
Identification and Characterization of Glutathione S-transferase Genes in Spodoptera frugiperda (Lepidoptera: Noctuidae) under Insecticides Stress

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. S. frugiperda Rearing
2.2. Bioassay of Tested Insecticides against Third-Instar S. frugiperda Larvae
2.3. Quantitative Real-Time (PCR qRT-PCR) Analysis
2.4. Bioinformatics Analyses
2.5. In Silico Molecular Docking Assay
2.6. Statistical Analysis
3. Results
3.1. Insecticidal Activity of Tested Insecticides against the Third Larval Instars of S. frugiperda
3.2. Identification and Classification of S. frugiperda GSTs
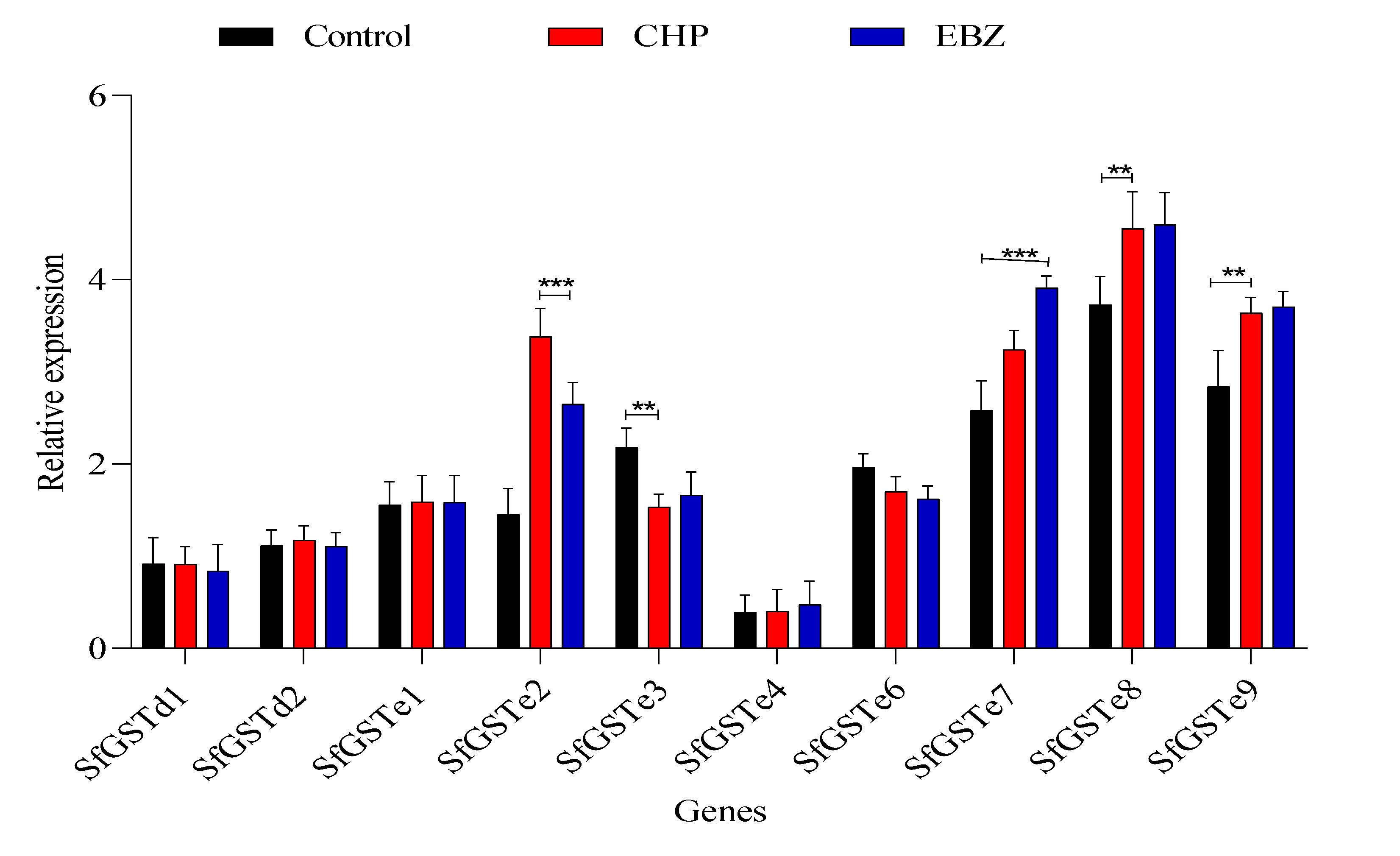
3.3. Expression Profiling of sfGSTs in S. frugiperda Larvae Exposed to Tested Insecticides
3.4. Molecular Docking Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, J.-B.; Li, G.-Q.; Wan, P.-J.; Zhu, T.-T.; Meng, Q.-W. Identification of glutathione S-transferase genes in Leptinotarsa decemlineata and their expression patterns under stress of three insecticides. Pestic. Biochem. Physiol. 2016, 133, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Qie, X.; Lu, W.; Aioub, A.A.; Li, Y.; Wu, W.; Hu, Z. Insight into the detoxification of haedoxan A and the synergistic effects of phrymarolin I against Mythimna separata. Ind. Crops Prod. 2020, 158, 112967. [Google Scholar] [CrossRef]
- Xiao, L.-F.; Zhang, W.; Jing, T.-X.; Zhang, M.-Y.; Miao, Z.-Q.; Wei, D.-D.; Yuan, G.-R.; Wang, J.-J. Genome-wide identification, phylogenetic analysis, and expression profiles of ATP-binding cassette transporter genes in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 25, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Yang, H.; Wang, Z.; Long, G.-Y.; Jin, D.-C. Protective and detoxifying enzyme activity and ABCG subfamily gene expression in Sogatella furcifera under insecticide stress. Front. Physiol. 2019, 9, 1890. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Chakrabarty, S.; Jin, M.; Liu, K.; Xiao, Y. Insect ATP-binding cassette (ABC) transporters: Roles in xenobiotic detoxification and Bt insecticidal activity. Int. J. Mol. Sci. 2019, 20, 2829. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Cheng, Y.; Guo, X.; Li, M.; Chakrabarty, S.; Liu, K.; Wu, K.; Xiao, Y. Down-regulation of lysosomal protein ABCB6 increases gossypol susceptibility in Helicoverpa armigera. Insect Biochem. Mol. Biol. 2020, 122, 103387. [Google Scholar] [CrossRef]
