
A Reassessment of the Therapeutic Potential of a Dopamine Receptor 2 Agonist (D2-AG) in Endometriosis by Comparison against a Standardized Antiangiogenic Treatment

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Heterologous Mouse Model of Endometriosis
2.2.1. Preparation of Endometriosis Recipient Mice
2.2.2. Obtention and Preprocessing of Human Endometrial Biopsies
2.2.3. Labeling of Human Endometrial Preprocessed Biopsies
2.2.4. Implantation of Labeled Endometrial Fragments in Recipient Mice
2.3. Pharmacologic Interventions during the Time Course
2.4. Non-Invasive Assessment of Lesion Size
2.4.1. Obtention of Raw Fluorescent Signal
2.4.2. Quantification of Lesion Fluorescent Signal
2.5. Immunofluorescence and Immunohistochemistry
Quantification of Immunofluorescent/Immunohistochemical Images
2.6. Statistical Analysis
3. Results
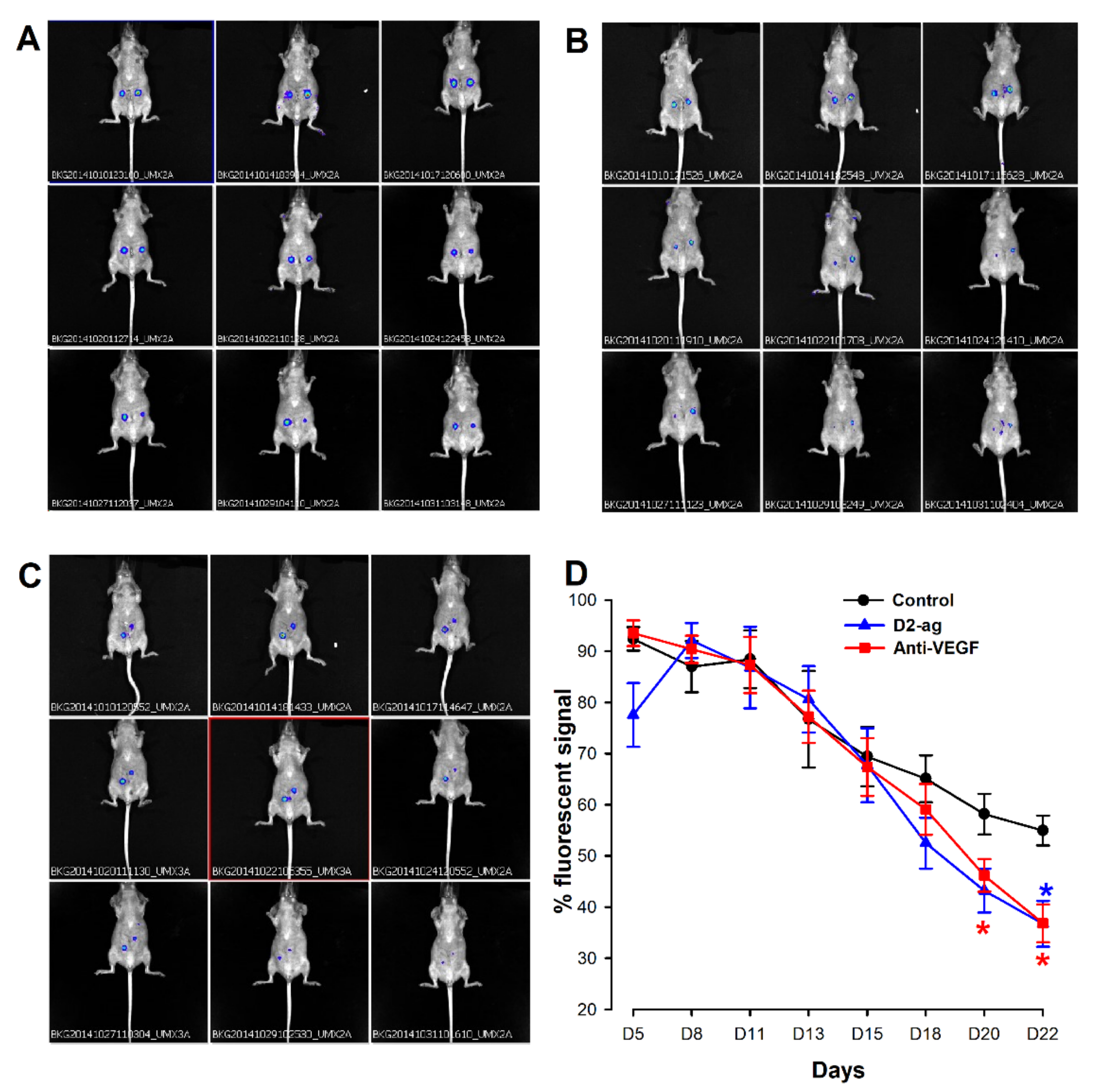
3.1. Assessment of Endometriotic Lesions by FLI
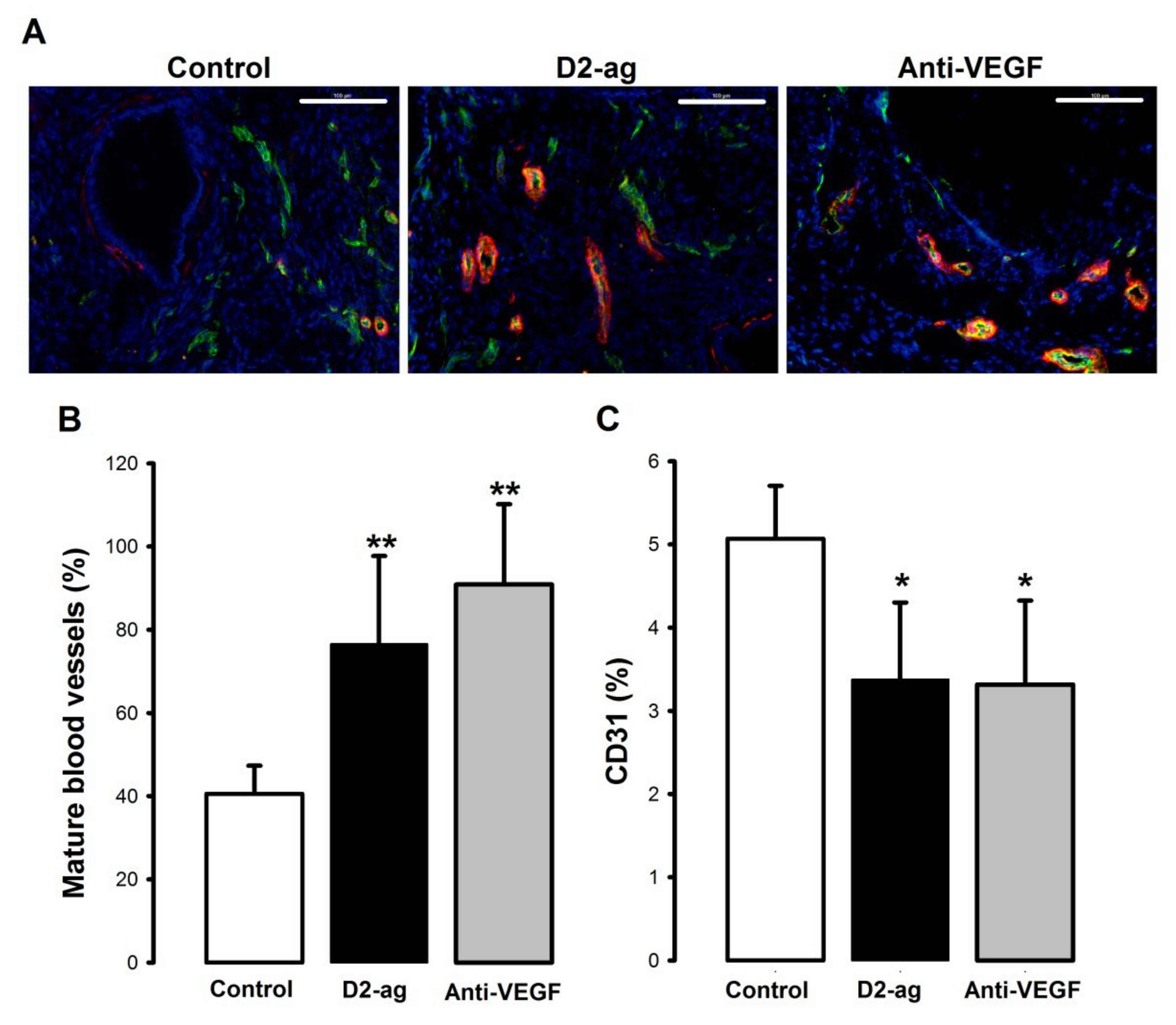
3.2. Vascular Density and Maturity in Endometriotic Lesions
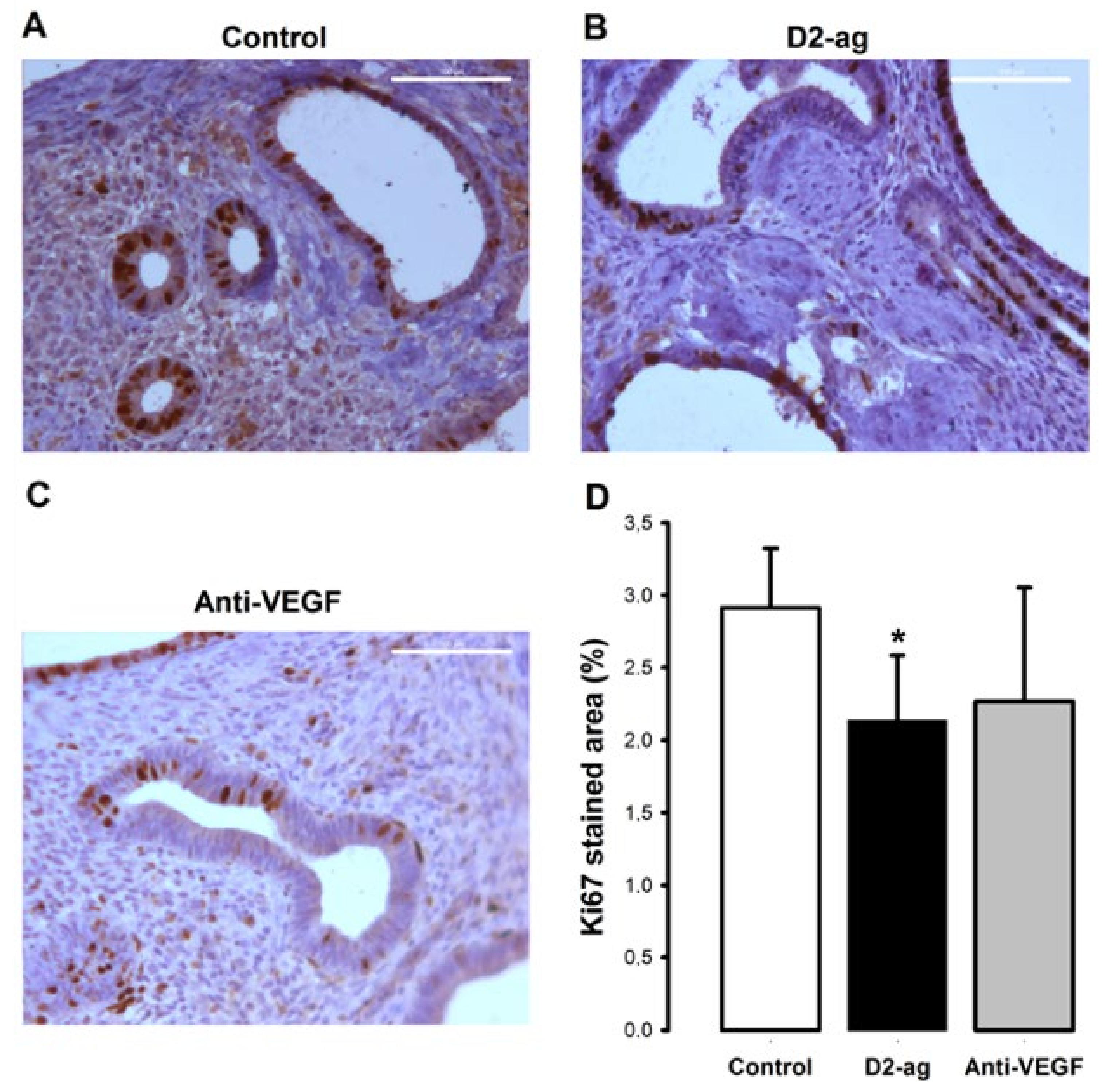
3.3. Proliferation
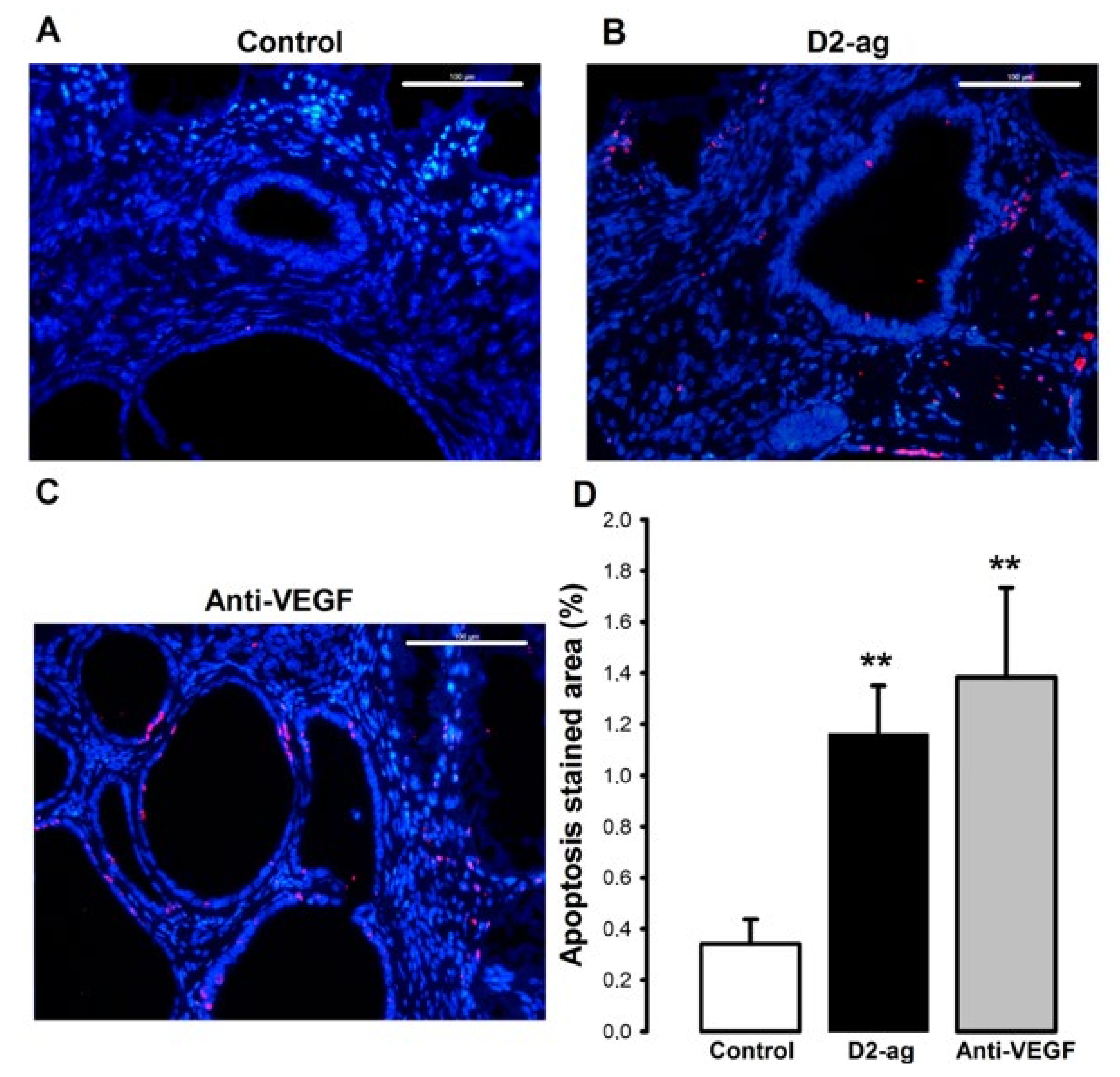
3.4. Apoptosis
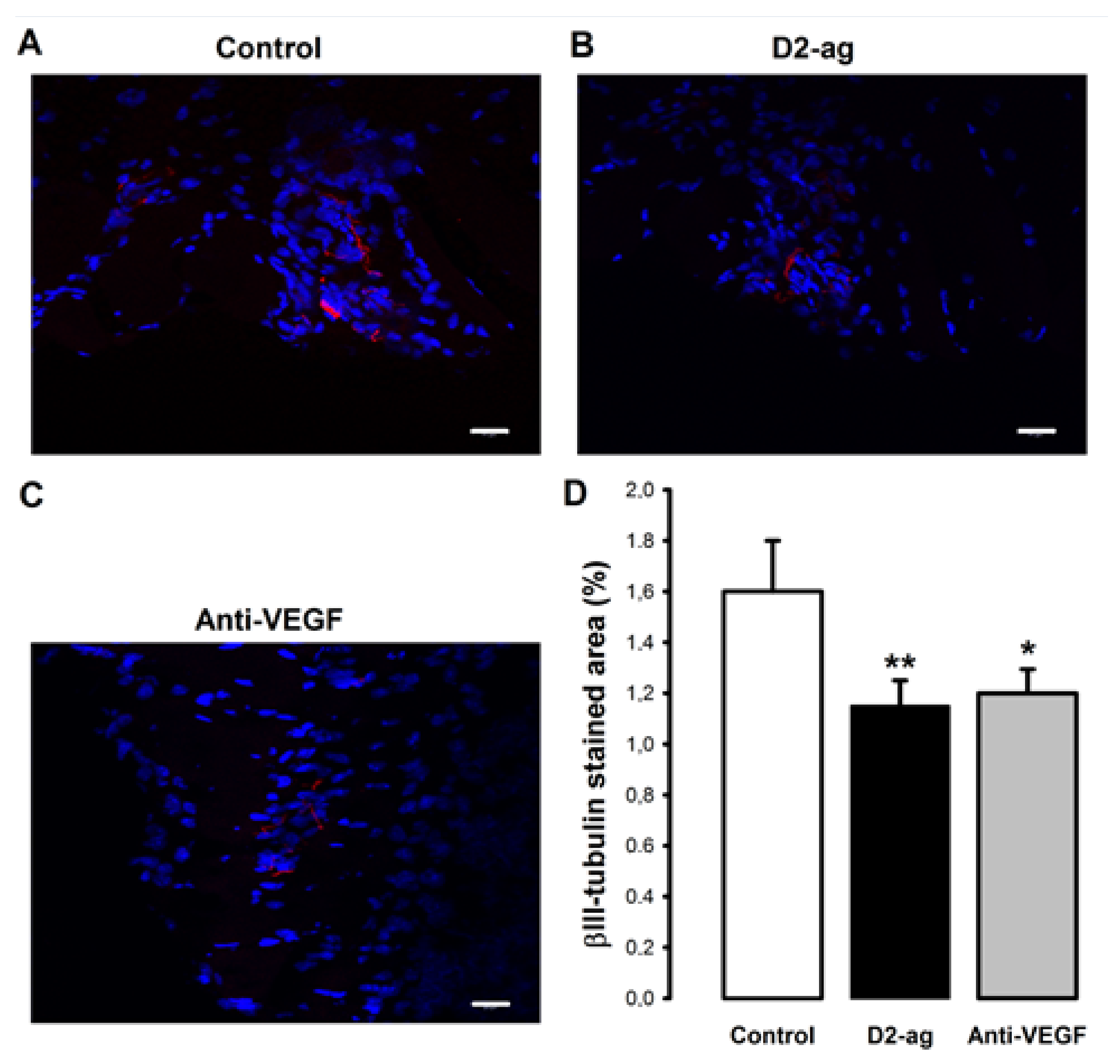
3.5. Nerve Fiber Density
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrero, S.; Evangelisti, G.; Barra, F. Current and emerging treatment options for endometriosis. Expert Opin. Pharmacother. 2018, 19, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Carlyle, D.; Khader, T.; Lam, D.; Vadivelu, N.; Shiwlochan, D.; Yonghee, C. Endometriosis Pain Management: A Review. Curr. Pain Headache Rep. 2020, 24, 1–9. [Google Scholar] [CrossRef]
- Zheng, W.; Cao, L.; Xu, Z.; Ma, Y.; Liang, X. Anti-Angiogenic Alternative and Complementary Medicines for the Treatment of Endometriosis: A Review of Potential Molecular Mechanisms. Evidence-Based Complement. Altern. Med. 2018, 2018, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.-P. The Role of Vascular Endothelial Growth Factor in Angiogenesis. Acta Haematol. 2001, 106, 148–156. [Google Scholar] [CrossRef]
- Gargett, C.E.; Rogers, P.A. Human endometrial angiogenesis. Reproduction 2001, 121, 181–186. [Google Scholar] [CrossRef]
