
Toxic Effects of Two Representative Rare Earth Elements (La and Gd) on Danio rerio Based on Transcriptome Analysis
by
Shu Kang
1,2,3,†,
Cheng Guo
3,†,
Chenyang Xue
2,
Chenshu Ma
1,4,
Huaizhong Mu
1 and
Lizong Sun
2,* 1
School of Public Management, Liaoning University, Shenyang 110016, China
2
Key Laboratory of Pollution Ecology and Environmental Engineering, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016, China
3
School of Environmental and Safety Engineering, Liaoning Petrochemical University, Fushun 113001, China
4
Liaoning Economic Vocational Technological Institute, Shenyang 110016, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Toxics 2022, 10(9), 519; https://doi.org/10.3390/toxics10090519
Submission received: 17 July 2022
/
Revised: 25 August 2022
/
Accepted: 30 August 2022
/
Published: 31 August 2022
(This article belongs to the Section Toxicology)
Abstract
:The expanding applications of rare earth elements (REEs) in various fields have raised concerns about their biosafety. However, previous studies are insufficient to elucidate their toxic effects and mechanisms of action and whether there are uniform or predictable toxicity patterns among REEs. Herein, we investigated the toxic effects of two representative REEs (lanthanum (La) and gadolinium (Gd)) on zebrafish (Danio rerio) through toxicity experiments and transcriptome analysis. The results of the toxicity experiments showed that the two REEs have similar lethality, with half-lethal concentrations (LC50) at micromolar levels and mixed toxicity showing additive effects. Differential expression gene screening and functional group enrichment analysis showed that La and Gd might affect the growth and development of Danio rerio by interfering with some biological molecules. The two REEs showed significant effects on the metabolic pathways of exogenous or endogenous substances, including glutathione sulfotransferase and acetaldehyde dehydrogenase. Moreover, some basic biological processes, such as DNA replication, the insulin signaling pathway, and the p53 signaling pathway, were significantly enriched. Overall, the toxicity patterns of La and Gd may affect some biological processes with different intensities; however, there are many similarities in their toxicity mechanisms and modes of action. The concentrations investigated in this study were comparable to those of REE residues at highly contaminated sites, thus mimicking the ecotoxicological effects at environmentally relevant concentrations.
1. Introduction
Rare earth elements (REEs) include 15 lanthanides (Ln) (ranging from lanthanum (Z = 57) to lutetium (Z = 71)), as well as scandium and yttrium. Owing to their irreplaceable material properties, REEs have become an important strategic resource and are widely used in the fields of electronics, metallurgy, energy, medicine, and agriculture [1,2]. In the past few decades, the global demand for REEs has increased exponentially, in turn disrupting their biogeochemical cycle [3,4]. Moreover, the high demand for REEs has triggered rapid growth in the mining industry. Nonetheless, REEs readily deposit into the environment and are, thus, regarded as emerging contaminants [5,6,7]. Recent studies have shown that REEs are abundantly enriched in soils and plants in mining areas [8]. Their concentrations in topsoil are more than 10 times the background value and are also increasingly deposited in aquatic ecosystems [9]. Owing to this, their biosafety and related ecotoxicological effects are receiving increasing attention [10,11,12,13,14,15,16].
Compared to extensive studies that have been performed on heavy metals (Cu, Zn, Cd, and Pb), only a few studies have reported on the ecotoxicity of REEs [9]. Most existing studies on REEs have focused on geological, mineralogical, and material properties, with their environmental risks receiving little attention [17]. Presently, the reported toxicity data for REEs are mostly concentrated on single elements, especially lanthanum (La) and cerium (Ce) [18,19,20]. To predict and explain the biological effects of other REEs, it is important to understand and determine whether extrapolation can be applied to deduce the properties of other REEs based on the ecotoxicity of one REE. Gonzalez et al. [21] statistically analyzed hundreds of reports on the toxicity of REEs in aquatic organisms and found that the toxicity data in literature immensely varied, failing to determine any form of regular pattern or correlation that could be inferred from similarities in their atomic properties.
To date, few studies have reported on the mixed toxic effects of REEs on aquatic organisms [22,23,24,25,26,27]. Tai et al. [22] found that the biological toxicities of 13 REEs to unicellular algae (Chlorella autotrophica) were essentially the same, and the toxicity of the mixture of components was similar to that of a single element. Blinova et al. [23] found that the lethal concentrations (LC50) of five REEs in freshwater crustaceans were between 0.3 and 0.5 mg L−1, suggesting that REEs can be regarded as a uniform group of elements with an additive toxicity model despite their mechanisms of action revealed. However, Romero-Freire et al. [24,25] found that REEs are more toxic to low-trophic organisms, with mixed toxicity having an additive effect on low-trophic organisms (such as bacteria) and an antagonistic effect on high-trophic organisms (such as zebrafish). Bing et al. [26] found that three REEs (Y, La, and Ce) had similar Km and Jmax values through comparison using a simple biological ligand model. However, Yang et al. [27] concluded that the ligand model could not adequately explain the toxicity of REEs because the mixtures of different components are unlikely to have exactly the same or independent modes of action. Thus, it is difficult to compare the general patterns of REEs in terms of toxicity data from the currently available literature. Owing to this, a comparative study on the toxicity of REEs should not rely exclusively on model predictions and toxicity data. Along with paying attention to exposure conditions, tested species, and test endpoints, it is also necessary to clarify the toxicity mechanism of REEs at the molecular level to determine whether it shows a predictable model of the toxicity effects and mechanisms [21,26,27].
Transcriptome analysis has been efficiently applied in risk assessments of chemical substances and ecotoxicological mechanism investigations [28]. Compared to traditional toxicological analysis, transcriptome analysis has obvious advantages in distinguishing toxicity differences and action patterns of harmful substances [29,30]. Therefore, in this study, two representative REEs, La and gadolinium (Gd), were selected as test elements to explore their ecotoxic effects and mechanisms in zebrafish (Danio rerio). This study aimed: (1) to assess the toxic effects and modes of action of the two REEs and their mixtures on Danio rerio, (2) to clarify their toxic mechanisms based on transcriptome analysis, and (3) to investigate the toxicological differences and potential toxicity patterns between the two REEs.
2. Materials and Methods
2.1. Materials and Reagents
Zebrafish (Danio rerio, AB strain), about 2 months old with body lengths of 2.0 ± 0.5 cm were purchased from the Wuhan Institute of Aquatic Biology, Chinese Academy of Sciences. The fish were raised in an aquatic system. They were housed under the following conditions: a light/dark cycle of 14/l0 h, water temperature maintained at 26 ± 1 °C, and a pH of 7.0 ± 0.5. After domestication in the experimental environment for 2 weeks, no obvious diseases or deformities were visible. The selected two representative REEs (LaCl36H2O and GdCl36H2O (purity > 99.9%)) were purchased from Sterem Chemicals (MA, USA). These two REEs, which are relatively rich in the earth’s crust, are representative of odd REEs (atomic number: 57La) and even REEs (64Gd) and are representative of light and heavy REEs, respectively.