- Jin, M.; Liao, C.; Chakrabarty, S.; Zheng, W.; Wu, K.; Xiao, Y. Transcriptional response of ATP-binding cassette (ABC) transporters to insecticides in the cotton bollworm, Helicoverpa armigera. Pestic. Biochem. Physiol. 2019, 154, 46–59. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Berenbaum, M.R.; Johnson, R.M. Xenobiotic detoxification pathways in honey bees. Curr. Opin. Insect Sci. 2015, 10, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Moural, T.; Koirala BK, S.; Hernandez, J.; Shen, Z.; Alyokhin, A.; Zhu, F. Structural and functional characterization of one unclassified glutathione S-transferase in xenobiotic adaptation of Leptinotarsa decemlineata. Int. J. Mol. Sci. 2021, 22, 11921. [Google Scholar] [CrossRef]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Koirala BK, S.; Moural, T.; Zhu, F. Functional and structural diversity of insect Glutathione S-transferases in xenobiotic adaptation. Int. J. Biol. Sci. 2022, 18, 5713–5723. [Google Scholar] [CrossRef]
- You, Y.; Xie, M.; Ren, N.; Cheng, X.; Li, J.; Ma, X.; Zou, M.; Vasseur, L.; Gurr, G.M.; You, M. Characterization and expression profiling of glutathione S-transferases in the diamondback moth, Plutella xylostella (L.). Bmc Genom. 2015, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Low, W.Y.; Ng, H.L.; Morton, C.J.; Parker, M.W.; Batterham, P.; Robin, C. Molecular evolution of glutathione S-transferases in the genus Drosophila. Genetics 2007, 177, 1363–1375. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Pei, L.; Gu, S.; Zhu, S.; Wang, Y.; Zhang, Y.; Li, B. Glutathione S-transferase (GST) genes in the red flour beetle, Tribolium castaneum, and comparative analysis with five additional insects. Genomics 2012, 100, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Raisuddin, S.; Rhee, J.-S.; Lee, Y.-M.; Han, K.-N.; Lee, J.-S. Molecular cloning, phylogenetic analysis and expression of a MAPEG superfamily gene from the pufferfish Takifugu obscurus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 358–362. [Google Scholar] [CrossRef]
- Kostaropoulos, I.; Papadopoulos, A.I.; Metaxakis, A.; Boukouvala, E.; Papadopoulou-Mourkidou, E. Glutathione S–transferase in the defence against pyrethroids in insects. Insect Biochem. Mol. Biol. 2001, 31, 313–319. [Google Scholar] [CrossRef]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef]
- Zhu, B.; Li, L.; Wei, R.; Liang, P.; Gao, X. Regulation of GSTu1-mediated insecticide resistance in Plutella xylostella by miRNA and lncRNA. PLoS Genet. 2021, 17, e1009888. [Google Scholar] [CrossRef]
- Wei, S.; Clark, A.; Syvanen, M. Identification and cloning of a key insecticide-metabolizing glutathione S-transferase (MdGST-6A) from a hyper insecticide-resistant strain of the housefly Musca domestica. Insect Biochem. Mol. Biol. 2001, 31, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Prapanthadara, L.-a.; Hemingway, J.; Ketterman, A.J. DDT-resistance in Anopheles gambiae (Diptera: Culicidae) from Zanzibar, Tanzania, based on increased DDT-dehydrochlorinase activity of glutathione S-transferases. Bull. Entomol. Res. 1995, 85, 267–274. