lower Urinary Tract & Pelvic Floor Symptoms and Dysfunction. The International Continence Society (ICS) report on the terminology for adult male lower urinary tract and pelvic floor symptoms and dysfunction. Neurourol. Urodyn. 2019, 38, 433–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Daleeneshgari, F. Diabetic bladder dysfunction. Chin. Med. J. 2014, 127, 1357–1364. [Google Scholar] [PubMed]
- Chapple, C.R.; Osman, N.I.; Birder, L.; van Koeveringe, G.A.; Oelke, M.; Nitti, V.W.; Drake, M.J.; Yamaguchi, O.; Abrams, P.; Smith, P.P. The underactive bladder: A new clinical concept? Eur. Urol. 2015, 68, 351–353. [Google Scholar] [CrossRef]
- Cohn, J.A.; Brown, E.T.; Kaufman, M.R.; Dmochowski, R.R.; Reynolds, W.S. Underactive bladder in women: Is there any evidence? Curr. Opin. Urol. 2016, 26, 309–314. [Google Scholar] [CrossRef]
- Wang, J.; Ren, L.; Liu, X.; Liu, J.; Ling, Q. Underactive bladder and detrusor underactivity: New advances and prospectives. Int. J. Mol. Sci. 2023, 24, 15517. [Google Scholar] [CrossRef]
- Yoshimura, N.; Chancellor, M.B.; Andersson, K.E.; Christ, G.J. Recent advances in understanding the biology of diabetes-associated bladder complications and novel therapy. BJU Int. 2005, 95, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Daneshgari, F.; Liu, G.; Hanna-Mitchell, A.T. Path of translational discovery of urological complications of obesity and diabetes. Am. J. Physiol. Renal Physiol. 2017, 312, F887–F896. [Google Scholar] [CrossRef] [PubMed]
- Galì, A.; Mucciardi, G.; Butticè, S.; Subba, E.; D’Amico, C.; Lembo, F.; Magno, C. Correlation between advanced glycation end-products, lower urinary tract symptoms and bladder dysfunctions in patients with type 2 diabetes mellitus. Low. Urin. Tract. Symptoms 2017, 9, 15–20. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.G.; Medeiros, M.L.; Tavares, E.B.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal, a reactive glucose metabolite, induces bladder overactivity in addition to inflammation in mice. Front. Physiol. 2020, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; Medeiros, M.L.; de Oliveira, M.G.; Teixeira, C.J.; Mónica, F.Z.; Antunes, E. Enhanced RAGE expression and excess reactive-oxygen species production mediates Rho Kinase-dependent detrusor overactivity after methylglyoxal exposure. Front. Physiol. 2022, 13, 860342. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; Medeiros, M.L.; Ghezzi, A.C.; Dos Santos, G.A.; Mello, G.C.; Mónica, F.Z.; Antunes, E. Evidence that methylglyoxal and receptor for advanced glycation end products are implicated in bladder dysfunction of obese diabetic ob/ob mice. Am. J. Physiol. Renal Physiol. 2023, 325, F436–F447. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero-Lopez, D.; Lecomte, M.; Moinet, G.; Patereau, G.; Lagarde, M.; Wiernsperger, N. Reaction of metformin with dicarbonyl compounds. Possible implication in the inhibition of advanced glycation end product formation. Biochem. Pharmacol. 1999, 58, 1765–1773. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, P.J.; Howell, S.K.; Touchette, A.D.; Lal, S.; Szwergold, B.S. Metformin reduces systemic methylglyoxal levels in type 2 diabetes. Diabetes 1999, 48, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kinsky, O.R.; Hargraves, T.L.; Anumol, T.; Jacobsen, N.E.; Dai, J.; Snyder, S.A.; Monks, T.J.; Lau, S.S. Metformin scavenges methylglyoxal to form a novel imidazolinone metabolite in humans. Chem. Res. Toxicol. 2016, 29, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Huo, X.; Wang, S.; Yin, Z. The inhibitory effects of natural antioxidants on protein glycation as well as aggregation induced by methylglyoxal and underlying mechanisms. Colloids Surf. B Biointerfaces 2022, 212, 112360. [Google Scholar] [CrossRef]
- Roy, H.A.; Green, A.L. The central autonomic network and regulation of bladder function. Front. Neurosci. 2019, 13, 535. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.; Rovner, E.S. Lower urinary tract symptoms in women: Epidemiology, diagnosis, and management. Curr. Opin. Urol. 2016, 26, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.E.; Milsom, I.; Hunskaar, S.; Reilly, K.; Kopp, Z.; Herschorn, S.; Coyne, K.; Kelleher, C.; Hampel, C.; Artibani, W.; et al. Population-based survey of urinary incontinence, overactive bladder, and other lower urinary tract symptoms in five countries: Results of the EPIC study. Eur. Urol. 2006, 50, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Milsom, I.; Abrams, P.; Cardozo, L.; Roberts, R.G.; Thüroff, J.; Wein, A.J. How widespread are the symptoms of an overactive bladder and how are they managed? A population-based prevalence study. BJU Int. 2001, 87, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.F.; Van Rooyen, J.B.; Cundiff, G.W.; Abrams, P.; Herzog, A.R.; Corey, R.; Hunt, T.L.; Wein, A.J. Prevalence and burden of overactive bladder in the United States. World J. Urol. 2003, 20, 327–336. [Google Scholar] [CrossRef]
- Teloken, C.; Caraver, F.; Weber, F.A.; Teloken, P.E.; Moraes, J.F.; Sogari, P.R.; Graziottin, T.M. Overactive bladder: Prevalence and implications in Brazil. Eur. Urol. 2006, 49, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.D., Jr.; Neves, R.C.; Neto, A.F.; Duarte, F.G.; Moreira, T.L.; Lobo, C.F.; Glasser, D.B. A population-based survey of lower urinary tract symptoms (LUTS) and symptom-specific bother: Results from the Brazilian LUTS epidemiology study (BLUES). World J. Urol. 2013, 31, 1451–1458. [Google Scholar] [CrossRef]
- Coyne, K.S.; Kvasz, M.; Ireland, A.M.; Milsom, I.; Kopp, Z.S.; Chapple, C.R. Urinary incontinence and its relationship to mental health and health-related quality of life in men and women in Sweden, the United Kingdom, and the United States. Eur. Urol. 2012, 61, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.M.; Averbeck, M.A.; Koyama, M.; Soler, R. Impact of OAB symptoms on work, quality of life and treatment-seeking behavior in Brazil. Curr. Med. Res. Opin. 2020, 36, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Bai, H.; Mather, B.; Hill, M.A.; Jia, G.; Sowers, J.R. Diabetic vasculopathy: Molecular mechanisms and clinical insights. Int. J. Mol. Sci. 2024, 25, 804. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, S.; Smit, E.; Giovannucci, E.; Platz, E.A. Association between markers of the metabolic syndrome and lower urinary tract symptoms in the Third National Health and Nutrition Examination Survey (NHANES III). Int. J. Obes. 2005, 29, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Hunskaar, S. A systematic review of overweight and obesity as risk factors and targets for clinical intervention for urinary incontinence in women. Neurourol. Urodyn. 2008, 27, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.C.; Chung, S.D.; Ho, C.H.; Tai, T.Y.; Yang, W.S.; Tseng, C.H.; Wu, H.P.; Yu, H.J. Metabolic syndrome components worsen lower urinary tract symptoms in women with type 2 diabetes. J. Clin. Endocrinol. Metab. 2010, 95, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Uzun, H.; Zorba, O.U. Metabolic syndrome in female patients with overactive bladder. Urology 2012, 79, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Bang, W.J.; Lee, J.Y.; Koo, K.C.; Hah, Y.S.; Lee, D.H.; Cho, K.S. Is type-2 diabetes mellitus associated with overactive bladder symptoms in men with lower urinary tract symptoms? Urology 2014, 84, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Bunn, F.; Kirby, M.; Pinkney, E.; Cardozo, L.; Chapple, C.; Chester, K.; Cruz, F.; Haab, F.; Kelleher, C.; Milsom, I.; et al. Is there a link between overactive bladder and the metabolic syndrome in women? A systematic review of observational studies. Int. J. Clin. Pract. 