The Glucosinolates: A Sulphur Glucoside Family of Mustard Anti-Tumour and Antimicrobial Phytochemicals of Potential Therapeutic Application


Abstract
1. Introduction
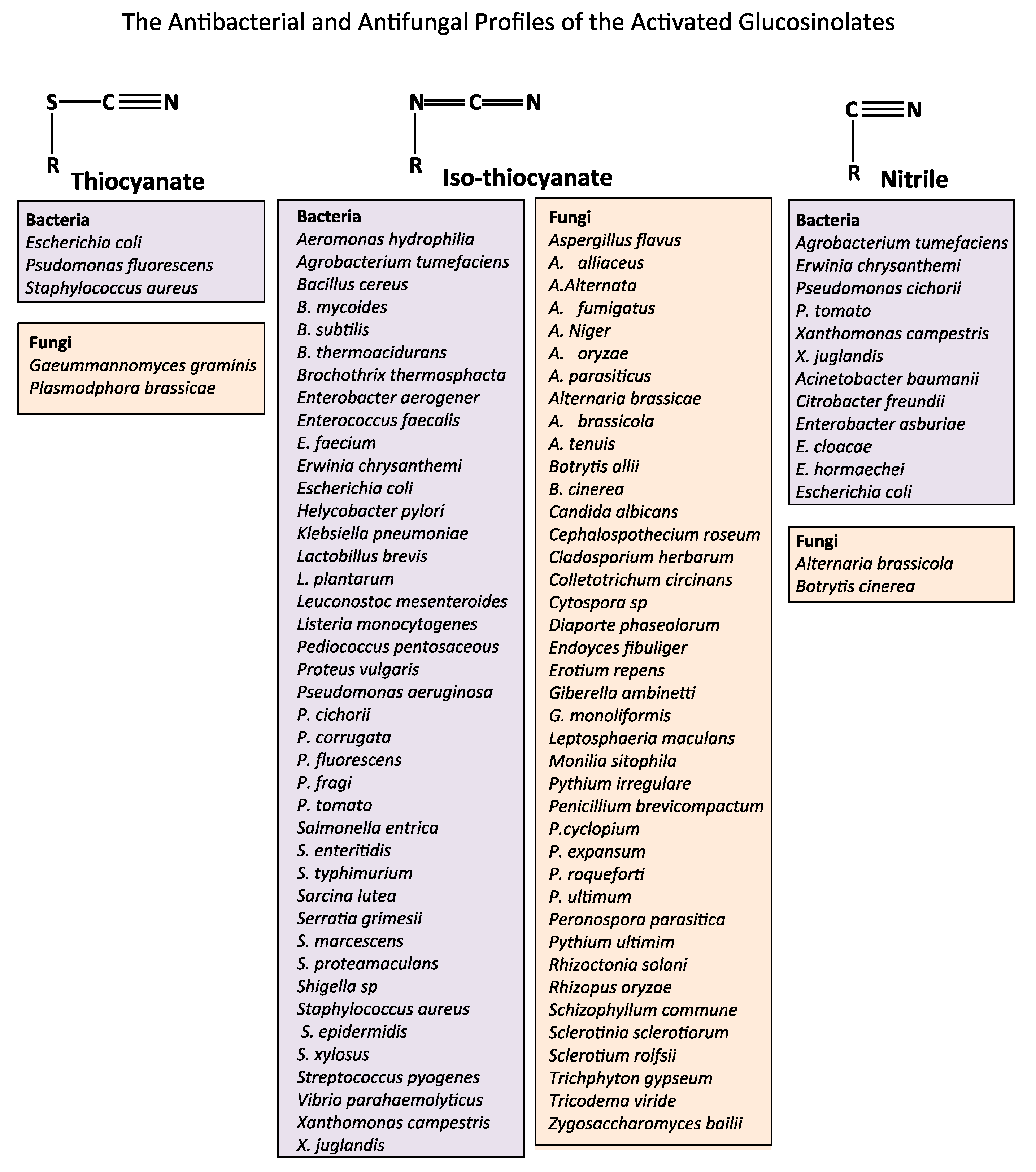
2. The Natural Anti-Microbial Activity of Glucosinolate Rich Foods
3. The Brassicaceae Family of Plants
4. Public Health Concern over the Impact of Antibiotic-Resistant Bacteria
4.1. Treatment of Antibiotic-Resistant Bacterial Infections
4.2. Commercial Development of Antibiotics
5. WHO, United Nation and World Bank Programmes and Coordinated Interagency Collaborations Designed to Combat Antibiotic-Resistant Bacteria
6. Application of the Myrosinase-Glucosinolate System in Biomedicine
6.1. The Bioactivity of Glucosinolates
6.2. Cancer and Dietary SFN and AITC Levels
6.3. The Beneficial Bioactivities of Sinigrin and Their Applications in Biomedicine
7. Concluding Remarks
- Antibiotics, anti-fungal and anti-viral agents
- Biofilm prevention in medical implants, catheters and industrial plant equipment
- Nutritive additives with anti-cancer properties
- Advanced food packaging technology to improve shelf-life of food products.
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AKT | A serine/threonine-specific protein kinase |
| ARE | Antioxidant response element |
| EPA | Environment Protection Agency |
| ESBL | Extended Spectrum Beta-Lactamases |
| Keap-1-Nrf2-ARE | Kelch-like erythroid cell-derived protein with CNC homology (ECH)-associated protein 1–anti-oxidant response element |
| AITC | Allyl isothiocyanate |
| GARDP | Global Antibiotic Research and Development Partnership |
| GSK | Glycogen Synthase Kinase |
| GST | Glutathione-S-transferase |
| DNDI | Drugs for Neglected Diseases initiative |
| IACG | Interagency Coordination Group on Antimicrobial Resistance |
| LPS | Lipopolysaccharide |
| MAPK | A mitogen-activated protein kinase |
| NFκB | Nuclear factor kappa light chain enhancer of activated B cells |
| NLRPR3 | Nucleotide-binding domain and leucine-rich repeat–containing protein 3 |
| NSAID | Non-Steroidal anti-inflammatory |
| PDGF | Platelet derived growth factor |
| ROS | Reactive oxygen species |
| SMC | Smooth muscle cell |
| TNF | Tumour necrosis factor-alpha |
References
- Briones-Herrera, A.; Eugenio-Perez, D.; Reyes-Ocampo, J.G.; Rivera-Mancia, S.; Pedraza-Chaverri, J. New highlights on the health-improving effects of sulforaphane. Food. Funct. 2018, 9, 2589–2606. [Google Scholar] [CrossRef] [PubMed]
- Palliyaguru, D.L.; Yuan, J.M.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, e1700965. [Google Scholar] [CrossRef] [PubMed]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from Broccoli, Sulforaphane, and Its Properties. J. Med. Food 2019, 22, 121–126. [Google Scholar] [CrossRef]
- Aires, A.; Mota, V.R.; Saavedra, M.J.; Rosa, E.A.; Bennett, R.N. The antimicrobial effects of glucosinolates and their respective enzymatic hydrolysis products on bacteria isolated from the human intestinal tract. J. Appl. Microbiol. 2009, 106, 2086–2095. [Google Scholar] [CrossRef] [PubMed]
- Baskar, V.; Park, S.W.; Nile, S.H. An Update on Potential Perspectives of Glucosinolates on Protection against Microbial Pathogens and Endocrine Dysfunctions in Humans. Crit. Rev. Food Sci. Nutr. 2016, 56, 2231–2249. [Google Scholar] [CrossRef]
- Becker, T.M.; Juvik, J.A. The Role of Glucosinolate Hydrolysis Products from Brassica Vegetable Consumption in Inducing Antioxidant Activity and Reducing Cancer Incidence. Diseases 2016, 4, 22. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Ferreira, C.; Saavedra, M.J.; Simoes, L.C.; Simoes, M. Antibacterial activity and mode of action of selected glucosinolate hydrolysis products against bacterial pathogens. J. Food Sci. Technol. 2015, 52, 4737–4748. [Google Scholar] [CrossRef]
- Carpenter, E.; Mai, N.; Miranda, C.L.; Reed, R.L.; Stevens, J.F.; Indra, A.K.; Indra, G. Photoprotective Properties of Isothiocyanate and Nitrile Glucosinolate Derivatives from Meadowfoam (Limnanthes alba) against UVB Irradiation in Human Skin Equivalent. Front. Pharmacol. 2018, 9, 477. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Dufour, V.; Alazzam, B.; Ermel, G.; Thepaut, M.; Rossero, A.; Tresse, O.; Baysse, C. Antimicrobial activities of isothiocyanates against Campylobacter jejuni isolates. Front. Cell Infect. Microbiol. 2012, 2, 53. [Google Scholar] [CrossRef]
- Dufour, V.; Stahl, M.; Baysse, C. The antibacterial properties of isothiocyanates. Microbiology 2015, 161, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Gamet-Payrastre, L.; Li, P.; Lumeau, S.; Cassar, G.; Dupont, M.A.; Chevolleau, S.; Gasc, N.; Tulliez, J.; Terce, F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000, 60, 1426–1433. [Google Scholar] [PubMed]
- Attokaran, M. Mustard, Natural Food Flavors and Colorants; Wiley-Blackwell: Hoboken, NJ, USA, 2011. [Google Scholar]
- Ekanayake, A.; Strife, R.J.; Zehentbauer, G.N.; David, J.R.D. Chapter 98 - Yellow or White Mustard (Sinapis alba L.) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 857–863. [Google Scholar]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Quiles, J.M.; Manyes, L.; Luciano, F.B.; Manes, J.; Meca, G. Effect of the oriental and yellow mustard flours as natural preservative against aflatoxins B1, B2, G1 and G2 production in wheat tortillas. J. Food Sci. Technol. 2015, 52, 8315–8321. [Google Scholar] [CrossRef] [PubMed]
- Suhr, K.I.; Nielsen, P.V. Antifungal activity of essential oils evaluated by two different application techniques against rye bread spoilage fungi. J. Appl. Microbiol. 2003, 94, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Suhr, K.I.; Nielsen, P.V. Inhibition of Fungal Growth on Wheat and Rye Bread by Modified Atmosphere Packaging and Active Packaging Using Volatile Mustard Essential Oil. J. Food Sci. 2005, 70, M37–M44. [Google Scholar] [CrossRef]
- Nielsen, P.V.; Rios, R. Inhibition of fungal growth on bread by volatile components from spices and herbs, and the possible application in active packaging, with special emphasis on mustard essential oil. Int. J. Food Microbiol. 2000, 60, 219–229. [Google Scholar] [CrossRef]
- Steinbrecher, A.; Linseisen, J. Dietary intake of individual glucosinolates in participants of the EPIC-Heidelberg cohort study. Ann. Nutr. Metab. 2009, 54, 87–96. [Google Scholar] [CrossRef]
- Ngala, B.M.; Haydock, P.P.; Woods, S.; Back, M.A. Biofumigation with Brassica juncea, Raphanus sativus and Eruca sativa for the management of field populations of the potato cyst nematode Globodera pallida. Pest Manag. Sci. 2015, 71, 759–769. [Google Scholar] [CrossRef]
- Eutrema japonicum (Miq) Koidz—The plant list. Available online: http://www.theplantlist.org/tpl1.1/record/kew-2805492 (accessed on 31 May 2019).