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Htain, N.N.; Boughton, D.; Zhang, L.; Xiao, Y.; Nagoshi, B.Y.; Mota-Sanchez, D. Southeastern Asia fall armyworms are closely related to populations in Africa and India, consistent with common origin and recent migration. Sci. Rep. 2020, 10, 1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Yang, Y.; Shan, Y.; Chakrabarty, S.; Cheng, Y.; Soberón, M.; Bravo, A.; Liu, K.; Wu, K.; Xiao, Y. Two ABC transporters are differentially involved in the toxicity of two Bacillus thuringiensis Cry1 toxins to the invasive crop-pest Spodoptera frugiperda (JE Smith). Pest Manag. Sci. 2021, 77, 1492–1501. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A. Genetic structure and insecticide resistance characteristics of fall armyworm populations invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef]
- Jamil, S.Z.; Saranum, M.M.; Saleh Hudin, L.J.; Anuar Wan Ali, W.K. First incidence of the invasive fall armyworm, Spodoptera frugiperda (JE Smith, 1797) attacking maize in Malaysia. BioInvasions Rec. 2021, 10, 81–90. [Google Scholar]
- Padhee, A.; Prasanna, B. The emerging threat of Fall Armyworm in India. Indian Farming 2019, 69, 51–54. [Google Scholar]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Dahi, H.F.; Salem, S.A.; Gamil, W.E.; Mohamed, H.O. Heat requirements for the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) as a new invasive pest in Egypt. Egypt. Acad. J. Biol. Sci. A Entomol. 2020, 13, 73–85. [Google Scholar]
- Lima, M.; Silva, P.; Oliveira, O.; Silva, K.; Freitas, F. Corn yield response to weed and fall armyworm controls. Planta Daninha 2010, 28, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Whalon, M.; Mota-Sanchez, D.; Hollingworth, R.; Duynslager, L. Arthropod Pesticide Resistance Database; Michigan State University: East Lansing, MI, USA, 2012; p. 38. [Google Scholar]
- Yu, S.; Nguyen, S.; Abo-Elghar, G. Biochemical characteristics of insecticide resistance in the fall armyworm, Spodoptera frugiperda (JE Smith). Pestic. Biochem. Physiol. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification-a tool for resistance management. Pestic. Biochem. Physiol. 2020, 167, 104587. [Google Scholar] [CrossRef]
- Liu, J.; Hao, Z.; Yang, S.; Lin, Y.; Zhong, H.; Jin, T. Insecticide resistance and its underlying synergism in field populations of Spodoptera frugiperda (JE Smith) from Hainan Island, China. Phytoparasitica 2022, 50, 933–945. [Google Scholar] [CrossRef]
- Roy, D.; Biswas, S.; Sarkar, S.; Adhikary, S.; Chakraborty, G.; Sarkar, P.K.; Al-Shuraym, L.A.; Sayed, S.; Gaber, A.; Hossain, A. Risk Assessment of Fluxametamide Resistance and Fitness Costs in Fall Armyworm (Spodoptera frugiperda). Toxics 2023, 11, 307. [Google Scholar] [CrossRef]
- Chen, H.; Xie, M.; Lin, L.; Zhong, Y.; Zhang, F.; Su, W. Transcriptome analysis of detoxification-related genes in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Insect Sci. 2022, 22, 11. [Google Scholar] [CrossRef]
