
The Effects of the Coating and Aging of Biodegradable Polylactic Acid Membranes on In Vitro Primary Human Retinal Pigment Epithelium Cells
 , , , and
, , , and 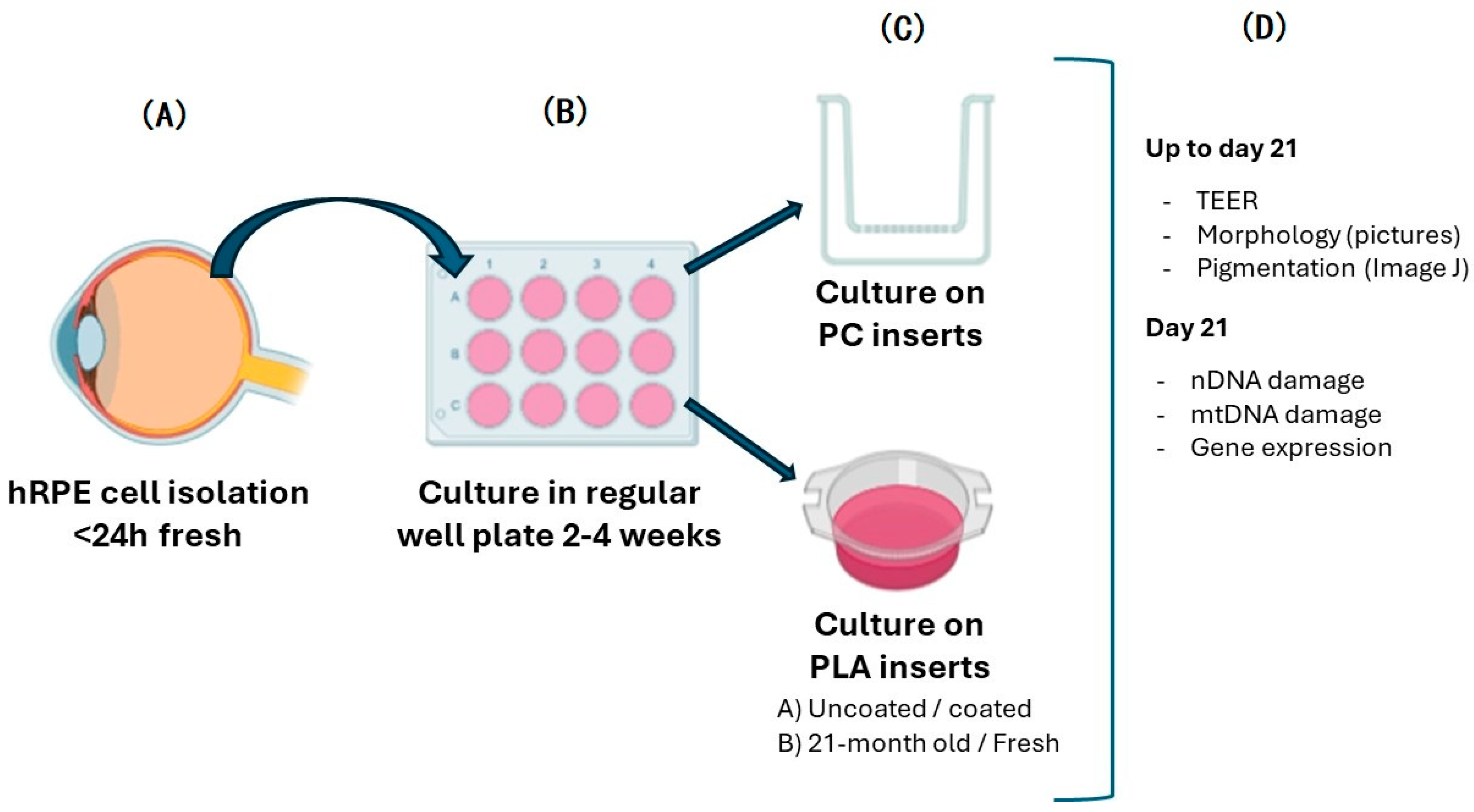{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation, Culture, Passage, and Seeding
2.2. Morphology and Pigmentation
2.3. TEER Measurements
2.4. DNA Damage and Mitochondrial DNA Copy Number Estimation
2.5. Gene Expression
3. Results and Discussion
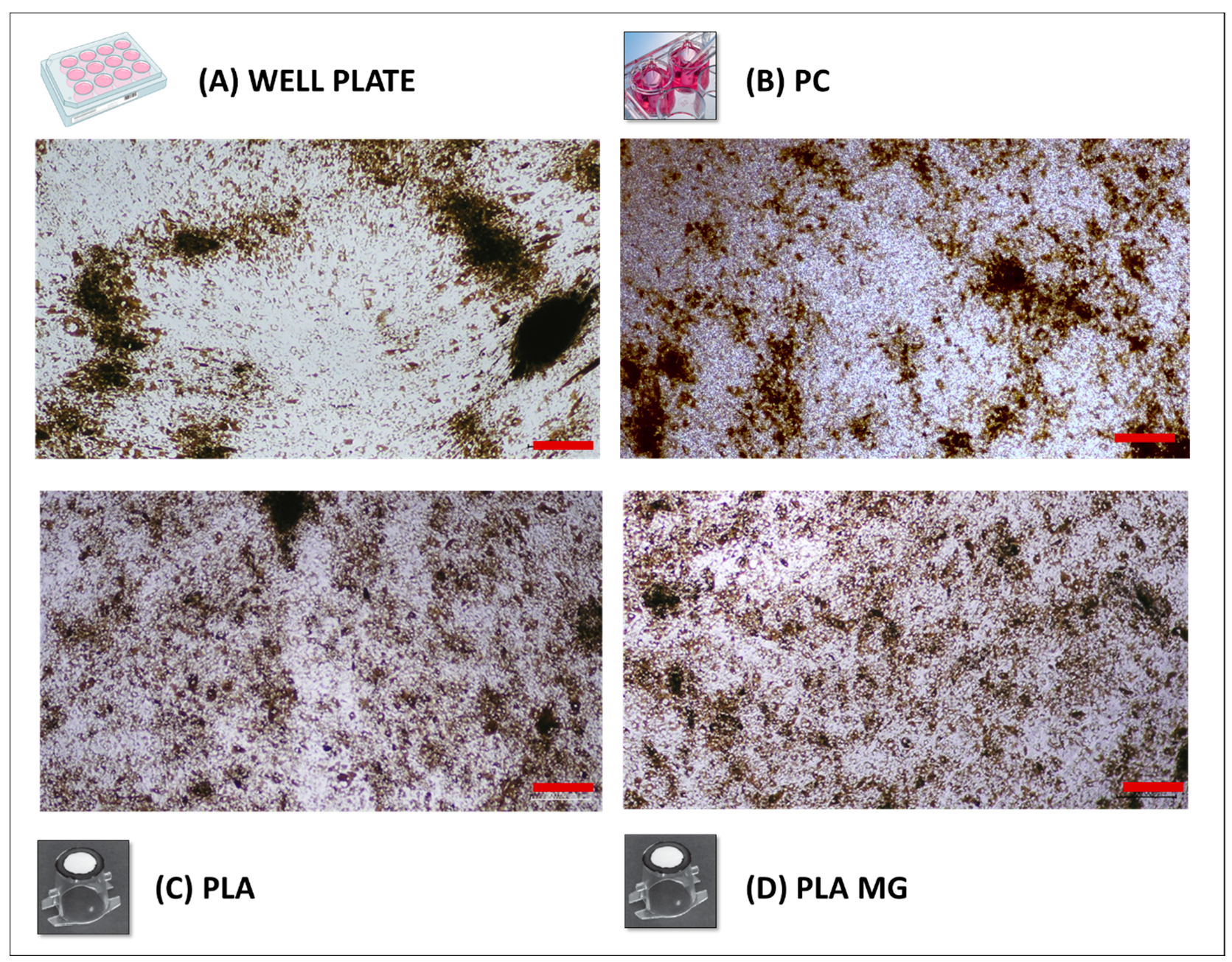
3.1. Morphology and Pigmentation
3.2. Transepithelial Resistance and Cell Detachment Analysis
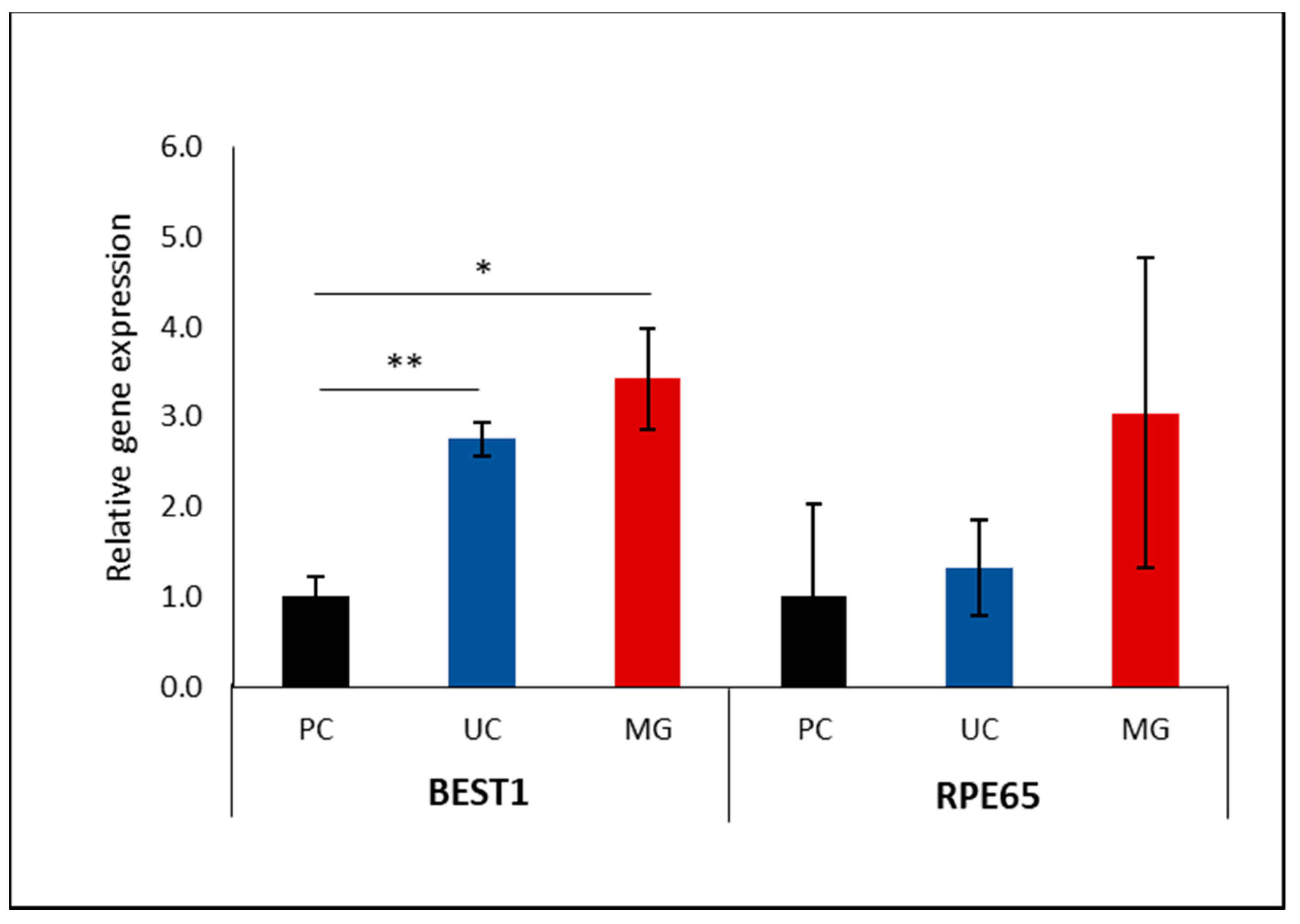
3.3. Impact of Supporting Membrane on RPE Gene Programming
3.4. DNA Damage and mtDNA Copy Number Analysis
3.5. Coating Rescues Old PLA Membrane Functionality
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ab/Am | Antibiotic/antimycotic |
| Am | Area of the membrane |
| AMD | Age-related macular degeneration |
| cDNA | Complementary desoxyribonucleic acid |
| DNA | Desoxyribonucleic acid |
| DPBS | Dulbecco phosphate-buffered saline |
| EMT | Epithelial to mesenchymal transition |
| FBS | Fetal bovine serum |
| hRPE | Human retinal pigment epithelium/al |
| MG | Matrigel |
| mtDNA | Mitochondrial desoxyribonucleic acid |
| mtDNA-CN | Mitochondrial desoxyribonucleic acid copy number |
| nDNA | Nuclear desoxyribonucleic acid |
| PC | Polycarbonate |
| PLA | Polylactide |
| Rb | Resistance in absence of cells |
| RPE | Retinal pigment epithelium/al |
| Rt | Resistance in the presence of cells |
| RT-qPCR | Reverse transcription–quantitative polymerase chain reaction |
| SEM | Standard error of the mean |
| TEER | Transepithelial electrical resistance |
References
- Nashine, S.; Nesburn, A.B.; Kuppermann, B.D.; Kenney, M.C. Age-related macular degeneration (AMD) mitochondria modulate epigenetic mechanisms in retinal pigment epithelial cells. Exp. Eye Res. 2019, 189, 107701. [Google Scholar] [CrossRef]
- Popelka, Š.; Studenovská, H.; Abelová, L.; Ardan, T.; Studený, P.; Straňák, Z.; Klíma, J.; Dvořánková, B.; Kotek, J.; Hodan, J.; et al. A frame-supported ultrathin electrospun polymer membrane for transplantation of retinal pigment epithelial cells. Biomed. Mater. 2015, 10, 045022. [Google Scholar] [CrossRef]
