

Impact of Electronic Cigarettes, Heated Tobacco Products and Conventional Cigarettes on the Generation of Oxidative Stress and Genetic and Epigenetic Lesions in Human Bronchial Epithelial BEAS-2B Cells

 , , ,
, , , 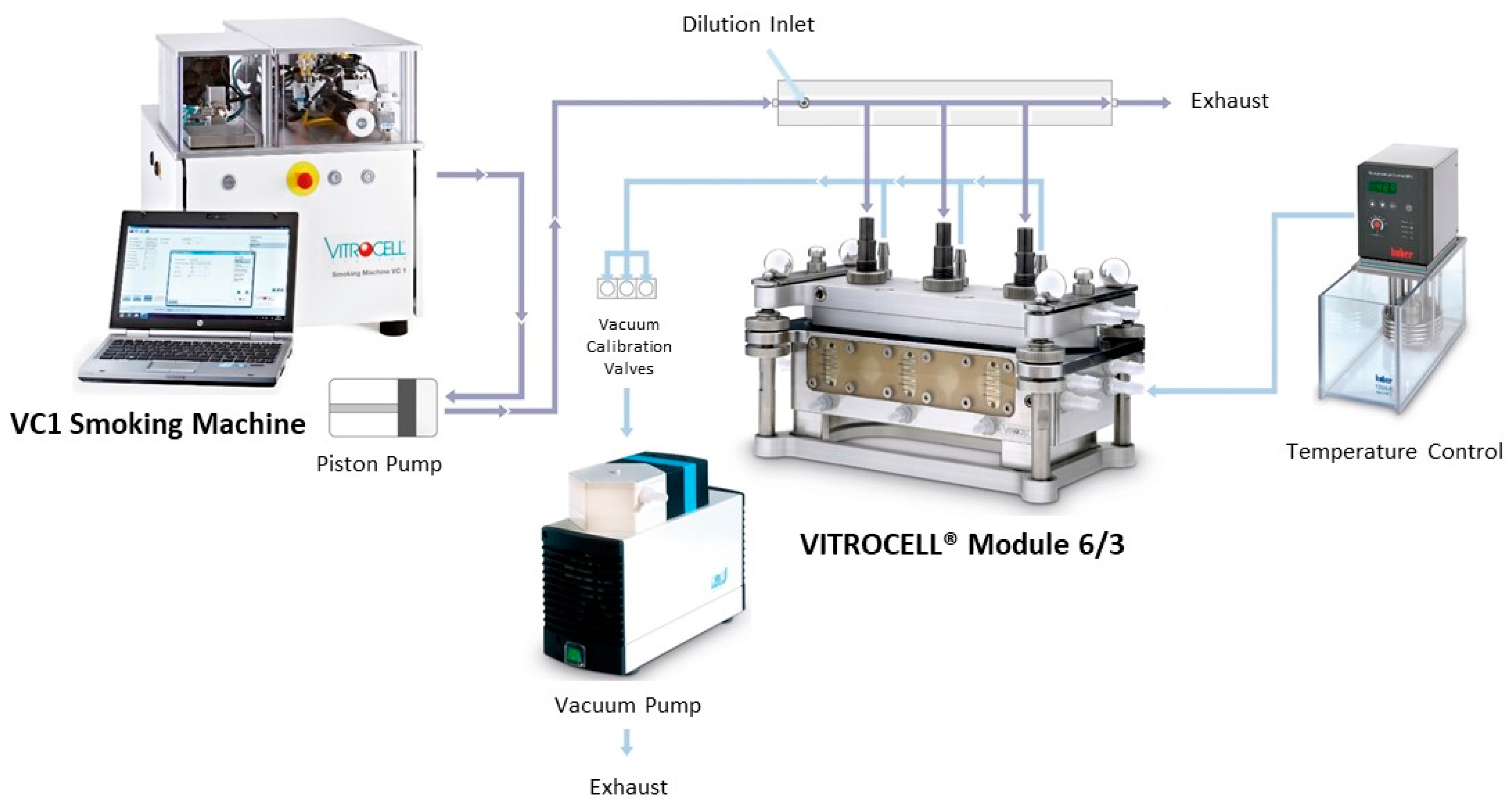{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. E-Cig and Tobacco Products
2.2. Cell Culture
2.3. Aerosol Generation and Cell Exposure
2.4. Intracellular ATP Assay
2.5. Measure of the Nuclear Activity of the Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2)
2.6. Gene Expression Analysis
2.7. Western Blot Analyses
2.8. Comet Assay
2.9. Micronucleus Test
2.10. Global DNA Methylation
2.11. ELISA Assay to Detect Histone Modifications
2.12. Statistical Analyses
3. Results
3.1. Evaluation of the Cytotoxicity after Exposure to E-Cig or Tobacco Product Emissions
3.2. Study of Oxidative Stress after Exposure to E-Cig or Tobacco Product Emissions
3.3. Assessment of Genotoxicity and Mutagenicity after Exposure to E-Cig or Tobacco Product Emissions
3.4. Study of Impact of E-Cig or Tobacco Product Emissions in Global DNA Methylation and Histone Modulations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALI | air–liquid interface |
| ARE | antioxidant response element |
| CS | cigarette smoke |
| EC50 | dose decreasing to 50% cell viability |
| e-cig | electronic cigarettes |
| HTP | heated tobacco products |
| Mb-18W | Modbox e-cig model set at 18 W |
| Mb-30W | Modbox e-cig model set at 30 W |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| PAHs | polycyclic aromatic hydrocarbons |
References
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of Lung Cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Soleimani, F.; Dobaradaran, S.; De-la-Torre, G.E.; Schmidt, T.C.; Saeedi, R. Content of Toxic Components of Cigarette, Cigarette Smoke vs Cigarette Butts: A Comprehensive Systematic Review. Sci. Total Environ. 2022, 813, 152667. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.; Stevenson, M.; Wieczorek, R.; Pani, J.; Trelles-Sticken, E.; Dethloff, O.; Czekala, L.; Simms, L.; Buchanan, F.; O’Connell, G.; et al. Chemical Composition and In Vitro Toxicity Profile of a Pod-Based E-Cigarette Aerosol Compared to Cigarette Smoke. Appl. In Vitro Toxicol. 2020, 6, 11–41. [Google Scholar] [CrossRef]
- Vivarelli, F.; Granata, S.; Rullo, L.; Mussoni, M.; Candeletti, S.; Romualdi, P.; Fimognari, C.; Cruz-Chamorro, I.; Carrillo-Vico, A.; Paolini, M.; et al. On the Toxicity of E-Cigarettes Consumption: Focus on Pathological Cellular Mechanisms. Pharmacol. Res. 2022, 182, 106315. [Google Scholar] [CrossRef]