2.2. Acute Toxicity Test
The culture and acute toxicity tests of Danio rerio were performed as per the protocol outlined in the fish acute toxicity test (OECD 203) and method for determination of acute toxicity of water quality substances to freshwater fish (zebrafish) (GB/T13267-91). Standard dilution water was used after aeration for 48 h. For the configuration method, 11.76 g calcium chloride (CaCl2·2H2O) was dissolved in water and diluted to 1 L, 4.93 g magnesium sulfate (MgSO4·7H2O) was dissolved in water and diluted to 1 L, 2.59 g sodium bicarbonate (NaHCO3) was dissolved in water and diluted to 1 L, and 0.23 g potassium chloride (KCl) was dissolved in water and diluted to 1 L. Then, 25 mL of each of the above four solutions was taken, mixed, and diluted to 1 L with distilled water and aerated until the dissolved oxygen concentration reached the air saturation value. The newly configured standard dilution water had a pH of 7.8 ± 0.2 and a hardness of 250 mg L−1. The molarity of each ion in the standard water is shown in Table S1.
Based on the pre-experiment results, a static exposure toxicity test was used to set the concentrations of REEs to 0, 20, 40, 80, 160, and 320 μmol L−1, with three replicates being performed for each treatment. Eight zebrafish were placed in each tank (20 × 20 × 25 cm, 10 L) with a water volume of 2 L. Before the beginning of the toxicity test, the 24 h-LC50 of K2Cr2O7 in zebrafish was determined to be 298.7 ± 23.7 mg L−1, confirming that the zebrafish were not in distress. In order to ensure that the pH of all treatments was maintained at 7.0, 1 mol L−1 NaOH and HCL were used to regulate pH before the toxicity test. Before the test, zebrafish were domesticated in continuously aerated water for more than 7 days under environmental conditions similar to those of the test in terms of water quality and lighting. Feeding was stopped 24 h before the test. The Ln concentrations in the test solution were measured at the beginning and end of the test to detect that the chemical properties of Ln were stable. Dissolved oxygen (DO) concentration, pH, and temperature were measured in each vessel at the beginning and end of the test and once a day. The exposure water was changed every 24 h. Ammonia concentrations were monitored at the beginning and end of each water change. Ammonia concentration measurements during 72 h acute toxicity are shown in Table S2. During this period, fish activity and poisoning symptoms were observed, and dead fish were removed immediately. The number of dead animals was recorded after 72 h. The semilethal concentrations (72 h-LC50) following each treatment were calculated according to the probability unit-logarithmic graphic method.
2.3. Bioaccumulation
Based on Section 2.2, three treatments were set up: 0 (control), 0.1 LC50 (low-concentration group, 15 μmol L−1), and 0.2 LC50 (high-concentration group, 30 μmol L−1). Three replicates were set up for each treatment, and 50 zebrafish were placed in each tank. According to the fish prolonged toxicity test: 14-day study (OECD 204) guidelines, a semistatic test was used for the 28 d toxicity experiment. The test medium was changed once every 2 days, which is fully satisfactory for the chemically stable Ln that the concentration of the test solution fluctuates within 20% of the theoretical concentration value during the change in the solution. The DO concentration, pH, and temperature of each vessel solution were measured twice a week. The Ln concentration in the test solution was measured at the beginning of the test, before the first update of the test solution, and at the end of the test to check that the Ln remained relatively stable during the time of changing the test solution. Feeding was performed once a day, and the feeding amount did not exceed the food requirement of the test fish at one time. The activity of the animals and symptoms of poisoning were observed, and waste and dead fish were cleaned up immediately. At the beginning and the end of the test, the body weight and body length of all surviving zebrafish were measured.
Samples were taken after 28 days of exposure, and muscle, liver, and gills of zebrafish were separated with a scalpel and refrigerated at −80 °C. The Ln content in each treatment was determined by microwave digestion and inductively coupled plasma mass spectrometry (ICP-MS, NexION 300X, Waltham, MA, USA). The pretreatment methods for the determination of Ln in fish tissue and solution were as follows: the former weighed 0.1 g of zebrafish liver tissue dried by gauze (accurate to 0.001 g), and 20 mL of the solution and 5 mL of HNO3 were added to the autoclave. The lid was screwed tightly, left for 1 h, and the standard operation procedure of microwave digestion apparatus (APL Touchwin 2.0, Wuhan, China) was followed. After cooling, the lid was slowly opened to vent the air, and the autoclave was placed on a temperature-controlled hot plate to drive out the acid at 140 °C. The digestion solution was transferred to a 10–25 mL volumetric flask, washed three times with a small amount of water, combined in a volumetric flask, and fixed to the scale. The Ln content was determined by ICP-MS. The method was carried out according to the instruction manual of the instrument. The microwave digestion method was based on a previous study [19].
2.4. Toxicity of Mixture
The toxic effect of the mixture (Mix) containing La and Gd was determined from an equimolar concentration combination (Mix = ½ La + ½ Gd, Mix = ¼ La + ¾ Gd, and Mix = ¾ La + ¼ Gd). The concentration and toxicity test methods are described in Section 2.2 and Section 2.3. The 72 h-LC50 of zebrafish in each treatment group and the accumulation of La, Gd, and the Mix were measured at the end of the test.
2.5. Transcriptome High-Throughput Sequencing
Transcriptome analysis was performed on the livers of the zebrafish collected, as described in Section 2.3. We used the rapid freezing method of 2–4 °C proposed by the American Veterinary Medical Association (2013) for zebrafish euthanized. Liver tissue (100 mg) was ground in liquid nitrogen, and total RNA was extracted using an RNA extraction kit (Takara, Bao Biological Engineering Co., Ltd., Dalian, China). After testing the purity and integrity of the extracted RNA, it was reverse transcribed using the Prime Script TM 1st Strand cDNA Synthesis Kit (Takara) to synthesize cDNA. Specific procedures were performed according to the manufacturer’s instructions.
Based on second-generation sequencing technology, the cDNA library was sequenced using the Illumina high-throughput sequencing platform and transformed by the sequencing platform’s own software to generate raw data for FASTQ. Subsequently, the raw data were filtered to remove low-quality and adaptor sequences to obtain clean data, and sequence comparison was performed with the specified zebrafish reference genome to obtain the comparison data. This process was performed at Shanghai Personalbio Technology Co., Ltd. (Shanghai, China).
Differential gene expression analysis was performed using DESeq to identify differentially expressed genes (DEGs) in zebrafish livers after different treatments. The DEGs were entered in the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases using BLAST software, and their annotation information was obtained.
2.6. Quantitative Real-Time Polymerase Chain Reaction
Quantitative real-time polymerase chain reaction (qPCR) was performed using an SYBR Green Mix (20 µL reaction system) amplification system with the following inputs: SYBR (buffer) 10 µL, RF (primer) 2 µL (20 ng), cDNA template (1 µL), and sterile water (7 µL). The PCR amplification process was as follows: predenaturation at 95 °C for 30 s, denaturation at 95 °C for 5 s, annealing at 60 °C for 30 s, annealing at 75 °C for 5 s, 40 cycles, extending at 60 ℃ for 10 min. Fluorescence was measured at the end of the reaction, and real-time amplification and lysis curves were established with three replicates for each treatment. Twelve DEGs (including CDKA;1, CDK2, GSTP2, UGT5a1, ALDH3B1, PCNA1, CYP1A, CDC45, TP53, BRCA1, MRE11, and MDM2) in zebrafish liver were screened using transcriptome high-throughput sequencing. Gene ACT2 (accession number: 821411) was as an internal reference gene, and the DEGs induced by La and Gd were further verified by qPCR analysis. The genes’ names and their primer sequences are listed in Table S3. The Quant Studio™ Design analysis software automatically calculates the relative expression levels of genes using algorithms based on the 2−ΔΔCt method.