2015, 69, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Zacche, M.M.; Giarenis, I.; Thiagamoorthy, G.; Robinson, D.; Cardozo, L. Is there an association between aspects of the metabolic syndrome and overactive bladder? A prospective cohort study in women with lower urinary tract symptoms. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 217, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.H.; Helmuth, M.E.; Smith, A.R.; Wiseman, J.B.; Gillespie, B.W.; Kirkali, Z. Symptoms of lower urinary tract dysfunction research network (LURN). relationship between central Obesity, general obesity, overactive bladder syndrome and urinary incontinence among male and female patients seeking care for their lower urinary tract symptoms. Urology 2019, 123, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Lee, Y.G.; Han, K.; Han, J.H. Obesity, metabolic health, and urological disorders in adults: A nationwide population-based study. Sci. Rep. 2021, 11, 8687. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.A.; D’ancona, C.A.L.; Cândido, E.C.; Achermann, A.P.P.; Chaim, E.A. Prevalence of LUTS and urodynamics results in obese women. Neurourol. Urodyn. 2022, 41, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.E.; Kim, D.G.; Yoo, J.W.; Lee, K.S. Metabolic syndrome-related factors as possible targets for lower urinary tract symptoms in Korean males. Aging Male 2023, 26, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Chilaka, C.; Toozs-Hobson, P.; Chilaka, V. Pelvic floor dysfunction and obesity. Best Pract. Res. Clin. Obstet. Gynaecol. 2023, 90, 102389. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zou, J.; Wang, Z.; Wang, M.; Yuan, Y.; Lv, H. Correlation Between Insulin resistance and urinary incontinence in female patients with type 2 Diabetes Mellitus. Int. Urogynecol. J. 2024, 35, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Alsannan, B.; Laganà, A.S.; Alhermi, J.; Almansoor, S.; Ayed, A.; Venezia, R.; Etrusco, A. Prevalence of overactive bladder among overweight and obese women: A prospective cross-sectional cohort study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2024, 295, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Park, H.K.; Lee, H.W.; Lee, K.S.; Byun, S.S.; Jeong, S.J.; Hong, S.K.; Lee, S.E.; Park, J.H.; Lee, S.B.; Kim, K.W.; et al. Relationship between lower urinary tract symptoms and metabolic syndrome in a community-based elderly population. Urology 2008, 72, 556. [Google Scholar] [CrossRef] [PubMed]
- Eom, C.S.; Park, J.H.; Cho, B.L.; Choi, H.C.; Oh, M.J.; Kwon, H.T. Metabolic syndrome and accompanying hyperinsulinemia have favorable effects on lower urinary tract symptoms in a generally healthy screened population. J. Urol. 2011, 186, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Di, X.P.; He, S.X.; Zeng, X.; Shen, H.; Zhu, H.L.; Luo, D.Y. Metabolic syndrome in women with and without interstitial cystitis/bladder pain syndrome. Int. Urogynecol. J. 2021, 32, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, G.; Liccardi, A.; Graziadio, C.; Barrea, L.; Muscogiuri, G.; Colao, A. Obesity and infectious diseases: Pathophysiology and epidemiology of a double pandemic condition. Int. J. Obes. 2022, 46, 449–465. [Google Scholar] [CrossRef]
- Pari, B.; Gallucci, M.; Ghigo, A.; Brizzi, M.F. Insight on Infections in diabetic setting. Biomedicines 2023, 11, 971. [Google Scholar] [CrossRef] [PubMed]
- Kirby, M.G.; Wagg, A.; Cardozo, L.; Chapple, C.; Castro-Diaz, D.; de Ridder, D.; Espuna-Pons, M.; Haab, F.; Kelleher, C.; Kölbl, H.; et al. Overactive bladder: Is there a link to the metabolic syndrome in men? Neurourol. Urodyn. 2010, 29, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Calogero, A.E.; Burgio, G.; Condorelli, R.A.; Cannarella, R.; La Vignera, S. Epidemiology and risk factors of lower urinary tract symptoms/benign prostatic hyperplasia and erectile dysfunction. Aging Male 2019, 22, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Omran, A.; Leca, B.M.; Oštarijaš, E.; Graham, N.; Da Silva, A.S.; Zaïr, Z.M.; Miras, A.D.; le Roux, C.W.; Vincent, R.P.; Cardozo, L.; et al. Metabolic syndrome is associated with prostate enlargement: A systematic review, meta-analysis, and meta-regression on patients with lower urinary tract symptom factors. Ther. Adv. Endocrinol. Metab. 2021, 12, 20420188211066210. [Google Scholar] [CrossRef] [PubMed]
- Subak, L.L.; Richter, H.E.; Hunskaar, S. Obesity and urinary incontinence: Epidemiology and clinical research update. J. Urol. 2009, 182, S2–S7. [Google Scholar] [CrossRef]
- Bauer, S.R.; Harrison, S.L.; Cawthon, P.M.; Senders, A.; Kenfield, S.A.; Suskind, A.M.; McCulloch, C.E.; Covinsky, K.; Marshall, L.M. Longitudinal changes in adiposity and lower urinary tract symptoms among older men. J. Gerontol. Ser. A 2022, 77, 2102–2109. [Google Scholar] [CrossRef] [PubMed]
- Daneshgari, F.; Liu, G.; Birder, L.; Hanna-Mitchell, A.T.; Chacko, S. Diabetic bladder dysfunction: Current translational knowledge. J. Urol. 2009, 182, S18–S26. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Chancellor, M.B.; Lin, J.M.; Hsieh, J.H.; Yu, H.J. Type 2 diabetes but not metabolic syndrome is associated with an increased risk of lower urinary tract symptoms and erectile dysfunction in men aged <45 years. BJU Int. 2010, 105, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Moul, S.; McVary, K.T. Lower urinary tract symptoms, obesity and the metabolic syndrome. Curr. Opin. Urol. 2010, 20, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Arrellano-Valdez, F.; Urrutia-Osorio, M.; Arroyo, C.; Soto-Vega, E. A comprehensive review of urologic complications in patients with diabetes. Springerplus 2014, 3, 549. [Google Scholar] [CrossRef] [PubMed]
- Blair, Y.; Wessells, H.; Pop-Busui, R.; Ang, L.; Sarma, A.V. Urologic complications in diabetes. J. Diabetes Complicat. 2022, 36, 108288. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Tang, Z.; He, C.; Tang, W. Diabetic cystopathy: A review. J. Diabetes 2015, 7, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Gasbarro, G.; Lin, D.L.; Vurbic, D.; Quisno, A.; Kinley, B.; Daneshgari, F.; Damaser, M.S. Voiding function in obese and type 2 diabetic female rats. Am. J. Physiol. Renal Physiol. 2010, 298, F72–F77. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.H.; Daneshgari, F.; Thor, K.; Drake, M.; Eccles, R.; Kanai, A.J.; Birder, L.A. Animal models and their use in understanding lower urinary tract dysfunction. Neurourol. Urodyn. 2010, 29, 603–608. [Google Scholar] [CrossRef]
