
Acetyl Tributyl Citrate Exposure at Seemingly Safe Concentrations Induces Adverse Effects in Different Genders of Type 2 Diabetes Mice, Especially Brain Tissue

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Main Reagents and Kits
2.3. Establishment of Mouse T2DM Model
2.4. ATBC Exposure Protocol
2.5. Increase in Body Weight, Food Consumed, and Water Intake
2.6. Determination of FBG Level and Glucose Tolerance
2.7. TC, TG, LDL-C, HDL-C, and Insulin Content Assessment
2.8. Morris Water Maze (MWM)
2.9. Liver, Pancreas, and Brain Sample Histological Examination
2.10. Tissue Sample Preparation
2.11. ACh, 5-HT, and Aβ1–42 Assessment
2.12. Western Blot Analysis of Tau Phosphorylation
2.13. Assessment of Reactive Oxygen Species (ROS), Malondialdehyde (MDA), and Glutathione (GSH)
2.14. Analysis of 4-HNE and 8-OHdG Content, and SOD Activity
2.15. Cytokine Array
2.16. Immunofluorescence (IF) Analysis of GFAP and IBA1
2.17. Statistical Analyses
3. Results
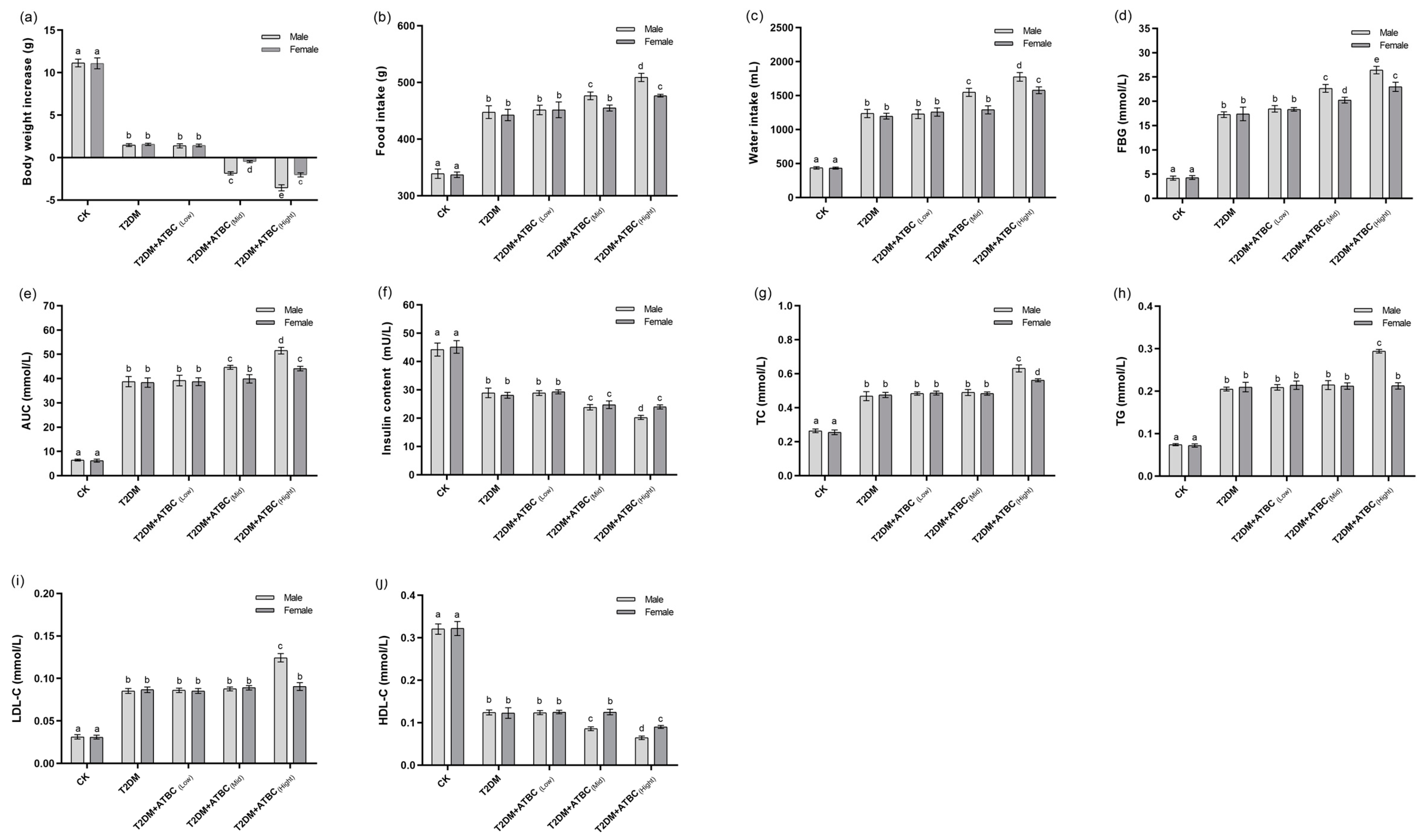
3.1. Glucose and Lipid Metabolism Index Changes in Mice after ATBC Exposure