- Wasabia japonica. Multilingual multiscript plant nama database—University of Melbourne. Available online: http://www.plantnames.unimelb.edu.au/ (accessed on 31 May 2019).
- Verboven, P.; Herremans, E.; Borisjuk, L.; Helfen, L.; Ho, Q.T.; Tschiersch, H.; Fuchs, J.; Nicolai, B.M.; Rolletschek, H. Void space inside the developing seed of Brassica napus and the modelling of its function. New Phytol. 2013, 199, 936–947. [Google Scholar] [CrossRef]
- Linnaeus, C. Classification of Brassica napus. Species Plantarum 1753, 2, 666. [Google Scholar]
- Harper, D. Derivation of the name rape-seed. OnlineEtymology Dictionary. 2016. Available online: www.etymonline.com (accessed on 12 May 2019).
- Saul, H.; Madella, M.; Fischer, A.; Glykou, A.; Hartz, S.; Craig, O.E. Phytoliths in pottery reveal the use of spice in European prehistoric cuisine. PLoS ONE 2013, 8, e70583. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.K.; Yadav, S.; Vats, V. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Gupta, R.; Bajpai, K.G.; Johri, S.; Saxena, A.M. An overview of Indian novel traditional medicinal plants with anti-diabetic potentials. Afr. J. Tradit. Complement. Altern. Med. 2007, 5, 1–17. [Google Scholar] [PubMed]
- Mali, R.G. Cleome viscosa (wild mustard): A review on ethnobotany, phytochemistry, and pharmacology. Pharm. Biol. 2010, 48, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Y.; Maibam, B.C.; Biswas, D.; Laisharm, S.; Deb, L.; Talukdar, N.C.; Borah, J.C. Anti-diabetic potential of selected ethno-medicinal plants of north east India. J. Ethnopharmacol. 2015, 171, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, M. Crismer values and erucic scid contents of rapeseed oils. J. Am. Oil Chem. Soc. 1977, 54, 323–324. [Google Scholar] [CrossRef]
- Potts, D.; Rakow, G.W.; Males, D.R. Canola quality Brassica juncea, a new oilseed crop for the Canadian prairies. New Horizons for an old crop. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999. [Google Scholar]
- Chen, N.G.; Chen, K.T.; Lu, C.C.; Lan, Y.H.; Lai, C.H.; Chung, Y.T.; Yang, J.S.; Lin, Y.C. Allyl isothiocyanate triggers G2/M phase arrest and apoptosis in human brain malignant glioma GBM 8401 cells through a mitochondria-dependent pathway. Oncol. Rep. 2010, 24, 449–455. [Google Scholar]
- Giacoppo, S.; Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. (RS)-glucoraphanin purified from Tuscan black kale and bioactivated with myrosinase enzyme protects against cerebral ischemia/reperfusion injury in rats. Fitoterapia 2014, 99, 166–177. [Google Scholar] [CrossRef]
- Giacoppo, S.; Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. The protective effects of bioactive (RS)-glucoraphanin on the permeability of the mice blood-brain barrier following experimental autoimmune encephalomyelitis. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 194–204. [Google Scholar]
- Giacoppo, S.; Galuppo, M.; Montaut, S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. An overview on neuroprotective effects of isothiocyanates for the treatment of neurodegenerative diseases. Fitoterapia 2015, 106, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Beltran, C.E.; Calderon-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Sanadgol, N.; Zahedani, S.S.; Sharifzadeh, M.; Khalseh, R.; Barbari, G.R.; Abdollahi, M. Recent Updates in Imperative Natural Compounds for Healthy Brain and Nerve Function: A Systematic Review of Implications for Multiple Sclerosis. Curr. Drug Targets 2017, 18, 1499–1517. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Fujita, Y.; Hashimoto, R.; Ohi, K.; Yamamori, H.; Yasuda, Y.; Ishima, T.; Suganuma, H.; Ushida, Y.; Takeda, M.; et al. Dietary Intake of Sulforaphane-Rich Broccoli Sprout Extracts during Juvenile and Adolescence Can Prevent Phencyclidine-Induced Cognitive Deficits at Adulthood. PLoS ONE 2015, 10, e0127244. [Google Scholar] [CrossRef] [PubMed]
- Tarozzi, A.; Angeloni, C.; Malaguti, M.; Morroni, F.; Hrelia, S.; Hrelia, P. Sulforaphane as a potential protective phytochemical against neurodegenerative diseases. Oxid. Med. Cell Longev. 2013, 2013, 415078. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Buonfiglio, M.; Corona, G.; Chirafisi, J.; Vafeiadou, K.; Angeloni, C.; Hrelia, S.; Hrelia, P.; Spencer, J.P. Sulforaphane protects cortical neurons against 5-S-cysteinyl-dopamine-induced toxicity through the activation of ERK1/2, Nrf-2 and the upregulation of detoxification enzymes. Mol. Nutr. Food Res. 2010, 54, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Zhang, J.C.; Ishima, T.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Suganuma, H.; et al. Role of Keap1-Nrf2 signaling in depression and dietary intake of glucoraphanin confers stress resilience in mice. Sci. Rep. 2016, 6, 30659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Ushida, Y.; Suganuma, H.; et al. Prophylactic effects of sulforaphane on depression-like behavior and dendritic changes in mice after inflammation. J. Nutr. Biochem. 2017, 39, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Freitas, H.R.; Ferreira, G.D.C.; Trevenzoli, I.H.; Oliveira, K.J.; de Melo Reis, R.A. Fatty Acids, Antioxidants and Physical Activity in Brain Aging. Nutrients 2017, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.; Aires, A.; Bennett, R.N.; Rosa, E.A.; Saavedra, M.J. First study on antimicriobial activity and synergy between isothiocyanates and antibiotics against selected Gram-negative and Gram-positive pathogenic bacteria from clinical and animal source. Med. Chem. 2012, 8, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Ardal, C.; Baraldi, E.; Theuretzbacher, U.; Outterson, K.; Plahte, J.; Ciabuschi, F.; Rottingen, J.A. Insights into early stage of antibiotic development in small- and medium-sized enterprises: A survey of targets, costs, and durations. J. Pharm. Policy Pract. 2018, 11, 8. [Google Scholar] [CrossRef]
- Anonymous. Wanted: A reward for antibiotic development. Nat. Biotechnol. 2018, 36, 555. [Google Scholar] [CrossRef] [PubMed]
- Renwick, M.J.; Simpkin, V.; Mossialos, E. Targeting innovation in antibiotic drug discovery and development: The need for a One Health – One Europe – One World Framework; European Observatory on Health Systems and Policies: Copenhagen, Denmark, 2016. Available online: http://www.ncbi.nlm.nih.gov/books/NBK447337/ (accessed on 19 August 2019).
- Cruz-Muniz, M.Y.; Lopez-Jacome, L.E.; Hernandez-Duran, M.; Franco-Cendejas, R.; Licona-Limon, P.; Ramos-Balderas, J.L.; Martinez-Vazquez, M.; Belmont-Diaz, J.A.; Wood, T.K.; Garcia-Contreras, R. Repurposing the anticancer drug mitomycin C for the treatment of persistent Acinetobacter baumannii infections. Int. J. Antimicrob. Agents 2017, 49, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Vega, A.; Bernstein, L.R.; Mandujano-Tinoco, E.A.; Garcia-Contreras, S.J.; Garcia-Contreras, R. Drug repurposing as an alternative for the treatment of recalcitrant bacterial infections. Front. Microbiol. 2015, 6, 282. [Google Scholar] [CrossRef] [PubMed]
- Soo, V.W.; Kwan, B.W.; Quezada, H.; Castillo-Juarez, I.; Perez-Eretza, B.; Garcia-Contreras, S.J.; Martinez-Vazquez, M.; Wood, T.K.; Garcia-Contreras, R. Repurposing of Anticancer Drugs for the Treatment of Bacterial Infections. Curr. Top. Med. Chem. 2017, 17, 1157–1176. [Google Scholar] [CrossRef] [PubMed]
- Van Nuffel, A.M.; Sukhatme, V.; Pantziarka, P.; Meheus, L.; Sukhatme, V.P.; Bouche, G. Repurposing Drugs in Oncology (ReDO)-clarithromycin as an anti-cancer agent. Ecancermedicalscience 2015, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Fry, D.E.; Barie, P.S. The changing face of Staphylococcus aureus: A continuing surgical challenge. Surg. Infect. 2011, 12, 191–203. [Google Scholar] [CrossRef]
- Velazquez-Meza, M.E.; Hernandez-Salgado, M.; Contreras-Cordero, J.F.; Perez-Cortes, P.; Villarreal-Trevino, L. Surveillance of methicillin-resistant Staphylococcus aureus causing nosocomial infections in five medical centers of Monterrey, Nuevo Leon, Mexico from 2005-2009. Arch. Med. Res. 2013, 44, 570–574. [Google Scholar] [CrossRef]
- Reid, K.C.; Cockerill, I.F.; Patel, R. Clinical and epidemiological features of Enterococcus casseliflavus/flavescens and Enterococcus gallinarum bacteremia: A report of 20 cases. Clin. Infect. Dis. 2001, 32, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.J.; Chen, C.C.; Zhang, C.C.; Su, B.A.; Li, C.M.; Weng, T.C.; Chiang, S.R.; Ko, W.C.; Chuang, Y.C. In vitro efficacy of fosfomycin-based combinations against clinical vancomycin-resistant Enterococcus isolates. Diagn. Microbiol. Infect. Dis. 2013, 77, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.C.; Borges, A.; Simoes, L.C.; Saavedra, M.J.; Simoes, M. Antibacterial activity of phenyl isothiocyanate on Escherichia coli and Staphylococcus aureus. Med. Chem. 2013, 9, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; Nicola, G.R.; Iori, R.; Dell’utri, P.; Bramanti, P.; Mazzon, E. Antibacterial activity of glucomoringin bioactivated with myrosinase against two important pathogens affecting the health of long-term patients in hospitals. Molecules 2013, 18, 14340–14348. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Holley, R.A. Inhibition of Listeria monocytogenes and Salmonella by combinations of oriental mustard, malic acid, and EDTA. J. Food Sci. 2014, 79, M614–M621. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.J.; Borges, A.; Dias, C.; Aires, A.; Bennett, R.N.; Rosa, E.S.; Simoes, M. Antimicrobial activity of phenolics and glucosinolate hydrolysis products and their synergy with streptomycin against pathogenic bacteria. Med. Chem. 2010, 6, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.J.; Dias, C.S.; Martinez-Murcia, A.; Bennett, R.N.; Aires, A.; Rosa, E.A. Antibacterial effects of glucosinolate-derived hydrolysis products against enterobacteriaceae and enterococci isolated from pig ileum segments. Foodborne Pathog. Dis. 2012, 9, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, T.; Lema, M.; Soengas, P.; Cartea, M.E.; Velasco, P. In vitro activity of glucosinolates and their degradation products against brassica-pathogenic bacteria and fungi. Appl. Environ. Microbiol. 2015, 81, 432–440. [Google Scholar] [CrossRef]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef]
- World Health Organisation. Guidelines for the prevention and control of carbapenem-resistant Enterobacteriaceae, Acinetobacter baumannii and Pseudomonas aeruginosa in health care facilities. 2017. Available online: https://www.who.int/infection-prevention/publications/guidelines-cre/en/ (accessed on 19 August 2019).