- Wu, W.; Tu, Y.; Liu, H.; Zhu, J. Celangulins II, III, and IV: New insecticidal sesquiterpenoids from Celastrus angulatus. J. Nat. Prod. 1992, 55, 1294–1298. [Google Scholar] [CrossRef]
- Zhou, L.; Meng, J.Y.; Ruan, H.Y.; Yang, C.L.; Zhang, C.Y. Expression stability of candidate RT-qPCR housekeeping genes in Spodoptera frugiperda (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2021, 108, e21831. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Labute, P. Molecular Operating Environment; Chemical Computing Group. Inc.: Montreal, QC, Canada, 2008. [Google Scholar]
- Damayanthi Devi, I. Comparative binding mode of organophosphates, pyrethroids against modelled structures of acetylcholinesterase and alpha amylase in Blattella germanica. J. Entomol. Zool. Stud. 2015, 3, 233–238. [Google Scholar]
- Elkanzi, N.A.; Hrichi, H.; Bakr, R.B. Antioxidant, Antimicrobial, and Molecular Docking Studies of Novel Chalcones and Schiff Bases Bearing 1, 4-naphthoquinone Moiety. Lett. Drug Des. Discov. 2022, 19, 654–673. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, J.; Chen, S.N.; Huang, L.H.; Feng, Q.L.; Zheng, S.C. Expression profiles of glutathione S-transferase superfamily in Spodoptera litura tolerated to sublethal doses of chlorpyrifos. Insect Sci. 2016, 23, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, Y.-X.; Wang, W.-L.; Zhang, B.-X.; Li, S.-G. Identification and characterisation of seventeen glutathione S-transferase genes from the cabbage white butterfly Pieris rapae. Pestic. Biochem. Physiol. 2017, 143, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-W.; Liang, Q.-M.; Xu, Y.; Gurr, G.M.; Bao, Y.-Y.; Zhou, X.-P.; Zhang, C.-X.; Cheng, J.; Zhu, Z.-R. Genomic insights into the glutathione S-transferase gene family of two rice planthoppers, Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). PLoS ONE 2013, 8, e56604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, F.; Haubruge, E.; Gaspar, C.; Dierickx, P.J. Glutathione S-transferases of Aulacorthum solani and Acyrthosiphon pisum: Partial purification and characterization. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Ortelli, F.; Rossiter, L.C.; Hemingway, J.; Ranson, H. The Anopheles gambiae glutathione transferase supergene family: Annotation, phylogeny and expression profiles. BMC Genom. 2003, 4, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Lu, C.; Li, B.; Fang, S.; Zuo, W.; Dai, F.; Zhang, Z.; Xiang, Z. Identification, genomic organization and expression pattern of glutathione S-transferase in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1158–1164. [Google Scholar] [CrossRef]
- Prasanna, B.; Huesing, J.; Eddy, R.; Peschke, V. Fall Armyworm in Africa: A Guide for Integrated Pest Management; USAID: Washington, DC, USA; CIMMYT: Texcoco, Mexico, 2018.