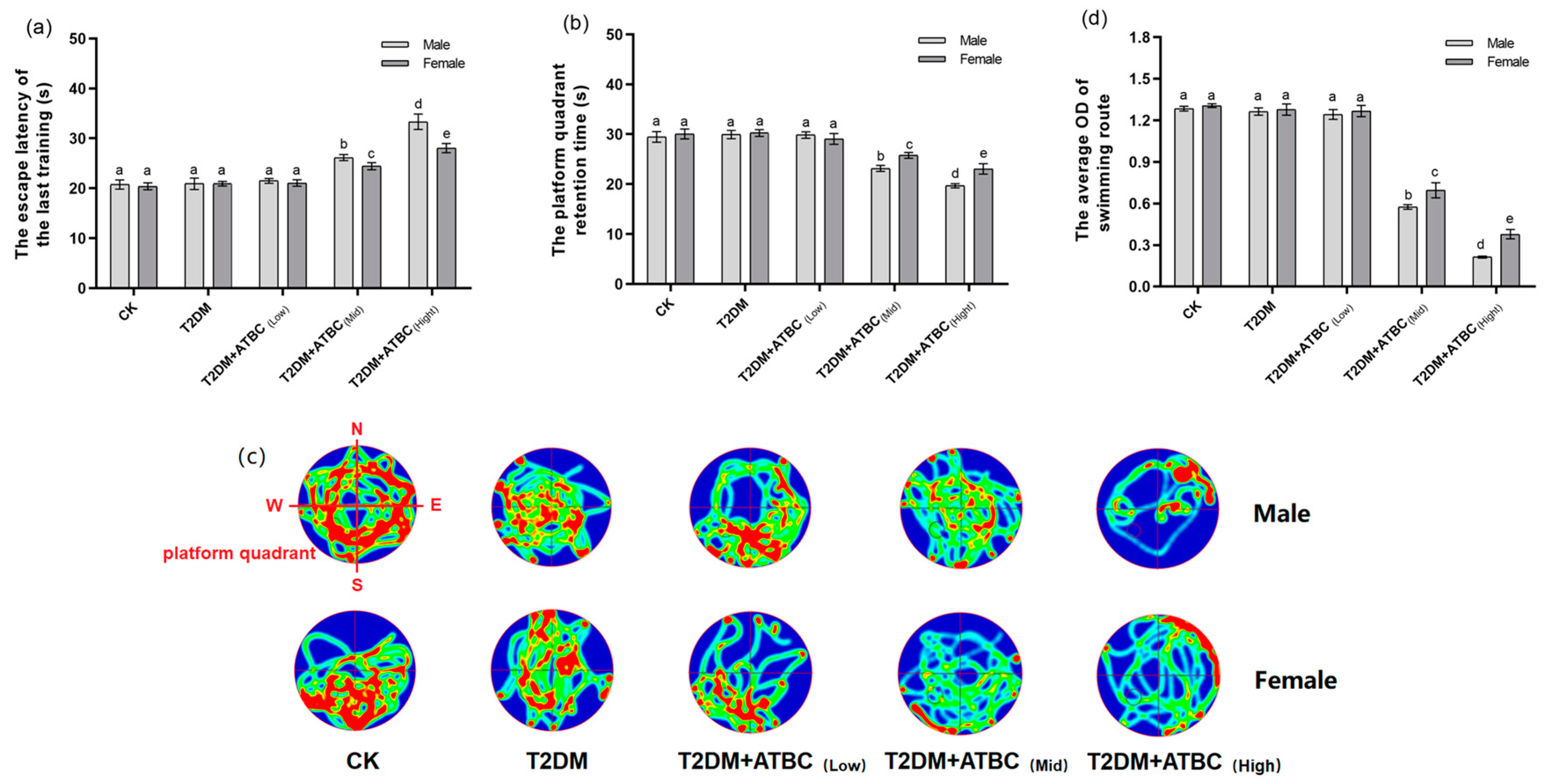
3.2. Cognitive Ability Changes after ATBC Exposure
3.3. Histological Analysis of Liver and Pancreas after ATBC Exposure
3.4. Histological Analysis of Hippocampus after ATBC Exposure
3.5. ACh, 5-HT, Aβ1–42, and Tau Phosphorylation Level Changes in Mouse Brains after ATBC Exposure
3.6. Oxidative-Stress Level Changes in Mouse Brains after ATBC Exposure
3.7. Cytokine Level Changes in Mouse Brains after ATBC Exposure
3.8. Glial Activation in Mouse Brains after ATBC Exposure
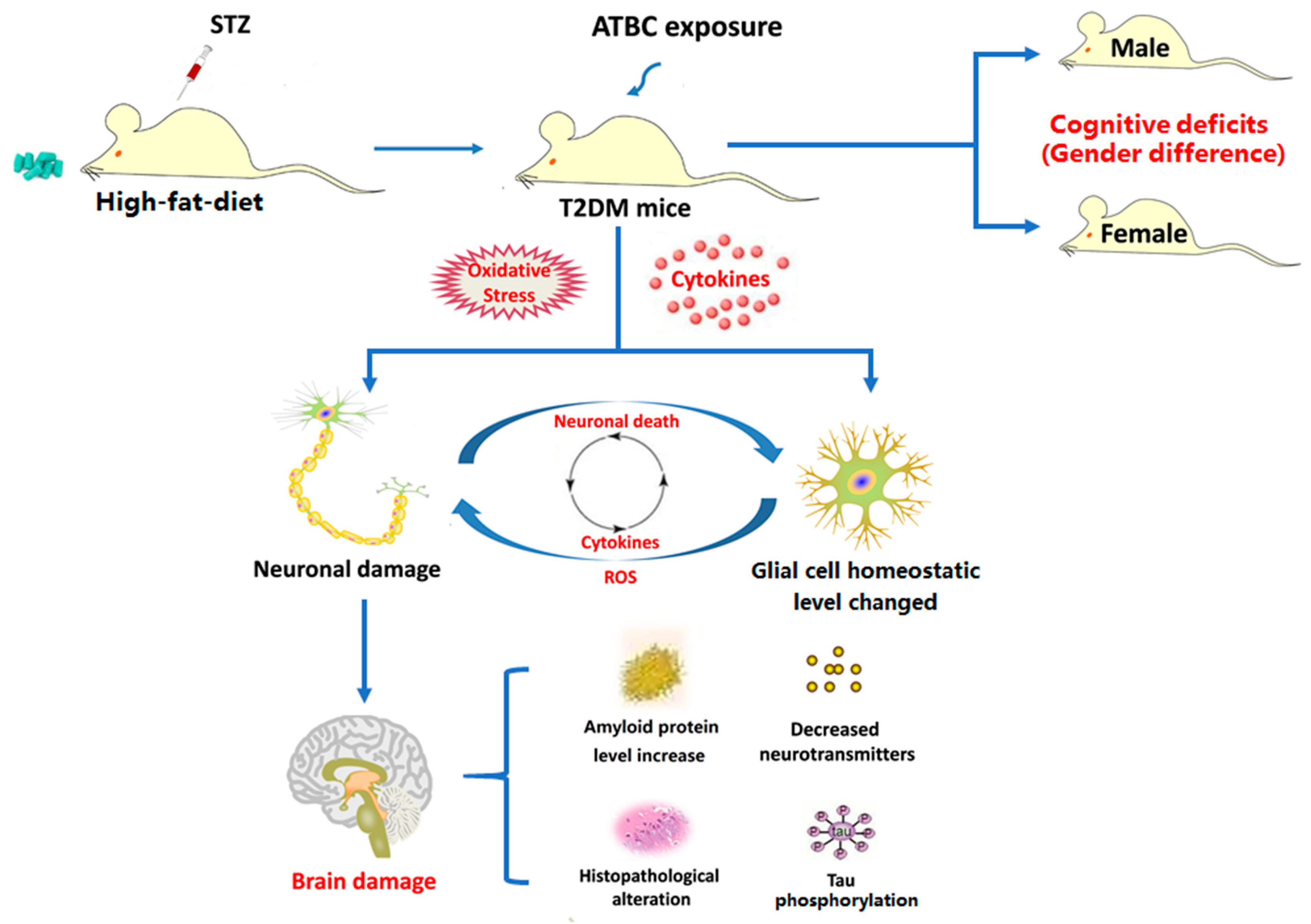
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Report on Diabetes. 2016. Available online: https://www.who.int/publications-detail/global-report-on-diabetes (accessed on 2 March 2020).
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Prattichizzo, F. Variability of risk factors and diabetes complications. Cardiovasc. Diabetol. 2021, 20, 101. [Google Scholar] [CrossRef] [PubMed]
- Donato, J.J. The central nervous system as a promising target to treat diabetes mellitus. Curr. Top. Med. Chem. 2012, 12, 2070–2081. [Google Scholar] [CrossRef]