- Adegoke, A.A.; Faleye, A.C.; Singh, G.; Stenstrom, T.A. Antibiotic Resistant Superbugs: Assessment of the Interrelationship of Occurrence in Clinical Settings and Environmental Niches. Molecules 2016, 22, 29. [Google Scholar] [CrossRef]
- Mohammed, N.; Savardekar, A.R.; Patra, D.P.; Narayan, V.; Nanda, A. The 21st-century challenge to neurocritical care: The rise of the superbug Acinetobacter baumannii. A meta-analysis of the role of intrathecal or intraventricular antimicrobial therapy in reduction of mortality. Neurosurg. Focus 2017, 43, E8. [Google Scholar] [CrossRef] [PubMed]
- Rello, J.; Kalwaje Eshwara, V.; Lagunes, L.; Alves, J.; Wunderink, R.G.; Conway-Morris, A.; Rojas, J.N.; Alp, E.; Zhang, Z. A global priority list of the TOp TEn resistant Microorganisms (TOTEM) study at intensive care: A prioritization exercise based on multi-criteria decision analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Righi, E.; Carnelutti, A.; Graziano, E.; Russo, A. Multidrug-resistant Klebsiella pneumoniae: Challenges for treatment, prevention and infection control. Expert. Rev. Anti-Infect. Ther. 2018, 16, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Goldstein, E.J.; Wise, J.; Bilker, W.B.; Tolomeo, P.; Lautenbach, E. Epidemiology of Carbapenem-Resistant Klebsiella pneumoniae in a Network of Long-Term Acute Care Hospitals. Clin. Infect. Dis. 2017, 64, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Gordon, M.; Villarreal, E.; Frasquet, J.; Sanchez, M.A.; Martin, M.; Castellanos, A.; Ramirez, P. Influence of antibiotic pressure on multi-drug resistant Klebsiella pneumoniae colonisation in critically ill patients. Antimicrob. Resist. Infect. Control. 2019, 8, 38. [Google Scholar] [CrossRef]
- Akturk, H.; Sutcu, M.; Somer, A.; Aydin, D.; Cihan, R.; Ozdemir, A.; Coban, A.; Ince, Z.; Citak, A.; Salman, N. Carbapenem-resistant Klebsiella pneumoniae colonization in pediatric and neonatal intensive care units: Risk factors for progression to infection. Braz. J. Infect. Dis. 2016, 20, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Justo-da-Silva, L.H.; De-Azeredo, A.N.; Bueno, A.C.; Montezzi, L.F.; Leobons, M.; Alves, M.S.; de Souza Inhaquite, P.; Santos, R.R.; Girao, V.B.C.; da Cunha, A.; et al. Diversity of clonal types of Klebsiella pneumoniae causing infections in intensive care neonatal patients in a large urban setting. Braz. J. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Vergadi, E.; Bitsori, M.; Maraki, S.; Galanakis, E. Community-onset carbapenem-resistant Klebsiella pneumoniae urinary tract infections in infancy following NICU hospitalisation. J. Pediatr. Urol. 2017, 13, 495.e491–495.e496. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.L.; da Silva, B.C.M.; Rezende, G.S.; Nakamura-Silva, R.; Pitondo-Silva, A.; Campanini, E.B.; Brito, M.C.A.; da Silva, E.M.L.; Freire, C.C.M.; da Cunha, A.F.; et al. High Prevalence of Multidrug-Resistant Klebsiella pneumoniae Harboring Several Virulence and beta-Lactamase Encoding Genes in a Brazilian Intensive Care Unit. Front. Microbiol. 2018, 9, 3198. [Google Scholar] [CrossRef] [PubMed]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef]
- Shankar, C.; Nabarro, L.E.; Anandan, S.; Ravi, R.; Babu, P.; Munusamy, E.; Jeyaseelan, V.; Rupali, P.; Verghese, V.P.; Veeraraghavan, B. Extremely High Mortality Rates in Patients with Carbapenem-resistant, Hypermucoviscous Klebsiella pneumoniae Blood Stream Infections. J. Assoc. Physicians India 2018, 66, 13–16. [Google Scholar] [PubMed]
- Aygun, F.; Aygun, F.D.; Varol, F.; Durak, C.; Cokugras, H.; Camcioglu, Y.; Cam, H. Infections with Carbapenem-Resistant Gram-Negative Bacteria are a Serious Problem Among Critically Ill Children: A Single-Centre Retrospective Study. Pathogens 2019, 8, 69. [Google Scholar] [CrossRef]
- Brinkac, L.M.; White, R.; D’Souza, R.; Nguyen, K.; Obaro, S.K.; Fouts, D.E. Emergence of New Delhi Metallo-beta-Lactamase (NDM-5) in Klebsiella quasipneumoniae from Neonates in a Nigerian Hospital. mSphere 2019, 4. [Google Scholar] [CrossRef]
- Karampatakis, T.; Tsergouli, K.; Politi, L.; Diamantopoulou, G.; Iosifidis, E.; Antachopoulos, C.; Karyoti, A.; Mouloudi, E.; Tsakris, A.; Roilides, E. Molecular Epidemiology of Endemic Carbapenem-Resistant Gram-Negative Bacteria in an Intensive Care Unit. Microb. Drug Resist. 2019, 25, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Sirijatuphat, R.; Sripanidkulchai, K.; Boonyasiri, A.; Rattanaumpawan, P.; Supapueng, O.; Kiratisin, P.; Thamlikitkul, V. Implementation of global antimicrobial resistance surveillance system (GLASS) in patients with bacteremia. PLoS ONE 2018, 13, e0190132. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, P.; Unni, S.; Krishnappa, G.; Padmanabhan, B. The Keap1-Nrf2 pathway: Promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys. Rev. 2017, 9, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Fimognari, C.; Nusse, M.; Cesari, R.; Iori, R.; Cantelli-Forti, G.; Hrelia, P. Growth inhibition, cell-cycle arrest and apoptosis in human T-cell leukemia by the isothiocyanate sulforaphane. Carcinogenesis 2002, 23, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Lai, B.; Yuan, Q. Sulforaphane induces cell-cycle arrest and apoptosis in cultured human lung adenocarcinoma LTEP-A2 cells and retards growth of LTEP-A2 xenografts in vivo. J. Nat. Prod. 2008, 71, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.A.; Jacobberger, J.W.; Schimmer, A.D.; Cao, P.; Gronda, M.; Hedley, D.W. The dietary isothiocyanate sulforaphane targets pathways of apoptosis, cell cycle arrest, and oxidative stress in human pancreatic cancer cells and inhibits tumor growth in severe combined immunodeficient mice. Mol. Cancer Ther. 2004, 3, 1239–1248. [Google Scholar] [PubMed]
- Tang, L.; Zhang, Y.; Jobson, H.E.; Li, J.; Stephenson, K.K.; Wade, K.L.; Fahey, J.W. Potent activation of mitochondria-mediated apoptosis and arrest in S and M phases of cancer cells by a broccoli sprout extract. Mol. Cancer Ther. 2006, 5, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Xiao, D.; Lew, K.L.; Dhir, R.; Singh, S.V. Sulforaphane induces caspase-mediated apoptosis in cultured PC-3 human prostate cancer cells and retards growth of PC-3 xenografts in vivo. Carcinogenesis 2004, 25, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Azarenko, O.; Okouneva, T.; Singletary, K.W.; Jordan, M.A.; Wilson, L. Suppression of microtubule dynamic instability and turnover in MCF7 breast cancer cells by sulforaphane. Carcinogenesis 2008, 29, 2360–2368. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.J.; Singletary, K.W. Sulforaphane: A naturally occurring mammary carcinoma mitotic inhibitor, which disrupts tubulin polymerization. Carcinogenesis 2004, 25, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Dashwood, R.H.; Ho, E. Dietary agents as histone deacetylase inhibitors: Sulforaphane and structurally related isothiocyanates. Nutr. Rev. 2008, 66 (Suppl. 1), S36–S38. [Google Scholar] [CrossRef]
- Gibbs, A.; Schwartzman, J.; Deng, V.; Alumkal, J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc. Natl. Acad. Sci. USA 2009, 106, 16663–16668. [Google Scholar] [CrossRef]
- Myzak, M.C.; Hardin, K.; Wang, R.; Dashwood, R.H.; Ho, E. Sulforaphane inhibits histone deacetylase activity in BPH-1, LnCaP and PC-3 prostate epithelial cells. Carcinogenesis 2006, 27, 811–819. [Google Scholar] [CrossRef]
- Myzak, M.C.; Karplus, P.A.; Chung, F.L.; Dashwood, R.H. A novel mechanism of chemoprotection by sulforaphane: Inhibition of histone deacetylase. Cancer Res. 2004, 64, 5767–5774. [Google Scholar] [CrossRef]
- Desai, J.V.; Mitchell, A.P.; Andes, D.R. Fungal biofilms, drug resistance, and recurrent infection. Cold Spring Harb. Perspect. Med. 2014, 4, a019729. [Google Scholar] [CrossRef]
- Deva, A.K.; Adams, W.P., Jr.; Vickery, K. The role of bacterial biofilms in device-associated infection. Plast Reconstr. Surg. 2013, 132, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.R.; McGillicuddy, E.; Kaplan, L.J. Biofilm: Basic principles, pathophysiology, and implications for clinicians. Surg. Infect. 2014, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Holban, A.M.; Gestal, M.C.; Grumezescu, A.M. New molecular strategies for reducing implantable medical devices associated infections. Curr. Med. Chem. 2014, 21, 3375–3382. [Google Scholar] [CrossRef] [PubMed]