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhang, L.; Yang, C.; Zong, M.; Huang, Q.; Tao, L. Detection on emamectin benzoate-induced apoptosis and DNA damage in Spodoptera frugiperda Sf-9 cell line. Pestic. Biochem. Physiol. 2016, 126, 6–12. [Google Scholar] [CrossRef]
- Fanigliulo, A.; Sacchetti, M. Emamectin benzoate: New insecticide against Helicoverpa armigera. Commun. Agric. Appl. Biol. Sci. 2008, 73, 651–653. [Google Scholar]
- Lahm, G.P.; Stevenson, T.M.; Selby, T.P.; Freudenberger, J.H.; Cordova, D.; Flexner, L.; Bellin, C.A.; Dubas, C.M.; Smith, B.K.; Hughes, K.A. Rynaxypyr™: A new insecticidal anthranilic diamide that acts as a potent and selective ryanodine receptor activator. Bioorganic Med. Chem. Lett. 2007, 17, 6274–6279. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Huang, J.-M.; Ni, H.; Guo, D.; Yang, F.-X.; Wang, X.; Wu, S.-F.; Gao, C.-F. Susceptibility of fall armyworm, Spodoptera frugiperda (JE Smith), to eight insecticides in China, with special reference to lambda-cyhalothrin. Pestic. Biochem. Physiol. 2020, 168, 104623. [Google Scholar] [CrossRef]
- Hardke, J.T.; Temple, J.H.; Leonard, B.R.; Jackson, R.E. Laboratory toxicity and field efficacy of selected insecticides against fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 2011, 94, 272–278. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, J.; Wang, K.; Zhang, Y.; Liu, Z.; Yu, N. A Binary Mixture of Emamectin Benzoate and Chlorantraniliprole Supplemented with an Adjuvant Effectively Controls Spodoptera frugiperda. Insects 2022, 13, 1157. [Google Scholar] [CrossRef]
- Chen, H.-L.; Hasnain, A.; Cheng, Q.-H.; Xia, L.-J.; Cai, Y.-H.; Hu, R.; Gong, C.-W.; Liu, X.-M.; Pu, J.; Zhang, L. Resistance monitoring and mechanism in the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) for chlorantraniliprole from Sichuan Province, China. Front. Physiol. 2023, 14, 694. [Google Scholar] [CrossRef]
- El-Sayed, M.H.; Ibrahim, M.M.; Elsobki, A.E.; Aioub, A.A. Enhancing the Toxicity of Cypermethrin and Spinosad against Spodoptera littoralis (Lepidoptera: Noctuidae) by Inhibition of Detoxification Enzymes. Toxics 2023, 11, 215. [Google Scholar] [CrossRef]
- Liu, S.; Rao, X.J.; Li, M.Y.; Feng, M.F.; He, M.Z.; Li, S.G. Glutathione S-transferase genes in the rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae): Identification and expression profiles. Arch. Insect Biochem. Physiol. 2015, 90, 1–13. [Google Scholar] [CrossRef]
- Deng, H.; Huang, Y.; Feng, Q.; Zheng, S. Two epsilon glutathione S-transferase cDNAs from the common cutworm, Spodoptera litura: Characterization and developmental and induced expression by insecticides. J. Insect Physiol. 2009, 55, 1174–1183. [Google Scholar] [CrossRef]
- Gui, Z.; Hou, C.; Liu, T.; Qin, G.; Li, M.; Jin, B. Effects of insect viruses and pesticides on glutathione S-transferase activity and gene expression in Bombyx mori. J. Econ. Entomol. 2009, 102, 1591–1598. [Google Scholar] [CrossRef]