- Anderson, J.L.; Waller, D.K.; Canfield, M.A.; Shaw, G.M.; Watkins, M.L.; Werler, M.M. Maternal obesity, gestational diabetes, and central nervous system birth defects. Epidemiology 2005, 16, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.J. Final report on the safety assessment of acetyl triethyl citrate, acetyl tributyl citrate, acetyl trihexyl citrate, and acetyl trioctyl citrate. Int. J. Toxicol. 2002, 2, 1–17. [Google Scholar] [CrossRef]
- Kandimalla, R.; Thirumala, V.; Reddy, P.H. Is Alzheimer’s disease a Type 3 Diabetes? A critical appraisal. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1078–1089. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; Xie, Z.; Wang, Y.; Wang, X.; Li, S.; Yan, J.; Zhan, G.; Zhou, Z.; Zhao, Y.; Li, S. Type 2 diabetes mellitus-associated cognitive dysfunction: Advances in potential mechanisms and therapies. Neurosci. Biobehav. Rev. 2022, 137, 104642. [Google Scholar] [CrossRef] [PubMed]
- Mirzadeh, Z.; Faber, C.L.; Schwartz, M.W. Central Nervous System Control of Glucose Homeostasis: A Therapeutic Target for Type 2 Diabetes? Annu. Rev. Pharmacol. Toxicol. 2022, 62, 55–84. [Google Scholar] [CrossRef]
- Lioy, P.J.; Hauser, R.; Gennings, C.; Koch, H.M.; Mirkes, P.E.; Schwetz, B.A.; Kortenkamp, A. Assessment of phthalates/phthalate alternatives in children’s toys and childcare articles: Review of the report including conclusions and recommendation of the Chronic Hazard Advisory Panel of the Consumer Product Safety Commission. J. Expo. Sci. Environ. Epidemiol. 2015, 25, 343–353. [Google Scholar] [CrossRef]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar] [CrossRef]
- Katsikantami, I.; Sifakis, S.; Tzatzarakis, M.N.; Vakonaki, E.; Kalantzi, O.; Tsatsakis, A.M.; Rizos, A.K. A global assessment of phthalates burden and related links to health effects. Environ. Int. 2016, 97, 212–236. [Google Scholar] [CrossRef] [PubMed]
- Review of Exposure and Toxicity Data for Phthalate Substitutes. 2010. Available online: https://www.cpsc.gov/zh-CN/content/review-of-exposure-and-toxicity-data-for-phthalate-substitutes (accessed on 15 February 2020).