- Nicolle, L.E. Urinary catheter-associated infections. Infect. Dis. Clin. North Am. 2012, 26, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yu, Q.; Sun, H. Novel strategies for the prevention and treatment of biofilm related infections. Int. J. Mol. Sci. 2013, 14, 18488–18501. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 207–228. [Google Scholar] [PubMed]
- Otto, M. Staphylococcal infections: Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis pathogenesis. Methods Mol. Biol. 2014, 1106, 17–31. [Google Scholar]
- Agarwal, A.; Singh, K.P.; Jain, A. Medical significance and management of staphylococcal biofilm. FEMS Immunol. Med. Microbiol. 2010, 58, 147–160. [Google Scholar] [CrossRef]
- Kleinschmidt, S.; Huygens, F.; Faoagali, J.; Rathnayake, I.U.; Hafner, L.M. Staphylococcus epidermidis as a cause of bacteremia. Future Microbiol. 2015, 10, 1859–1879. [Google Scholar] [CrossRef]
- Shah, S.; Tatara, A.M.; D’Souza, R.N.; Mikos, A.G.; Kurtis, F.K. Evolving strategies for preventing biofilm on implantable materials. Mater. Today 2013, 16, 177–182. [Google Scholar] [CrossRef]
- Sulemankhil, I.; Ganopolsky, J.G.; Dieni, C.A.; Dan, A.F.; Jones, M.L.; Prakash, S. Prevention and treatment of virulent bacterial biofilms with an enzymatic nitric oxide-releasing dressing. Antimicrob. Agents Chemother. 2012, 56, 6095–6103. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.J.; Mutters, N.T.; Blessing, B.; Gunther, F. Natural isothiocyanates express antimicrobial activity against developing and mature biofilms of Pseudomonas aeruginosa. Fitoterapia 2017, 119, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Lara-Lledo, M.; Olaimat, A.; Holley, R.A. Inhibition of Listeria monocytogenes on bologna sausages by an antimicrobial film containing mustard extract or sinigrin. Int. J. Food Microbiol. 2012, 156, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.; Dwivedi, A.; du Plessis, J. Sinigrin and Its Therapeutic Benefits. Molecules 2016, 21, 416. [Google Scholar] [CrossRef] [PubMed]
- Feskanich, D.; Ziegler, R.G.; Michaud, D.S.; Giovannucci, E.L.; Speizer, F.E.; Willett, W.C.; Colditz, G.A. Prospective study of fruit and vegetable consumption and risk of lung cancer among men and women. J. Natl. Cancer Inst. 2000, 92, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.A.; Moysich, K.B.; Freudenheim, J.L.; Shields, P.G.; Bowman, E.D.; Zhang, Y.; Marshall, J.R.; Ambrosone, C.B. Cruciferous vegetables, genetic polymorphisms in glutathione S-transferases M1 and T1, and prostate cancer risk. Nutr. Cancer 2004, 50, 206–213. [Google Scholar] [CrossRef]
- Neuhouser, M.L.; Patterson, R.E.; Thornquist, M.D.; Omenn, G.S.; King, I.B.; Goodman, G.E. Fruits and vegetables are associated with lower lung cancer risk only in the placebo arm of the beta-carotene and retinol efficacy trial (CARET). Cancer Epidemiol. Biomarkers Prev. 2003, 12, 350–358. [Google Scholar]
- Verhoeven, D.T.; Goldbohm, R.A.; van Poppel, G.; Verhagen, H.; van den Brandt, P.A. Epidemiological studies on brassica vegetables and cancer risk. Cancer Epidemiol. Biomarkers Prev. 1996, 5, 733–748. [Google Scholar]
- Voorrips, L.E.; Goldbohm, R.A.; van Poppel, G.; Sturmans, F.; Hermus, R.J.; van den Brandt, P.A. Vegetable and fruit consumption and risks of colon and rectal cancer in a prospective cohort study: The Netherlands Cohort Study on Diet and Cancer. Am. J. Epidemiol. 2000, 152, 1081–1092. [Google Scholar] [CrossRef]
- Smith, G.A.; Lin, T.H.; Sheehan, A.E.; Van der Goes van Naters, W.; Neukomm, L.J.; Graves, H.K.; Bis-Brewer, D.M.; Zuchner, S.; Freeman, M.R. Glutathione S-Transferase Regulates Mitochondrial Populations in Axons through Increased Glutathione Oxidation. Neuron 2019. [Google Scholar] [CrossRef] [PubMed]
- Agbas, A.; Krishnamurthy, P.; Michaelis, M.L.; Michaelis, E.K. Mitochondrial Electron Transfer Cascade Enzyme Activity Assessment in Cultured Neurons and Select Brain Regions. Curr. Protoc. Toxicol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Gill, R.; Mailloux, R.J. Protein S-glutathionylation: The linchpin for the transmission of regulatory information on redox buffering capacity in mitochondria. Chem. Biol. Interact. 2019, 299, 151–162. [Google Scholar] [CrossRef]
- Krug, P.; Mielczarek, L.; Wiktorska, K.; Kaczynska, K.; Wojciechowski, P.; Andrzejewski, K.; Ofiara, K.; Szterk, A.; Mazur, M. Sulforaphane-conjugated selenium nanoparticles: Towards a synergistic anticancer effect. Nanotechnology 2019, 30, 065101. [Google Scholar] [CrossRef] [PubMed]
- Kan, S.F.; Wang, J.; Sun, G.X. Sulforaphane regulates apoptosis- and proliferationrelated signaling pathways and synergizes with cisplatin to suppress human ovarian cancer. Int. J. Mol. Med. 2018, 42, 2447–2458. [Google Scholar] [PubMed]
- Rakariyatham, K.; Wu, X.; Tang, Z.; Han, Y.; Wang, Q.; Xiao, H. Synergism between luteolin and sulforaphane in anti-inflammation. Food Funct. 2018, 9, 5115–5123. [Google Scholar] [CrossRef] [PubMed]
- Lubecka, K.; Kaufman-Szymczyk, A.; Fabianowska-Majewska, K. Inhibition of breast cancer cell growth by the combination of clofarabine and sulforaphane involves epigenetically mediated CDKN2A upregulation. Nucleosides Nucleotides Nucleic Acids 2018, 37, 280–289. [Google Scholar] [CrossRef]
- Bose, C.; Awasthi, S.; Sharma, R.; Benes, H.; Hauer-Jensen, M.; Boerma, M.; Singh, S.P. Sulforaphane potentiates anticancer effects of doxorubicin and attenuates its cardiotoxicity in a breast cancer model. PLoS ONE 2018, 13, e0193918. [Google Scholar] [CrossRef]
- Chirumbolo, S.; Bjorklund, G. Sulforaphane and 5-fluorouracil synergistically inducing autophagy in breast cancer: A possible role for the Nrf2-Keap1-ARE signaling? Food Chem. Toxicol. 2018, 112, 414–415. [Google Scholar] [CrossRef]
- Schillheim, B.; Jansen, I.; Baum, S.; Beesley, A.; Bolm, C.; Conrath, U. Sulforaphane Modifies Histone H3, Unpacks Chromatin, and Primes Defense. Plant Physiol. 2018, 176, 2395–2405. [Google Scholar] [CrossRef]
- Royston, K.J.; Udayakumar, N.; Lewis, K.; Tollefsbol, T.O. A Novel Combination of Withaferin A and Sulforaphane Inhibits Epigenetic Machinery, Cellular Viability and Induces Apoptosis of Breast Cancer Cells. Int. J. Mol. Sci. 2017, 18, 1092. [Google Scholar] [CrossRef] [PubMed]
- Chaiprasongsuk, A.; Lohakul, J.; Soontrapa, K.; Sampattavanich, S.; Akarasereenont, P.; Panich, U. Activation of Nrf2 Reduces UVA-Mediated MMP-1 Upregulation via MAPK/AP-1 Signaling Cascades: The Photoprotective Effects of Sulforaphane and Hispidulin. J. Pharmacol. Exp. Ther. 2017, 360, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Rhee, Y.H.; Ahn, J.C. Sulforaphene-Carboplatin Combination Synergistically Enhances Apoptosis by Disruption of Mitochondrial Membrane Potential and Cell Cycle Arrest in Human Non-Small Cell Lung Carcinoma. J. Med. Food 2016, 19, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tao, C.; Yu, Y.; Yu, F.; Zhang, H.; Gao, J.; Wang, D.; Chen, Y.; Zhang, G.; Zhou, G.; et al. Simultaneous Targeting of Differentiated Breast Cancer Cells and Breast Cancer Stem Cells by Combination of Docetaxel- and Sulforaphane-Loaded Self-Assembled Poly(D, L-lactide-co-glycolide)/Hyaluronic Acid Block Copolymer-Based Nanoparticles. J. Biomed. Nanotechnol. 2016, 12, 1463–1477. [Google Scholar] [CrossRef] [PubMed]
- Kaczynska, A.; Herman-Antosiewicz, A. Combination of lapatinib with isothiocyanates overcomes drug resistance and inhibits migration of HER2 positive breast cancer cells. Breast Cancer 2017, 24, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Erzinger, M.M.; Bovet, C.; Hecht, K.M.; Senger, S.; Winiker, P.; Sobotzki, N.; Cristea, S.; Beerenwinkel, N.; Shay, J.W.; Marra, G.; et al. Sulforaphane Preconditioning Sensitizes Human Colon Cancer Cells towards the Bioreductive Anticancer Prodrug PR-104A. PLoS ONE 2016, 11, e0150219. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xiong, B.; Zhang, B.; Li, S.; Huang, N.; Zhan, G.; Jiang, R.; Yang, L.; Wu, Y.; Miao, L.; et al. Sulforaphane Alleviates Lipopolysaccharide-induced Spatial Learning and Memory Dysfunction in Mice: The Role of BDNF-mTOR Signaling Pathway. Neuroscience 2018, 388, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Micheli, L.; Trallori, E.; Citi, V.; Martelli, A.; Testai, L.; De Nicola, G.R.; Iori, R.; Calderone, V.; Ghelardini, C.; et al. Effect of glucoraphanin and sulforaphane against chemotherapy-induced neuropathic pain: Kv7 potassium channels modulation by H2 S release in vivo. Phytother. Res. 2018, 32, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.M.H.; Amin, A.; Prastowo, S.; Linares-Otoya, L.; Hoelker, M.; Schellander, K.; Tesfaye, D. Correction to: Sulforaphane protects granulosa cells against oxidative stress via activation of NRF2-ARE pathway. Cell Tissue Res. 2018, 374, 679–685. [Google Scholar] [CrossRef]