- Somasundaran, S.; Constable, I.J.; Mellough, C.B.; Carvalho, L.S. Retinal pigment epithelium and age-related macular degeneration: A review of major disease mechanisms. Clin. Exp. Ophthalmol. 2020, 48, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, P.G.; Kannan, R. Mechanisms of protection of retinal pigment epithelial cells from oxidant injury by humanin and other mitochondrial-derived peptides: Implications for age-related macular degeneration. Redox Biol. 2020, 37, 101663. [Google Scholar] [CrossRef]
- Sharma, R.; Bose, D.; Maminishkis, A.; Bharti, K. Retinal pigment epithelium replacement therapy for age-related macular degeneration: Are we there yet? Annu. Rev. Pharmacol. Toxicol. 2020, 60, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Michelet, F.; Balasankar, A.; Teo, N.; Stanton, L.W.; Singhal, S. Rapid generation of purified human RPE from pluripotent stem cells using 2D cultures and lipoprotein uptake-based sorting. Stem. Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Shadforth, A.M.A.; George, K.A.; Kwan, A.S.; Chirila, T.V.; Harkin, D.G. The cultivation of human retinal pigment epithelial cells on Bombyx mori silk fibroin. Biomaterials 2012, 33, 4110–4117. [Google Scholar] [CrossRef]
- Petrovski, G.; Lytvynchuk, L.; Ebbert, A.; Studenovska, H.; Nagymihály, R.; Josifovska, N.; Rais, D.; Popelka, Š.; Tichotová, L.; Nemesh, Y.; et al. Subretinal Implantation of Human Primary RPE Cells Cultured on Nanofibrous Membranes in Minipigs. Biomedicines 2022, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Khristov, V.; Rising, A.; Jha, B.S.; Dejene, R.; Hotaling, N.; Li, Y.; Stoddard, J.; Stankewicz, C.; Wan, Q.; et al. Clinical-grade stem cell–derived retinal pigment epithelium patch rescues retinal degeneration in rodents and pigs. Sci. Transl. Med. 2019, 11, eaat5580. [Google Scholar] [CrossRef]
- Tichotová, L.; Studenovska, H.; Petrovski, G.; Popelka, Š.; Nemesh, Y.; Sedláčková, M.; Drutovič, S.; Rohiwal, S.; Jendelová, P.; Erceg, S.; et al. Advantages of nanofibrous membranes for culturing of primary RPE cells compared to commercial scaffolds. Acta Ophthalmol. 2022, 100, E1172–E1185. [Google Scholar] [CrossRef]