- Liu, S.; Xin, X.; Hua, T.; Shi, R.; Chi, S.; Jin, Z.; Wang, H. Efficacy of Anti-VEGF/VEGFR Agents on Animal Models of Endometriosis: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0166658. [Google Scholar] [CrossRef] [Green Version]
- Schmidinger, M. Understanding and managing toxicities of vascular endothelial growth factor (VEGF) inhibitors. Eur. J. Cancer Suppl. 2013, 11, 172–191. [Google Scholar] [CrossRef] [Green Version]
- Wulff, C.; Dickson, S.E.; Duncan, W.C.; Fraser, H.M. Angiogenesis in the human corpus luteum: Simulated early pregnancy by HCG treatment is associated with both angiogenesis and vessel stabilization. Hum. Reprod. 2001, 16, 2515–2524. [Google Scholar] [CrossRef]
- Zimmermann, R.C.; Hartman, T.; Bohlen, P.; Sauer, M.V.; Kitajewski, J. Preovulatory Treatment of Mice with Anti-VEGF Receptor 2 Antibody Inhibits Angiogenesis in Corpora Lutea. Microvasc. Res. 2001, 62, 15–25. [Google Scholar] [CrossRef]
- Zimmermann, R.C.; Xiao, E.; Husami, N.; Sauer, M.V.; Lobo, R.; Kitajewski, J.; Ferin, M. Short-Term Administration of Antivascular Endothelial Growth Factor Antibody in the Late Follicular Phase Delays Follicular Development in the Rhesus Monkey 1. J. Clin. Endocrinol. Metab. 2001, 86, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, S.A.; Tang, H.; Wang, J.; Bohlen, P.; Posser, R.; Hartman, T.; Sauer, M.V.; Kitajewski, J.; Zimmermann, R.C. The Vascular Endothelial Growth Factor (VEGF)/VEGF Receptor 2 Pathway Is Critical for Blood Vessel Survival in Corpora Lutea of Pregnancy in the Rodent. Endocrinology 2005, 146, 1301–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockwell, L.C.; Pillai, S.; Olson, C.E.; Koos, R.D. Inhibition of Vascular Endothelial Growth Factor/Vascular Permeability Factor Action Blocks Estrogen-Induced Uterine Edema and Implantation in Rodents1. Biol. Reprod. 2002, 67, 1804–1810. [Google Scholar] [CrossRef] [Green Version]
- Heryanto, B.; Lipson, K.E.; Rogers, P.A.W. Effect of angiogenesis inhibitors on oestrogen-mediated endometrial endothelial cell proliferation in the ovariectomized mouse. Reproduction 2003, 125, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.C.; Hartman, T.; Kavic, S.; Pauli, S.A.; Bohlen, P.; Sauer, M.V.; Kitajewski, J. Vascular endothelial growth factor receptor 2-mediated angiogenesis is essential for gonadotropin-dependent follicle development. J. Clin. Invest. 2003, 5, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Robert, E.; Musatti, L.; Piscitelli, G.; Ferrari, C.I. Pregnancy outcome after treatment with the ergot derivative, cabergoline. Reprod. Toxicol. 1996, 10, 333–337. [Google Scholar] [CrossRef]