- Liu, G.; Li, M.; Vasanji, A.; Daneshgari, F. Temporal diabetes and diuresis-induced alteration of nerves and vasculature of the urinary bladder in the rat. BJU Int. 2011, 107, 1988–1993. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, J.; Tyagi, P.; Chuang, Y.C.; Lee, W.C.; Yoshimura, N.; Huang, C.C.; Rajaganapathy, B.; Chancellor, M.B. Functional and molecular characterization of hyposensitive underactive bladder tissue and urine in streptozotocin-induced diabetic rat. PLoS ONE 2014, 9, e102644. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Babcook, M.A.; Shukla, S.; Shankar, E.; Wang, Z.; Liu, G.; Erokwu, B.O.; Flask, C.A.; Lu, L.; Daneshgari, F.; et al. Obesity-initiated metabolic syndrome promotes urinary voiding dysfunction in a mouse model. Prostate 2016, 76, 964–976. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.G.; Nascimento, D.M.; Alexandre, E.C.; Bonilla-Becerra, S.M.; Zapparoli, A.; Mónica, F.Z.; Antunes, E. Menthol ameliorates voiding dysfunction in types I and II diabetic mouse model. Neurourol. Urodyn. 2018, 37, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Blaha, I.; López-Oliva, M.E.; Martínez, M.P.; Recio, P.; Agis-Torres, Á.; Martínez, A.C.; Benedito, S.; García-Sacristán, A.; Prieto, D.; Fernandes, V.S.; et al. Bladder dysfunction in an obese Zucker rat: The role of TRPA1 channels, oxidative Stress, and hydrogen sulfide. Oxid. Med. Cell Longev. 2019, 2019, 5641645. [Google Scholar] [CrossRef] [PubMed]
- Eleazu, C.O.; Eleazu, K.C.; Chukwuma, S.; Essien, U.N. Review of the mechanism of cell death resulting from streptozotocin challenge in experimental animals, its practical use and potential risk to humans. J. Diabetes Metab. Disord. 2013, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Chmelir, T.; Chottova, D.M. Animal models in diabetic research-history, presence, and future perspectives. Biomedicines 2023, 11, 2852. [Google Scholar] [CrossRef] [PubMed]
- Andersson, P.O.; Malmgren, A.; Uvelius, B. Cystometrical and in vitro evaluation of urinary bladder function in rats with STZ-induced diabetes. J. Urol. 1988, 139, 1359–1362. [Google Scholar] [CrossRef]
- Eika, B.; Levin, R.M.; Longhurst, P.A. Comparison of urinary bladder function in rats with hereditary diabetes insipidus, streptozotocin-induced diabetes mellitus, and nondiabetic osmotic diuresis. J. Urol. 1994, 151, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Tammela, T.L.; Leggett, R.E.; Levin, R.M.; Longhurst, P.A. Temporal changes in micturition and bladder contractility after sucrose diuresis and streptozotocin-induced diabetes mellitus in rats. J. Urol. 1995, 153, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.H.; Brading, A.F. Smooth muscle of the bladder in the normal and the diseased state: Pathophysiology, diagnosis and treatment. Pharmacol. Ther. 1997, 75, 77–110. [Google Scholar] [CrossRef] [PubMed]
- Pitre, D.A.; Ma, T.; Wallace, L.J.; Bauer, J.A. Time-dependent urinary bladder remodeling in the streptozotocin-induced diabetic rat model. Acta Diabetol. 2002, 39, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Arioglu, I.E.; Ellenbroek, J.H.; Michel, M.C. A systematic review of urinary bladder hypertrophy in experimental diabetes: Part I. Streptozotocin-induced rat models. Neurourol. Urodyn. 2018, 37, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Daneshgari, F. Alterations in neurogenically mediated contractile responses of urinary bladder in rats with diabetes. Am. J. Physiol. Renal Physiol. 2005, 288, F1220–F1226. [Google Scholar] [CrossRef] [PubMed]
- Daneshgari, F.; Huang, X.; Liu, G.; Bena, J.; Saffore, L.; Powell, C.T. Temporal differences in bladder dysfunction caused by diabetes, diuresis, and treated diabetes in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1728–R1735. [Google Scholar] [CrossRef] [PubMed]
- Daneshgari, F.; Liu, G.; Imrey, P.B. Time dependent changes in diabetic cystopathy in rats include compensated and decompensated bladder function. J. Urol. 2006, 176, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Huang, Y.; Kavran, M.; Elrashidy, R.A.; Liu, G. Short-term diabetes- and diuresis-induced alterations of the bladder are mostly reversible in rats. Int. J. Urol. 2015, 22, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Yesilyurt, Z.E.; Matthes, J.; Hintermann, E.; Castañeda, T.R.; Elvert, R.; Beltran-Ornelas, J.H.; Silva-Velasco, D.L.; Xia, N.; Kannt, A.; Christen, U.; et al. Analysis of 16 studies in nine rodent models does not support the hypothesis that diabetic polyuria is a main reason of urinary bladder enlargement. Front. Physiol. 2022, 13, 923555. [Google Scholar] [CrossRef] [PubMed]
- Cao, N.; Alexandre, E.C.; Gotoh, D.; Kurobe, M.; Mizoguchi, S.; Gu, B.; Yoshimura, N. Urethral dysfunction and alterations of nitric oxide mechanisms in streptozotocin-induced diabetic rats with or without low-dose insulin treatment. Life Sci. 2020, 249, 117537. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, D.; Cao, N.; Alexandre, E.C.; Saito, T.; Morizawa, Y.; Hori, S.; Miyake, M.; Torimoto, K.; Fujimoto, K.; Yoshimura, N. Effects of low-dose insulin or a soluble guanylate cyclase activator on lower urinary tract dysfunction in streptozotocin-induced diabetic rats. Life Sci. 2021, 286, 120001. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, B.R.; Liu, G.; Arioglu-Inan, E.; Michel, M.C. Established and emerging treatments for diabetes-associated lower urinary tract dysfunction. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Mónica, F.Z.; Carvalho, F.D.; Claudino, M.A.; Franco-Penteado, C.F.; Schenka, A.; Grant, A.D.; De Nucci, G.; Antunes, E. Functional, morphological and molecular characterization of bladder dysfunction in streptozotocin-induced diabetic mice: Evidence of a role for L-type voltage-operated Ca2+ channels. Br. J. Pharmacol. 2011, 163, 1276–1288. [Google Scholar] [CrossRef]
- Yu, W.; Ackert-Bicknell, C.; Larigakis, J.D.; MacIver, B.; Steers, W.D.; Churchill, G.A.; Hill, W.G.; Zeidel, M.L. Spontaneous voiding by mice reveals strain-specific lower urinary tract function to be a quantitative genetic trait. Am. J. Physiol. Renal Physiol. 2014, 306, F1296–F1307. [Google Scholar] [CrossRef] [PubMed]
- Bjorling, D.E.; Wang, Z.; Vezina, C.M.; Ricke, W.A.; Keil, K.P.; Yu, W.; Guo, L.; Zeidel, M.L.; Hill, W.G. Evaluation of voiding assays in mice: Impact of genetic strains and sex. Am. J. Physiol. Renal Physiol. 2015, 308, F1369–F1378. [Google Scholar] [CrossRef] [PubMed]
- Mossa, A.H.; Galan, A.; Cammisotto, P.G.; Velasquez Flores, M.; Shamout, S.; Barcelona, P.; Saragovi, H.U.; Campeau, L. Antagonism of proNGF or its receptor p75NTR reverses remodelling and improves bladder function in a mouse model of diabetic voiding dysfunction. Diabetologia 2020, 63, 1932–1946. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Hayashi, Y.; Yoshida, A.; Yoshida, S.; Ito, Y.; Yamaguchi, K.; Yamada, S.; Takahashi, S. Concomitant alteration in number and affinity of P2X and muscarinic receptors are associated with bladder dysfunction in early stage of diabetic rats. Int. Urol. Nephrol. 2018, 50, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M., Jr.; Allkanjari, A.; Odom, M.R.; Jin, H.; Purves, J.T. Diabetic bladder dysfunction progresses from an overactive to an underactive phenotype in a type-1 diabetic mouse model (Akita female mouse) and is dependent on NLRP3. Life Sci. 2022, 299, 120528. [Google Scholar] [CrossRef] [PubMed]
- Hughes, F.M., Jr.; Allkanjari, A.; Odom, M.R.; Mulcrone, J.E.; Jin, H.; Purves, J.T. Male Akita mice develop signs of bladder underactivity independent of NLRP3 as a result of a decrease in neurotransmitter release from efferent neurons. Am. J. Physiol. Renal Physiol. 2023, 325, F61–F72. [Google Scholar] [CrossRef]