2.7. Quality Control and Data Processing
Three biological replicates were performed for all experimental treatments. A multiparameter water quality analyzer (YSI Professional Plus) was used to ensure the stability of the water quality conditions, including pH, temperature, and dissolved oxygen, during the toxicity test. Tukey’s HSD of one-way analysis of variance (ANOVA) was used to analyze significant differences in the control and La- and Gd-exposed bioaccumulation and qRT-PCR validation of DEGs. A value of p < 0.05 was considered to indicate a significant difference. Results are given as mean ± standard deviation (SD). The plots were created using Microsoft Excel 2017 and Origin 2017.
3. Results
3.1. Toxic Effects and Associated Mechanisms of La and Gd on Danio rerio
Table S4 shows that the variation range of Ln concentration was less than 10%, the DO concentration was not less than 80% of the air saturation value (ASV), the temperature did not vary more than 1 °C, and the pH did not vary more than 0.2 during the test. The acute toxicity test showed that the 72 h-LC50 of La and Gd on Danio rerio were 170.73 (161.55–179.56) μmol L−1 and 148.86 (141.96–156.32) μmol L−1, respectively (Figure 1a). Stressed by the high concentrations of REEs, the death of Danio rerio was significantly higher than that observed in the low-concentration and control groups, and the individual action was obviously slow. Compared with La and Gd, the toxicity of the latter on Danio rerio was slightly higher than that of the former. As shown in Figure 1b, the 72 h-LC50 of the Mix on Danio rerio was 158.57 (149.96–166.73) μmol L−1, which is similar to that of a single element, indicating that the Mix had an additive toxic mode.
The survival rate of Danio rerio in each treatment group was greater than 95% when they were stressed with 15 μmol L−1 and 30 μmol L−1 La and Gd for 28 days. The enrichment amounts and bioconcentration factors (BCF) of La and Gd in liver, gills, and muscle tissues are shown in Table 1: liver (7.73–9.33) > gills (3.23–4.49) > muscle (0.17–0.33). These findings indicate that REEs were mainly enriched in the livers of Danio rerio, which also provided the basis for subsequent transcriptome experiments.
3.2. Transcriptome Analysis of Danio rerio under La and Gd Stress
Figure 2 and Table S5 show the transcriptome high-throughput sequencing of Danio rerio stressed by La and Gd. Compared with the control (Figure 2a), La (30 μmol L−1) induced 418 DEGs, of which 189 were upregulated and 229 were downregulated, and Gd (30 μmol L−1) induced 465 DEGs, of which 212 were upregulated and 253 were downregulated. GO classification results showed that, as shown in Figure 2b, multiple genes related to oxidation reduction, DNA replication, cell cycle processes, metabolism of xenobiotics by cytochrome, signaling pathways, extracellular matrix, and response to external biotic stimuli were significantly enriched following exposure to La and Gd. In addition, differentially expressed genes were involved in transport, regulation of transcription, and catalytic activity.
KEGG pathway enrichment analysis was performed for the DEGs in each treatment group. As indicated in Figure 3, most of the DEGs in Danio rerio were enriched in glutathione metabolism, cell cycle, biosynthesis, metabolism of xenobiotics by cytochrome P450, DNA replication, mismatch repair, homologous recombination, p53 signaling pathway, and insulin signaling pathway. Interestingly, we found that DEGs induced by La and Gd share a considerable number of common enrichment pathways. These results suggest that La and Gd may share similar biological mechanisms of action.
3.3. Validation of DEGs Induced by La and Gd
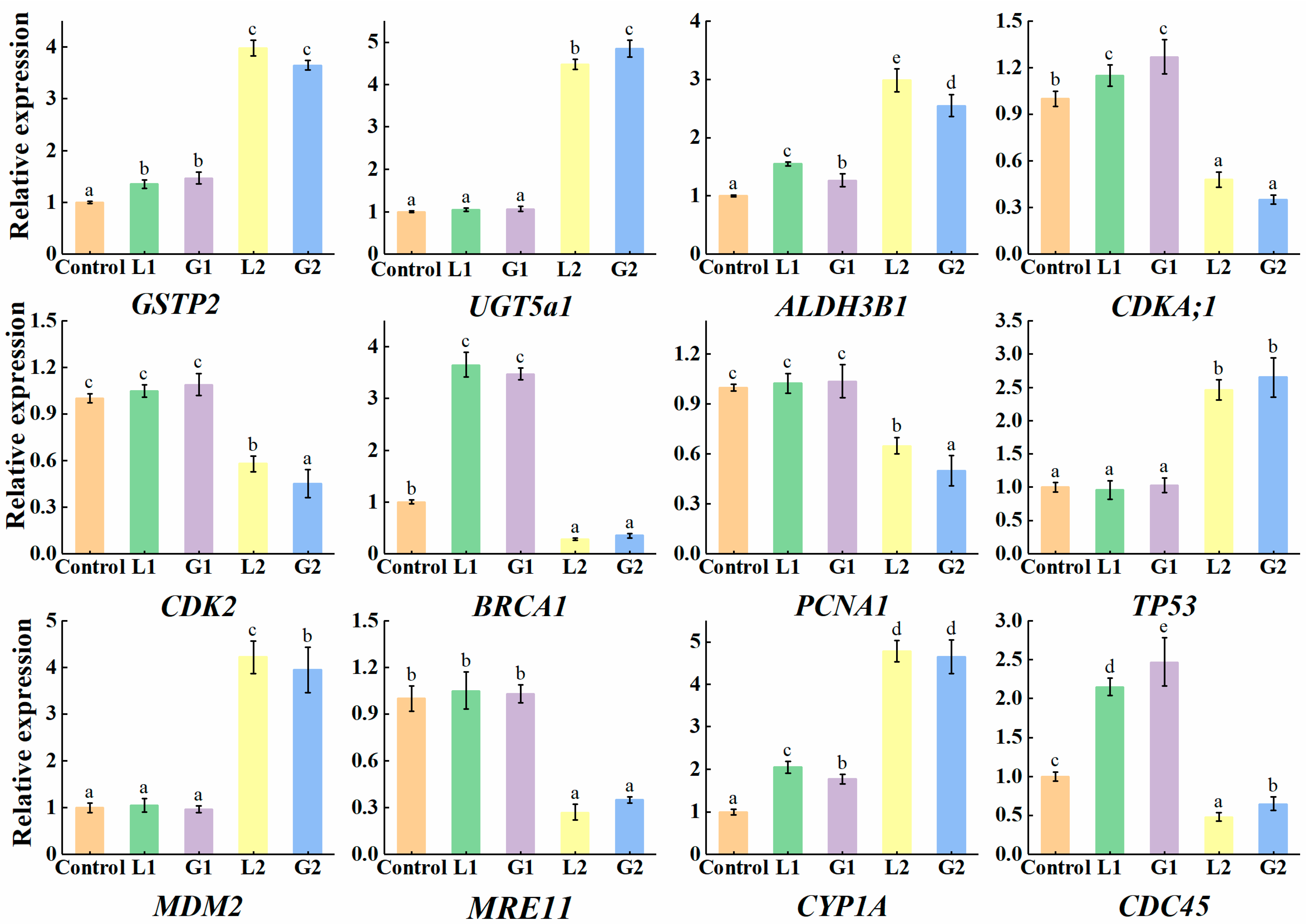
qRT-PCR was used to further verify that La and Gd induced the DEGs of Danio rerio liver. Twelve DEGs were successfully amplified and compared with those in the transcriptome (Figure 4). In the 30 μmol L−1 La treatment group, the relative expression of GSTP2, UGT5a1, ALDH3B1, CYP1A, TP53, and MDM2 showed a significant upregulation trend and were 3.98-, 4.45-, 2.95-, 4.73-, 2.51-, and 4.27-times higher than the control, respectively, while the relative expression levels of CDKA;1, CDK2, BRCA1, PCNA1, MRE11, and CDC45 were significantly downregulated, showing 0.43-, 0.65-, 0.32-, 0.62-, 0.27-, and 0.49-fold reductions, respectively, compared with the control. The changes in these genes in qPCR and transcriptome sequencing were the same, indicating that the transcriptome sequencing analysis was reliable. Furthermore, compared with La and Gd, there was no significant difference in the change trends of the 12 DEGs, with only a slight difference in the amount of expression (Gd treatment induced a slightly higher number of DEGs than La treatment).