- Stanzel, B.V.; Liu, Z.; Somboonthanakij, S.; Wongsawad, W.; Brinken, R.; Eter, N.; Corneo, B.; Holz, F.G.; Temple, S.; Stern, J.H.; et al. Human RPE Stem Cells Grown into Polarized RPE Monolayers on a Polyester Matrix Are Maintained after Grafting into Rabbit Subretinal Space. Stem Cell Rep. 2014, 2, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, G.L.; Benedicto, I.; Philp, N.J.; Rodriguez-Boulan, E. Plasma membrane protein polarity and trafficking in RPE cells: Past, present and future. Exp. Eye Res. 2014, 126, 5–15. [Google Scholar] [CrossRef]
- Brandl, C.; Zimmermann, S.J.; Milenkovic, V.M.; Rosendahl, S.M.G.; Grassmann, F.; Milenkovic, A.; Hehr, U.; Federlin, M.; Wetzel, C.H.; Helbig, H.; et al. In-depth characterisation of retinal pigment epithelium (RPE) cells derived from human induced pluripotent stem cells (hiPSC). NeuroMolecular Med. 2014, 16, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Blenkinsop, T.A.; Salero, E.; Stern, J.H.; Temple, S. The culture and maintenance of functional retinal pigment epithelial monolayers from adult human eye. Methods Mol. Biol. 2013, 945, 45–65. [Google Scholar] [CrossRef]
- Hu, X.; Calton, M.A.; Tang, S.; Vollrath, D. Depletion of Mitochondrial DNA in Differentiated Retinal Pigment Epithelial Cells. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Abberton, K.; Bortolotto, S.; Woods, A.; Findlay, M.; Morrison, W.; Thompson, E.; Messina, A. Myogel, a Novel, Basement Membrane-Rich, Extracellular Matrix Derived from Skeletal Muscle, Is Highly Adipogenic in vivo and in vitro. Cells Tissues Organs 2008, 188, 347–358. [Google Scholar] [CrossRef]
- Kelly, D.J.; Prendergast, P.J. Prediction of the optimal mechanical properties for a scaffold used in osteochondral defect repair. Tissue Eng.—Part B Rev. 2006, 12, 2509–2519. [Google Scholar] [CrossRef]
- Anghelina, M.; Krishnan, P.; Moldovan, L.; Moldovan, N.I. Monocytes/Macrophages Cooperate with Progenitor Cells during Neovascularization and Tissue Repair. Am. J. Pathol. 2006, 168, 529–541. [Google Scholar] [CrossRef]
- Bischoff, J. Approaches to studying cell adhesion molecules in angiogenesis. Trends Cell Biol. 1995, 5, 69–74. [Google Scholar] [CrossRef]
- Wiencke, A.K.; Kiilgaard, J.F.; Nicolini, J.; Bundgaard, M.; Röpke, C.; La Cour, M. Growth of cultured porcine retinal pigment epithelial cells Anne. Acta Ophthalmol. Scand. 2019, 60, 2384. [Google Scholar] [CrossRef]