- Wang, Y.; Mandal, A.K.; Son, Y.O.; Pratheeshkumar, P.; Wise, J.T.F.; Wang, L.; Zhang, Z.; Shi, X.; Chen, Z. Roles of ROS, Nrf2, and autophagy in cadmium-carcinogenesis and its prevention by sulforaphane. Toxicol. Appl. Pharmacol. 2018, 353, 23–30. [Google Scholar] [CrossRef]
- Corssac, G.B.; Campos-Carraro, C.; Hickmann, A.; da Rosa Araujo, A.S.; Fernandes, R.O.; Bello-Klein, A. Sulforaphane effects on oxidative stress parameters in culture of adult cardiomyocytes. Biomed. Pharmacother. 2018, 104, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Jaafaru, M.S.; Abd Karim, N.A.; Enas, M.E.; Rollin, P.; Mazzon, E.; Abdull Razis, A.F. Protective Effect of Glucosinolates Hydrolytic Products in Neurodegenerative Diseases (NDDs). Nutrients 2018, 10, 580. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, B.R.; Kim, J.; LaFerla, F.M.; Park, J.H.Y.; Han, J.S.; Lee, K.W. Sulforaphane Upregulates the Heat Shock Protein Co-Chaperone CHIP and Clears Amyloid-beta and Tau in a Mouse Model of Alzheimer’s Disease. Mol. Nutr. Food Res. 2018, 62, e1800240. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, P.E.; Abdelkader, N.F.; El Awdan, S.A.; El-Shabrawy, O.A.; Zaki, H.F. Extracellular Matrix Remodeling and Modulation of Inflammation and Oxidative Stress by Sulforaphane in Experimental Diabetic Peripheral Neuropathy. Inflammation 2018, 41, 1460–1476. [Google Scholar] [CrossRef] [PubMed]
- Silva Rodrigues, J.F.; Silva, E.S.C.; Franca Muniz, T.; de Aquino, A.F.; Neuza da Silva Nina, L.; Fialho Sousa, N.C.; Nascimento da Silva, L.C.; de Souza, B.; da Penha, T.A.; Abreu-Silva, A.L.; et al. Sulforaphane Modulates Joint Inflammation in a Murine Model of Complete Freund’s Adjuvant-Induced Mono-Arthritis. Molecules 2018, 23, 988. [Google Scholar] [CrossRef] [PubMed]
- Pu, D.; Zhao, Y.; Chen, J.; Sun, Y.; Lv, A.; Zhu, S.; Luo, C.; Zhao, K.; Xiao, Q. Protective Effects of Sulforaphane on Cognitive Impairments and AD-like Lesions in Diabetic Mice are Associated with the Upregulation of Nrf2 Transcription Activity. Neuroscience 2018, 381, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Jhang, K.A.; Park, J.S.; Kim, H.S.; Chong, Y.H. Sulforaphane rescues amyloid-beta peptide-mediated decrease in MerTK expression through its anti-inflammatory effect in human THP-1 macrophages. J. Neuroinflammation 2018, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhu, D.; Chen, D.; Zhang, Q.; Dong, H.; Wu, W.; Lu, H.; Wu, G. Sulforaphane, a Natural Isothiocyanate Compound, Improves Cardiac Function and Remodeling by Inhibiting Oxidative Stress and Inflammation in a Rabbit Model of Chronic Heart Failure. Med. Sci. Monit. 2018, 24, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Kee, H.J.; Jin, L.; Ryu, Y.; Sun, S.; Kim, G.R.; Jeong, M.H. Inhibition of class IIa histone deacetylase activity by gallic acid, sulforaphane, TMP269, and panobinostat. Biomed. Pharmacother. 2018, 101, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, G.; Tian, H.; Hu, Y.; Wu, S.; Geng, Y.; Lin, K.; Wu, W. Sulforaphane metabolites cause apoptosis via microtubule disruption in cancer. Endocr. Relat. Cancer 2018, 25, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Eren, E.; Tufekci, K.U.; Isci, K.B.; Tastan, B.; Genc, K.; Genc, S. Sulforaphane Inhibits Lipopolysaccharide-Induced Inflammation, Cytotoxicity, Oxidative Stress, and miR-155 Expression and Switches to Mox Phenotype through Activating Extracellular Signal-Regulated Kinase 1/2-Nuclear Factor Erythroid 2-Related Factor 2/Antioxidant Response Element Pathway in Murine Microglial Cells. Front. Immunol. 2018, 9, 36. [Google Scholar] [PubMed]
- Qin, S.; Yang, C.; Huang, W.; Du, S.; Mai, H.; Xiao, J.; Lu, T. Sulforaphane attenuates microglia-mediated neuronal necroptosis through down-regulation of MAPK/NF-kappaB signaling pathways in LPS-activated BV-2 microglia. Pharmacol. Res. 2018, 133, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Dulull, N.K.; Dias, D.A.; Thrimawithana, T.R.; Kwa, F.A.A. L-Sulforaphane Confers Protection Against Oxidative Stress in an In Vitro Model of Age-Related Macular Degeneration. Curr Mol Pharmacol 2018, 11, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Dokumacioglu, E.; Iskender, H.; Aktas, M.S.; Hanedan, B.; Dokumacioglu, A.; Sen, T.M.; Musmul, A. The effect of sulforaphane on oxidative stress and inflammation in rats with toxic hepatitis induced by acetaminophene. Bratisl Lek Listy 2017, 118, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, X.; Yin, Y.; Sun, H.; Ge, H.; Li, W. Effects of sulforaphane and vitamin E on cognitive disorder and oxidative damage in lead-exposed mice hippocampus at lactation. J. Trace Elem. Med. Biol. 2017, 44, 88–92. [Google Scholar] [CrossRef]
- Son, Y.H.; Jang, E.J.; Kim, Y.W.; Lee, J.H. Sulforaphane prevents dexamethasone-induced muscle atrophy via regulation of the Akt/Foxo1 axis in C2C12 myotubes. Biomed. Pharmacother. 2017, 95, 1486–1492. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Yuan, H.; Lin, C. Sulforaphane induces p53deficient SW480 cell apoptosis via the ROSMAPK signaling pathway. Mol. Med. Rep. 2017, 16, 7796–7804. [Google Scholar] [CrossRef]
- Dacosta, C.; Bao, Y. The Role of MicroRNAs in the Chemopreventive Activity of Sulforaphane from Cruciferous Vegetables. Nutrients 2017, 9, 902. [Google Scholar] [CrossRef]
- Psurski, M.; Janczewski, L.; Switalska, M.; Gajda, A.; Goszczynski, T.M.; Oleksyszyn, J.; Wietrzyk, J.; Gajda, T. Novel phosphonate analogs of sulforaphane: Synthesis, in vitro and in vivo anticancer activity. Eur. J. Med. Chem. 2017, 132, 63–80. [Google Scholar] [CrossRef]
- Yanaka, A. Role of Sulforaphane in Protection of Gastrointestinal Tract Against H. pylori and NSAID-Induced Oxidative Stress. Curr. Pharm. Des. 2017, 23, 4066–4075. [Google Scholar] [CrossRef]
- Nazmy, E.A.; El-Khouly, O.A.; Atef, H.; Said, E. Sulforaphane protects against sodium valproate-induced acute liver injury. Can. J. Physiol. Pharmacol. 2017, 95, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Bi, M.; Li, Q.; Guo, D.; Ding, X.; Bi, W.; Zhang, Y.; Zou, Y. Sulphoraphane Improves Neuronal Mitochondrial Function in Brain Tissue in Acute Carbon Monoxide Poisoning Rats. Basic Clin. Pharmacol. Toxicol. 2017, 120, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Li, W.; Liu, Y.; Xiong, Z.; Zhang, X.; Zhou, S.; Palko, O.; Chen, H.; Kapita, M.; Prigge, J.R.; et al. Chemoprevention of oxidative stress-associated oral carcinogenesis by sulforaphane depends on NRF2 and the isothiocyanate moiety. Oncotarget 2016, 7, 53502–53514. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xu, H.; Fu, S.; Chen, Y.; Chen, Q.; Cai, Z.; Zhou, J.; Wang, Z. Sulforaphane Ameliorates Bladder Dysfunction through Activation of the Nrf2-ARE Pathway in a Rat Model of Partial Bladder Outlet Obstruction. Oxid. Med. Cell Longev. 2016, 2016, 7598294. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Bai, Y.; Sun, W.; Cui, W.; Xin, Y.; Wang, Y.; Tan, Y.; Miao, L.; Fu, Y.; Su, G.; et al. Sulforaphane prevention of diabetes-induced aortic damage was associated with the up-regulation of Nrf2 and its down-stream antioxidants. Nutr. Metab. 2012, 9, 84. [Google Scholar] [CrossRef]
- An, Y.W.; Jhang, K.A.; Woo, S.Y.; Kang, J.L.; Chong, Y.H. Sulforaphane exerts its anti-inflammatory effect against amyloid-beta peptide via STAT-1 dephosphorylation and activation of Nrf2/HO-1 cascade in human THP-1 macrophages. Neurobiol. Aging. 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Koolivand, M.; Ansari, M.; Piroozian, F.; Moein, S.; MalekZadeh, K. Alleviating the progression of acute myeloid leukemia (AML) by sulforaphane through controlling miR-155 levels. Mol. Biol. Rep. 2018, 45, 2491–2499. [Google Scholar] [CrossRef]
- Misiewicz, I.; Skupinska, K.; Kasprzycka-Guttman, T. Sulforaphane and 2-oxohexyl isothiocyanate induce cell growth arrest and apoptosis in L-1210 leukemia and ME-18 melanoma cells. Oncol. Rep. 2003, 10, 2045–2050. [Google Scholar] [CrossRef]
- Prata, C.; Facchini, C.; Leoncini, E.; Lenzi, M.; Maraldi, T.; Angeloni, C.; Zambonin, L.; Hrelia, S.; Fiorentini, D. Sulforaphane Modulates AQP8-Linked Redox Signalling in Leukemia Cells. Oxid. Med. Cell Longev. 2018, 2018, 4125297. [Google Scholar] [CrossRef]