4. Discussion
REEs are metals located in the sixth period of the periodic table. According to previous studies, no significant physiological transformation processes associated with REEs have been found in animals; therefore, REEs are not considered essential life elements. Long-term exposure to or consumption of foods containing high levels of REEs may have biological effects. In the present study, the highest enrichment of La and Gd was found in the Danio rerio liver, which was 30–60 times higher than that in muscle. Similar results have also been reported in other studies. Tu [31] studied the accumulation of REEs in different organs of carp and found that the bioenrichment in carp was in the following order: liver > gills > bone > muscle. Bustamante [32] found that invertebrates have the highest enrichment of REEs in their digestive glands, followed by the gills, gonads, kidneys, and muscles. Therefore, the liver of Danio rerio was used as the target organ in this study to explore the toxic effects of the selected two REEs.
Whether REEs are genotoxic remains controversial. Generally, REEs have a significant hormesis effect: low doses promote biological growth, whereas high doses have a negative effect [33]. The growth-promoting effect of REEs is related to the promotion of cell proliferation, similar to that of calcium (Ca) [34]. At high doses, REEs cause various types of base damage, DNA breakage, mutation, and carcinogenesis [35]. Ca plays an important role in cell growth and proliferation [36,37]. REEs and Ca have the same ion radius; therefore, REEs can act as substitutes for extracellular or intracellular Ca, resulting in the disruption of Ca homeostasis and leading to disorders of Ca-related biological processes [37,38]. From the findings of the present study, La and Gd can be postulated to interfere with the normal operation of physiological activities involved in cytoplasmic Ca, thus interfering with the expression of biomolecules, including cyclin B, insulin growth factor, and cyclin-dependent kinase, further affecting the progression of cell meiosis.
Environmental stress can cause various forms of DNA damage. To maintain the integrity of the genome, organisms can activate the activity of DNA repair complexes through signal cascade reactions, regulate the process of the cell cycle, or start the process of cell death [39]. The expression of cell cycle regulatory genes (such as BRCA1, MRE11, and TP53) are regulated by ataxia telangiectasia mutated (ATM)/ataxia telangiectasia and Rad3 (ATR) related dependent signal transduction [40]. They play an important role in monitoring genomic integrity and DNA damage repair [41]. In this study, when La and Gd concentrations were 15 μmol L−1, the expression of BRCA1 in the liver of Danio rerio increased by 3.67 times compared with the control. The increased expression of genes related to the cell cycle and damage repair reduced the damage in the liver of Danio rerio. When La and Gd concentrations were 30 μmol L−1, the expression of BRCA1 decreased significantly, which indicated that a high concentration of La induced DNA damage. Similarly, the expression of MRE11, which is related to the terminal repair of DNA double-strand breaks (DSB) [42], decreased significantly in the 30 μmol L−1 La and Gd treatment groups. These results showed that under high concentrations of REEs treatments, the expression of damage repair-related genes decreased, and the dysfunctional repair system increased DNA damage in the liver of Danio rerio. At the same time, the increased expression of TP53 and MDM2 indicated that the cells might undergo apoptosis.
Herein, several processes related to exogenous substance metabolism (such as the metabolism of xenobiotics by cytochrome P450 and glutathione metabolism) were significantly enriched following exposure to La and Gd. Several important enzyme-encoding genes involved in the metabolism of exogenous substances, such as GSTP2, UGT5a1, and Aldh3b1, were significantly upregulated following exposure to La and Gd. Glutathione is an indispensable substrate for the catalytic detoxification of glutathione sulfotransferase [43,44]. Acetaldehyde dehydrogenase, the expression product of the Aldh3b1 gene, is also an important enzyme for cellular detoxification, especially for cytotoxicity caused by oxidants [45]. When an organism is under adverse stress or disturbed by foreign substances, the organism initiates relevant mechanisms for the repair or maintenance of homeostasis, and a series of detoxifying metabolic enzymes are activated, and their expression levels are elevated to increase cellular tolerance [46]. In the present study, the observation of enriched detoxification metabolic pathways and the upregulated expression levels of some key metabolic enzyme genes suggest that exposure to these two REEs stimulated defense mechanisms in the zebrafish. The results of GO annotation confirmed the aforementioned postulation, with the response to external biotic stimulus and the extracellular matrix significantly enriched in both the La and Gd treatment groups.
Whether there is a common toxicity mechanism or modes of action among REEs has always been a question. Because they have the same number of electronic layers and outer electronic structure, all REEs possess very similar chemical properties and are, thus, regarded as a group of elements with uniform chemical properties [11,47]. There has been speculation about the homogeneity of REEs; however, there are not enough comparable data to reach a consistent conclusion [21]. Previous acute toxicity experiments showed that there was no significant difference in lethality between the two elements. However, Gd may cause a stronger molecular response to some biological processes and induce more DEGs than La (Figure 4). Similarly, an experiment on sea urchin embryos showed that the levels of reactive oxygen species and malondialdehyde were higher following Ce than La exposure [48]. However, of the two, Ce did not have a stronger effect on most biological processes. In a study of two sea urchin species, La caused damage to embryos, whereas Ce did not [10]. Herein, both La and Gd had similar lethality; however, a comparison of their toxicity patterns needs to be based on the investigated species, toxicological endpoints, and physiological processes.