- Shang, P.; Stepicheva, N.A.; Hose, S.; Zigler, J.S., Jr.; Sinha, D. Primary cell cultures from the mouse retinal pigment epithelium. J. Vis. Exp. 2018, 133, e56997. [Google Scholar] [CrossRef]
- Jarrett, S.G.; Lewin, A.S.; Boulton, M.E. The importance of Mitochondria in age-related and inherited eye disorders. Ophthalmic Res. 2010, 44, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Hyttinen, J.M.T.; Viiri, J.; Kaarniranta, K.; Błasiak, J. Mitochondrial quality control in AMD: Does mitophagy play a pivotal role? Cell. Mol. Life Sci. 2018, 75, 2991–3008. [Google Scholar] [CrossRef]
- Karunadharma, P.P.; Nordgaard, C.L.; Olsen, T.W.; Ferrington, D.A. Mitochondrial DNA damage as a potential mechanism for age-related macular degeneration. Investig. Opthalmol. Vis. Sci. 2010, 51, 5470–5479. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Wahlin, K.; Wan, J.; Hu, J.; Maruotti, J.; Yang, X.; Iacovelli, J.; Wolkow, N.; Kist, R.; Dunaief, J.L.; et al. Transcription Factor SOX9 Plays a Key Role in the Regulation of Visual Cycle Gene Expression in the Retinal Pigment. J. Biol. Chem. 2014, 289, 12908–12921. [Google Scholar] [CrossRef] [PubMed]
- Jadeja, R.N.; Thounaojam, M.C.; Bartoli, M.; Martin, P.M. Implications of NAD+ Metabolism in the Aging Retina and Retinal Degeneration. Oxidative Med. Cell. Longev. 2020, 2020, 2692794. [Google Scholar] [CrossRef]
- Fernandez-Godino, R.; Garland, D.L.; A Pierce, E. Isolation, culture and characterization of primary mouse RPE cells. Nat. Protoc. 2016, 11, 1206–1218. [Google Scholar] [CrossRef]
- Subramaniam, M.D.; Iyer, M.; Nair, A.P.; Venkatesan, D.; Mathavan, S.; Eruppakotte, N.; Kizhakkillach, S.; Chandran, M.K.; Roy, A.; Gopalakrishnan, A.V.; et al. Oxidative stress and mitochondrial transfer: A new dimension towards ocular diseases. Genes Dis. 2020, 9, 610–637. [Google Scholar] [CrossRef] [PubMed]
- Sparrrow, J.R.; Hicks, D.; Hamel, C.P. The Retinal Pigment Epithelium in Health and Disease. Curr. Mol. Med. 2010, 10, 802–823. [Google Scholar] [CrossRef]
- Brown, E.E.; DeWeerd, A.J.; Ildefonso, C.J.; Lewin, A.S.; Ash, J.D. Mitochondrial oxidative stress in the retinal pigment epithelium (RPE) led to metabolic dysfunction in both the RPE and retinal photoreceptors. Redox Biol. 2019, 24, 101201. [Google Scholar] [CrossRef]