- Gomez, R.; Gonzalez-Izquierdo, M.; Zimmermann, R.C.; Novella-Maestre, E.; Alonso-Muriel, I.; Sanchez-Criado, J.; Remohi, J.; Simon, C.; Pellicer, A. Low-Dose Dopamine Agonist Administration Blocks Vascular Endothelial Growth Factor (VEGF)-Mediated Vascular Hyperpermeability without Altering VEGF Receptor 2-Dependent Luteal Angiogenesis in a Rat Ovarian Hyperstimulation Model. Endocrinology 2006, 147, 5400–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mornex, R.; Orgiazzi, J.; Hugues, B.; Gagnaire, J.-C.; Claustrat, B. Normal Pregnancies after Treatment of Hyperprolactinemia with Bromoergocryptine, Despite Suspected Pituitary Tumors. J. Clin. Endocrinol. Metab. 1978, 47, 290–295. [Google Scholar] [CrossRef]
- Basu, S.; Nagy, J.A.; Pal, S.; Vasile, E.; Eckelhoefer, I.A.; Bliss, V.S.; Manseau, E.J.; Dasgupta, P.S.; Dvorak, H.F.; Mukhopadhyay, D. The neurotransmitter dopamine inhibits angiogenesis induced by vascular permeability factor/vascular endothelial growth factor. Nat. Med. 2001, 7, 569–574. [Google Scholar] [CrossRef]
- Delgado-Rosas, F.; Gómez, R.; Ferrero, H.; Gaytan, F.; Garcia-Velasco, J.; Simon, C.; Pellicer, A. The effects of ergot and non-ergot-derived dopamine agonists in an experimental mouse model of endometriosis. Reproduction 2011, 142, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Grümmer, R.; Schwarzer, F.; Bainczyk, K.; Hess-Stumpp, H.; Regidor, P.-A.; Schindler, A.E.; Winterhager, E. Peritoneal endometriosis: Validation of an in-vivo model. Hum. Reprod. 2001, 16, 1736–1743. [Google Scholar] [CrossRef] [Green Version]
- Novella-Maestre, E.; Carda, C.; Noguera, I.; Ruiz-Saurí, A.; García-Velasco, J.A.; Simón, C.; Pellicer, A. Dopamine agonist administration causes a reduction in endometrial implants through modulation of angiogenesis in experimentally induced endometriosis. Hum. Reprod. 