
Acetylated Diacylglycerol 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol in Autoimmune Arthritis and Interstitial Lung Disease in SKG Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Induction of Arthritis and Interstitial Lung Disease in SKG Mice
2.2. Clinical Scoring of Peripheral Arthritis
2.3. Lung Histology
2.4. Analysis of the Serum Cytokines by a Luminex Multiplex Cytokine Assay
2.5. Immunofluorescent Staining
2.6. Statistical Analysis
3. Results
3.1. Effect of PLAG on Arthritis
3.2. Effect of PLAG on Lung Inflammation and Fibrosis
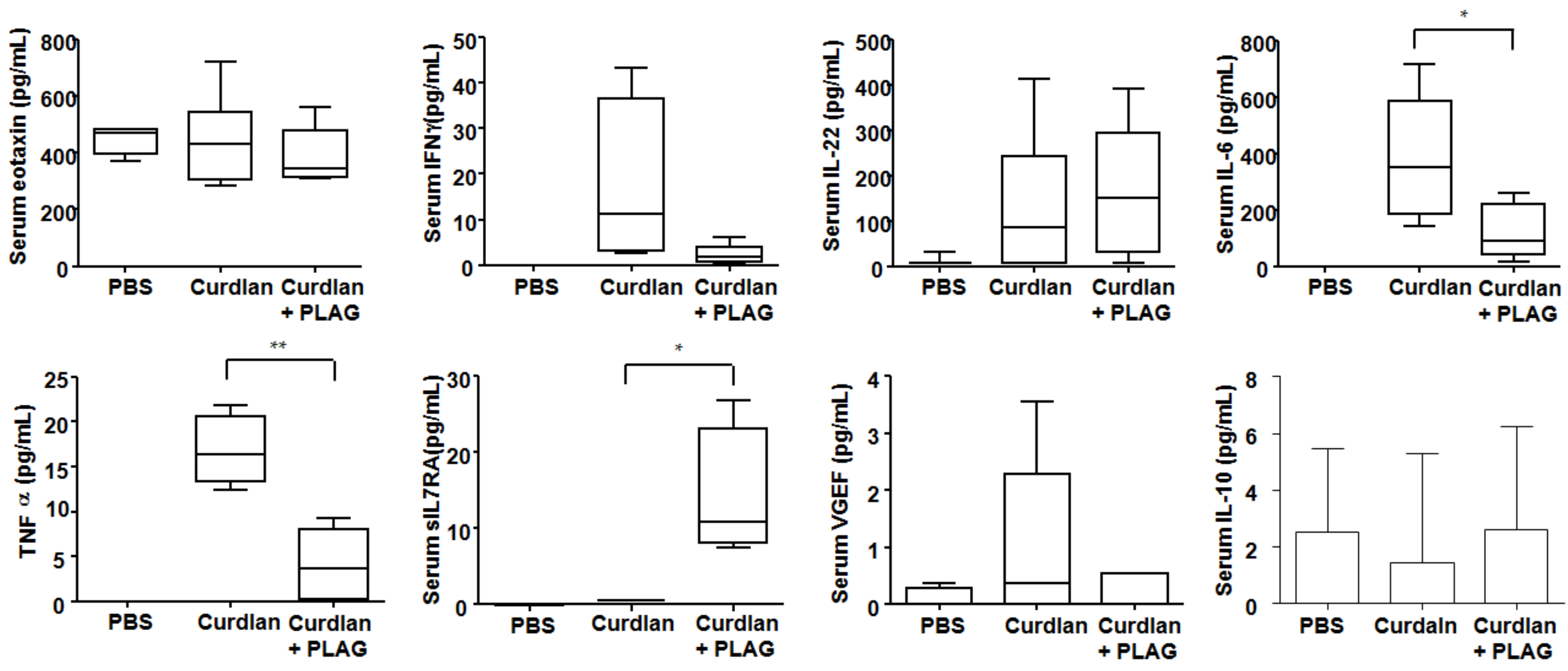
3.3. Changes of Serum Cytokines after PLAG Treatment
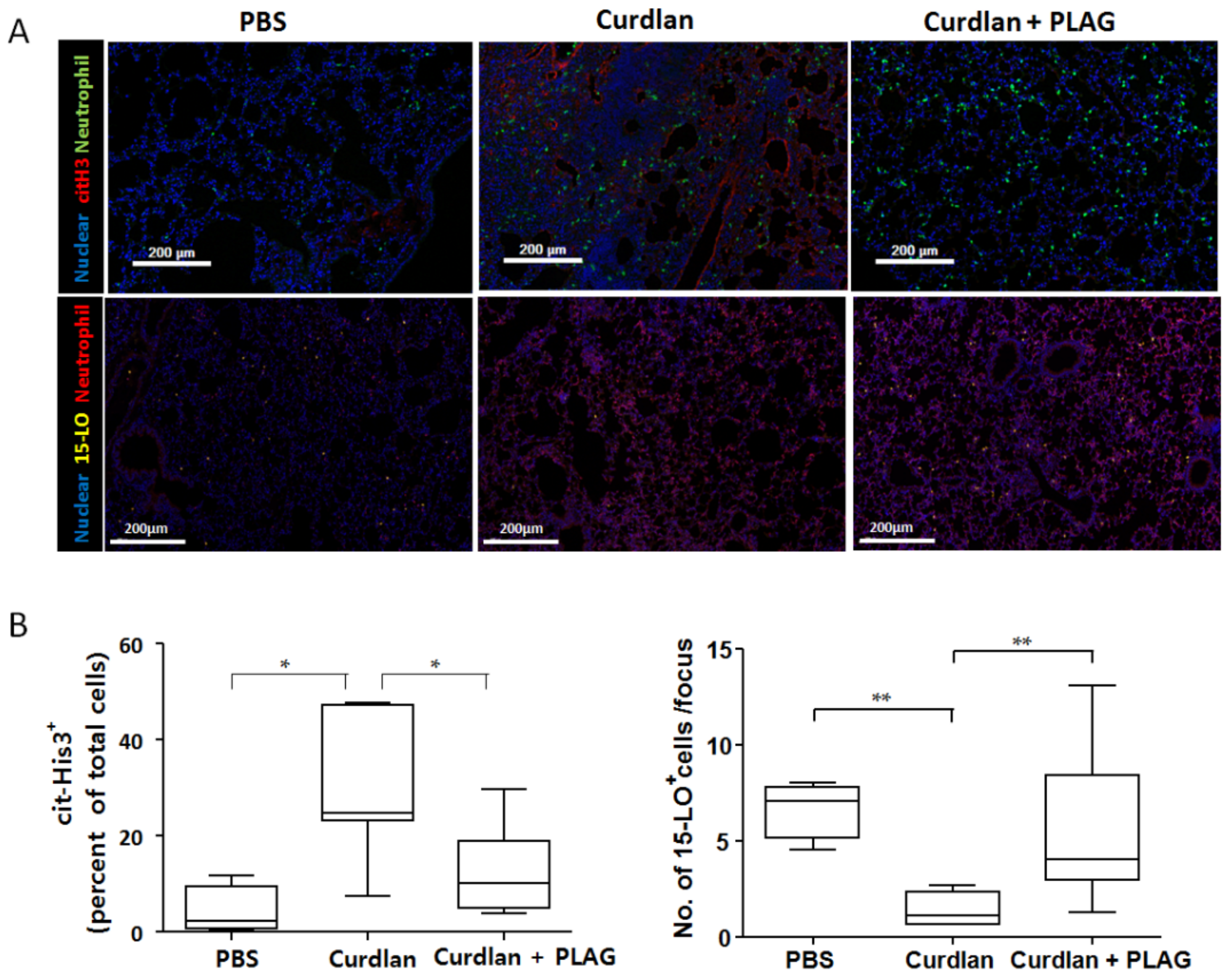
3.4. Citrullinated Histone 3 Expression (NETosis) in the Lung Tissue after PLAG Treatment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, A.U.; Denton, C.P. Interstitial lung disease in connective tissue disease--mechanisms and management. Nat. Rev. Rheumatol. 2014, 10, 728–739. [Google Scholar] [CrossRef]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Gutsche, M.; Rosen, G.D.; Swigris, J.J. Connective Tissue Disease-associated Interstitial Lung Disease: A review. Curr. Respir. Care Rep. 2012, 1, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Klareskog, L.; Stolt, P.; Lundberg, K.; Källberg, H.; Bengtsson, C.; Grunewald, J.; Rönnelid, J.; Harris, H.E.; Ulfgren, A.K.; Rantapää-Dahlqvist, S.; et al. A new model for an etiology of rheumatoid arthritis: Smoking may trigger HLA-DR (shared epitope)-restricted immune reactions to autoantigens modified by citrullination. Arthritis Rheum. 2006, 54, 38–46. [Google Scholar] [CrossRef]
- Castelino, F.V.; Varga, J. Interstitial lung disease in connective tissue diseases: Evolving concepts of pathogenesis and management. Arthritis Res. Ther. 2010, 12, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Sung, Y.-K. Epidemiology of Rheumatoid Arthritis in Korea. J. Rheum Dis. 2021, 28, 60–67. [Google Scholar] [CrossRef]
- Olson, A.L.; Swigris, J.J.; Sprunger, D.B.; Fischer, A.; Fernandez-Perez, E.R.; Solomon, J.; Murphy, J.; Cohen, M.; Raghu, G.; Brown, K.K. Rheumatoid arthritis-interstitial lung disease-associated mortality. J. Respir. Crit. Care Med. 2011, 183, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, M.; Maini, S.R. Role of cytokines in rheumatoid arthritis: An education in pathophysiology and therapeutics. Immunol. Rev. 2008, 223, 7–19. [Google Scholar] [CrossRef]