- Esteban-Lopez, M.; Perry, M.D.; Garbinski, L.D.; Manevski, M.; Andre, M.; Ceyhan, Y.; Caobi, A.; Paul, P.; Lau, L.S.; Ramelow, J.; et al. Health Effects and Known Pathology Associated with the Use of E-Cigarettes. Toxicol. Rep. 2022, 9, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Chen, J.; Liu, P.; Li, M. A Review of Toxicity Mechanism Studies of Electronic Cigarettes on Respiratory System. Int. J. Mol. Sci. 2022, 23, 5030. [Google Scholar] [CrossRef]
- Smith, M.R.; Clark, B.; Lüdicke, F.; Schaller, J.-P.; Vanscheeuwijck, P.; Hoeng, J.; Peitsch, M.C. Evaluation of the Tobacco Heating System 2.2. Part 1: Description of the System and the Scientific Assessment Program. Regul. Toxicol. Pharmacol. 2016, 81 (Suppl. 2), S17–S26. [Google Scholar] [CrossRef]
- Kim, S.C.; Friedman, T.C. A New Ingenious Enemy: Heat-Not-Burn Products. Tob. Use Insights 2022, 15, 1179173X221076419. [Google Scholar] [CrossRef]
- Znyk, M.; Jurewicz, J.; Kaleta, D. Exposure to Heated Tobacco Products and Adverse Health Effects, a Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 6651. [Google Scholar] [CrossRef]
- Bravo-Gutiérrez, O.A.; Falfán-Valencia, R.; Ramírez-Venegas, A.; Sansores, R.H.; Ponciano-Rodríguez, G.; Pérez-Rubio, G. Lung Damage Caused by Heated Tobacco Products and Electronic Nicotine Delivery Systems: A Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 4079. [Google Scholar] [CrossRef]
- Schaller, J.-P.; Keller, D.; Poget, L.; Pratte, P.; Kaelin, E.; McHugh, D.; Cudazzo, G.; Smart, D.; Tricker, A.R.; Gautier, L.; et al. Evaluation of the Tobacco Heating System 2.2. Part 2: Chemical Composition, Genotoxicity, Cytotoxicity, and Physical Properties of the Aerosol. Regul. Toxicol. Pharmacol. 2016, 81 (Suppl. 2), S27–S47. [Google Scholar] [CrossRef] [PubMed]
- Dusautoir, R.; Zarcone, G.; Verriele, M.; Garçon, G.; Fronval, I.; Beauval, N.; Allorge, D.; Riffault, V.; Locoge, N.; Lo-Guidice, J.-M.; et al. Comparison of the Chemical Composition of Aerosols from Heated Tobacco Products, Electronic Cigarettes and Tobacco Cigarettes and Their Toxic Impacts on the Human Bronchial Epithelial BEAS-2B Cells. J. Hazard. Mater. 2021, 401, 123417. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, Y.; Jiang, X.; Zhang, H.; Zhu, F.; Hu, S.; Hou, H.; Hu, Q.; Pang, Y. Chemical Analysis and Simulated Pyrolysis of Tobacco Heating System 2.2 Compared to Conventional Cigarettes. Nicotine Tob. Res. 2019, 21, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Nohmi, T. Thresholds of Genotoxic and Non-Genotoxic Carcinogens. Toxicol. Res. 2018, 34, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.-S.; Lee, H.-W.; Weng, M.-W.; Wang, H.-T.; Hu, Y.; Chen, L.-C.; Park, S.-H.; Chan, H.-W.; Xu, J.; Wu, X.-R.; et al. DNA Damage, DNA Repair and Carcinogenicity: Tobacco Smoke versus Electronic Cigarette Aerosol. Mutat. Res. Rev. Mutat. Res. 2022, 789, 108409. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Khan, S.; Tollefsbol, T.O.; Meeran, S.M. Genetics and Epigenetics of Lung Cancer: Mechanisms and Future Perspectives. Curr. Cancer Ther. Rev. 2013, 9, 97–110. [Google Scholar] [CrossRef]