2009, 24, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Novella-Maestre, E.; Carda, C.; Ruiz-Sauri, A.; Garcia-Velasco, J.A.; Simon, C.; Pellicer, A. Identification and Quantification of Dopamine Receptor 2 in Human Eutopic and Ectopic Endometrium: A Novel Molecular Target for Endometriosis Therapy. Biol. Reprod. 2010, 83, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Novella-Maestre, E.; Herraiz, S.; Vila-Vives, J.M.; Carda, C.; Ruiz-Sauri, A.; Pellicer, A. Effect of antiangiogenic treatment on peritoneal endometriosis-associated nerve fibers. Fertil. Steril. 2012, 98, 1209–1217. [Google Scholar] [CrossRef]
- Gomez, R.; Abad, A.; Delgado, F.; Tamarit, S.; Simon, C.; Pellicer, A. Effects of hyperprolactinemia treatment with the dopamine agonist quinagolide on endometriotic lesions in patients with endometriosis-associated hyperprolactinemia. Fertil. Steril. 2011, 95, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Menger, M.D. Basic mechanisms of vascularization in endometriosis and their clinical implications. Hum. Reprod. Updat. 2018, 24, 207–224. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, Y.M.; Dutta, S.; Iyer, A.K.V.; Wright, C.A.; Ramesh, V.; Kaushik, V.; Semmes, O.J.; Azad, N. A Lipidomics Approach to Identifying Key Lipid Species Involved in VEGF-Inhibitor Mediated Attenuation of Bleomycin-Induced Pulmonary Fibrosis. Proteom. Clin. Appl. 2018, 12, e1700086. [Google Scholar] [CrossRef] [PubMed]
- García-Pascual, C.M.; Martinez, J.; Calvo, P.; Ferrero, H.; Villanueva, A.; Pozuelo-Rubio, M.; Soengas, M.; Tormo, D.; Simon, C.; Pellicer, A.; et al. Evaluation of the potential therapeutic effects of a double-stranded RNA mimic complexed with polycations in an experimental mouse model of endometriosis. Fertil. Steril. 2015, 104, 1310–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.; Bisbal, V.; Marin, N.; Cano, A.; Gómez, R. Noninvasive Monitoring of Lesion Size in a Heterologous Mouse Model of Endometriosis. J. Vis. Exp. 2019, e58358. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, L.; Shinkaruk, S.; Lequin, O.; Rousseau, B.; Hagedorn, M.; Costa, F.; Caronzolo, D.; Balke, M.; Canron, X.; Convert, O.; et al. Structure and Inhibitory Effects on Angiogenesis and Tumor Development of a New Vascular Endothelial Growth Inhibitor. J. Biol. Chem. 2003, 278, 