- D’Elia, T. Methotrexate-induced pneumonitis: Heterogeneity of bronchoalveolar lavage and differences between cancer and rheumatoid arthritis. Inflamm. Allergy Drug Targets 2014, 13, 25–33. [Google Scholar] [CrossRef]
- Ostör, A.J.; Chilvers, E.R.; Somerville, M.F.; Lim, A.Y.; Lane, S.E.; Crisp, A.J.; Scott, D.G. Pulmonary complications of infliximab therapy in patients with rheumatoid arthritis. J. Rheumatol. 2006, 33, 622–628. [Google Scholar] [PubMed]
- Peno-Green, L.; Lluberas, G.; Kingsley, T.; Brantley, S. Lung injury linked to etanercept therapy. Chest 2002, 122, 1858–1860. [Google Scholar] [CrossRef]
- Nagasawa, Y.; Takada, T.; Shimizu, T.; Narita, J.; Moriyama, H.; Terada, M.; Suzuki, E.; Gejyo, F. Inflammatory cells in lung disease associated with rheumatoid arthritis. Intern. Med. 2009, 48, 1209–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, O.C.; Lee, E.J.; Chang, E.J.; Youn, J.; Ghang, B.; Hong, S.; Lee, C.K.; Yoo, B.; Kim, Y.G. IL-17A(+)GM-CSF(+) Neutrophils Are the Major Infiltrating Cells in Interstitial Lung Disease in an Autoimmune Arthritis Model. Front. Immunol. 2018, 9, 1544. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, P.; Salaffi, F.; Pesci, A. Clinical and subclinical alveolitis in connective tissue diseases assessed by bronchoalveolar lavage. Semin. Arthritis Rheum. 1997, 26, 740–754. [Google Scholar] [CrossRef]
- Garcia, J.G.; Parhami, N.; Killam, D.; Garcia, P.L.; Keogh, B.A. Bronchoalveolar lavage fluid evaluation in rheumatoid arthritis. Am. Rev. Respir. Dis. 1986, 133, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.G.; James, H.L.; Zinkgraf, S.; Perlman, M.B.; Keogh, B.A. Lower respiratory tract abnormalities in rheumatoid interstitial lung disease. Potential role of neutrophils in lung injury. Am. Rev. Respir. Dis. 1987, 136, 811–817. [Google Scholar] [CrossRef]