In conclusion, in this study, there were numerous DEGs and biological pathways co-enriched by La and Gd, while only a few biological processes were affected by one element. The expression of genes related to cell division, exogenous substance metabolism, and some basic physiological activities, such as DNA replication and the cell cycle, differed slightly between the two treatments; however, the complete physiological process was subjected to the same interference under La and Gd stress. Taken together, La and Gd may affect some biological processes with different intensities; nonetheless, there are many similarities in their overall toxicity mechanisms and modes of action. The current data help clarify the potential toxicity and modes of action of REEs, which are not yet fully recognized, thereby contributing to their environmental risk assessment.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/toxics10090519/s1, Table S1: Molarity of each ion in the standard water; Table S2: Ammonia concentration (mg/L) measurements during 72-h acute toxicity; Table S3: Primer sequences used in the experiment; Table S4: Analysis of the water quality of the solution during the toxicity test; Table S5: Gene ontology and gene symbol in zebra fish livers induced by La and Gd. References [49,50,51,52] are cited in the supplementary materials.
Author Contributions
Conceptualization, L.S. and C.G.; methodology, S.K.; software, C.X.; validation, S.K., C.G. and L.S.; formal analysis, C.M.; investigation, S.K.; resources, L.S.; data curation, S.K.; writing—original draft preparation, S.K. and C.G.; writing—review and editing, L.S.; supervision, H.M.; funding acquisition, L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially sponsored by the National Key Research and Development Program of China (2018YFC1800703, 2016YFD0800305) and the National Natural Science Foundation of China (41877146, 31600372).
Institutional Review Board Statement
This study was approved by LABORATORY ANIMAL CENTER OF SHENYANG PHARMACEUTICAL UNIVERSITY (Approval Code: SYPU-IVCUC-2022-0303-001; Approval Date: 3.2022—12.2027) and has been performed in accordance with the ethical guidelines of the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.
Informed Consent Statement
The study did not involve humans.
Data Availability Statement
Data can be made available upon reasonable request.
Acknowledgments
We thank the National Natural Science Foundation of China for the financial support and Shenyang Pharmaceutical University for the help in animal toxicology experiments.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Migaszewski, Z.M.; Galuszka, A. The characteristics, occurrence, and geochemical behavior of rare earth elements in the environment: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 429–471. [Google Scholar] [CrossRef]
- Sun, L.; Dong, H.; Zhang, P.; Yan, C. Up conversion of rare earth nanomaterials. Annu. Rev. Phys. Chem. 2015, 66, 619–642. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Chang, L.; Finkelman, R.B.; Wang, W.; Liu, J.; Li, J.; Xing, J.; Hou, C. Distribution of rare earth elements in PM10 emitted from burning coals and soil-mixed coal briquettes. J. Environ. Sci. 2020, 97, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Li, Z.; Liu, T.; Chen, J.; Wu, T.; Feng, X. Rare earth elements in street dust and associated health risk in a municipal industrial base of central China. Environ. Geochem. Health. 2017, 39, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Justyna, R.; Ewa, O.; Wojciech, R.; Lidia, W. Gadolinium as a New Emerging Contaminant of Aquatic Environments. Environ. Toxicol. Chem. 2018, 37, 1523–1534. [Google Scholar] [CrossRef]
- Pagano, G.; Guida, M.; Tommasi, F.; Oral, R. Health effects and toxicity mechanisms of rare earth elements-knowledge gaps and research prospects. Ecotoxicolo. Environ. Saf. 2015, 115, 40–48. [Google Scholar] [CrossRef]
- Willis, G.; Lynda, M.; Concilia, D.; Nhamo, C.; Nothando, D.; Edmond, S. Sources, behaviour, and environmental and human health risks of hightechnology rare earth elements as emerging contaminants. Sci. Total Environ. 2018, 636, 299–313. [Google Scholar] [CrossRef]
- Li, X.F.; Che, Z.B.; Chen, Z.Q.; Zhang, Y. A human health risk assessment of rare earth elements in soil and vegetables from a mining area in Fujian Province, Southeast China. Chemosphere 2013, 93, 1240–1246. [Google Scholar] [CrossRef]
- Herrmann, H.; Nolde, J.; Berger, S.; Heise, S. Aquatic ecotoxicity of lanthanum: A review and anattempt to derive water and sediment quality criteria. Ecotoxicol. Environ. Saf. 2016, 124, 213–238. [Google Scholar] [CrossRef]
- Marco, T.; Giovanni, P.; Marco, G.; Anna, P.; Antonietta, S.; Maria, G.; Daniel, M.L.; Petra, B.; Maja, L.; Philippe, J.T.; et al. Comparative toxicity of seven rare earth elements in sea urchin early life stages. Environ. Sci. Pollut. Res. 2017, 24, 20803–20810. [Google Scholar] [CrossRef] [Green Version]
- Dahle, J.T.; Arai, Y. Environmental geochemistry of cerium: Applications and toxicology of cerium oxide nanoparticles. Int. J. Environ. Res. Public Health 2015, 12, 1253–1278. [Google Scholar] [CrossRef] [PubMed]
- Giovanni, P.; Philippe, J.; Aldo, D.; Macro, T. Human exposures to rare earth elements: Present knowledge and research prospects. Environ. Res. 2019, 171, 493–500. [Google Scholar] [CrossRef]
- Kulaksız, S.; Bau, M. Rare earth elements in the Rhine River, Germany: First case of anthropogenic lanthanum as a dissolved microcontaminant in the hydrosphere. Environ. Int. 2011, 37, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.G. Detection of anthropogenic gadolinium in the Brisbane River plume in Moreton Bay, Queensland, Australia. Mar. Pollut. Bull. 2010, 60, 1113–1116. [Google Scholar] [CrossRef]
- Jacinto, J.; Henriques, B.; Duarte, A.C.; Vale, C.; Pereira, E. Removal and recovery of Critical Rare Elements from contaminated waters by living Gracilaria gracilis. J. Hazard. Mater. 2018, 344, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.B.; Liang, J.H.; Meng, H.Y.; Yin, Y.; Zhang, K. Rare Earth Elements Lanthanum and Praseodymium Adversely Affect Neural and Cardiovascular Development in Zebrafish (Danio rerio). Environ. Sci. Technol. 2021, 55, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Rim, K.T. Effects of rare earth elements on the environment and human health: A literature review. Toxicol. Environ. Health Sci. 2016, 8, 189–200. [Google Scholar] [CrossRef]
- Carpenter, D.; Boutin, C.; Allison, J.E.; Parsons, J.L.; Ellis, D.M. Uptake and effects of six rare earth elements (rees) on selected native and crop species growing in contaminated soils. PLoS ONE 2015, 10, e0129936. [Google Scholar] [CrossRef]
- Liu, Z.H.; Guo, C.; Tai, P.D.; Sun, L.Z.; Chen, Z.B. The exposure of gadolinium at environmental relevant levels induced genotoxic effects in Arabidopsis thaliana (L.). Ecotoxicol. Environ. Saf. 2021, 215, 112–138. [Google Scholar] [CrossRef]
- De Oliveira, C.; Ramos, S.J.; Siqueira, J.O.; Faquin, V.; De Castro, E.M.; Amaral, D.C.; Techio, V.H.; Coelho, L.C.; ESilva, P.H.; Schnug, E.; et al. Bioaccumulation and effects of lanthanum on growth and mitotic index in soybean plants. Ecotoxicological Environ. Saf. 2015, 122, 136–144. [Google Scholar] [CrossRef]
- Gonzalez, V.; Vignati, D.A.L.; Leyval, C.; Giamberini, L. Environmental fate and ecotoxicity of lanthanides: Are they a uniform group beyond chemistry? Environ. Int. 2014, 71, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.D.; Zhao, Q.; Su, D.; Li, P.J.; Stagnitti, F. Biological toxicity of lanthanide elements on algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef]