- Wen, J.; Fu, J.; Zhang, W.; Guo, M. Genetic and Epigenetic Changes in Lung Carcinoma and Their Clinical Implications. Mod. Pathol. 2011, 24, 932–943. [Google Scholar] [CrossRef]
- Caliri, A.W.; Tommasi, S.; Besaratinia, A. Relationships among Smoking, Oxidative Stress, Inflammation, Macromolecular Damage, and Cancer. Mutat. Res. 2021, 787, 108365. [Google Scholar] [CrossRef]
- Xie, Z.; Rahman, I.; Goniewicz, M.L.; Li, D. Perspectives on Epigenetics Alterations Associated with Smoking and Vaping. Function 2021, 2, zqab022. [Google Scholar] [CrossRef]
- Szulakowski, P.; Crowther, A.J.L.; Jiménez, L.A.; Donaldson, K.; Mayer, R.; Leonard, T.B.; MacNee, W.; Drost, E.M. The Effect of Smoking on the Transcriptional Regulation of Lung Inflammation in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2006, 174, 41–50. [Google Scholar] [CrossRef]
- Ito, K.; Lim, S.; Caramori, G.; Chung, K.F.; Barnes, P.J.; Adcock, I.M. Cigarette Smoking Reduces Histone Deacetylase 2 Expression, Enhances Cytokine Expression, and Inhibits Glucocorticoid Actions in Alveolar Macrophages. FASEB J. 2001, 15, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- Caliri, A.W.; Caceres, A.; Tommasi, S.; Besaratinia, A. Hypomethylation of LINE-1 Repeat Elements and Global Loss of DNA Hydroxymethylation in Vapers and Smokers. Epigenetics 2020, 15, 816–829. [Google Scholar] [CrossRef] [PubMed]
- Marwick, J.A.; Kirkham, P.A.; Stevenson, C.S.; Danahay, H.; Giddings, J.; Butler, K.; Donaldson, K.; Macnee, W.; Rahman, I. Cigarette Smoke Alters Chromatin Remodeling and Induces Proinflammatory Genes in Rat Lungs. Am. J. Respir. Cell Mol. Biol. 2004, 31, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Choukrallah, M.-A.; Sierro, N.; Martin, F.; Baumer, K.; Thomas, J.; Ouadi, S.; Hoeng, J.; Peitsch, M.C.; Ivanov, N.V. Tobacco Heating System 2.2 Has a Limited Impact on DNA Methylation of Candidate Enhancers in Mouse Lung Compared with Cigarette Smoke. Food Chem. Toxicol. 2019, 123, 501–510. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Luo, F.; Xu, Y.; Qin, Y.; Lu, X.; Xu, W.; Shi, L.; Liu, Q.; Xiang, Q. Epigenetic Silencing of microRNA-218 via EZH2-Mediated H3K27 Trimethylation Is Involved in Malignant Transformation of HBE Cells Induced by Cigarette Smoke Extract. Arch. Toxicol. 2016, 90, 449–461. [Google Scholar] [CrossRef]
- Huang, H.; Ji, Y.; Zhang, J.; Su, Z.; Liu, M.; Tong, J.; Ge, C.; Chen, T.; Li, J. Aberrant DNA Methylation in Radon and/or Cigarette Smoke-Induced Malignant Transformation in BEAS-2B Human Lung Cell Line. J. Toxicol. Environ. Health A 2017, 80, 1321–1330. [Google Scholar] [CrossRef]