- Zhang, S.; Shu, X.; Tian, X.; Chen, F.; Lu, X.; Wang, G. Enhanced formation and impaired degradation of neutrophil extracellular traps in dermatomyositis and polymyositis: A potential contributor to interstitial lung disease complications. Clin. Exp. Immunol. 2014, 177, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.O.; Kim, S.H.; Cho, S.H.; Kim, M.G.; Seo, J.Y.; Park, J.S.; Jhon, G.J.; Han, S.Y. Purification and structural determination of hematopoietic stem cell-stimulating monoacetyldiglycerides from Cervus nippon (deer antler). Chem. Pharm. Bull. 2004, 52, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.O.; Park, J.S.; Cho, S.H.; Yoon, J.Y.; Kim, M.G.; Jhon, G.J.; Han, S.Y.; Kim, S.H. Stimulatory effects of monoacetyldiglycerides on hematopoiesis. Biol. Pharm. Bull. 2004, 27, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Shin, J.M.; Shin, S.H.; Kim, J.H.; Sohn, K.Y.; Kim, H.J.; Kang, J.K.; Yoon, S.Y.; Kim, J.W. 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol ameliorates arthritic joints through reducing neutrophil infiltration mediated by IL-6/STAT3 and MIP-2 activation. Oncotarget 2017, 8, 96636–96648. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.R.; Shin, S.H.; Kim, J.H.; Sohn, K.Y.; Yoon, S.Y.; Kim, J.W. 1-Palmitoyl-2-Linoleoyl-3-Acetyl-rac-Glycerol (PLAG) Rapidly Resolves LPS-Induced Acute Lung Injury Through the Effective Control of Neutrophil Recruitment. Front. Immunol. 2019, 10, 2177. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.H.; Lee, E.J.; Kwon, O.C.; Hong, S.; Lee, C.K.; Yoo, B.; Youn, J.; Kim, T.H.; Kim, Y.G. Effect of tumor necrosis factor inhibition on spinal inflammation and spinal ankylosis in SKG mice. Sci. Rep. 2019, 9, 18000. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Hashimoto, M.; Ito, Y.; Matsuura, M.; Ito, H.; Tanaka, M.; Watanabe, H.; Kondoh, G.; Tanaka, A.; Yasuda, K.; et al. Autoimmune Th17 Cells Induced Synovial Stromal and Innate Lymphoid Cell Secretion of the Cytokine GM-CSF to Initiate and Augment Autoimmune. Arthritis Immunol. 2018, 48, 1220.e1225–1232.e1225. [Google Scholar] [CrossRef] [PubMed]
- Ruutu, M.; Thomas, G.; Steck, R.; Degli-Esposti, M.A.; Zinkernagel, M.S.; Alexander, K.; Velasco, J.; Strutton, G.; Tran, A.; Benham, H.; et al. β-glucan triggers spondylarthritis and Crohn’s disease-like ileitis in SKG mice. Arthritis Rheum. 2012, 64, 2211–2222. [Google Scholar] [CrossRef]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakasa-Morimoto, C.; Toyosaki-Maeda, T.; Matsutani, T.; Yoshida, R.; Nakamura-Kikuoka, S.; Maeda-Tanimura, M.; Yoshitomi, H.; Hirota, K.; Hashimoto, M.; Masaki, H.; et al. Arthritis and pneumonitis produced by the same T cell clones from mice with spontaneous autoimmune arthritis. Int. Immunol. 