- Hu, B.; Hu, S.; Huang, H.; Wei, Q.; Ren, M.; Huang, S.; Tian, X.; Su, J. Insecticides induce the co-expression of glutathione S-transferases through ROS/CncC pathway in Spodoptera exigua. Pestic. Biochem. Physiol. 2019, 155, 58–71. [Google Scholar] [CrossRef]
- Hu, F.; Dou, W.; Wang, J.J.; Jia, F.X.; Wang, J.J. Multiple glutathione S-transferase genes: Identification and expression in oriental fruit fly, Bactrocera dorsalis. Pest Manag. Sci. 2014, 70, 295–303. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Jia, Y.; Li, J.; Li, Z.; Liu, X. Molecular identification of glutathione S-transferase genes and their potential roles in insecticides susceptibility of Grapholita molesta. J. Appl. Entomol. 2023, 147, 249–260. [Google Scholar] [CrossRef]
- Chen, X.e.; Zhang, Y.l. Identification and characterisation of multiple glutathione S-transferase genes from the diamondback moth, Plutella xylostella. Pest Manag. Sci. 2015, 71, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, X.; Liu, J.; Hu, M.; Zhong, G.; Geng, P.; Yi, X. Molecular cloning, expression and molecular modeling of chemosensory protein from Spodoptera litura and its binding properties with Rhodojaponin III. PLoS ONE 2012, 7, e47611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, C.M.; Jiang, X.; Oldfield, T.; Waldman, M. LigandFit: A novel method for the shape-directed rapid docking of ligands to protein active sites. J. Mol. Graph. Model. 2003, 21, 289–307. [Google Scholar] [CrossRef]
- Chu, Y.-H.; Li, Y.; Wang, Y.-T.; Li, B.; Zhang, Y.-H. Investigation of interaction modes involved in alkaline phosphatase and organophosphorus pesticides via molecular simulations. Food Chem. 2018, 254, 80–86. [Google Scholar] [CrossRef]
- Jiang, L.; Li, Y.; Shi, W.; Chen, W.; Ma, Z.; Feng, J.; Hashem, A.S.; Wu, H. Cloning and expression of the mitochondrial cytochrome c oxidase subunit II gene in Sitophilus zeamais and interaction mechanism with allyl isothiocyanate. Pestic. Biochem. Physiol. 2023, 192, 105392. [Google Scholar] [CrossRef]
- Alanazi, M.A.; Arafa, W.A.; Althobaiti, I.O.; Altaleb, H.A.; Bakr, R.B.; Elkanzi, N.A. Green Design, Synthesis, and Molecular Docking Study of Novel Quinoxaline Derivatives with Insecticidal Potential against Aphis craccivora. ACS Omega 2022, 7, 27674–27689. [Google Scholar] [CrossRef]
- Yang, S.; Peng, H.; Zhu, J.; Zhao, C.; Xu, H. Design, synthesis, insecticidal activities, and molecular docking of novel pyridylpyrazolo carboxylate derivatives. J. Heterocycl. Chem. 2022, 59, 1366–1375. [Google Scholar] [CrossRef]
- Wiktelius, E.; Stenberg, G. Novel class of glutathione transferases from cyanobacteria exhibit high catalytic activities towards naturally occurring isothiocyanates. Biochem. J. 2007, 406, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Yang, Y.; Wen, X.; Ou, S.; Zhang, P.; Yu, Q.; Zhang, Y.; Shen, G.; Xu, Z.; Li, J. Stability of cyflumetofen resistance in Tetranychus cinnabarinus and its correlation with glutathione-S-transferase gene expression. Pest Manag. Sci. 2019, 75, 2802–2809. [Google Scholar] [CrossRef]