- Mónica, F.Z.; Antunes, E. Stimulators and activators of soluble guanylate cyclase for urogenital disorders. Nat. Rev. Urol. 2018, 15, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Torimoto, K.; Fraser, M.O.; Hirao, Y.; De Groat, W.C.; Chancellor, M.B.; Yoshimura, N. Urethral dysfunction in diabetic rats. J. Urol. 2004, 171, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Melman, A.; Zotova, E.; Kim, M.; Arezzo, J.; Davies, K.; DiSanto, M.; Tar, M. Longitudinal studies of time-dependent changes in both bladder and erectile function after streptozotocin-induced diabetes in Fischer 344 male rats. BJU Int. 2009, 104, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Radenković, M.; Stojanović, M.; Prostran, M. Experimental diabetes induced by alloxan and streptozotocin: The current state of the art. J. Pharmacol. Toxicol. Methods 2016, 78, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Uvelius, B. Detrusor smooth muscle in rats with alloxan-induced diabetes. J. Urol. 1986, 136, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Paro, M.; Italiano, G.; Travagli, R.A.; Petrelli, L.; Zanoni, R.; Prosdocimi, M.; Fiori, M.G. Cystometric changes in alloxan diabetic rats: Evidence for functional and structural correlates of diabetic autonomic neuropathy. J. Auton. Nerv. Syst. 1990, 30, 1–11. [Google Scholar] [CrossRef]
- Rodrigues, A.A., Jr.; Suaid, H.J.; Tucci, S., Jr.; Fazan, V.P.; Foss, M.C.; Cologna, A.J.; Martins, A.C. Long term evaluation of functional and morphological bladder alterations on alloxan-induced diabetes and aging: Experimental study in rats. Acta Cir. Bras. 2008, 23 (Suppl. S1), 53–58; discussion 58. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.N. Functional and biochemical characteristics of urinary bladder muscarinic receptors in long-term alloxan diabetic rats. Einstein 2015, 13, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Hypolite, J.A.; DiSanto, M.E.; Changolkar, A.; Wein, A.J.; Chacko, S. Increased basal phosphorylation of detrusor smooth muscle myosin in alloxan-induced diabetic rabbit is mediated by upregulation of Rho-kinase beta and CPI-17. Am. J. Physiol. Renal Physiol. 2006, 290, F650–F656. [Google Scholar] [CrossRef]
- Ichiyanagi, N.; Tsujii, T.; Masuda, H.; Kihara, K.; Goto, M.; Azuma, H. Changed responsiveness of the detrusor in rabbits with alloxan induced hyperglycemia: Possible role of 5-hydroxytryptamine for diabetic bladder dysfunction. J. Urol. 2002, 168, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Aravani, D.; Kassi, E.; Chatzigeorgiou, A.; Vakrou, S. Cardiometabolic syndrome: An update on available mouse models. Thromb. Haemost. 2021, 121, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahrén, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53 (Suppl. S3), S215–S219. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.U.; Phonsombat, S.; Bochinski, D.; Carrion, R.E.; Nunes, L.; Lue, T.F. An animal model to study lower urinary tract symptoms and erectile dysfunction: The hyperlipidaemic rat. BJU Int. 2007, 100, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, E.C.; Calmasini, F.B.; de Oliveira, M.G.; Silva, F.H.; da Silva, C.P.V.; André, D.M.; Leonardo, F.C.; Delbin, M.A.; Antunes, E. Chronic treatment with resveratrol improves overactive bladder in obese mice via antioxidant activity. Eur. J. Pharmacol. 2016, 788, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Calmasini, F.B.; de Oliveira, M.G.; Alexandre, E.C.; da Silva, F.H.; da Silva, C.P.V.; Candido, T.Z.; Antunes, E.; Mónica, F.Z. Long-term treatment with the beta-3 adrenoceptor agonist, mirabegron ameliorates detrusor overactivity and restores cyclic adenosine monophosphate (cAMP) levels in obese mice. Neurourol. Urodyn. 2017, 36, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Sollon, C.; Calixto, M.C.; Lintomen, L.; Mónica, F.Z.; Anhê, G.F.; De Nucci, G.; Zanesco, A.; Grant, A.D.; Antunes, E. Role of PKC and CaV1.2 in detrusor overactivity in a model of obesity associated with insulin resistance in mice. PLoS ONE 2012, 7, e48507. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Sollon, C.; Báu, F.R.; Mónica, F.Z.; D’Ancona, C.L.; De Nucci, G.; Grant, A.D.; Anhê, G.F.; Antunes, E. Insulin relaxes bladder via PI3K/AKT/eNOS pathway activation in mucosa: Unfolded protein response-dependent insulin resistance as a cause of obesity-associated overactive bladder. J. Physiol. 2013, 591, 2259–2273. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Silva, F.H.; Davel, A.P.; Alexandre, E.C.; Calixto, M.C.; De Nucci, G.; Mónica, F.Z.; Antunes, E. The soluble guanylyl cyclase activator BAY 60-2770 ameliorates overactive bladder in obese mice. J. Urol. 2014, 191, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Malysz, J.; Petkov, G.V. Urinary bladder smooth muscle ion channels: Expression, function, and regulation in health and disease. Am. J. Physiol. Renal Physiol. 2020, 319, F257–F283. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Luttrell, I.; Chitaley, K.; Yang, C.C. T- and L-type voltage-gated calcium channels: Their role in diabetic bladder dysfunction. Neurourol. Urodyn. 2014, 33, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, E.C.; Leiria, L.O.; Silva, F.H.; Mendes-Silvério, C.B.; Calmasini, F.B.; Davel, A.P.; Mónica, F.Z.; De Nucci, G.; Antunes, E. Soluble guanylyl cyclase (sGC) degradation and impairment of nitric oxide-mediated responses in urethra from obese mice: Reversal by the sGC activator BAY 60-2770. J. Pharmacol. Exp. Ther. 2014, 349, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, E.C.; Calmasini, F.B.; Sponton, A.C.D.S.; de Oliveira, M.G.; André, D.M.; Silva, F.H.; Delbin, M.A.; Mónica, F.Z.; Antunes, E. Influence of the periprostatic adipose tissue in obesity-associated mouse urethral dysfunction and oxidative stress: Effect of resveratrol treatment. Eur. J. Pharmacol. 2018, 836, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Calmasini, F.B.; McCarthy, C.G.; Wenceslau, C.F.; Priviero, F.B.M.; Antunes, E.; Webb, R.C. Toll-like receptor 9 regulates metabolic profile and contributes to obesity-induced benign prostatic hyperplasia in mice. Pharmacol. Rep. 2020, 72, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Lin, G.; Wang, G.; Reed, A.; Lu, Z.; Wang, L.; Banie, L.; Lue, T.F. Impaired contractility of the circular striated urethral sphincter muscle may contribute to stress urinary incontinence in female zucker fatty rats. Neurourol. Urodyn. 2017, 36, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.K.; Hamadani, C.; Zeidel, M.L.; Hill, W.G. Urological complications of obesity and diabetes in males and females of three mouse models: Temporal manifestations. Am. J. Physiol. Renal Physiol. 2020, 318, F160–F174. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, M.; Maher, S.; Fu, P.; Cai, D.; Wang, B.; Gupta, S.; Hijaz, A.; Daneshgari, F.; Liu, G. Effects of different diets used to induce obesity/metabolic syndrome on bladder function in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2023, 324, R70–R81. [Google Scholar] [CrossRef] [PubMed]
- Oswal, A.; Yeo, G. Leptin and the control of body weight: A review of its diverse central targets, signaling mechanisms, and role in the pathogenesis of obesity. Obesity 2010, 18, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, J.H.; Arioglu, E.; Michel, M.C. A systematic review of urinary bladder hypertrophy in experimental diabetes: Part 2. Comparison of animal models and functional consequences. Neurourol. Urodyn. 2018, 37, 2346–2360. [Google Scholar] [CrossRef] [PubMed]