- Sundar, I.K.; Nevid, M.Z.; Friedman, A.E.; Rahman, I. Cigarette Smoke Induces Distinct Histone Modifications in Lung Cells: Implications for the Pathogenesis of COPD and Lung Cancer. J. Proteome Res. 2014, 13, 982–996. [Google Scholar] [CrossRef]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in Cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef]
- Yu, V.; Rahimy, M.; Korrapati, A.; Xuan, Y.; Zou, A.E.; Krishnan, A.R.; Tsui, T.; Aguilera, J.A.; Advani, S.; Crotty Alexander, L.E.; et al. Electronic Cigarettes Induce DNA Strand Breaks and Cell Death Independently of Nicotine in Cell Lines. Oral. Oncol. 2016, 52, 58–65. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Elkhatib, R.; Al-Rajoudi, T.; Al-Qudaihi, G.; Manogarannogaran, P.; Eltabache, C.; Alotaibi, A.; Mummer, A.B.; Almugbel, S. Cytotoxic and Genotoxic Effects of E-Liquids and Their Potential Associations with Nicotine, Menthol and Phthalate Esters. Chemosphere 2020, 249, 126153. [Google Scholar] [CrossRef]
- Tellez, C.S.; Juri, D.E.; Phillips, L.M.; Do, K.; Yingling, C.M.; Thomas, C.L.; Dye, W.W.; Wu, G.; Kishida, S.; Kiyono, T.; et al. Cytotoxicity and Genotoxicity of E-Cigarette Generated Aerosols Containing Diverse Flavoring Products and Nicotine in Oral Epithelial Cell Lines. Toxicol. Sci. 2021, 179, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Misra, M.; Leverette, R.D.; Cooper, B.T.; Bennett, M.B.; Brown, S.E. Comparative in Vitro Toxicity Profile of Electronic and Tobacco Cigarettes, Smokeless Tobacco and Nicotine Replacement Therapy Products: E-Liquids, Extracts and Collected Aerosols. Int. J. Environ. Res. Public Health 2014, 11, 11325–11347. [Google Scholar] [CrossRef]
- Wieczorek, R.; Phillips, G.; Czekala, L.; Trelles Sticken, E.; O’Connell, G.; Simms, L.; Rudd, K.; Stevenson, M.; Walele, T. A Comparative in Vitro Toxicity Assessment of Electronic Vaping Product E-Liquids and Aerosols with Tobacco Cigarette Smoke. Toxicol. In Vitro 2020, 66, 104866. [Google Scholar] [CrossRef] [PubMed]
- Thorne, D.; Leverette, R.; Breheny, D.; Lloyd, M.; McEnaney, S.; Whitwell, J.; Clements, J.; Bombick, B.; Gaça, M. Genotoxicity Evaluation of Tobacco and Nicotine Delivery Products: Part Two. In Vitro Micronucleus Assay. Food Chem. Toxicol. 2019, 132, 110546. [Google Scholar] [CrossRef] [PubMed]
- Thorne, D.; Leverette, R.; Breheny, D.; Lloyd, M.; McEnaney, S.; Whitwell, J.; Clements, J.; Bombick, B.; Gaca, M. Genotoxicity Evaluation of Tobacco and Nicotine Delivery Products: Part One. Mouse Lymphoma Assay. Food Chem. Toxicol. 2019, 132, 110584. [Google Scholar] [CrossRef]
- Thorne, D.; Whitwell, J.; Clements, J.; Walker, P.; Breheny, D.; Gaca, M. The Genotoxicological Assessment of a Tobacco Heating Product Relative to Cigarette Smoke Using the in Vitro Micronucleus Assay. Toxicol. Rep. 2020, 7, 1010–1019. [Google Scholar] [CrossRef]