- Ablonczy, Z.; Dahrouj, M.; Tang, P.H.; Liu, Y.; Sambamurti, K.; Marmorstein, A.D.; Crosson, C.E. Human retinal pigment epithelium cells as functional models for the RPE in vivo. Investig. Opthalmology Vis. Sci. 2011, 52, 8614–8620. [Google Scholar] [CrossRef] [PubMed]
- Storm, T.; Burgoyne, T.; Futter, C.E. Membrane trafficking in the retinal pigment epithelium at a glance. J. Cell Sci. 2020, 133, jcs238279. [Google Scholar] [CrossRef]
- Caceres, P.S.; Rodriguez-Boulan, E. Retinal pigment epithelium polarity in health and blinding diseases. Curr. Opin. Cell Biol. 2020, 62, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Muckersie, E.; Robertson, M.; Fraczek, M.; Forrester, J.V.; Xu, H. Characterization of a spontaneous mouse retinal pigment epithelial cell line B6-RPE07. Investig. Opthalmology Vis. Sci. 2008, 49, 3699–3706. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Elbrecht, D.H.; Long, C.J.; Hickman, J.J. Transepithelial/endothelial Electrical Resistance (TEER) theory and applications for microfluidic body-on-a-chip devices. J. Rare Dis. Res. Treat. 2016, 1, 46–52. [Google Scholar] [CrossRef]
- Yu-Wai-Man, P.; Newman, N.J. Inherited eye-related disorders due to mitochondrial dysfunction. Hum. Mol. Genet. 2017, 26, R12–R20. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of mitochondrial dna damage in ros-mediated pathogenesis of age-related macular degeneration (Amd). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Xu, H.; Liang, F.Q.; Liang, H.; Gupta, P.; Havey, A.N.; Boulton, M.E.; Godley, B.F. Mitochondrial DNA damage and repair in rpe associated with aging and age-related macular degeneration. Investig. Opthalmology Vis. Sci. 2011, 52, 3521–3529. [Google Scholar] [CrossRef]
- Terluk, M.R.; Kapphahn, R.J.; Soukup, L.M.; Gong, H.; Gallardo, C.; Montezuma, S.R.; Ferrington, D.A. Investigating mitochondria as a target for treating age-related macular degeneration. J. Neurosci. 2015, 35, 7304–7311. [Google Scholar] [CrossRef]
- Heller, J.P.; Kwok, J.C.F.; Vecino, E.; Martin, K.R.; Fawcett, J.W. A method for the isolation and culture of adult rat retinal pigment epithelial (RPE) cells to study retinal diseases. Front. Cell. Neurosci. 2015, 9, 449. [Google Scholar] [CrossRef]