35564–35573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, S.; Ishii, K.; Yamane, J.; Iwanami, A.; Ikegami, T.; Katoh, H.; Iwamoto, Y.; Nakamura, M.; Miyoshi, H.; Okano, H.J.; et al. In vivo imaging of engrafted neural stem cells: Its application in evaluating the optimal timing of transplantation for spinal cord injury. FASEB J. 2005, 19, 1839–1841. [Google Scholar] [CrossRef] [Green Version]
- Fortin, M.; Lépine, M.; Merlen, Y.; Thibeault, I.; Rancourt, C.; Gosselin, D.; Hugo, P.; Steff, A.-M. Quantitative Assessment of Human Endometriotic Tissue Maintenance and Regression in a Noninvasive Mouse Model of Endometriosis. Mol. Ther. 2004, 9, 540–547. [Google Scholar] [CrossRef]
- Masuda, H.; Maruyama, T.; Hiratsu, E.; Yamane, J.; Iwanami, A.; Nagashima, T.; Ono, M.; Miyoshi, H.; Okano, H.J.; Ito, M.; et al. Noninvasive and real-time assessment of reconstructed functional human endometrium in NOD/SCID/ Formula immunodeficient mice. Proc. Natl. Acad. Sci. USA 2007, 104, 1925–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Pascual, C.M.; Zimmermann, R.C.; Ferrero, H.; Shawber, C.J.; Kitajewski, J.; Simon, C.; Pellicer, A.; Gomez, R. Delta-like ligand 4 regulates vascular endothelial growth factor receptor 2–driven luteal angiogenesis through induction of a tip/stalk phenotype in proliferating endothelial cells. Fertil. Steril. 2013, 100, 1768–1776. [Google Scholar] [CrossRef]
- García-Pascual, C.M.; Ferrero, H.; Juarez, I.; Martínez, J.; Villanueva, A.; Pozuelo-Rubio, M.; Soengas, M.; Tormo, D.; Simón, C.; Gómez, R.; et al. Evaluation of the antiproliferative, proapoptotic, and antiangiogenic effects of a double-stranded RNA mimic complexed with polycations in an experimental mouse model of leiomyoma. Fertil. Steril. 2016, 105, 529–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.-W.; Kim, D.-S.; Suhito, I.R.; Han, D.K.; Lee, T.; Kim, T.-H. Enhancing Neurogenesis of Neural Stem Cells Using Homogeneous Nanohole Pattern-Modified Conductive Platform. Int. J. Mol. Sci. 2019, 21, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherwood, L.M.; Parris, E.E.; Folkman, J. Tumor Angiogenesis: Therapeutic Implications. New Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef]