2008, 20, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Turesson, C.; Matteson, E.L.; Colby, T.V.; Vuk-Pavlovic, Z.; Vassallo, R.; Weyand, C.M.; Tazelaar, H.D.; Limper, A.H. Increased CD4+ T cell infiltrates in rheumatoid arthritis-associated interstitial pneumonitis compared with idiopathic interstitial pneumonitis. Arthritis Rheum. 2005, 52, 73–79. [Google Scholar] [CrossRef]
- Weiss, S.J. Tissue destruction by neutrophils. N. Engl. J. Med. 1989, 320, 365–376. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Fattman, C.L.; Tan, R.J.; Oury, T.D. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am. J. Respir. Crit. Care Med. 2005, 172, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Qi, S.; den Hartog, G.J.; Bast, A. Superoxide radicals increase transforming growth factor-beta1 and collagen release from human lung fibroblasts via cellular influx through chloride channels. Toxicol. Appl. Pharmacol. 2009, 237, 111–118. [Google Scholar] [CrossRef]
- Henson, P.M.; Johnston, R.B., Jr. Tissue injury in inflammation. Oxidants, proteinases, and cationic proteins. J. Clin. Investig. 1987, 79, 669–674. [Google Scholar] [CrossRef] [Green Version]
- Malech, H.L.; Gallin, J.I. Current concepts: Immunology. Neutrophils in human diseases. N. Engl. J. Med. 1987, 317, 687–694. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Leppkes, M.; Munoz, L.E.; Schley, G.; Schett, G.; Herrmann, M. Extracellular DNA traps in inflammation, injury and healing. Nat. Rev. Nephrol. 2019, 15, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Corsiero, E.; Pratesi, F.; Prediletto, E.; Bombardieri, M.; Migliorini, P. NETosis as Source of Autoantigens in Rheumatoid Arthritis. Front. Immunol. 2016, 7, 485. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, S.; Zhao, Y.; Liu, Y.; Yan, B. Neutrophil extracellular traps may contribute to interstitial lung disease associated with anti-MDA5 autoantibody positive dermatomyositis. Clin. Rheumatol. 2018, 37, 107–115. [Google Scholar] [CrossRef]
- Trapnell, B.C.; Whitsett, J.A. Gm-CSF regulates pulmonary surfactant homeostasis and alveolar macrophage-mediated innate host defense. Annu. Rev. Physiol. 2002, 64, 775–802. [Google Scholar] [CrossRef]