- Liggett, T.E.; Griffiths, T.D.; Gaillard, E.R. Isolation and characterization of a spontaneously immortalized bovine retinal pigmented epithelial cell line. BMC Cell Biol. 2009, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Conley, S.M.; Naash, M.I. RPE65: Role in the visual cycle, human retinal disease, and gene therapy. Ophthalmic Genet. 2009, 30, 57–62. [Google Scholar] [CrossRef]
- Szatmári-Tóth, M.; Kristóf, E.; Veréb, Z.; Akhtar, S.; Facskó, A.; Fésüs, L.; Kauppinen, A.; Kaarniranta, K.; Petrovski, G. Clearance of autophagy-associated dying retinal pigment epithelial cells—A possible source for inflammation in age-related macular degeneration. Cell Death Dis. 2016, 7, e2367. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Scheffler, K.; Esbensen, Y.; Eide, L. Quantification of DNA damage by real-time qPCR. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; pp. 27–32. [Google Scholar] [CrossRef]
- Wang, W.; Esbensen, Y.; Scheffler, K.; Eide, L. Analysis of Mitochondrial DNA and RNA Integrity by a Real-Time qPCR-Based Method. In Mitochondrial Medicine; Humana Press: New York, NY, USA, 2015; pp. 97–106. [Google Scholar] [CrossRef]
- Castellani, C.A.; Longchamps, R.J.; Sun, J.; Guallar, E.; Arking, D.E. Thinking outside the nucleus: Mitochondrial DNA copy number in health and disease. Mitochondrion 2020, 53, 214–223. [Google Scholar] [CrossRef]
- Longchamps, R.J.; Castellani, C.A.; Yang, S.Y.; Newcomb, C.E.; Sumpter, J.A.; Lane, J.; Grove, M.L.; Guallar, E.; Pankratz, N.; Taylor, K.D.; et al. Evaluation of mitochondrial DNA copy number estimation techniques. PLoS ONE 2020, 15, 1–14. [Google Scholar] [CrossRef]
- Vaajasaari, H.; Ilmarinen, T.; Juuti-Uusitalo, K.; Rajala, K.; Onnela, N.; Narkilahti, S.; Suuronen, R.; Hyttinen, J.; Uusitalo, H.; Skottman, H. Toward the defined and xeno-free differentiation of functional human pluripotent stem cell-derived retinal pigment epithelial cells. Mol. Vis. 2011, 22, 558–575. [Google Scholar]
- Carr, A.-J.; Vugler, A.A.; Yu, L.; Semo, M.; Coffey, P.; Moss, S.E.; Greenwood, J. The expression of retinal cell markers in human retinal pigment epithelial cells and their augmentation by the synthetic retinoid fenretinide. Mol. Vis. 2011, 17, 1701–1715. [Google Scholar] [PubMed]