- Nap, A.W.; Griffioen, A.W.; Dunselman, G.A.J.; Steege, J.C.A.B.-T.; Thijssen, V.L.J.L.; Evers, J.L.H.; Groothuis, P.G. Antiangiogenesis Therapy for Endometriosis. J. Clin. Endocrinol. Metab. 2004, 89, 1089–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, M.L.; Charnock-Jones, D.S.; Chan, C.L.K.; Bruner-Tran, K.L.; Osteen, K.G.; Tom, B.D.M.; Fan, T.-P.D.; Smith, S.K. Antiangiogenic Agents Are Effective Inhibitors of Endometriosis. J. Clin. Endocrinol. Metab. 2003, 88, 2889–2899. [Google Scholar] [CrossRef] [Green Version]
- Douglas, N.C.; Tang, H.; Gomez, R.; Pytowski, B.; Hicklin, D.J.; Sauer, C.M.; Kitajewski, J.; Sauer, M.V.; Zimmermann, R.C. Vascular Endothelial Growth Factor Receptor 2 (VEGFR-2) Functions to Promote Uterine Decidual Angiogenesis during Early Pregnancy in the Mouse. Endocrinology 2009, 150, 3845–3854. [Google Scholar] [CrossRef] [Green Version]
- García-Pascual, C.; Ferrero, H.; Zimmermann, R.; Simón, C.; Pellicer, A.; Gómez, R. Inhibition of Delta-like 4 mediated signaling induces abortion in mice due to deregulation of decidual angiogenesis. Placenta 2014, 35, 501–508. [Google Scholar] [CrossRef]
- Boehm, S.; Rothermundt, C.; Hess, D.; Joerger, M. Antiangiogenic Drugs in Oncology: A Focus on Drug Safety and the Elderly—A Mini-Review. Gerontology 2010, 56, 303–309. [Google Scholar] [CrossRef]
- Riondino, S.; Del Monte, G.; Fratangeli, F.; Guadagni, F.; Roselli, M.; Ferroni, P. Anti-Angiogenic Drugs, Vascular Toxicity and Thromboembolism in Solid Cancer. Cardiovasc. Hematol. Agents Med. Chem. 2017, 15, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Van Langendonckt, A.; Donnez, J.; Defrère, S.; Dunselman, G.A.; Groothuis, P.G. Antiangiogenic and vascular-disrupting agents in endometriosis: Pitfalls and promises. Mol. Hum. Reprod. 2008, 14, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, K.; Becker, C.M. Endometriosis and angiogenesis. Minerva Ginecol. 2008, 60, 245–254. [Google Scholar] [PubMed]