- Shang, H.S.; Shih, Y.L.; Lee, C.H.; Hsueh, S.C.; Liu, J.Y.; Liao, N.C.; Chen, Y.L.; Huang, Y.P.; Lu, H.F.; Chung, J.G. Sulforaphane-induced apoptosis in human leukemia HL-60 cells through extrinsic and intrinsic signal pathways and altering associated genes expression assayed by cDNA microarray. Environ. Toxicol. 2017, 32, 311–328. [Google Scholar] [CrossRef]
- Shih, Y.L.; Wu, L.Y.; Lee, C.H.; Chen, Y.L.; Hsueh, S.C.; Lu, H.F.; Liao, N.C.; Chung, J.G. Sulforaphane promotes immune responses in a WEHI3induced leukemia mouse model through enhanced phagocytosis of macrophages and natural killer cell activities in vivo. Mol. Med. Rep. 2016, 13, 4023–4029. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Chen, F.; Liu, A.; Sun, D.; Wu, J.; Kong, F.; Luan, Y.; Qu, X.; Wang, R. Reversal of the multidrug resistance of human ileocecal adenocarcinoma cells by acetyl-11-keto-beta-boswellic acid via downregulation of P-glycoprotein signals. Biosci. Trends 2016, 10, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Dogan Sigva, Z.O.; Balci Okcanoglu, T.; Biray Avci, C.; Yilmaz Susluer, S.; Kayabasi, C.; Turna, B.; Dodurga, Y.; Nazli, O.; Gunduz, C. Investigation of the synergistic effects of paclitaxel and herbal substances and endemic plant extracts on cell cycle and apoptosis signal pathways in prostate cancer cell lines. Gene 2019, 687, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Kim, S.H.; Hahm, E.R.; Pore, S.K.; Jacobs, B.L.; Singh, S.V. Prostate cancer chemoprevention by sulforaphane in a preclinical mouse model is associated with inhibition of fatty acid metabolism. Carcinogenesis 2018, 39, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A.R.; Moura, M.B.; Hahm, E.R.; Singh, K.B.; Singh, S.V. Sulforaphane Inhibits c-Myc-Mediated Prostate Cancer Stem-Like Traits. J. Cell Biochem. 2016, 117, 2482–2495. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Tsai, S.H.; Wu, C.C. The chemopreventive isothiocyanate sulforaphane reduces anoikis resistance and anchorage-independent growth in non-small cell human lung cancer cells. Toxicol. Appl. Pharmacol. 2019, 362, 116–124. [Google Scholar] [CrossRef]
- Zuryn, A.; Litwiniec, A.; Safiejko-Mroczka, B.; Klimaszewska-Wisniewska, A.; Gagat, M.; Krajewski, A.; Gackowska, L.; Grzanka, D. The effect of sulforaphane on the cell cycle, apoptosis and expression of cyclin D1 and p21 in the A549 non-small cell lung cancer cell line. Int. J. Oncol. 2016, 48, 2521–2533. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Tan, K.N.; Gotteland, M.; Borges, K. Sulforaphane Protects against High Cholesterol-Induced Mitochondrial Bioenergetics Impairments, Inflammation, and Oxidative Stress and Preserves Pancreatic beta-Cells Function. Oxid. Med. Cell Longev. 2017, 2017, 3839756. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Z.; Zhou, C.; Chen, K.; Li, X.; Wang, Z.; Wu, Z.; Ma, J.; Ma, Q.; Duan, W. Activation of Nrf2 by Sulforaphane Inhibits High Glucose-Induced Progression of Pancreatic Cancer via AMPK Dependent Signaling. Cell Physiol. Biochem. 2018, 50, 1201–1215. [Google Scholar] [CrossRef]
- Dong, Z.; Shang, H.; Chen, Y.Q.; Pan, L.L.; Bhatia, M.; Sun, J. Sulforaphane Protects Pancreatic Acinar Cell Injury by Modulating Nrf2-Mediated Oxidative Stress and NLRP3 Inflammatory Pathway. Oxid. Med. Cell Longev. 2016, 2016, 7864150. [Google Scholar] [CrossRef]
- Park, Y.K.; Ramalingam, M.; Kim, S.; Jang, B.C.; Park, J.W. Sulforaphane inhibits the interferon-gamma-induced expression of MIG, IP-10 and I-TAC in INS1 pancreatic beta-cells through the downregulation of IRF-1, STAT-1 and PKB. Int. J. Mol. Med. 2017, 40, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Wu, H.; Vasilatos, S.N.; Chandran, U.; Qin, Y.; Wan, Y.; Oesterreich, S.; Davidson, N.E.; Huang, Y. HDAC5-LSD1 axis regulates antineoplastic effect of natural HDAC inhibitor sulforaphane in human breast cancer cells. Int. J. Cancer 2018, 143, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Danafar, H.; Sharafi, A.; Kheiri Manjili, H.; Andalib, S. Sulforaphane delivery using mPEG-PCL co-polymer nanoparticles to breast cancer cells. Pharm. Dev. Technol. 2017, 22, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Gianfredi, V.; Nucci, D.; Vannini, S.; Villarini, M.; Moretti, M. In vitro Biological Effects of Sulforaphane (SFN), Epigallocatechin-3-gallate (EGCG), and Curcumin on Breast Cancer Cells: A Systematic Review of the Literature. Nutr. Cancer 2017, 69, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Gianfredi, V.; Vannini, S.; Moretti, M.; Villarini, M.; Bragazzi, N.L.; Izzotti, A.; Nucci, D. Sulforaphane and Epigallocatechin Gallate Restore Estrogen Receptor Expression by Modulating Epigenetic Events in the Breast Cancer Cell Line MDA-MB-231: A Systematic Review and Meta-Analysis. J. Nutrigenet. Nutrigenomics 2017, 10, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Jaman, M.S.; Sayeed, M.A. Ellagic acid, sulforaphane, and ursolic acid in the prevention and therapy of breast cancer: Current evidence and future perspectives. Breast Cancer 2018, 25, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.M.; Nazzal, S. Novel sulforaphane-enabled self-microemulsifying delivery systems (SFN-SMEDDS) of taxanes: Formulation development and in vitro cytotoxicity against breast cancer cells. Int. J. Pharm. 2018, 536, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Adamczyk-Grochala, J.; Deregowska, A.; Wnuk, M. Sulforaphane-Induced Cell Cycle Arrest and Senescence are accompanied by DNA Hypomethylation and Changes in microRNA Profile in Breast Cancer Cells. Theranostics 2017, 7, 3461–3477. [Google Scholar] [CrossRef]
- Yang, F.; Wang, F.; Liu, Y.; Wang, S.; Li, X.; Huang, Y.; Xia, Y.; Cao, C. Sulforaphane induces autophagy by inhibition of HDAC6-mediated PTEN activation in triple negative breast cancer cells. Life Sci. 2018, 213, 149–157. [Google Scholar] [CrossRef]
- Abbaoui, B.; Lucas, C.R.; Riedl, K.M.; Clinton, S.K.; Mortazavi, A. Cruciferous Vegetables, Isothiocyanates, and Bladder Cancer Prevention. Mol. Nutr. Food Res. 2018, 62, e1800079. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Li, Y.; Wade, K.L.; Paonessa, J.D.; Fahey, J.W.; Zhang, Y. Allyl isothiocyanate-rich mustard seed powder inhibits bladder cancer growth and muscle invasion. Carcinogenesis 2010, 31, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Huang, L.; Lei, P.; Liu, X.; Li, B.; Shan, Y. Sulforaphane Normalizes Intestinal Flora and Enhances Gut Barrier in Mice with BBN-Induced Bladder Cancer. Mol. Nutr. Food Res. 2018, 62, e1800427. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Y.; Molagoda, I.M.N.; Karunarathne, W.; Kang, S.H.; Park, C.; Kim, G.Y.; Choi, Y.H. TRAIL attenuates sulforaphane-mediated Nrf2 and sustains ROS generation, leading to apoptosis of TRAIL-resistant human bladder cancer cells. Toxicol. Appl. Pharmacol. 2018, 352, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Diorio, G.; Sexton, W.; Schell, M.; Alexandrow, M.; Fahey, J.W.; Kumar, N.B. Sulforaphane for the chemoprevention of bladder cancer: Molecular mechanism targeted approach. Oncotarget 2017, 8, 35412–35424. [Google Scholar] [CrossRef] [PubMed]
- Veeranki, O.L.; Bhattacharya, A.; Tang, L.; Marshall, J.R.; Zhang, Y. Cruciferous vegetables, isothiocyanates, and prevention of bladder cancer. Curr. Pharmacol. Rep. 2015, 1, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Kntayya, S.B.; Ibrahim, M.D.; Mohd Ain, N.; Iori, R.; Ioannides, C.; Abdull Razis, A.F. Induction of Apoptosis and Cytotoxicity by Isothiocyanate Sulforaphene in Human Hepatocarcinoma HepG2 Cells. Nutrients 2018, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, W.; Zhou, Z.; Smith, A.J.O.; Bowater, R.P.; Wormstone, I.M.; Chen, Y.; Bao, Y. Chemopreventive Activities of Sulforaphane and Its Metabolites in Human Hepatoma HepG2 Cells. Nutrients 2018, 10, 585. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yuan, L.; Wang, Y.; Chen, G.; Hu, K. Benzyl sulforaphane is superior to sulforaphane in inhibiting the Akt/MAPK and activating the Nrf2/ARE signalling pathways in HepG2 cells. J. Pharm. Pharmacol. 2018, 70, 1643–1653. [Google Scholar] [CrossRef]
- Ren, K.; Li, Z.; Li, Y.; Zhang, W.; Han, X. Sulforaphene enhances radiosensitivity of hepatocellular carcinoma through suppression of the NF-kappaB pathway. J. Biochem. Mol. Toxicol. 2017, 31, e21917. [Google Scholar] [CrossRef]