- Gonzalez, D.; Fraichard, S.; Grassein, P.; Delarue, P.; Senet, P.; Nicolaï, A.; Chavanne, E.; Mucher, E.; Artur, Y.; Ferveur, J.-F. Characterization of a Drosophila glutathione transferase involved in isothiocyanate detoxification. Insect Biochem. Mol. Biol. 2018, 95, 33–43. [Google Scholar] [CrossRef]
- Tan, X.; Hu, X.-M.; Zhong, X.-W.; Chen, Q.-M.; Xia, Q.-Y.; Zhao, P. Antenna-specific glutathione S-transferase in male silkmoth Bombyx mori. Int. J. Mol. Sci. 2014, 15, 7429–7443. [Google Scholar] [CrossRef] [Green Version]
- Naine, S.J.; Devi, C.S.; Mohanasrinivasan, V.; Doss, C.G.P.; Kumar, D.T. Binding and molecular dynamic studies of sesquiterpenes (2R-acetoxymethyl-1, 3, 3-trimethyl-4t-(3-methyl-2-buten-1-yl)-1t-cyclohexanol) derived from marine Streptomyces sp. VITJS8 as potential anticancer agent. Appl. Microbiol. Biotechnol. 2016, 100, 2869–2882. [Google Scholar] [CrossRef]
- Hashem, A.S.; Ramadan, M.M.; Abdel-Hady, A.A.; Sut, S.; Maggi, F.; Dall’Acqua, S. Pimpinella anisum essential oil nanoemulsion toxicity against Tribolium castaneum? Shedding light on its interactions with aspartate aminotransferase and alanine aminotransferase by molecular docking. Molecules 2020, 25, 4841. [Google Scholar] [CrossRef]
- Da Silva Mesquita, R.; Kyrylchuk, A.; Grafova, I.; Kliukovskyi, D.; Bezdudnyy, A.; Rozhenko, A.; Tadei, W.P.; Leskelä, M.; Grafov, A. Synthesis, molecular docking studies, and larvicidal activity evaluation of new fluorinated neonicotinoids against Anopheles darlingi larvae. PLoS ONE 2020, 15, e0227811. [Google Scholar] [CrossRef]
- Tiwari, N.; Mishra, A. Computational perspectives on Chlorpyrifos and its degradants as human glutathione S-transferases inhibitors: DFT calculations, molecular docking study and MD simulations. Comput. Toxicol. 2023, 26, 100264. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Toxicity Regression Equation | LC50 (mg/L) | 95% Fiducial Limits (mg/L) | r | df |
|---|---|---|---|---|---|
| EBZ | y = 0.059x + 3.2355 | 0.029 | 0.019–0.045 | 0.97 | 4 |
| CHP | y = 0.058x + 2.6388 | 1.250 | 0.973–1.455 | 0.96 | 4 |
| Group | Gene Name | ORF (bp) | Protein (aa) | Molecular Weight (kDa) | Theoretical pI | NCBI ID |
|---|---|---|---|---|---|---|
| Delta | SfGSTd1 | 723 | 240 | 27.23 | 5.64 | MZ673611 |
| SfGSTd2 | 651 | 216 | 24.22 | 6.90 | MZ673619 | |
| Epsilon | SfGSTe1 | 594 | 197 | 23.01 | 5.75 | MZ673615 |
| SfGSTe2 | 654 | 217 | 25.17 | 6.23 | MZ673616 | |
| SfGSTe3 | 654 | 217 | 25.14 | 6.53 | MZ673617 | |
| SfGSTe4 | 348 | 115 | 94.38 | 5.71 | MZ673620 | |
| SfGSTe5 | 291 | 96 | 11.16 | 6.26 | MZ673621 | |
| SfGSTe6 | 669 | 222 | 25.11 | 7.81 | MZ673622 | |
| SfGSTe7 | 693 | 230 | 25.75 | 8.42 | MZ673623 | |
| SfGSTe8 | 429 | 142 | 15.95 | 6.90 | MZ673625 | |
| SfGSTe9 | 432 | 143 | 23.83 | 6.72 | MZ673626 | |
| SfGSTe10 | 657 | 218 | 24.98 | 6.97 | MZ673629 | |
| SfGSTe11 | 657 | 218 | 25.06 | 7.10 | MZ673630 | |
| SfGSTe12 | 420 | 139 | 15.73 | 6.90 | MZ673632 | |
| SfGSTe13 | 474 | 157 | 18.05 | 5.19 | MZ673633 | |
| SfGSTe14 | 540 | 179 | 20.59 | 5.86 | MZ673635 | |