- Haned, Z.; Moulay, S.; Lacorte, S. Migration of plasticizers from poly(vinyl chloride) and multilayer infusion bags using selective extraction and GC-MS. J. Pharm. Biomed. Anal. 2018, 156, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Erythropel, H.C.; Maric, M.; Nicell, J.A.; Leask, R.L.; Yargeau, V. Leaching of the plasticizer di(2-ethylhexyl)phthalate (DEHP) from plastic containers and the question of human exposure. Appl. Microbiol. Biotechnol. 2014, 98, 9967–9981. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Zhang, Y.C. Effects of chronic exposure to tributyl acetyl citrate on cognitive ability and oxidative stress level of brain tissue in mice. J. Environ. Health 2021, 1, 26–29, (In Chinese with English abstract). [Google Scholar]
- Liu, X.D.; Zhang, Z.H.; Yang, J.B.; Liu, L.Y.; Zhu, S.J.; Zhang, Y.C.; Yang, X. Effects of Plasticizer Acetyl Tributyl Citrate Exposure on Learning and Memory Abilities and Protective Effects of Vitmain E. Sci. Technol. Food Ind. 2021, 12, 233–240, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Li, D.; Jiang, C.; Mei, G.; Zhao, Y.; Chen, L.; Liu, J.; Tang, Y.; Gao, C.; Yao, P. Quercetin Alleviates Ferroptosis of Pancreatic β Cells in Type 2 Diabetes. Nutrients 2020, 12, 2954. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ma, L.; Ren, X.; Zhang, W.; Shi, D.; Huo, Y.; Ba, Y.; Bai, Y.; Cheng, N. A new mouse model of type 2 diabetes mellitus established through combination of high-fat diet, streptozotocin and glucocorticoid. Life Sci. 2021, 286, 120062. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Liu, X.; Wu, J.; Yan, B.; Zhang, Y.; Lu, Y.; Wu, Y.; Liu, C.; Guo, J.; Nanberg, E.; et al. Cognitive deficits and anxiety induced by diisononyl phthalate in mice and the neuroprotective effects of melatonin. Sci. Rep. 2015, 5, 14676. [Google Scholar] [CrossRef]
- Garaulet, M.; Lopez-Minguez, J.; Dashti, H.S.; Vetter, C.; Hernández-Martínez, A.M.; Pérez-Ayala, M.; Baraza, J.C.; Wang, W.; Florez, J.C.; Scheer, F.A.J.L.; et al. Interplay of Dinner Timing and MTNR1B Type 2 Diabetes Risk Variant on Glucose Tolerance and Insulin Secretion: A Randomized Crossover Trial. Diabetes Care 2022, 45, 512–519. [Google Scholar] [CrossRef]
- Yoshida, M.; Watanabe, C.; Honda, A.; Satoh, M.; Akira Yasutake, A. Emergence of delayed behavioral effects in offspring mice exposed to low levels of mercury vapor during the lactation period. J Toxicol Sci 2013, 38, 1–6. [Google Scholar] [CrossRef]