- Becker, C.M.; Wright, R.D.; Satchi-Fainaro, R.; Funakoshi, T.; Folkman, J.; Kung, A.L.; D’Amato, R.J. A Novel Noninvasive Model of Endometriosis for Monitoring the Efficacy of Antiangiogenic Therapy. Am. J. Pathol. 2006, 168, 2074–2084. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, D.; Sarkar, C.; Yu, H.; Wang, J.; Liu, Z.; Dasgupta, P.S.; Basu, S. Dopamine stabilizes tumor blood vessels by up-regulating angiopoietin 1 expression in pericytes and Kruppel-like factor-2 expression in tumor endothelial cells. Proc. Natl. Acad. Sci. USA 2011, 108, 20730–20735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, T.; Wang, C.; Chen, X.; Duan, C.; Zhang, X.; Zhang, J.; Chai, H.; Tang, T.; Chen, H.; Yue, J.; et al. Dopamine induces growth inhibition and vascular normalization through reprogramming M2-polarized macrophages in rat C6 glioma. Toxicol. Appl. Pharmacol. 2015, 286, 112–123. [Google Scholar] [CrossRef]
- Alarcon-Martinez, L.; Yilmaz-Ozcan, S.; Yemisci, M.; Schallek, J.; Kılıç, K.; Can, A.; Di Polo, A.; Dalkara, T. Capillary pericytes express α-smooth muscle actin, which requires prevention of filamentous-actin depolymerization for detection. eLife 2018, 7, e34861. [Google Scholar] [CrossRef]
- Sarkar, C.; Chakroborty, D.; Mitra, R.B.; Banerjee, S.; Dasgupta, P.S.; Basu, S. Dopamine in vivo inhibits VEGF-induced phosphorylation of VEGFR-2, MAPK, and focal adhesion kinase in endothelial cells. Am. J. Physiol. Circ. Physiol. 2004, 287, H1554–H1560. [Google Scholar] [CrossRef]
- Laschke, M.; Giebels, C.; Menger, M. Vasculogenesis: A new piece of the endometriosis puzzle. Hum. Reprod. Updat. 2011, 17, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, D.; Chowdhury, U.R.; Sarkar, C.; Baral, R.; Dasgupta, P.S.; Basu, S. Dopamine regulates endothelial progenitor cell mobilization from mouse bone marrow in tumor vascularization. J. Clin. Investig. 2008, 118, 1380–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laschke, M.W.; Giebels, C.; Nickels, R.M.; Scheuer, C.; Menger, M.D. Endothelial Progenitor Cells Contribute to the Vascularization of Endometriotic Lesions. Am. J. Pathol. 2011, 178, 442–450. [Google Scholar] [CrossRef]
- Clinicaltrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03749109 (accessed on 11 September 2020).
- Hamid, A.M.S.A.; Madkour, W.A.I.; Moawad, A.; Elzaher, M.A.; Roberts, M.P. Does Cabergoline help in decreasing endometrioma size compared to LHRH agonist? A prospective randomized study. Arch. Gynecol. Obstet. 2014, 290, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinskaya, M.; Suslova, E.; Tkachenko, N.; Molotkov, A.; Kogan, I. Dopamine agonists as genital endometriosis target therapy. Gynecol. Endocrinol. 2020, 36, 7–11. [Google Scholar] [CrossRef]