| SfGSTe15 | 657 | 218 | 24.90 | 6.96 | MZ673636 | |
| SfGSTe16 | 420 | 139 | 15.76 | 6.90 | MZ673637 | |
| SfGSTe17 | 192 | 63 | 7.05 | 5.44 | MZ673638 | |
| Omega | SfGSTo1 | 723 | 240 | 28.77 | 7.01 | MZ673610 |
| SfGSTo2 | 849 | 282 | 32.48 | 7.01 | MZ673624 | |
| Sigma | SfGSTs1 | 618 | 205 | 23.98 | 6.98 | MZ673614 |
| SfGSTs2 | 615 | 204 | 23.09 | 4.92 | MZ673618 | |
| SfGSTs3 | 639 | 212 | 24.19 | 6.11 | MZ673628 | |
| SfGSTs4 | 639 | 212 | 20.07 | 5.44 | MZ673631 | |
| SfGSTs5 | 108 | 35 | 10.09 | 4.82 | MZ673634 | |
| SfGSTs6 | 633 | 210 | 23.87 | 5.61 | MZ673639 | |
| Microsomal | SfGSTm1 | 516 | 171 | 16.59 | 9.62 | MZ673612 |
| SfGSTm2 | 456 | 151 | 16.64 | 9.79 | MN480699 | |
| SfGSTm3 | 453 | 150 | 16.64 | 9.65 | MZ673627 | |
| Theta | SfGSTt1 | 687 | 228 | 26.42 | 7.66 | MN480695 |
| Species | Delta | Epsilon | Omega | Sigma | Theta | Zeta | Unclassified | Microsomal | Total |
|---|---|---|---|---|---|---|---|---|---|
| Spodoptera frugiperda | 2 | 17 | 2 | 6 | 1 | 0 | 0 | 3 | 31 |
| Spodoptera litura | 3 | 15 | 3 | 6 | 1 | 2 | 1 | 5 | 36 |
| Pieris rapae | 3 | 3 | 4 | 4 | 1 | 2 | 0 | 0 | 17 |
| Nilaparvata lugens | 2 | 1 | 1 | 3 | 1 | 0 | 1 | 2 | 11 |
| Sogatella furcifera | 2 | 1 | 1 | 1 | 1 | 1 | 0 | 2 | 9 |
| Leptinotarsa decemlineata | 3 | 10 | 5 | 4 | 4 | 1 | 2 | 1 | 30 |
| Drosophila melanogaster | 11 | 14 | 4 | 1 | 4 | 2 | 1 | 3 | 40 |
| Acyrthosiphon pisum | 16 | 1 | 2 | 6 | 2 | 0 | 2 | 3 | 32 |
| Anopheles gambiae | 17 | 8 | 1 | 1 | 2 | 1 | 2 | 3 | 35 |
| Bombyx mori | 5 | 7 | 4 | 2 | 1 | 2 | 0 | 1 | 22 |
| Genes | EBZ | CHP | ||
|---|---|---|---|---|
| Binding Energy (kcal/M) | RMSD (A ◦) | Binding Energy (kcal/M) | RMSD (A ◦) | |
| SfGSTe10 | −24.41 | 4.47 | −26.72 | 2.94 |
| SfGSTe13 | −26.85 | 2.96 | −26.78 | 2.36 |
| SfGSTs1 | −23.16 | 1.49 | −18.45 | 4.61 |
| SfGSTe2 | −21.19 | 2.36 | −19.52 | 4.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aioub, A.A.A.; Hashem, A.S.; El-Sappah, A.H.; El-Harairy, A.; Abdel-Hady, A.A.A.; Al-Shuraym, L.A.; Sayed, S.; Huang, Q.; Abdel-Wahab, S.I.Z. Identification and Characterization of Glutathione S-transferase Genes in Spodoptera frugiperda (Lepidoptera: Noctuidae) under Insecticides Stress. Toxics 2023, 11, 542. https://doi.org/10.3390/toxics11060542
Aioub AAA, Hashem AS, El-Sappah AH, El-Harairy A, Abdel-Hady AAA, Al-Shuraym LA, Sayed S, Huang Q, Abdel-Wahab SIZ. Identification and Characterization of Glutathione S-transferase Genes in Spodoptera frugiperda (Lepidoptera: Noctuidae) under Insecticides Stress. Toxics. 2023; 11(6):542. https://doi.org/10.3390/toxics11060542
Chicago/Turabian StyleAioub, Ahmed A. A., Ahmed S. Hashem, Ahmed H. El-Sappah, Amged El-Harairy, Amira A. A. Abdel-Hady, Laila A. Al-Shuraym, Samy Sayed, Qiulan Huang, and Sarah I. Z. Abdel-Wahab. 2023. "Identification and Characterization of Glutathione S-transferase Genes in Spodoptera frugiperda (Lepidoptera: Noctuidae) under Insecticides Stress" Toxics 11, no. 6: 542. https://doi.org/10.3390/toxics11060542