- Ning, N.; Dang, X.Q.; Bai, C.Y.; Zhang, C.; Wang, K.Z. Panax notoginsenoside produces neuroprotective effects in rat model of acute spinal cord ischemia-reperfusion injury. J. Ethnopharmacol. 2012, 139, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, D.P.; Puig, K.L.; Gorr, M.W.; Wold, L.E.; Combs, C.K. A pilot study to assess effects of long-term inhalation of airborne particulate matter on early Alzheimer-like changes in the mouse brain. PLoS ONE 2015, 10, e0127102. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Zhang, Y.C.; Li, J.Q.; Wang, D.; Wu, Y.; Li, Y.; Lu, Z.S.; Yu, S.C.; Li, R.; Yang, X. Cognitive deficits and decreased locomotor activity induced by single-walled carbon nanotubes and neuroprotective effects of ascorbic acid. Int. J. Nanomed. 2014, 9, 823–839. [Google Scholar] [CrossRef]
- Li, J.; Jia, Z.; Xu, W.; Guo, W.D.; Zhang, M.C.; Bi, J.; Cao, Y.; Fan, Z.K.; Li, G. TGN-020 alleviates edema and inhibits astrocyte activation and glial scar formation after spinal cord compression injury in rats. Life Sci. 2019, 222, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.M.; Sen, N.; Liu, X.S.; Craig, Z.R. Effects of oral exposure to the phthalate substitute acetyl tributyl citrate on female reproduction in mice. J. Appl. Toxicol. 2017, 37, 668–675. [Google Scholar] [CrossRef]
- Subedi, B.; Sullivan, K.D.; Dhungana, B. Phthalate and non-phthalate plasticizersin indoor dust from childcare facilities, salons, and homes across the USA. Environ. Pollut. 2017, 230, 701–708. [Google Scholar] [CrossRef]
- Takeshita, A.; Igarashi-Migitaka, J.; Nishiyama, K.; Takahashi, H.; Takeuchi, Y.; Koibuchi, N. Acetyl tributyl citrate, the most widely used phthalate substitute plasticizer, induces cytochrome p450 3a through steroid and xenobiotic receptor. Toxicol. Sci. 2011, 123, 460–470. [Google Scholar] [CrossRef]
- Athyros, V.G.; Doumas, M.; Imprialos, K.P.; Stavropoulos, K.; Georgianou, E.; Katsimardou, A.; Karagiannis, A. Diabetes and lipid metabolism. Hormones 2018, 1, 61–67. [Google Scholar] [CrossRef]
- Cui, X.; Qian, D.W.; Jiang, S.; Shang, E.X.; Zhu, Z.H.; Duan, J.A. Scutellariae Radix and Coptidis Rhizoma Improve Glucose and Lipid Metabolism in T2DM Rats via Regulation of the Metabolic Profiling and MAPK/PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 3634. [Google Scholar] [CrossRef]
- Xiong, X.Y.; Tang, Y.; Yang, Q.W. Metabolic changes favor the activity and heterogeneity of reactive astrocytes. Trends Endocrinol. Metab. 2022, 33, 390–400. [Google Scholar] [CrossRef]
- Luger, S.; Schwebler, A.; Vutukuri, R.; Bouzas, N.F.; Labocha, S.; Schreiber, Y.; Brunkhorst, R.; Steinmetz, H.; Pfeilschifter, J.; Pfeilschifter, W. Beta adrenoceptor blockade ameliorates impaired glucose tolerance and alterations of the cerebral ceramide metabolism in an experimental model of ischemic stroke. Ther. Adv. Neurol. Disord. 2018, 11, 1756286418769830. [Google Scholar] [CrossRef] [PubMed]
- Strosznajder, A.K.; Wójtowicz, S.; Jeżyna, M.J.; Sun, G.Y.; Strosznajder, J.B. Recent Insights on the Role of PPAR-β/δ in Neuroinflammation and Neurodegeneration, and Its Potential Target for Therapy. NeuroMolecular Med. 2021, 23, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Lawal, S.K.; Olojede, S.O.; Dare, A.; Faborode, O.S.; Naidu, E.C.S.; Rennie, C.O.; Azu, O.O. Silver Nanoparticles Conjugate Attenuates Highly Active Antiretroviral Therapy-Induced Hippocampal Nissl Substance and Cognitive Deficits in Diabetic Rats. J. Diabetes Res. 2021, 2021, 2118538. [Google Scholar] [CrossRef] [PubMed]