- Laan, M.; Prause, O.; Miyamoto, M.; Sjöstrand, M.; Hytönen, A.M.; Kaneko, T.; Lötvall, J.; Lindén, A. A role of GM-CSF in the accumulation of neutrophils in the airways caused by IL-17 and TNF-alpha. Eur. Respir. J. 2003, 21, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef]
- Jones, H.R.; Robb, C.T.; Perretti, M.; Rossi, A.G. The role of neutrophils in inflammation resolution. Semin. Immunol. 2016, 28, 137–145. [Google Scholar] [CrossRef]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid mediator class switching during acute inflammation: Signals in resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Fierro, I.M.; Colgan, S.P.; Bernasconi, G.; Petasis, N.A.; Clish, C.B.; Arita, M.; Serhan, C.N. Lipoxin A4 and aspirin-triggered 15-epi-lipoxin A4 inhibit human neutrophil migration: Comparisons between synthetic 15 epimers in chemotaxis and transmigration with microvessel endothelial cells and epithelial cells. J. Immunol. 2003, 170, 2688–2694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.E.; Yoon, S.Y.; Ly, S.Y.; Kim, J.H.; Sohn, K.Y.; Kim, J.W. 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol (PLAG) reduces hepatic injury in concanavalin A-treated mice. J. Cell Biochem. 2018, 119, 1392–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, D.-H.; Lee, E.-J.; Lee, H.-S.; Kim, D.H.; Lee, J.-H.; Jeong, M.R.; Hong, S.; Lee, C.-K.; Yoo, B.; Youn, J.; et al. Acetylated Diacylglycerol 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol in Autoimmune Arthritis and Interstitial Lung Disease in SKG Mice. Biomedicines 2021, 9, 1095. https://doi.org/10.3390/biomedicines9091095
Lim D-H, Lee E-J, Lee H-S, Kim DH, Lee J-H, Jeong MR, Hong S, Lee C-K, Yoo B, Youn J, et al. Acetylated Diacylglycerol 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol in Autoimmune Arthritis and Interstitial Lung Disease in SKG Mice. Biomedicines. 2021; 9(9):1095. https://doi.org/10.3390/biomedicines9091095
Chicago/Turabian StyleLim, Doo-Ho, Eun-Ju Lee, Hee-Seop Lee, Do Hoon Kim, Jae-Hyun Lee, Mi Ryeong Jeong, Seokchan Hong, Chang-Keun Lee, Bin Yoo, Jeehee Youn, and et al. 2021. "Acetylated Diacylglycerol 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol in Autoimmune Arthritis and Interstitial Lung Disease in SKG Mice" Biomedicines 9, no. 9: 1095. https://doi.org/10.3390/biomedicines9091095