- Nobe, K.; Yamazak, T.; Tsumita, N.; Hashimoto, T.; Honda, K. Glucose-dependent enhancement of diabetic bladder contraction is associated with a rho kinase-regulated protein kinase C pathway. J. Pharmacol. Exp. Ther. 2009, 328, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, X.; Xiao, N.; Huang, Y.; Kavran, M.; Elrashidy, R.A.; Wang, M.; Daneshgari, F.; Liu, G. Functional and morphological alterations of the urinary bladder in type 2 diabetic FVB (db/db) mice. J. Diabetes Complicat. 2016, 30, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cheng, Z.; Cristofaro, V.; Li, J.; Xiao, X.; Gomez, P.; Ge, R.; Gong, E.; Strle, K.; Sullivan, M.P.; et al. Inhibition of TNF-α improves the bladder dysfunction that is associated with type 2 diabetes. Diabetes 2012, 61, 2134–2145. [Google Scholar] [CrossRef] [PubMed]
- Riboulet, C.A.; Pierron, I.; Durand, J.; Murdaca, J.; Giudicelli, E.; Van, O. Methylglyoxal impairs the insulin signaling pathways independently of the formation of intracellular reactive oxygen species. Diabetes 2006, 55, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, O.; Niessen, P.M.; Ferreira, I.; Miyata, T.; Scheffer, P.G.; Teerlink, T.; Schrauwen, P.; Brownlee, M.; Stehouwer, C.D.; Schalkwijk, C.G. Overexpression of glyoxalase-I reduces hyperglycemia-induced levels of advanced glycation end products and oxidative stress in diabetic rats. J. Biol. Chem. 2011, 286, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Shamsaldeen, Y.A.; Mackenzie, L.S.; Lione, L.A.; Benham, C.D. Methylglyoxal, a metabolite increased in diabetes is associated with insulin resistance, vascular dysfunction and neuropathies. Curr. Drug Metab. 2016, 17, 359–367. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz-Ares, S.; Cardelo, M.P.; Gutiérrez-Mariscal, F.M.; Torres-Peña, J.D.; García-Rios, A.; Katsiki, N.; Malagón, M.M.; López-Miranda, J.; Pérez-Martínez, P.; Yubero-Serrano, E.M. Endothelial dysfunction and advanced glycation end products in patients with newly diagnosed versus established diabetes: From the CORDIOPREV Study. Nutrients 2020, 12, 238. [Google Scholar] [CrossRef] [PubMed]
- Heber, S.; Haller, P.M.; Kiss, A.; Jäger, B.; Huber, K.; Fischer, M.J.M. Association of plasma methylglyoxal increase after myocardial infarction and the left ventricular ejection fraction. Biomedicines 2024, 10, 605. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Langborg, A.; Minhas, H.S. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochem. J. 1999, 344 Pt 1, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Kalapos, M.P.; de Bari, L. Hidden biochemical fossils reveal an evolutionary trajectory for glycolysis in the prebiotic era. FEBS Lett. 2022, 596, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.W.T.; Lopez Gonzalez, E.J.; Zoukari, T.; Ki, P.; Shuck, S.C. Methylglyoxal and Its Adducts: Induction, Repair, and Association with Disease. Chem. Res. Toxicol. 2022, 35, 1720–1746. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D.E.; Stehouwer, C.D.; Schalkwijk, C.G. The role of methylglyoxal and the glyoxalase system in diabetes and other age-related diseases. Clin. Sci. 2015, 28, 839–861. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, N.M.J.; Stehouwer, C.D.A.; Schalkwijk, C.G. Methylglyoxal stress, the glyoxalase system, and diabetic chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2019, 28, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Miranda, E.R.; Haus, J.M. Glyoxalase I is a novel target for the prevention of metabolic derangement. Pharmacol. Ther. 2023, 250, 108524. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, O.; Niessen, P.M.; Haenen, G.; Miyata, T.; Brownlee, M.; Stehouwer, C.D.; De Mey, J.G.; Schalkwijk, C.G. Hyperglycaemia-induced impairment of endothelium-dependent vasorelaxation in rat mesenteric arteries is mediated by intracellular methylglyoxal levels in a pathway dependent on oxidative stress. Diabetologia 2010, 53, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Waris, S.; Fleming, T.; Santarius, T.; Larkin, S.J.; Winklhofer-Roob, B.M.; Stratton, M.R.; Rabbani, N. Imidazopurinones are markers of physiological genomic damage linked to DNA instability and glyoxalase 1-associated tumour multidrug resistance. Nucleic Acids Res. 2010, 38, 5432–5442. [Google Scholar] [CrossRef] [PubMed]
- Ban, I.; Sugawa, H.; Nagai, R. Protein Modification with Ribose Generates Nδ-(5-hydro-5-methyl-4-imidazolone-2-yl)-ornithine. Int. J. Mol. Sci. 2022, 23, 1224. [Google Scholar] [CrossRef] [PubMed]
- Irshad, Z.; Xue, M.; Ashour, A.; Larkin, J.R.; Thornalley, P.J.; Rabbani, N. Activation of the unfolded protein response in high glucose treated endothelial cells is mediated by methylglyoxal. Sci. Rep. 2019, 9, 7889. [Google Scholar] [CrossRef]
- Zhang, X.; Scheijen, J.L.; Stehouwer, C.D.A.; Wouters, K.; Schalkwijk, C. Increased methylglyoxal formation in plasma and tissues during a glucose tolerance test is derived from exogenous glucose. Clin. Sci. 2023, 20, CS20220753. [Google Scholar] [CrossRef] [PubMed]
- Erusalimsky, J.D. The use of the soluble receptor for advanced glycation-end products (sRAGE) as a potential biomarker of disease risk and adverse outcomes. Redox Biol. 2021, 42, 101958. [Google Scholar] [CrossRef]
- Jangde, N.; Ray, R.; Rai, V. RAGE and its ligands: From pathogenesis to therapeutics. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 555–575. [Google Scholar] [CrossRef] [PubMed]
- Dozio, E.; Vettoretti, S.; Caldiroli, L.; Nerini-Molteni, S.; Tacchini, L.; Ambrogi, F.; Messa, P.; Corsi Romanelli, M.M. Advanced glycation end products (AGE) and soluble forms of AGE receptor: Emerging role as mortality risk factors in CKD. Biomedicines 2020, 8, 638. [Google Scholar] [CrossRef]
- Ganbaatar, B.; Fukuda, D.; Shinohara, M.; Yagi, S.; Kusunose, K.; Yamada, H.; Soeki, T.; Hirata, K.I.; Sata, M. Empagliflozin ameliorates endothelial dysfunction and suppresses atherogenesis in diabetic apolipoprotein E-deficient mice. Eur. J. Pharmacol. 2020, 875, 173040. [Google Scholar] [CrossRef] [PubMed]
- Korac, B.; Kalezic, A.; Pekovic-Vaughan, V.; Korac, A.; Jankovic, A. Redox changes in obesity, metabolic syndrome, and diabetes. Redox Biol. 2021, 42, 101887. [Google Scholar] [CrossRef] [PubMed]
- Seryogina, E.S.; Kamynina, A.V.; Koroev, D.O.; Volpina, O.M.; Vinokurov, A.Y.; Abramov, A.Y. RAGE induces physiological activation of NADPH oxidase in neurons and astrocytes and neuroprotection. FEBS J. 2024. [Google Scholar] [CrossRef]
- Dobrucki, I.T.; Miskalis, A.; Nelappana, M.; Applegate, C.; Wozniak, M.; Czerwinski, A.; Kalinowski, L.; Dobrucki, L.W. Receptor for advanced glycation end-products: Biological significance and imaging applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2023, 5, e1935. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P. Oxidative Stress in Health and Disease. Biomedicines 2023, 11, 2925. [Google Scholar] [CrossRef] [PubMed]