- Chapman, F.; Sticken, E.T.; Wieczorek, R.; Pour, S.J.; Dethloff, O.; Budde, J.; Rudd, K.; Mason, E.; Czekala, L.; Yu, F.; et al. Multiple Endpoint in Vitro Toxicity Assessment of a Prototype Heated Tobacco Product Indicates Substantially Reduced Effects Compared to Those of Combustible Cigarette. Toxicol. In Vitro 2023, 86, 105510. [Google Scholar] [CrossRef]
- Vivarelli, F.; Canistro, D.; Cirillo, S.; Elias, R.J.; Granata, S.; Mussoni, M.; Burattini, S.; Falcieri, E.; Turrini, E.; Fimognari, C.; et al. Unburned Tobacco Cigarette Smoke Alters Rat Ultrastructural Lung Airways and DNA. Nicotine Tob. Res. 2021, 23, 2127–2134. [Google Scholar] [CrossRef]
- Crooks, I.; Neilson, L.; Scott, K.; Reynolds, L.; Oke, T.; Forster, M.; Meredith, C.; McAdam, K.; Proctor, C. Evaluation of Flavourings Potentially Used in a Heated Tobacco Product: Chemical Analysis, in Vitro Mutagenicity, Genotoxicity, Cytotoxicity and in Vitro Tumour Promoting Activity. Food Chem. Toxicol. 2018, 118, 940–952. [Google Scholar] [CrossRef]
- Anthérieu, S.; Garat, A.; Beauval, N.; Soyez, M.; Allorge, D.; Garçon, G.; Lo-Guidice, J.-M. Comparison of Cellular and Transcriptomic Effects between Electronic Cigarette Vapor and Cigarette Smoke in Human Bronchial Epithelial Cells. Toxicol. In Vitro 2017, 45, 417–425. [Google Scholar] [CrossRef]
- World Health Organization & WHO Tobacco Free Initiative. Standard operating procedure for intense smoking of cigarettes. In WHO TobLabNet Official Method SOP 01; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- AFNOR. Norme XP D90-300-3 Cigarettes Électroniques et e-Liquides. In Partie 3: Exigences et méthodes d’essais Reatives; Association Française de Normalisation (AFNOR): Paris, France, 2016. [Google Scholar]
- Beauval, N.; Antherieu, S.; Soyez, M.; Gengler, N.; Grova, N.; Howsam, M.; Hardy, E.M.; Fischer, M.; Appenzeller, B.M.R.; Goossens, J.-F.; et al. Chemical Evaluation of Electronic Cigarettes: Multicomponent Analysis of Liquid Refills and Their Corresponding Aerosols. J. Anal. Toxicol. 2017, 41, 670–678. [Google Scholar] [CrossRef]
- Platel, A.; Privat, K.; Talahari, S.; Delobel, A.; Dourdin, G.; Gateau, E.; Simar, S.; Saleh, Y.; Sotty, J.; Antherieu, S.; et al. Study of in Vitro and in Vivo Genotoxic Effects of Air Pollution Fine (PM2.5-0.18) and Quasi-Ultrafine (PM0.18) Particles on Lung Models. Sci. Total Environ. 2020, 711, 134666. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A Simple Technique for Quantitation of Low Levels of DNA Damage in Individual Cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single Cell Gel/Comet Assay: Guidelines for in Vitro and in Vivo Genetic Toxicology Testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.C.; O’Donovan, M.R.; Martin, E.A. hOGG1 Recognizes Oxidative Damage Using the Comet Assay with Greater Specificity than FPG or ENDOIII. Mutagenesis 2006, 21, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Chang, W.P.; Kirsch-Volders, M.; Holland, N.; Bonassi, S.; Zeiger, E.; HUman MicronNucleus Project. HUMN Project: Detailed Description of the Scoring Criteria for the Cytokinesis-Block Micronucleus Assay Using Isolated Human Lymphocyte Cultures. Mutat. Res. 2003, 534, 65–75. [Google Scholar] [CrossRef]