- Blinova, I.; Lukjanova, A.; Muna, M.; Vija, H.; Kahru, A. Evaluation of the potential hazard of lanthanides to freshwater microcrustaceans. Sci. Total Environ. 2018, 642, 1100–1107. [Google Scholar] [CrossRef]
- Romero-Freire, A.; Joonas, E.; Muna, M.; Cossu-Leguille, C.; Vignati, D.A.L.; Giamberini, L. Assessment of the toxic effects of mixtures of three lanthanides (Ce, Gd, Lu) to aquatic biota. Sci. Total Environ. 2019, 661, 276–284. [Google Scholar] [CrossRef]
- Romero-Freire, A.; Minguez, L.; Pelletier, M.; Cayer, A.; Caillet, C.; Devin, S.; Gross, E.M.; Guérold, F.; Pain-Devin, S.; Vignati, D.A.L.; et al. Assessment of baseline ecotoxicity of sediments from a prospective mining area enriched in light rare earth elements. Sci. Total Environ. 2018, 612, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Bing, G.; Erkai, H.; Hao, Q.; Jianqiu, L.; Jie, J.; Ling, Z.; Xinde, C. Phytotoxicity of individual and binary mixtures of rare earth elements (Y, La, and Ce) in relation to bioavailability. Environ. Pollut. 2019, 246, 114–121. [Google Scholar] [CrossRef]
- Yang, G.; Kevin, J.W. Bio-uptake of a rare earth metal (Nd) by Chlamydomonas reinhardtii bioavailability of small organic complexes and role of hardness ions. Environ. Pollut. 2018, 243, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Webster, T.M.U.; Bury, N.; Van, A.R.; Santos, E.M. Global transcriptome profiling reveals molecular mechanisms of metal tolerance in a chronically exposed wild population of Brown Trout. Environ. Sci. Technol. 2013, 47, 8869–8877. [Google Scholar] [CrossRef]
- Loughery, J.R.; Marentette, J.R.; Frank, R.A.; Hewitt, L.M.; Martyniuk, C.J. Transcriptome profiling in larval fathead minnow exposed to commercial naphthenic acids and extracts from fresh and aged oil sands process-affected water. Environ. Sci. Technol. 2019, 53, 10435–10444. [Google Scholar] [CrossRef]
- Fuertes, I.; Campos, B.; Rivetti, C.; Pia, B.; Barata, C. Effects of single and combined low concentrations of neuroactive drugs on Daphnia magna reproduction and transcriptomic Responses. Environ. Sci. Technol. 2019, 53, 11979–11987. [Google Scholar] [CrossRef]
- Tu, Q.; Wang, X.R.; Tian, L.Q.; Dai, L.M. Bioaccumulation of the rare earth elements lanthanum, gadolinium and yttrium in carp (Cyprinus carpio). Environ. Pollut. 1994, 85, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, P.; Miramand, P. Subcellular and body distributions of 17 trace elements in the variegated scallop Chlamys varia from the French coast of the Bay of Biscay. Sci. Total Environ. 2005, 337, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.J.; Carpenter, D.; Boutin, C.; Allison, J.E. Rare earth elements (REEs): Effects on germination and growth of selected crop and native plant species. Chemosphere 2014, 96, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, M.; Chu, Y.; Chen, D.D.Y.; Huang, X.; Zhou, Q. Responses of plant calmodulin to endocytosis induced by rare earth elements. Chemosphere 2016, 154, 408–415. [Google Scholar] [CrossRef]
- Cheng, J.; Li, N.; Cheng, Z.; Hua, R.; Cai, J.; Si, W.; Hong, F. Splenocyte apopotic pathway in mice following exposure to cerium chloride. Chemosphere 2011, 83, 612–617. [Google Scholar] [CrossRef]
- Silvestre, F.; Boni, R.; Fissore, R.A.; Tosti, E. Ca2+ signaling during maturation of cumulus-oocyte complex in mammals. Mol. Reprod. Dev. 2011, 78, 744–756. [Google Scholar] [CrossRef]
- Boni, R.; Gualtieri, R.; Talevi, R.; Tosti, E. Calcium and other ion dynamics during gamete maturation and fertilization. Theriogenology 2007, 68, 156–164. [Google Scholar] [CrossRef]
- Martin, B.; Richardson, F.S. Lanthanides as probes for calcium in biological systems. Q. Rev. Biophys. 1979, 12, 181–209. [Google Scholar] [CrossRef]
- Hu, Z.; Cools, T.; De Veylder, L. Mechanisms used by plants to cope with DNA damage. Annu. Rev. Plant Biol. 2016, 67, 439–462. [Google Scholar] [CrossRef]
- Lario, S.; Casalots, A.; Sanfeliu, E.; Boix, L.; García-Iglesias, P.; Sánchez-Delgado, J.; Montserrat, A. Real-time PCR improves helicobacter pylori detection in patients with peptic ulcer bleeding. PLoS ONE 2011, 6, e20009. [Google Scholar] [CrossRef] [Green Version]
- Cools, T.; Veylder, D.L. DNA stress checkpoint control and plant development. Curr. Opin. Plant Biol. 2009, 12, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Minamisawa, K.; Okushima, Y.; Inagaki, S.; Yoshiyama, K.; Kondou, Y.; Kaminuma, E.; Kawashima, M.; Toyoda, T.; Matsui, M.; et al. Programmed induction of endoreduplication by DNA double-strand breaks in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 10004–10009. [Google Scholar] [CrossRef]
- Glisic, B.; Mihaljevic, I.; Popovic, M.; Zaja, R.; Loncar, J.; Fent, K.; Kovacevic, R.; Smital, T. Characterization of glutathione-S-transferases in zebrafish (Danio rerio). Aquat. Toxicol. 2015, 158, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, G.D.; Bridge, W.J. The glutathione system and the related thiol network in Caenorhabditis elegans. Redox Biol. 2019, 24, 101171. [Google Scholar] [CrossRef] [PubMed]
- Marchitti, S.A.; Brocker, C.; Orlicky, D.J.; Vasiliou, V. Molecular characterization, expression analysis, and role of ALDH3B1 in the cellular protection against oxidative stress. Free Radic. Biol. Med. 2010, 49, 1432–1443. [Google Scholar] [CrossRef]
- Li, Z.L.; Liu, Z.H.; Chen, R.J.; Li, X.J.; Tai, P.D.; Gong, Z.Q.; Jia, C.Y.; Liu, W. DNA damage and genetic methylation changes caused by cd in Arabidopsis thaliana seedlings. Environ. Toxicol. Chem. 2015, 34, 2095–2103. [Google Scholar] [CrossRef]
- Banin, A.; Navrot, J. Origin of life: Clues from relations between chemical compositions of living organisms and natural environments. Science 1975, 189, 550–551. [Google Scholar] [CrossRef]
- Pagano, G.; Guida, M.; Siciliano, A.; Oral, R.; Kocbas, F.; Palumbo, A.; Castellano, I.; Migliaccio, O.; Thomas, P.J.; Trifuoggi, M. Comparative toxicities of selected rare earth elements: Sea urchin embryogenesis and fertilization damage with redox and cytogenetic effects. Environ. Res. 2016, 147, 453–460. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Tai, P.; Sun, L.; Yang, X. Toxicity of ammonia, cadmium, and nitrobenzene to four local fishes in the Liao River, China and the derivation of site-specific water quality criteria. Ecotoxicol. Environ. Saf. 2017, 147, 656–663. [Google Scholar] [CrossRef]
- Liu, Z.; Tai, P.; Li, X.; Kong, L.; Matthews, T.G.; Lester, R.E.; Mondon, J.A. Deriving site-specific water quality criteria for ammonia from national versus international toxicity data. Ecotoxicol. Environ. Saf. 2019, 171, 665–676. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Ahmad, S.; Sun, M.; Zhang, Q. Selection of optimal reference genes for qRT-PCR analysis of shoot development and graviresponse in prostrate and erect chrysanthemums. PLoS ONE 2019, 14, e0225241. [Google Scholar] [CrossRef]
- Louren, A.P.; Mackert, A.; Cristino, A. Validation of reference genes for gene expression studies in the honey bee, apis mellifera, by quantitative real-time rt-pcr. Apidologie 2008, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Toxic effects of La, Gd, and the Mix on Danio rerio: (a) acute toxicity (72 h-LC50) of La and Gd on Danio rerio; (b) acute toxicity of the mixture (Mix) containing La and Gd determined from an equimolar concentration combination.