- Odani, H.; Shinzato, T.; Matsumoto, Y.; Usami, J.; Maeda, K. Increase in three alpha, beta-dicarbonyl compound levels in human uremic plasma: Specific in vivo determination of intermediates in advanced Maillard reaction. Biochem. Biophys. Res. Commun. 1999, 256, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Lapolla, A.; Flamini, R.; Dalla, V.A.; Senesi, A.; Reitano, R.; Fedele, D.; Basso, E.; Seraglia, R.; Traldi, P. Glyoxal and methylglyoxal levels in diabetic patients: Quantitative determination by a new GC/MS method. Clin. Chem. Lab. Med. 2003, 41, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Nakayama, K.; Nakayama, M.; Mori, T.; Matsushima, M.; Okamura, M.; Senda, M.; Nako, K.; Miyata, T.; Ito, S. Methylglyoxal is a predictor in type 2 diabetic patients of intima-media thickening and elevation of blood pressure. Hypertension 2010, 56, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Randell, E.; Han, Y.; Adeli, K.; Krahn, J.; Meng, Q.H. Increased plasma methylglyoxal level, inflammation, and vascular endothelial dysfunction in diabetic nephropathy. Clin. Biochem. 2011, 44, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Terawaki, H.; Nakao, M.; Nakayama, K.; Nakayama, M.; Kimura, A.; Takane, K.; Mitome, J.; Hamaguchi, A.; Ogura, M.; Yokoyama, K.; et al. Peritoneal clearance and transport of methylglyoxal. Nephrol. Dial. Transplant. 2011, 26, 753–754. [Google Scholar] [CrossRef] [PubMed]
- Senda, M.; Ogawa, S.; Nako, K.; Okamura, M.; Sakamoto, T.; Ito, S. The strong relation between post-hemodialysis blood methylglyoxal levels and post-hemodialysis blood glucose concentration rise. Clin. Exp. Nephrol. 2015, 9, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Ramachandra, B.L.; Vedantham, S.; Krishnan, U.M.; Rayappan, J.B.B. Methylglyoxal—An emerging biomarker for diabetes mellitus diagnosis and its detection methods. Biosens. Bioelectron. 2019, 133, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Mariscal, F.M.; Cardelo, M.P.; de la Cruz, S.; Alcala-Diaz, J.F.; Roncero-Ramos, I.; Guler, I.; Vals-Delgado, C.; López-Moreno, A.; Luque, R.M.; Delgado-Lista, J.; et al. Reduction in circulating advanced glycation end products by mediterranean diet is associated with increased likelihood of yype 2 diabetes remission in patients with coronary heart disease: From the Cordioprev Study. Mol. Nutr. Food Res. 2021, 65, e1901290. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D.E.; Hanssen, N.M.; Lips, M.A.; Scheijen, J.L.; Willems, D.K.; Pijl, H.; Stehouwer, C.D.; Schalkwijk, C.G. Energy restriction and Roux-en-Y gastric bypass reduce postprandial α-dicarbonyl stress in obese women with type 2 diabetes. Diabetologia 2016, 59, 2013–2017. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Belda, M.; Fernández-García, A.J.; Campillo, N.; Pérez-Cárceles, M.D.; Motas, M.; Hernández-Córdoba, M.; Viñas, P. Glyoxal and methylglyoxal as urinary markers of diabetes. Determination using a dispersive liquid-liquid microextraction procedure combined with gas chromatography-mass spectrometry. J. Chromatogr. A 2017, 1509, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bora, S.; Shankarrao Adole, P.; Vishwanath Vinod, K.; Ananthkrishna Pillai, A.; Ahmed, S. The genetic polymorphisms and activity of glyoxalase 1 as a risk factor for acute coronary syndrome in South Indians with type 2 diabetes mellitus. Gene 2023, 10, 147701. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J.; Yamaguchi, K.; Ishikura, H.; Tsubota, M.; Sekiguchi, F.; Seki, Y.; Tsujiuchi, T.; Murai, A.; Umemura, T.; Kawabata, A. Bladder pain relief by HMGB1 neutralization and soluble thrombomodulin in mice with cyclophosphamide-induced cystitis. Neuropharmacology 2014, 79, 112–118. [Google Scholar] [CrossRef]
- Hiramoto, S.; Tsubota, M.; Yamaguchi, K.; Okazaki, K.; Sakaegi, A.; Toriyama, Y.; Tanaka, J.; Sekiguchi, F.; Ishikura, H.; Wake, H.; et al. Cystitis-related bladder pain involves ATP-dependent HMGB1 release from macrophages and its downstream H2S/Cav3.2 signaling in mice. Cells 2020, 9, 1748. [Google Scholar] [CrossRef]
- Khorramdelazad, H.; Bagheri, V.; Hassanshahi, G.; Karami, H.; Moogooei, M.; Zeinali, M.; Abedinzadeh, M. S100A12 and RAGE expression in human bladder transitional cell carcinoma: A role for the ligand/RAGE axis in tumor progression? Asian Pac. J. Cancer Prev. 2015, 16, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Karkin, K.; İzol, V.; Kaplan, M.; Değer, M.; Akdoğan, N.; Tansuğ, M.Z. Demonstration of advanced glycation end product (AGE) expression in bladder cancer tissue in type-2 diabetic and non-diabetic patients and the relationship between AGE accumulation and endoplasmic reticulum stress with bladder cancer. Int. J. Clin. Pract. 2021, 75, e14526. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Fujiwara, Y.; Nagai, R.; Yoshida, M. Immunohistochemical detection of advanced glycation end products in human bladder with specific monoclonal antibody. Int. J. Urol. 2009, 16, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Emerging glycation-based therapeutics-glyoxalase 1 inducers and glyoxalase 1 inhibitors. Int. J. Mol. Sci. 2022, 23, 2453. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Aikawa, C.; Yoshida, R.; Matsui, T. Methylglyoxal-derived hydroimidazolone residue of plasma protein can behave as a predictor of prediabetes in Spontaneously Diabetic Torii rats. Physiol. Rep. 2015, 3, e12477. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, J.; Cibrian, D.; Guillén, I.; Freyre, F.; Alba, J.S.; Lopez-Saura, P.; Merino, N.; Aldama, A.; Quintela, A.M.; Triana, M.E.; et al. Methylglyoxal administration induces diabetes-like microvascular changes and perturbs the healing process of cutaneous wounds. Clin. Sci. 2005, 109, 83–95. [Google Scholar] [CrossRef]
- Sena, C.M.; Matafome, P.; Crisóstomo, J.; Rodrigues, L.; Fernandes, R.; Pereira, P.; Seiça, R.M. Methylglyoxal promotes oxidative stress and endothelial dysfunction. Pharmacol. Res. 2012, 65, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Tikellis, C.; Pickering, R.J.; Tsorotes, D.; Huet, O.; Cooper, M.E.; Jandeleit-Dahm, K.; Thomas, M.C. Dicarbonyl stress in the absence of hyperglycemia increases endothelial inflammation and atherogenesis similar to that observed in diabetes. Diabetes 2014, 63, 3915–3925. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, N.M.J.; Tikellis, C.; Pickering, R.J.; Dragoljevic, D.; Lee, M.K.S.; Block, T.; Scheijen, J.L.; Wouters, K.; Miyata, T.; Cooper, M.E.; et al. Pyridoxamine prevents increased atherosclerosis by intermittent methylglyoxal spikes in the aortic arches of ApoE-/- mice. Biomed. Pharmacother. 2023, 158, 114211. [Google Scholar] [CrossRef] [PubMed]
- Crisóstomo, J.; Matafome, P.; Santos-Silva, D.; Rodrigues, L.; Sena, C.M.; Pereira, P.; Seiça, R. Methylglyoxal chronic administration promotes diabetes-like cardiac ischaemia disease in Wistar normal rats. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1223–1230. [Google Scholar] [CrossRef]
- Peyret, H.; Konecki, C.; Terryn, C.; Dubuisson, F.; Millart, H.; Feliu, C.; Djerada, Z. Methylglyoxal induces cardiac dysfunction through mechanisms involving altered intracellular calcium handling in the rat heart. Chem. Biol. Interact. 2024, 394, 110949. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.; Matafome, P.; Crisóstomo, J.; Santos-Silva, D.; Sena, C.; Pereira, P.; Seiça, R. Advanced glycation end products and diabetic nephropathy: A comparative study using diabetic and normal rats with methylglyoxal-induced glycation. J. Physiol. Biochem. 2014, 70, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Kang, W.S.; Jo, K.; Kim, J. Ethyl pyruvate prevents renal damage induced by methylglyoxal-derived advanced glycation end products. J. Diabetes Res. 2019, 2019, 4058280. [Google Scholar] [CrossRef]
- Mojadami, S.; Ahangarpour, A.; Mard, S.A.; Khorsandi, L. Diabetic nephropathy induced by methylglyoxal: Gallic acid regulates kidney microRNAs and glyoxalase1-Nrf2 in male mice. Arch. Physiol. Biochem. 2023, 29, 655–662. [Google Scholar] [CrossRef]