- Fenech, M.; Bolognesi, C.; Kirsch-Volders, M.; Bonassi, S.; Zeiger, E.; Knasmüller, S.; Holland, N. Harmonisation of the Micronucleus Assay in Human Buccal Cells—A Human Micronucleus (HUMN) Project (Www.Humn.Org) Initiative Commencing in 2007. Mutagenesis 2007, 22, 3–4. [Google Scholar] [CrossRef]
- OECD. OECD Test Guideline No. 487: In Vitro Mammalian Cell Micronucleus Test, OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2016. [Google Scholar] [CrossRef]
- Matsushima, T.; Hayashi, M.; Matsuoka, A.; Ishidate, M.; Miura, K.F.; Shimizu, H.; Suzuki, Y.; Morimoto, K.; Ogura, H.; Mure, K.; et al. Validation Study of the in Vitro Micronucleus Test in a Chinese Hamster Lung Cell Line (CHL/IU). Mutagenesis 1999, 14, 569–580. [Google Scholar] [CrossRef]
- Santibáñez-Andrade, M.; Quezada-Maldonado, E.M.; Rivera-Pineda, A.; Chirino, Y.I.; García-Cuellar, C.M.; Sánchez-Pérez, Y. The Road to Malignant Cell Transformation after Particulate Matter Exposure: From Oxidative Stress to Genotoxicity. Int. J. Mol. Sci. 2023, 24, 1782. [Google Scholar] [CrossRef]
- Ďurovcová, I.; Kyzek, S.; Fabová, J.; Makuková, J.; Gálová, E.; Ševčovičová, A. Genotoxic Potential of Bisphenol A: A Review. Environ. Pollut. 2022, 306, 119346. [Google Scholar] [CrossRef]
- Tattan-Birch, H.; Hartmann-Boyce, J.; Kock, L.; Simonavicius, E.; Brose, L.; Jackson, S.; Shahab, L.; Brown, J. Heated Tobacco Products for Smoking Cessation and Reducing Smoking Prevalence. Cochrane Database Syst. Rev. 2022, 2022, CD013790. [Google Scholar] [CrossRef]
- Lam, C.; West, A. Are Electronic Nicotine Delivery Systems an Effective Smoking Cessation Tool? Can. J. Respir. Ther. 2015, 51, 93–98. [Google Scholar]
- Escobar, Y.-N.H.; Nipp, G.; Cui, T.; Petters, S.S.; Surratt, J.D.; Jaspers, I. In Vitro Toxicity and Chemical Characterization of Aerosol Derived from Electronic Cigarette Humectants Using a Newly Developed Exposure System. Chem. Res. Toxicol. 2020, 33, 1677–1688. [Google Scholar] [CrossRef]
- Bishop, E.; Haswell, L.; Adamson, J.; Costigan, S.; Thorne, D.; Gaca, M. An Approach to Testing Undiluted E-Cigarette Aerosol in Vitro Using 3D Reconstituted Human Airway Epithelium. Toxicol. In Vitro 2019, 54, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Canchola, A.; Ahmed, C.M.S.; Chen, K.; Chen, J.Y.; Lin, Y.-H. Formation of Redox-Active Duroquinone from Vaping of Vitamin E Acetate Contributes to Oxidative Lung Injury. Chem. Res. Toxicol. 2022, 35, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Giebe, S.; Hofmann, A.; Brux, M.; Lowe, F.; Breheny, D.; Morawietz, H.; Brunssen, C. Comparative Study of the Effects of Cigarette Smoke versus next Generation Tobacco and Nicotine Product Extracts on Endothelial Function. Redox Biol. 2021, 47, 102150. [Google Scholar] [CrossRef]