Figure 1.
Toxic effects of La, Gd, and the Mix on Danio rerio: (a) acute toxicity (72 h-LC50) of La and Gd on Danio rerio; (b) acute toxicity of the mixture (Mix) containing La and Gd determined from an equimolar concentration combination.
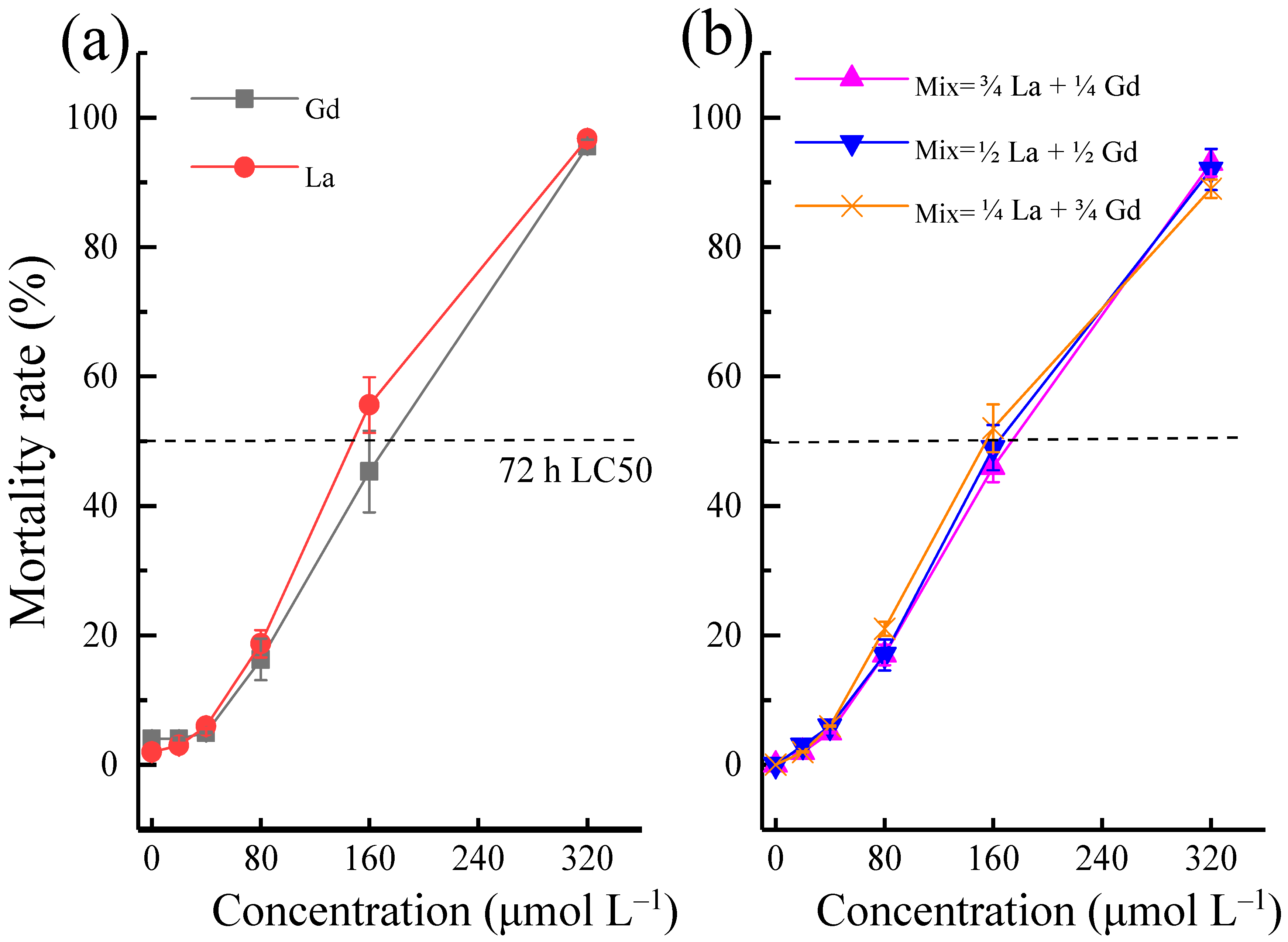
Figure 2.
Transcriptome high-throughput sequencing analysis in liver tissues of Danio rerio exposed to 30 μmol L−1 La and Gd: (a,b) volcano plots of DEGs in La and Gd, compared with the control, respectively. The red dots indicate upregulated genes, blue dots indicate downregulated genes, and gray dots indicate nonsignificant DEGs; (c) analysis of the shared GO classification of DEGs between La and Gd.
Figure 2.
Transcriptome high-throughput sequencing analysis in liver tissues of Danio rerio exposed to 30 μmol L−1 La and Gd: (a,b) volcano plots of DEGs in La and Gd, compared with the control, respectively. The red dots indicate upregulated genes, blue dots indicate downregulated genes, and gray dots indicate nonsignificant DEGs; (c) analysis of the shared GO classification of DEGs between La and Gd.

Figure 3.