- Medeiros, M.L.; de Oliveira, M.G.; Tavares, E.G.; Mello, G.C.; Anhê, G.F.; Mónica, F.Z.; Antunes, E. Long-term methylglyoxal intake aggravates murine Th2-mediated airway eosinophil infiltration. Int. Immunopharmacol. 2020, 81, 106254. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.L.; Oliveira, A.L.; de Oliveira, M.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal exacerbates lipopolysaccharide-induced acute lung injury via RAGE-induced ROS generation: Protective effects of metformin. J. Inflamm. Res. 2021, 14, 6477–6489. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.L.; Oliveira, A.L.; Mello, G.C.; Antunes, E. Metformin counteracts the deleterious effects of methylglyoxal on ovalbumin-induced airway eosinophilic inflammation and remodeling. Int. J. Mol. Sci. 2023, 24, 9549. [Google Scholar] [CrossRef] [PubMed]
- Zunkel, K.; Simm, A.; Bartling, B. Long-term intake of the reactive metabolite methylglyoxal is not toxic in mice. Food Chem. Toxicol. 2020, 141, 111333. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; de Oliveira, M.G.; Medeiros, M.L.; Mónica, F.; Antunes, E. Metformin abrogates the voiding dysfunction induced by prolonged methylglyoxal intake. Eur. J. Pharmacol. 2021, 910, 174502. [Google Scholar] [CrossRef] [PubMed]
- Al-Robaiy, S.; Navarrete, S.A.; Simm, A. RAGE-dependent effect of exogenous methylglyoxal intake on lung biomechanics in mice. Nutrients 2022, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, Y.; Wang, P.; Sang, S. Simultaneous determination of multiple reactive carbonyl species in high fat diet-induced metabolic disordered mice and the inhibitory effects of rosemary on carbonyl stress. J. Agric. Food Chem. 2021, 69, 1123–1131. [Google Scholar] [CrossRef]
- Oliveira, A.L.; Medeiros, M.L.; Gomes, E.T.; Mello, G.C.; Costa, S.K.; Mónica, F.Z.; Antunes, E. TRPA1 channel mediates methylglyoxal-induced mouse bladder dysfunction. Front. Physiol. 2023, 14, 1308077. [Google Scholar] [CrossRef] [PubMed]
- Hill, W.G.; Zeidel, M.L.; Bjorling, D.E.; Vezina, C.M. Void spot assay: Recommendations on the use of a simple micturition assay for mice. Am. J. Physiol. Renal Physiol. 2018, 315, F1422–F1429. [Google Scholar] [CrossRef] [PubMed]
- Wegner, K.A.; Abler, L.L.; Oakes, S.R.; Mehta, G.S.; Ritter, K.E.; Hill, W.G.; Zwaans, B.M.; Lamb, L.E.; Wang, Z.; Bjorling, D.E.; et al. Void spot assay procedural optimization and software for rapid and objective quantification of rodent voiding function, including overlapping urine spots. Am. J. Physiol. Renal Physiol. 2018, 315, F1067–F1080. [Google Scholar] [CrossRef] [PubMed]
- Laddha, A.P.; Kulkarni, Y.A. NADPH oxidase: A membrane-bound enzyme and its inhibitors in diabetic complications. Eur. J. Pharmacol. 2020, 881, 173206. [Google Scholar] [CrossRef] [PubMed]
- Poladia, D.P.; Bauer, J.A. Early cell-specific changes in nitric oxide synthases, reactive nitrogen species formation, and ubiquitinylation during diabetes-related bladder remodeling. Diabetes Metab. Res. Rev. 2003, 19, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Beshay, E.; Carrier, S. Oxidative stress plays a role in diabetes-induced bladder dysfunction in a rat model. Urology 2004, 64, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Kanika, N.D.; Chang, J.; Tong, Y.; Tiplitsky, S.; Li, J.; Yohannes, E.; Tar, M.; Chance, M.; Christ, G.J.; Melman, A.; et al. Oxidative stress status accompanying diabetic bladder cystopathy results in the activation of protein degradation pathways. BJU Int. 2011, 107, 1676–1684. [Google Scholar] [CrossRef]
- Tomechko, S.E.; Liu, G.; Tao, M.; Schlatzer, D.; Powell, C.T.; Gupta, S.; Chance, M.R.; Daneshgari, F. Tissue specific dysregulated protein subnetworks in type 2 diabetic bladder urothelium and detrusor muscle. Mol. Cell Proteom. 2015, 14, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, G.G.; Davies, K.P. Novel insights into development of diabetic bladder disorder provided by metabolomic analysis of the rat nondiabetic and diabetic detrusor and urothelial layer. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E471–E479. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.E. Oxidative stress and its possible relation to lower urinary tract functional pathology. BJU Int. 2018, 121, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.E. Oxidative stress and its relation to lower urinary tract symptoms. Int. Neurourol. J. 2022, 26, 261–267. [Google Scholar] [CrossRef]
- Song, Q.X.; Sun, Y.; Deng, K.; Mei, J.Y.; Chermansky, C.J.; Damaser, M.S. Potential role of oxidative stress in the pathogenesis of diabetic bladder dysfunction. Nat. Rev. Urol. 2022, 19, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Khosla, L.; Gong, S.; Weiss, J.P.; Birder, L.A. Oxidative stress biomarkers in age-related lower urinary tract disorders: A systematic review. Int. Neurourol. J. 2022, 26, 3–19. [Google Scholar] [CrossRef]
- Miyata, Y.; Matsuo, T.; Mitsunari, K.; Asai, A.; Ohba, K.; Sakai, H. A review of oxidative stress and urinary dysfunction caused by bladder outlet obstruction and treatments using antioxidants. Antioxidants 2019, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.; Monica, F.Z.; Passos, G.R.; Victorio, J.A.; Davel, A.P.; Oliveira, A.L.L.; Parada, C.A.; D’Ancona, C.A.L.; Hill, W.G.; Antunes, E. Selective pharmacological inhibition of NOX2 by GSK2795039 improves bladder dysfunction in cyclophosphamide-induced cystitis in mice. Antioxidants 2022, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Calmasini, F.B.; de Oliveira, M.G.; Alexandre, E.C.; Silva, F.H.; Tavares, E.B.G.; André, D.M.; Zapparoli, A.; Antunes, E. Obesity-induced mouse benign prostatic hyperplasia (BPH) is improved by treatment with resveratrol: Implication of oxidative stress, insulin sensitivity and neuronal growth factor. J. Nutr. Biochem. 2018, 55, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Elrashidy, R.A.; Kavran, M.; Asker, M.E.; Mohamed, H.E.; Daneshgari, F.; Liu, G. Smooth muscle-specific deletion of MnSOD exacerbates diabetes-induced bladder dysfunction in mice. Am. J. Physiol. Renal Physiol. 2019, 317, F906–F912. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Kihara, K.; Saito, K.; Matsuoka, Y.; Yoshida, S.; Chancellor, M.B.; de Groat, W.C.; Yoshimura, N. Reactive oxygen species mediate detrusor overactivity via sensitization of afferent pathway in the bladder of anaesthetized rats. BJU Int. 2008, 101, 775–780. [Google Scholar] [CrossRef]
- Frara, N.; Giaddui, D.; Braverman, A.S.; Jawawdeh, K.; Wu, C.; Ruggieri, M.R.; Barbe, M.F. Mechanisms involved in nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (Nox)-derived reactive oxygen species (ROS) modulation of muscle function in human and dog bladders. PLoS ONE 2023, 18, e0287212. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef] [PubMed]
- Coyle, C.H.; Philips, B.J.; Morrisroe, S.N.; Chancellor, M.B.; Yoshimura, N. Antioxidant effects of green tea and its polyphenols on bladder cells. Life Sci. 2008, 83, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Nomiya, M.; Andersson, K.E.; Yamaguchi, O. Chronic bladder ischemia and oxidative stress: New pharmacotherapeutic targets for lower urinary tract symptoms. Int. J. Urol. 2015, 22, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Szkudelska, K.; Szkudelski, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei-Malazy, O.; Lavari, N.; Abdollahi, M. Natural products in the clinical management of metabolic syndrome. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2024. [Google Scholar] [CrossRef]