- Khalil, C.; Chahine, J.B.; Haykal, T.; Al Hageh, C.; Rizk, S.; Khnayzer, R.S. E-Cigarette Aerosol Induced Cytotoxicity, DNA Damages and Late Apoptosis in Dynamically Exposed A549 Cells. Chemosphere 2021, 263, 127874. [Google Scholar] [CrossRef]
- Ganapathy, V.; Manyanga, J.; Brame, L.; McGuire, D.; Sadhasivam, B.; Floyd, E.; Rubenstein, D.A.; Ramachandran, I.; Wagener, T.; Queimado, L. Electronic Cigarette Aerosols Suppress Cellular Antioxidant Defenses and Induce Significant Oxidative DNA Damage. PLoS ONE 2017, 12, e0177780. [Google Scholar] [CrossRef]
- Li, X. In Vitro Toxicity Testing of Cigarette Smoke Based on the Air-Liquid Interface Exposure: A Review. Toxicol. In Vitro 2016, 36, 105–113. [Google Scholar] [CrossRef]
- Platel, A.; Dusautoir, R.; Kervoaze, G.; Dourdin, G.; Gateau, E.; Talahari, S.; Huot, L.; Simar, S.; Ollivier, A.; Laine, W.; et al. Comparison of the in Vivo Genotoxicity of Electronic and Conventional Cigarettes Aerosols after Subacute, Subchronic and Chronic Exposures. J. Hazard. Mater. 2022, 423, 127246. [Google Scholar] [CrossRef]
- Emma, R.; Caruso, M.; Campagna, D.; Pulvirenti, R.; Li Volti, G. The Impact of Tobacco Cigarettes, Vaping Products and Tobacco Heating Products on Oxidative Stress. Antioxidants 2022, 11, 1829. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarcone, G.; Lenski, M.; Martinez, T.; Talahari, S.; Simonin, O.; Garçon, G.; Allorge, D.; Nesslany, F.; Lo-Guidice, J.-M.; Platel, A.; et al. Impact of Electronic Cigarettes, Heated Tobacco Products and Conventional Cigarettes on the Generation of Oxidative Stress and Genetic and Epigenetic Lesions in Human Bronchial Epithelial BEAS-2B Cells. Toxics 2023, 11, 847. https://doi.org/10.3390/toxics11100847
Zarcone G, Lenski M, Martinez T, Talahari S, Simonin O, Garçon G, Allorge D, Nesslany F, Lo-Guidice J-M, Platel A, et al. Impact of Electronic Cigarettes, Heated Tobacco Products and Conventional Cigarettes on the Generation of Oxidative Stress and Genetic and Epigenetic Lesions in Human Bronchial Epithelial BEAS-2B Cells. Toxics. 2023; 11(10):847. https://doi.org/10.3390/toxics11100847
Chicago/Turabian StyleZarcone, Gianni, Marie Lenski, Thomas Martinez, Smaïl Talahari, Ophélie Simonin, Guillaume Garçon, Delphine Allorge, Fabrice Nesslany, Jean-Marc Lo-Guidice, Anne Platel, and et al. 2023. "Impact of Electronic Cigarettes, Heated Tobacco Products and Conventional Cigarettes on the Generation of Oxidative Stress and Genetic and Epigenetic Lesions in Human Bronchial Epithelial BEAS-2B Cells" Toxics 11, no. 10: 847. https://doi.org/10.3390/toxics11100847