- Burke, J.M. Epithelial phenotype and the RPE: Is the answer blowing in the Wnt? Prog. Retin. Eye Res. 2008, 27, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zonca, M.R.; Imbrogno, J.; Unser, A.M.; Sfakis, L.; Temple, S.; Belfort, G.; Xie, Y. Polarized, Cobblestone, Human Retinal Pigment Epithelial Cell Maturation on a Synthetic PEG Matrix. ACS Biomater. Sci. Eng. 2017, 3, 890–902. [Google Scholar] [CrossRef]
- Harris, T.I.; Paterson, C.A.; Farjood, F.; Wadsworth, I.D.; Caldwell, L.; Lewis, R.V.; Jones, J.A.; Vargis, E. Utilizing Recombinant Spider Silk Proteins to Develop a Synthetic Bruch’s Membrane for Modeling the Retinal Pigment Epithelium. ACS Biomater. Sci. Eng. 2019, 5, 4023–4036. [Google Scholar] [CrossRef]
- Rosales, M.A.B.; Shu, D.Y.; Iacovelli, J.; Saint-Geniez, M. Loss of PGC-1α in RPE induces mesenchymal transition and promotes retinal degeneration. Life Sci. Alliance 2019, 2, e201800212. [Google Scholar] [CrossRef]
- Youssef, P.N.; Sheibani, N.; Albert, D.M. Retinal light toxicity. Eye 2011, 25, 1–14. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, J.; Li, D. Functions and Diseases of the Retinal Pigment Epithelium. Front. Pharmacol. 2021, 12, 727870. [Google Scholar] [CrossRef]
- Yeste, J.; Illa, X.; Gutiérrez, C.; Solé, M.; Guimerà, A.; Villa, R. Geometric correction factor for transepithelial electrical resistance measurements in transwell and microfluidic cell cultures. J. Phys. D Appl. Phys. 2016, 49, 375401. [Google Scholar] [CrossRef]
- Masuda, T.; Esumi, N. SOX9, through interaction with microphthalmia-associated transcription factor (MITF) and OTX2, regulates BEST1 expression in the retinal pigment epithelium. J. Biol. Chem. 2010, 285, 26933–26944. [Google Scholar] [CrossRef]
- Seo, J.-M.; Kim, S.J.; Chung, H.; Kim, E.T.; Yu, H.G.; Yu, Y.S. Biocompatibility of polyimide microelectrode array for retinal stimulation. Mater. Sci. Eng. C 2004, 24, 185–189. [Google Scholar] [CrossRef]
- Kenney, M.C.; Riazi-Esfahani, M.; Kuppermann, B.D. The Role of Mitochondria in AMD: Current Knowledge and Future Applications. J. Ophthalmic Vis. Res. 2017, 12, 424–428. [Google Scholar] [CrossRef]
- Feher, J.; Kovacs, I.; Artico, M.; Cavallotti, C.; Papale, A.; Gabrieli, C.B. Mitochondrial alterations of retinal pigment epithelium in age-related macular degeneration. Neurobiol. Aging 2006, 27, 983–993. [Google Scholar] [CrossRef]