- Tokushige, N.; Markham, R.; Russell, P.; Fraser, I.S. High density of small nerve fibres in the functional layer of the endometrium in women with endometriosis. Hum. Reprod. 2006, 21, 782–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, L.K.; Thomas, J.R.; Lamps, L.W.; Smoller, B.R.; Folpe, A.L. Protein Gene Product 9.5 (PGP 9.5) Is Not a Specific Marker of Neural and Nerve Sheath Tumors: An Immunohistochemical Study of 95 Mesenchymal Neoplasms. Mod. Pathol. 2003, 16, 963–969. [Google Scholar] [CrossRef] [Green Version]
- Greaves, E.; Cousins, F.L.; Murray, A.; Esnal-Zufiaurre, A.; Fassbender, A.; Horne, A.W.; Saunders, P.T. A Novel Mouse Model of Endometriosis Mimics Human Phenotype and Reveals Insights into the Inflammatory Contribution of Shed Endometrium. Am. J. Pathol. 2014, 184, 1930–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Kim, Y.J.; Kim, M.J.; Lee, S.J.; Kwon, H.; Lee, J.H. Novel Medicine for Endometriosis and Its Therapeutic Effect in a Mouse Model. Biomedicines 2020, 8, 619. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejada, M.Á.; Santos-Llamas, A.I.; Fernández-Ramírez, M.J.; Tarín, J.J.; Cano, A.; Gómez, R. A Reassessment of the Therapeutic Potential of a Dopamine Receptor 2 Agonist (D2-AG) in Endometriosis by Comparison against a Standardized Antiangiogenic Treatment. Biomedicines 2021, 9, 269. https://doi.org/10.3390/biomedicines9030269
Tejada MÁ, Santos-Llamas AI, Fernández-Ramírez MJ, Tarín JJ, Cano A, Gómez R. A Reassessment of the Therapeutic Potential of a Dopamine Receptor 2 Agonist (D2-AG) in Endometriosis by Comparison against a Standardized Antiangiogenic Treatment. Biomedicines. 2021; 9(3):269. https://doi.org/10.3390/biomedicines9030269
Chicago/Turabian StyleTejada, Miguel Á., Ana I. Santos-Llamas, María José Fernández-Ramírez, Juan J. Tarín, Antonio Cano, and Raúl Gómez. 2021. "A Reassessment of the Therapeutic Potential of a Dopamine Receptor 2 Agonist (D2-AG) in Endometriosis by Comparison against a Standardized Antiangiogenic Treatment" Biomedicines 9, no. 3: 269. https://doi.org/10.3390/biomedicines9030269