- Franconi, F.; Campesi, I.; Occhioni, S.; Tonolo, G. Sex-gender differences in diabetes vascular complications and treatment. Endocr. Metab. Immune Disord. Drug Targets 2012, 12, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Kautzky-Willer, A.; Leutner, M.; Harreiter, J. Sex differences in type 2 diabetes. Diabetologia 2023, 66, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, B. Pathophysiology of oxidative stress in diabetes mellitus. J. Diabetes Complicat. 2001, 15, 203–210. [Google Scholar] [CrossRef]
- Scott, J.A.; King, G.L. Oxidative stress and antioxidant treatment in diabetes. Ann. N. Y. Acad. Sci. 2004, 1031, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender difference in oxidative stress: A new look at the mechanisms for cardiovascular diseases. J. Cell. Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef]
- Sumien, N.; Cunningham, J.T.; Davis, D.L.; Engelland, R.; Fadeyibi, O.; Farmer, G.E.; Mabry, S.; Mensah-Kane, P.; Trinh, O.T.P.; Vann, P.H.; et al. Neurodegenerative Disease: Roles for Sex, Hormones, and Oxidative Stress. Endocrinology 2021, 162, bqab185. [Google Scholar] [CrossRef]
- Ahmed, S.; Gull, A.; Khuroo, T.; Aqil, M.; Sultana, Y. Glial Cell: A Potential Target for Cellular and Drug Based Therapy in Various CNS Diseases. Curr. Pharm. Des. 2017, 23, 2389–2399. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Animal Number |
|---|---|---|
| CK (male/female) | ND + Normal saline | 10 |
| T2DM (male/female) | FD + Normal saline | 10 |
| T2DM + ATBC(Low) (male/female) | FD + 2 mg/kg/day ATBC | 10 |
| T2DM + ATBC(Mid) (male/female) | FD + 20 mg/kg/day ATBC | 10 |
| T2DM + ATBC(High) (male/female) | FD + 200 mg/kg/day ATBC | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, Z.; Zhu, S.; Liu, L.; Liu, X.; Yang, X. Acetyl Tributyl Citrate Exposure at Seemingly Safe Concentrations Induces Adverse Effects in Different Genders of Type 2 Diabetes Mice, Especially Brain Tissue. Toxics 2023, 11, 877. https://doi.org/10.3390/toxics11100877
Zhang Y, Zhang Z, Zhu S, Liu L, Liu X, Yang X. Acetyl Tributyl Citrate Exposure at Seemingly Safe Concentrations Induces Adverse Effects in Different Genders of Type 2 Diabetes Mice, Especially Brain Tissue. Toxics. 2023; 11(10):877. https://doi.org/10.3390/toxics11100877
Chicago/Turabian StyleZhang, Yuchao, Zhihuai Zhang, Sijie Zhu, Liangyu Liu, Xudong Liu, and Xu Yang. 2023. "Acetyl Tributyl Citrate Exposure at Seemingly Safe Concentrations Induces Adverse Effects in Different Genders of Type 2 Diabetes Mice, Especially Brain Tissue" Toxics 11, no. 10: 877. https://doi.org/10.3390/toxics11100877