- Zhang, R.; Engel, A.L.; Wang, Y.; Li, B.; Shen, W.; Gillies, M.C.; Chao, J.R.; Du, J. Inhibition of Mitochondrial Respiration Impairs Nutrient Consumption and Metabolite Transport in Human Retinal Pigment Epithelium. J. Proteome Res. 2021, 20, 909–922. [Google Scholar] [CrossRef]
- Strauß, O.; Müller, C.; Reichhart, N.; Tamm, E.R.; Gomez, N.M. The Role of Bestrophin-1 in Intracellular Ca2+ Signaling; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Hollyfield, J.G.; Anderson, R.E.; LaVail, M.M. Retinal Degenerative Diseases: Preface; Springer: New York, NY, USA, 2012. [Google Scholar]
- Liang, F.; Godley, B.F. Oxidative stress-induced mitochondrial DNA damage in human retinal pigment epithelial cells: A possible mechanism for RPE aging and age-related macular degeneration. Exp. Eye Res. 2003, 76, 397–403. [Google Scholar] [CrossRef]
- Rooney, J.P.; Ryde, I.T.; Sanders, L.H.; Howlett, E.H.; Colton, M.D.; Germ, K.E.; Mayer, G.D.; Greenamyre, J.T.; Meyer, J.N. PCR Based Determination of Mitochondrial DNA Copy Number in Multiple Species. Methods Mol. Biol. 2015, 1241, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Castellani, C.A.; Longchamps, R.J.; Sumpter, J.A.; Newcomb, C.E.; Lane, J.A.; Grove, M.L.; Bressler, J.; Brody, J.A.; Floyd, J.S.; Bartz, T.M.; et al. Mitochondrial DNA copy number can influence mortality and cardiovascular disease via methylation of nuclear DNA CpGs. Genome Med. Biomark. Health Sci. 2020, 12, 84. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faura, G.; Studenovska, H.; Sekac, D.; Ellederova, Z.; Petrovski, G.; Eide, L. The Effects of the Coating and Aging of Biodegradable Polylactic Acid Membranes on In Vitro Primary Human Retinal Pigment Epithelium Cells. Biomedicines 2024, 12, 966. https://doi.org/10.3390/biomedicines12050966
Faura G, Studenovska H, Sekac D, Ellederova Z, Petrovski G, Eide L. The Effects of the Coating and Aging of Biodegradable Polylactic Acid Membranes on In Vitro Primary Human Retinal Pigment Epithelium Cells. Biomedicines. 2024; 12(5):966. https://doi.org/10.3390/biomedicines12050966
Chicago/Turabian StyleFaura, Georgina, Hana Studenovska, David Sekac, Zdenka Ellederova, Goran Petrovski, and Lars Eide. 2024. "The Effects of the Coating and Aging of Biodegradable Polylactic Acid Membranes on In Vitro Primary Human Retinal Pigment Epithelium Cells" Biomedicines 12, no. 5: 966. https://doi.org/10.3390/biomedicines12050966