Analysis of the shared differentially expressed genes between La and Gd: (a) Venn diagrams showing the differentially expressed genes shared in response to La and Gd exposure; (b) top 10 significantly enriched KEGG pathways shared in response to La exposure. √ Indicates the pathways involved in Gd exposure.
Figure 3.
Analysis of the shared differentially expressed genes between La and Gd: (a) Venn diagrams showing the differentially expressed genes shared in response to La and Gd exposure; (b) top 10 significantly enriched KEGG pathways shared in response to La exposure. √ Indicates the pathways involved in Gd exposure.
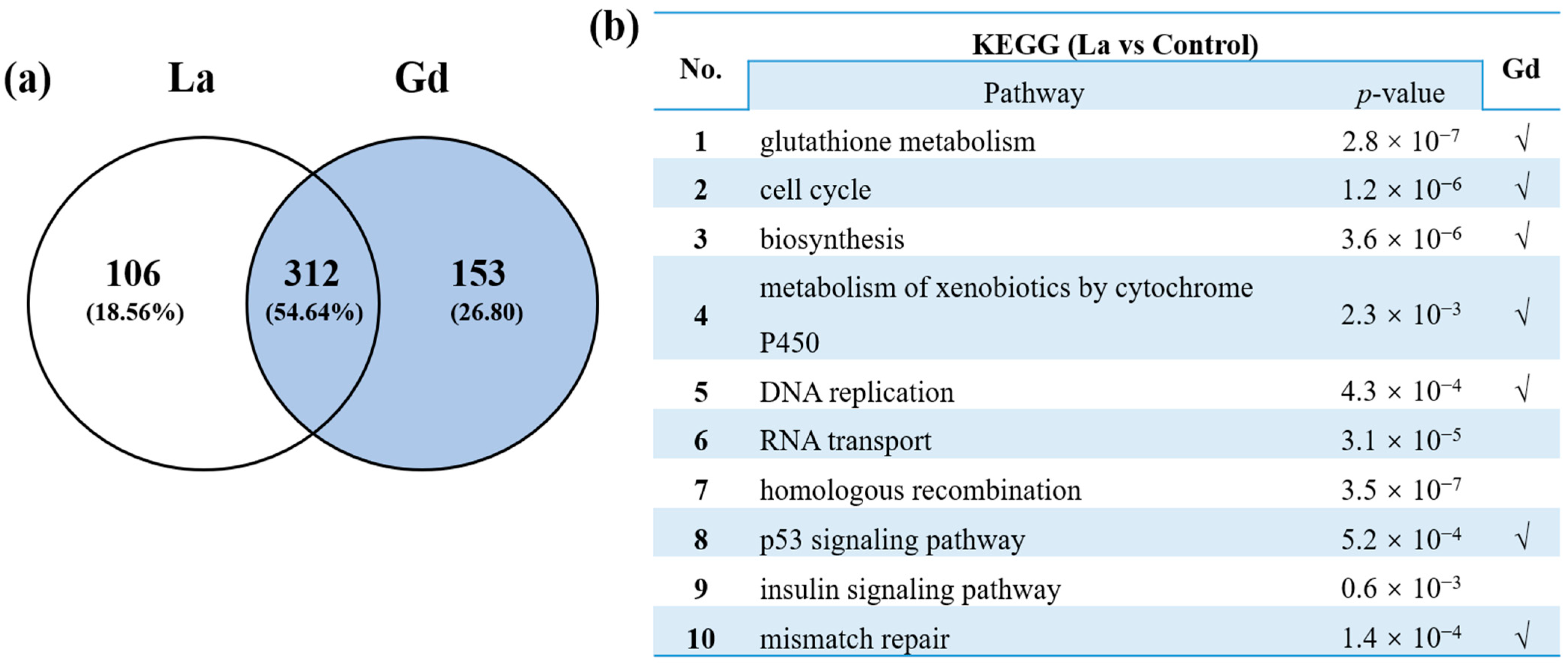
Figure 4.
Differential expression in liver tissues of Danio rerio exposed to La and Gd: L1, 15μmol L−1 La; L2, 30 μmol L−1 La; G1, 15μmol L−1 Gd; and G2, 30 μmol L−1 Gd. Data are shown as mean ± SD by qPCR. The expression of the control was set as 1. Data presented are the average of three replicates. Housekeeping gene ACT2 was used as an internal control, the same below. Different letters indicate significant differences in gene expression.
Figure 4.
Differential expression in liver tissues of Danio rerio exposed to La and Gd: L1, 15μmol L−1 La; L2, 30 μmol L−1 La; G1, 15μmol L−1 Gd; and G2, 30 μmol L−1 Gd. Data are shown as mean ± SD by qPCR. The expression of the control was set as 1. Data presented are the average of three replicates. Housekeeping gene ACT2 was used as an internal control, the same below. Different letters indicate significant differences in gene expression.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
La and Gd uptake and bioconcentration factors (BCF) in muscle, gills, and liver tissues of Danio rerio exposed to 15 and 30 μmol L−1 La and Gd for 28 days, respectively.
Table 1.
La and Gd uptake and bioconcentration factors (BCF) in muscle, gills, and liver tissues of Danio rerio exposed to 15 and 30 μmol L−1 La and Gd for 28 days, respectively.
| Concentration (μmol L−1) | Uptake (μg g−1, Fresh Weight) (BCF) | |||
|---|---|---|---|---|
| Muscle | Gills | Liver | ||
| 15 | La | 0.53 ± 0.14 b (0.25) | 9.36 ± 1.56 b (4.49) | 19.45 ± 3.21 b (9.33) |
| Gd | 0.41 ± 0.09 a (0.17) | 8.43 ± 1.22 a (3.58) | 18.21 ± 3.68 a (7.73) | |
| 30 | La | 1.38 ± 0.26 c (0.33) | 20.35 ± 3.72 d (4.88) | 35.41 ± 8.94 c (8.49) |
| Gd | 1.22 ± 0.35 c (0.26) | 15.21 ± 2.54 c (3.23) | 39.85 ± 7.62 d (8.46) | |
Mean values ± standard deviation are shown (n = 3); different letters next to the values indicate a significant difference (p < 0.05) among the different treatment groups, the same below: BCF, REEs concentration in muscle, gills, or liver/in solution.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kang, S.; Guo, C.; Xue, C.; Ma, C.; Mu, H.; Sun, L. Toxic Effects of Two Representative Rare Earth Elements (La and Gd) on Danio rerio Based on Transcriptome Analysis. Toxics 2022, 10, 519. https://doi.org/10.3390/toxics10090519
AMA Style
Kang S, Guo C, Xue C, Ma C, Mu H, Sun L. Toxic Effects of Two Representative Rare Earth Elements (La and Gd) on Danio rerio Based on Transcriptome Analysis. Toxics. 2022; 10(9):519. https://doi.org/10.3390/toxics10090519
Chicago/Turabian StyleKang, Shu, Cheng Guo, Chenyang Xue, Chenshu Ma, Huaizhong Mu, and Lizong Sun. 2022. "Toxic Effects of Two Representative Rare Earth Elements (La and Gd) on Danio rerio Based on Transcriptome Analysis" Toxics 10, no. 9: 519. https://doi.org/10.3390/toxics10090519
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.