- Zou, X.; Qu, Z.; Fang, Y.; Shi, X.; Ji, Y. Endoplasmic reticulum stress mediates sulforaphane-induced apoptosis of HepG2 human hepatocellular carcinoma cells. Mol. Med. Rep. 2017, 15, 331–338. [Google Scholar] [CrossRef][Green Version]
- Choi, Y.H. ROS-mediated activation of AMPK plays a critical role in sulforaphane-induced apoptosis and mitotic arrest in AGS human gastric cancer cells. Gen Physiol. Biophys. 2018, 37, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Kiani, S.; Akhavan-Niaki, H.; Fattahi, S.; Kavoosian, S.; Babaian Jelodar, N.; Bagheri, N.; Najafi Zarrini, H. Purified sulforaphane from broccoli (Brassica oleracea var. italica) leads to alterations of CDX1 and CDX2 expression and changes in miR-9 and miR-326 levels in human gastric cancer cells. Gene 2018, 678, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Elkashty, O.A.; Ashry, R.; Elghanam, G.A.; Pham, H.M.; Su, X.; Stegen, C.; Tran, S.D. Broccoli extract improves chemotherapeutic drug efficacy against head-neck squamous cell carcinomas. Med. Oncol. 2018, 35, 124. [Google Scholar] [CrossRef] [PubMed]
- Saha, K.; Fisher, M.L.; Adhikary, G.; Grun, D.; Eckert, R.L. Sulforaphane suppresses PRMT5/MEP50 function in epidermal squamous cell carcinoma leading to reduced tumor formation. Carcinogenesis 2017, 38, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Z.; Li, M.; Liu, M.; Bahena, A.; Zhang, Y.; Nambiar, C.; Liu, G. Sulforaphane promotes apoptosis, and inhibits proliferation and self-renewal of nasopharyngeal cancer cells by targeting STAT signal through miRNA-124-3p. Biomed. Pharmacother. 2018, 103, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Arcidiacono, P.; Ragonese, F.; Stabile, A.; Pistilli, A.; Kuligina, E.; Rende, M.; Bottoni, U.; Calvieri, S.; Crisanti, A.; Spaccapelo, R. Antitumor activity and expression profiles of genes induced by sulforaphane in human melanoma cells. Eur. J. Nutr. 2018, 57, 2547–2569. [Google Scholar] [CrossRef]
- Kumar, R.; de Mooij, T.; Peterson, T.E.; Kaptzan, T.; Johnson, A.J.; Daniels, D.J.; Parney, I.F. Modulating glioma-mediated myeloid-derived suppressor cell development with sulforaphane. PLoS ONE 2017, 12, e0179012. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Yang, G.; Tian, H.; Geng, Y.; Hu, Y.; Lin, K.; Wu, W. Sulforaphane-cysteine induces apoptosis by sustained activation of ERK1/2 and caspase 3 in human glioblastoma U373MG and U87MG cells. Oncol. Rep. 2017, 37, 2829–2838. [Google Scholar] [CrossRef]
- Sita, G.; Hrelia, P.; Graziosi, A.; Morroni, F. Sulforaphane from Cruciferous Vegetables: Recent Advances to Improve Glioblastoma Treatment. Nutrients 2018, 10, 1755. [Google Scholar] [CrossRef]
- Liu, K.C.; Shih, T.Y.; Kuo, C.L.; Ma, Y.S.; Yang, J.L.; Wu, P.P.; Huang, Y.P.; Lai, K.C.; Chung, J.G. Sulforaphane Induces Cell Death Through G2/M Phase Arrest and Triggers Apoptosis in HCT 116 Human Colon Cancer Cells. Am. J. Chin. Med. 2016, 44, 1289–1310. [Google Scholar] [CrossRef]
- Martin, S.L.; Kala, R.; Tollefsbol, T.O. Mechanisms for the Inhibition of Colon Cancer Cells by Sulforaphane through Epigenetic Modulation of MicroRNA-21 and Human Telomerase Reverse Transcriptase (hTERT) Down-regulation. Curr. Cancer Drug Targets 2018, 18, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Rojo de la Vega, M.; Chapman, E.; Ooi, A.; Zhang, D.D. The effects of NRF2 modulation on the initiation and progression of chemically and genetically induced lung cancer. Mol. Carcinog. 2018, 57, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, S.; Chen, Y.; Li, X.; Jiang, Y.; Yang, X.; Li, Y.; Wang, X.; Meng, Y.; Zhu, M.; et al. miR-19 targeting of GSK3beta mediates sulforaphane suppression of lung cancer stem cells. J. Nutr. Biochem. 2017, 44, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Fujita, T.; Stankovic, K.M.; Welling, D.B.; Moon, I.S.; Choi, J.Y.; Yun, J.; Kang, J.S.; Lee, J.D. Sulforaphane, a natural component of broccoli, inhibits vestibular schwannoma growth in vitro and in vivo. Sci. Rep. 2016, 6, 36215. [Google Scholar] [CrossRef] [PubMed]
- Lubelska, K.; Wiktorska, K.; Mielczarek, L.; Milczarek, M.; Zbroinska-Bregisz, I.; Chilmonczyk, Z. Sulforaphane Regulates NFE2L2/Nrf2-Dependent Xenobiotic Metabolism Phase II and Phase III Enzymes Differently in Human Colorectal Cancer and Untransformed Epithelial Colon Cells. Nutr. Cancer 2016, 68, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.M.; Tsai, C.C.; Hsu, Y.C. Sulforaphane, a Dietary Isothiocyanate, Induces G(2)/M Arrest in Cervical Cancer Cells through CyclinB1 Downregulation and GADD45beta/CDC2 Association. Int. J. Mol. Sci. 2016, 17, 1530. [Google Scholar] [CrossRef] [PubMed]
- Bauman, J.E.; Zang, Y.; Sen, M.; Li, C.; Wang, L.; Egner, P.A.; Fahey, J.W.; Normolle, D.P.; Grandis, J.R.; Kensler, T.W.; et al. Prevention of Carcinogen-Induced Oral Cancer by Sulforaphane. Cancer Prev. Res. 2016, 9, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Sukumar, S. Big punches come in nanosizes for chemoprevention. Cancer Prev. Res. 2013, 6, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of Glucosinolates and Their Breakdown Products: Impact of Processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef]
- Okunade, O.; Niranjan, K.; Ghawi, S.K.; Kuhnle, G.; Methven, L. Supplementation of the Diet by Exogenous Myrosinase via Mustard Seeds to Increase the Bioavailability of Sulforaphane in Healthy Human Subjects after the Consumption of Cooked Broccoli. Mol. Nutr. Food Res. 2018, 62, e1700980. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. Glycosaminoglycan and Proteoglycan Biotherapeutics in Articular Cartilage Protection and Repair Strategies: Novel Approaches to Visco-supplementation in Orthobiologics. Adv. Ther. 2019. [Google Scholar] [CrossRef]
- Kim, J.; Bang, H.; Ahn, M.; Choi, Y.; Kim, G.O.; Shin, T. Allyl isothiocyanate reduces liver fibrosis by regulating Kupffer cell activation in rats. J. Vet. Med. Sci. 2018, 80, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Lee, C.G.; Rhee, D.K.; Um, S.H.; Pyo, S. Sinigrin inhibits production of inflammatory mediators by suppressing NF-kappaB/MAPK pathways or NLRP3 inflammasome activation in macrophages. Int. Immunopharmacol. 2017, 45, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.; Dwivedi, A.; du Preez, J.L.; du Plessis, J. In vitro wound healing and cytotoxic effects of sinigrin-phytosome complex. Int. J. Pharm. 2016, 498, 283–293. [Google Scholar] [CrossRef]
- Awasthi, S.; Saraswathi, N.T. Elucidating the molecular interaction of sinigrin, a potent anticancer glucosinolate from cruciferous vegetables with bovine serum albumin: Effect of methylglyoxal modification. J. Biomol. Struct. Dyn. 2016, 34, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Jie, M.; Cheung, W.M.; Yu, V.; Zhou, Y.; Tong, P.H.; Ho, J.W. Anti-proliferative activities of sinigrin on carcinogen-induced hepatotoxicity in rats. PLoS ONE 2014, 9, e110145. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Lee, H.J. Allyl isothiocyanate and its N-acetylcysteine conjugate suppress metastasis via inhibition of invasion, migration, and matrix metalloproteinase-2/-9 activities in SK-Hep 1 human hepatoma cells. Exp. Biol. Med. 2006, 231, 421–430. [Google Scholar] [CrossRef]
- Smith, T.K.; Lund, E.K.; Clarke, R.G.; Bennett, R.N.; Johnson, I.T. Effects of Brussels sprout juice on the cell cycle and adhesion of human colorectal carcinoma cells (HT29) in vitro. J. Agric. Food Chem. 2005, 53, 3895–3901. [Google Scholar] [CrossRef]
- Smith, T.K.; Lund, E.K.; Parker, M.L.; Clarke, R.G.; Johnson, I.T. Allyl-isothiocyanate causes mitotic block, loss of cell adhesion and disrupted cytoskeletal structure in HT29 cells. Carcinogenesis 2004, 25, 1409–1415. [Google Scholar] [CrossRef]
- Musk, S.R.; Smith, T.K.; Johnson, I.T. On the cytotoxicity and genotoxicity of allyl and phenethyl isothiocyanates and their parent glucosinolates sinigrin and gluconasturtiin. Mutat. Res. 1995, 348, 19–23. [Google Scholar] [CrossRef]
- Brabban, A.D.; Edwards, C. The effects of glucosinolates and their hydrolysis products on microbial growth. J. Appl. Bacteriol. 1995, 79, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A.; Wang, C.X.; Amin, S.G.; Hecht, S.S.; Chung, F.L. Effects of dietary sinigrin or indole-3-carbinol on O6-methylguanine-DNA-transmethylase activity and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced DNA methylation and tumorigenicity in F344 rats. Carcinogenesis 1988, 9, 1891–1895. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Types |