- Tsounapi, P.; Honda, M.; Hikita, K.; Sofikitis, N.; Takenaka, A. Oxidative stress alterations in the bladder of a short-period type 2 diabetes rat model: Antioxidant treatment can be beneficial for the bladder. In Vivo 2019, 33, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Du, H.; Hou, J.; Li, N. Anti-inflammation properties of resveratrol in the detrusor smooth muscle of the diabetic rat. Int. Urol. Nephrol. 2022, 54, 2833–2843. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zeng, H.; Yu, Y.; Zhang, J.; Liu, Q.; Yang, B. Resveratrol improved detrusor fibrosis induced by mast cells during progression of chronic prostatitis in rats. Eur. J. Pharmacol. 2017, 815, 495–500. [Google Scholar] [CrossRef]
- Yu, Y.; Jiang, J.; He, Y.; Wang, W.; Shen, C.; Yang, B. Resveratrol improves urinary dysfunction in rats with chronic prostatitis and suppresses the activity of the stem cell factor/c-Kit signaling pathway. Mol. Med. Rep. 2017, 16, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.L.; Lin, K.L.; Wu, B.N.; Chuang, S.M.; Wu, W.J.; Lee, Y.C.; Ho, W.T.; Juan, Y.S. Epigallocatechin-3-gallate alleviates bladder overactivity in a rat model with metabolic syndrome and ovarian hormone deficiency through mitochondria apoptosis pathways. Sci. Rep. 2018, 8, 5358. [Google Scholar] [CrossRef] [PubMed]
- Juan, Y.S.; Chuang, S.M.; Lee, Y.L.; Long, C.Y.; Wu, T.H.; Chang, W.C.; Levin, R.M.; Liu, K.M.; Huang, C.H. Green tea catechins decrease oxidative stress in surgical menopause-induced overactive bladder in a rat model. BJU Int. 2012, 110, E236–E244. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.T.; Kuo, K.L.; Liu, S.H.; Shi, C.S.; Chang, H.C.; Lin, W.C.; Chou, C.T.; Hsu, C.H.; Liao, S.M.; Wang, Z.H.; et al. Epigallocatechin Gallate Attenuates Partial Bladder Outlet obstruction-induced bladder injury via suppression of endoplasmic reticulum stress-related apoptosis-in vivo study. Urology 2016, 91, 242.e1–242.e9. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Shao, X.; Bai, N.; Lo, C.Y.; Yang, C.S.; Ho, C.T. Tea polyphenol (-)-epigallocatechin-3-gallate: A new trapping agent of reactive dicarbonyl species. Chem. Res. Toxicol. 2007, 20, 1862–1870. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, L.; Zhou, Q.; Yan, S.; Li, Z.; Sheng, J.; Zhang, W. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol. Nutr. Food Res. 2014, 58, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Guigas, B.; Viollet, B. Metformin: Update on mechanisms of action and repurposing potential. Nat. Rev. Endocrinol. 2023, 19, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Schernthaner, G.; Schernthaner, G.H. The right place for metformin today. Diabetes Res. Clin. Pract. 2020, 159, 107946. [Google Scholar] [CrossRef] [PubMed]
- Mbara, K.C.; Mofo, P.E.; Driver, C.; Nzuza, S.; Mkhombo, N.T.; Gcwensa, S.K.; Mcobothi, E.N.; Owira, P.M. Metformin turns 62 in pharmacotherapy: Emergence of non-glycaemic effects and potential novel therapeutic applications. Eur. J. Pharmacol. 2021, 898, 173934. [Google Scholar] [CrossRef]
- Tobar, N.; Rocha, G.Z.; Santos, A.; Guadagnini, D.; Assalin, H.B.; Camargo, J.A.; Gonçalves, A.E.S.S.; Pallis, F.R.; Oliveira, A.G.; Rocco, S.A.; et al. Metformin acts in the gut and induces gut-liver crosstalk. Proc. Natl. Acad. Sci. USA 2023, 120, e2211933120. [Google Scholar] [CrossRef] [PubMed]
- Kender, Z.; Fleming, T.; Kopf, S.; Torzsa, P.; Grolmusz, V.; Herzig, S.; Schleicher, E.; Rácz, K.; Reismann, P.; Nawroth, P.P. Effect of metformin on methylglyoxal metabolism in patients with type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2014, 122, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Lv, L.; Zhang, L.; Gao, Z.; Liu, Y.; Wang, S.; Zhou, N.; Xia, Y.; Cui, J.; Jiang, X.; et al. Metformin ameliorates bladder dysfunction in a rat model of partial bladder outlet obstruction. Am. J. Physiol. Renal Physiol. 2021, 320, F838–F858. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.H.; Alexandre, E.C.; Calmasini, F.B.; Calixto, M.C.; Antunes, E. Treatment with metformin improves erectile dysfunction in a murine model of obesity associated with insulin resistance. Urology 2015, 86, 423.e1–423.e6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, H.; Yu, X.; Wu, Y.; Sui, D. Metformin ameliorates diabetic nephropathy in a rat model of low-dose streptozotocin-induced diabetes. Exp. Ther. Med. 2017, 14, 383–390. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference Number | Dose | Route of Administration | Animal and Strain | Sex | Treatment with MGO |
|---|---|---|---|---|---|
| [164] | 50–75 mg/kg | Intraperitoneal | Wistar rat | Male | Microvascular damage Microvessel degeneration |
| [165] | 50–75 mg/Kg | Drinking water | Spontaneously diabetic (GK) rats | Male | Endothelial dysfunction |
| [166] | 50 mmol/L | Drinking water | C57Bl6 ApoE-/- | Male | Atherosclerosis |
| [170] | 50–75 mg/kg | Drinking water | Goto-Kakizaki (GK), nonobese type 2 diabetic rats | ND | Renal disease |
| [171] | 17.25 mg/kg | Intraperitoneal | Sprague Dawley (SD) rats | ND | Renal disease |
| [172] | 600 mg/kg/day | Oral | NMRI mice | Male | Diabetic nephropathy |
| [176] | 500–2000 mg/kg | Drinking water | RAGE−/Glo1 ++ mice | Male Female | Renal dysfunction |
| [178] | 500 mg/kg | Drinking water | RAGE-KO mice | Male Female | Increased airway resistance/decreased maximal inspiratory flow |
| [173,175] | 0.5% | Drinking water | C57BL/6Junib mice | Male | Aggravation of allergic airway disease and acute lung injury |
| Parameter | Male | Female | |
|---|---|---|---|
| Urodynamic evaluation | Number of nonvoiding contractions (NVCs) |  |  |
| Frequency of voiding |  | | |
| Bladder capacity | | | |
| Bladder smooth Muscle contractility in vitro (presence of urothelium) | Neurogenic contractions (electrical-Field Stimulation, EFS) | | |
| Muscarinic-mediated contractions (carbachol) | | | |
| Purinergic-mediated contractions (α,β-methylene ATP) | | | |
| Void spot analysis | Total void volume | | |
| Volume per void | | | |
| Urine spot number | | | |
| Urine spot in center | | | |
| Urine spot in corner | | | |
| Histology | Collagen content | | |
increased; decreased or unaltered parameters.| Parameter | ob/ob | ob/ob + ALT-711 |
|---|---|---|
| Blood glucose | | |
| AGEs in bladder | | |
| MG-H1 content in bladder | | |
| RAGE content in bladder | | |
| Collagen content in the bladders | | |
| Volume per void | | |
| Number of voids | | |
| Void size | | |
increased; decreased or unaltered parameters.Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, A.L.; de Oliveira, M.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction? Biomedicines 2024, 12, 939. https://doi.org/10.3390/biomedicines12050939
Oliveira AL, de Oliveira MG, Mónica FZ, Antunes E. Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction? Biomedicines. 2024; 12(5):939. https://doi.org/10.3390/biomedicines12050939
Chicago/Turabian StyleOliveira, Akila Lara, Mariana Gonçalves de Oliveira, Fabíola Zakia Mónica, and Edson Antunes. 2024. "Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction?" Biomedicines 12, no. 5: 939. https://doi.org/10.3390/biomedicines12050939