|---|
| Brocolli |
| Brocolli Sprouts |
| Cabbage |
| Brussell Sprouts |
| Cauliflower |
| Daikon (Japanese radish) |
| Daikon sprouts |
| Garden Cress (Lepidum sativum) |
| Kale |
| Rapeseed (Brassica napus) |
| Wasabi (Wasabia japonica) |
| White Mustard (Sinapis alba) |
| Yellow Mustard (Brassica juncea) |
| Bok Choi |
| Arugula, Rocket (Eruca sativa) |
| Collard Greens |
| Horseradish |
| Kohlrabi |
| Radish |
| Rutabaga/turnip |
| Watercress |
| Mustard Greens |
| Glucosinolate Trivial Name | Aglycone Chemical Name | Aglycone Structure | Vegetable Source | Glucosinolate Content μmol/100 g) |
|---|---|---|---|---|
| A. Aliphatic | ||||
| Glucoibervirin | 3-Methylthiopropyl |  | Green Cauliflower White Cauliflower | 0–11.8 1.5–7.1 |
| Glucoerucin | 4-Methylthiobutyl |  | Rocket | 52–109 |
| Glucoiberin | 3-Methylsulfinylbutyl |  | Brocolli Sprouts Savoy Cabbage | 59–181 24–50 |
| Glucoraphanin | 4-Methylsulfinylbutyl |  | Brocolli Sprouts Brocolli | 233–676 24–285 |
| Sinigrin | Prop-2-enyl |  | Brussels Sprouts White Cauliflower | 46–91 57–121 |
| Gluconapin | But-2-enyl |  | Pak Choi | 24–157 |
| Glucovrassicanapin | Pent-2-enyl |  | Chinese Cabbage Pak Choi | 2.3–25 27–69 |
| Progoitrin | (2R)-Hydroxybut-3-enyl |  | Turnip Chinese Brocolli | 18–41 49 |
| B. Indolic | ||||
| Glucobrassicin | Indol-3-ylmethyl |  | Brocolli White Cauliflower | 13–29 11–33 |
| 4-Hydroxy-Glucobrassicin | 4-Hydroxy-3-indolylmethyl |  | Brocolli White Cauliflower | 0.1–3.3 0.2–2.8 |
| 4-Methoxy–Glucobrassicin | 4-Methoxy-3-indolylmethyl |  | Brocolli White Cauliflower | 0.9–2.8 0.7–3.2 |
| Neo-Glucobrassicin | 4-Methoxyindol-3-ylmethyl |  | Brocolli White Cauliflower | 1.8–13 0.9–3.0 |
| C. Aromatic | ||||
| Glucotropaeolin | Benzyl |  | Garden Cress | NA |
| Gluconasturtiin | Penylethyl |  | Water Cress | NA |
| Priority Category | Bacterium | Drug Resistance |
|---|---|---|
| Critical | Acinetobacter baumannii | |
| Pseudomonas aeruginosa | Carbapanem | |
| ESBL** producing members of the Enterobacteriaceae | ||
| High | Enterococcus faecium | Vancomycin |
| Staphylococcus aureus | Methicillin/Vancomycin | |
| Helicobacter pylori | Clathrimycin | |
| Campylobacter spp | Fluoroquinolone | |
| Salmonellae | Fluoroquinolone | |
| Neisseria gonorrhoeae | Cepalosporin/Fluoriquinolone | |
| Medium | Streptococcus pneumonia | Penicillin |
| Haemophilus influenzae | Ampicillin | |
| Shigella spp | Fluoriquinolone |
| Compound Used in Combination Therapy | Reference |
|---|---|
| SFN-Selenium nanoparticles | [122] |
| Paclitaxel | [9] |
| Cisplatin | [123] |
| Luteolin | [124] |
| Clofarabine | [125] |
| Doxorubicin | [126] |
| 5-fluorouracil | [127] |
| HistoneH3 | [128] |
| Withaferin A | [129] |
| Hispidulin | [130] |
| Carboplatin | [131] |
| Docetaxel | [132] |
| Lapatinib | [133] |
| PR-104A | [134] |
| Medical Conditions Treated with Sulphoraphane | |
|---|---|
| Spatial learning and memory dysfunction | [135] |
| Chemotherapy-induced neuropathic pain | [136] |
| Protection of granulosa cells against oxidative stress | [137] |
| Cadmium-mediated carcinogenesis | [138] |
| Oxidative stress in cultured adult cardiomyocytes | [139] |
| Protective effects of glucosinolate hydrolysis products in neurodegenerative diseases | [140] |
| Clearance of Amyloid-β and Tau protein in a mouse model of AD | [141] |
| Experimental diabetic peripheral neuropathy | [142] |
| Joint inflammation in a murine adjuvant-induced mono-arthritis | [143] |
| Protection against cognitive impairment in AD-like lesions in diabetes | [144] |
| Anti-inflammatory effect of SFN on human THP-1 macrophages in a murine AD model | [145] |
| Inhibition of oxidative stress/inflammation improves cardiac function in a Rabbit Model of Chronic Heart Failure | [146] |
| Inhibition of class IIa histone deacetylase activity | [147] |
| Apoptosis via microtubule disruption in cancer | [148] |
| Inhibition of LPS-Induced Inflammation/cytotoxicity/oxidative microglial stress | [149] |
| Down-regulation of MAPK/NF-κB signaling in LPS-activated BV-2 microglia | [150] |
| Inhibition of oxidative stress in an in vitro model of age-related macular degeneration | [151] |
| Modification of Histone H3, unpacking of chromatin, to prime defence | [128] |
| Modulation of oxidative stress and inflammation in rats with toxic hepatitis | [152] |
| Modulation of oxidative damage in lead exposed rat hippocampus | [153] |
| Prevention of dexamethasone-induced myotube atrophy via Akt/Foxo1 | [154] |
| Induction of p53 deficient SW480 cell apoptosis by ROS MAPK signaling | [155] |
| Role of microRNAs in the chemopreventive activity of SFN | [156] |
| Novel phosphonate analogs of SFN with in vitro and in vivo anti-cancer activity | [157] |
| Gastrointestinal protection against H. pylori and NSAID-Induced Oxidative Stress | [158] |
| Protection against sodium valproate-induced acute liver injury | [159] |
| Enhanced SFN cardioprotection against oxidative stress by 17β-Estradiol | [159] |
| Photoprotective Effects of SFN and Hispidulin | [130] |
| Improvement of neuronal mitochondrial function in brain tissue | [160] |
| Chemoprevention of oxidative stress-associated with oral carcinogenesis | [161] |
| Amelioration of bladder dysfunction via activation of Nrf2-ARE Pathway | [162] |
| Protection against aortic complications in diabetes | [163] |
| Anti-inflammatory effect against amyloid-β peptide via STAT-1 dephosphorylation and activation of Nrf2/HO-1 | [164] |
| Cancer Type | Reference |
|---|---|
| Leukemia | [87,165,166,167,168,169,170] |
| Prostate cancer | [91,95,171,172,173] |
| non-small cell lung cancer cells | [131,174,175] |
| Pancreatic cancer | [176,177,178,179] |
| Breast cancer | [92,93,125,126,127,129,132,133,180,181,182,183,184,185,186,187] |
| Bladder cancer | [162,188,189,190,191,192,193] |
| Ovarian cancer | [123] |
| HepG2 Carcinoma Cells | [194,195,196,197,198] |
| Gastric cancer | [199,200] |
| Squamous cell carcinoma | [201,202] |
| Nasopharangeal cancer | [203] |
| Melanoma | [204] |
| Glioma | [163,205,206,207] |
| Colon cancer | [134,208,209] |
| Lung cancer | [210,211] |
| Schwannoma | [212] |
| Colorectal cancer | [213] |
| Cervical cancer | [214] |
| Oral cancer | [215,216] |
| Application | Reference |
|---|---|
| Reduction of liver fibrosis | [220] |
| Suppression of NF-κB/MAPK and NLRP3 inflammasome activation in macrophages | [221] |
| Promotion of wound healing | [113,222] |
| Anti-cancer properties in methyl glyoxal modification | [223] |
| Anti-proliferative activity on carcinogen-induced hepatotoxicity | [224] |
| Biofumigation of potato cyst nematode | [21] |
| Inhibition of Listeria monocytogenes on bologna sausages | [112] |
| inhibition of invasion, migration, MMP-2/-9 activities in SK-Hep 1 human hepatoma cells | [225] |
| Brussel sprout juice mediated effects on cell cycle and adhesion of human colorectal carcinoma cells (HT29) in vitro | [226] |
| AITC mediated mitotic block, loss of cell adhesion/disrupted cytoskeleton in HT29 cells | [227] |
| Cytotoxicity and genotoxicity of allyl and phenethyl isothiocyanates, glucosinolates, sinigrin and gluconasturtiin | [228] |
| Inhibition of microbial growth | [47,65,229] |
| Effects of dietary sinigrin or indole-3-carbinol on O6-methylguanine-DNA-transmethylase activity and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced DNA methylation and tumorigenicity in F344 rats | [230] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melrose, J. The Glucosinolates: A Sulphur Glucoside Family of Mustard Anti-Tumour and Antimicrobial Phytochemicals of Potential Therapeutic Application. Biomedicines 2019, 7, 62. https://doi.org/10.3390/biomedicines7030062
Melrose J. The Glucosinolates: A Sulphur Glucoside Family of Mustard Anti-Tumour and Antimicrobial Phytochemicals of Potential Therapeutic Application. Biomedicines. 2019; 7(3):62. https://doi.org/10.3390/biomedicines7030062
Chicago/Turabian StyleMelrose, James. 2019. "The Glucosinolates: A Sulphur Glucoside Family of Mustard Anti-Tumour and Antimicrobial Phytochemicals of Potential Therapeutic Application" Biomedicines 7, no. 3: 62. https://doi.org/10.3390/biomedicines7030062
APA StyleMelrose, J. (2019). The Glucosinolates: A Sulphur Glucoside Family of Mustard Anti-Tumour and Antimicrobial Phytochemicals of Potential Therapeutic Application. Biomedicines, 7(3), 62. https://doi.org/10.3390/biomedicines7030062