
Reproductive and Developmental Effects of Sex-Specific Chronic Exposure to Dietary Arsenic in Zebrafish (Danio rerio)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
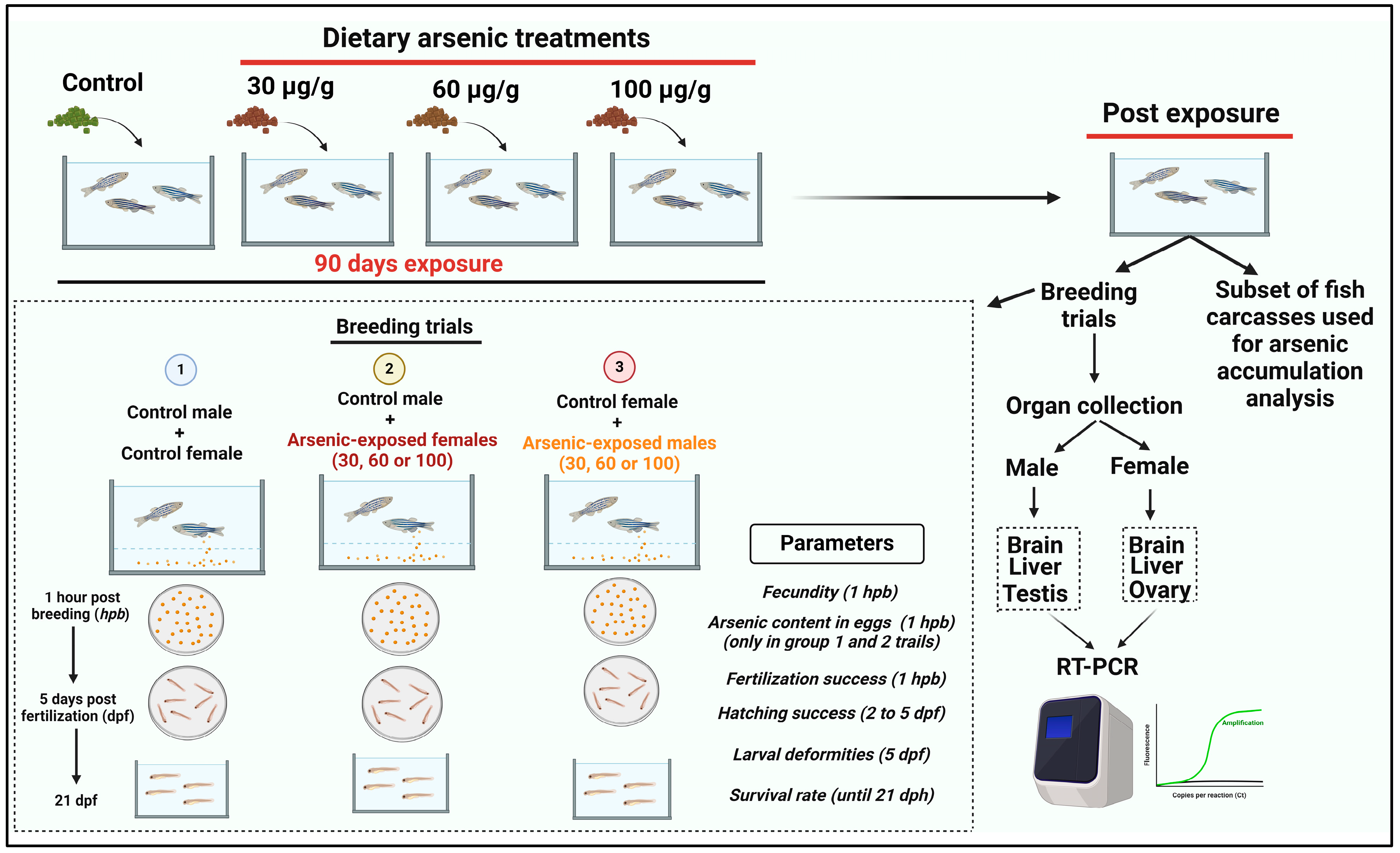
2.1. Fish Maintenance
2.2. Arsenic-Contaminated Diet Preparation and Experimental Exposure
2.3. Assessment of Reproductive Performance and Developmental Effects
2.4. Analysis of Arsenic Concentrations
2.5. Gene Expression Analysis
2.6. Data Processing and Statistical Analysis
3. Results
3.1. Mortality and Arsenic Accumulation in the Tissue and Eggs of Adult Zebrafish Exposed to Chronic Dietary Arsenic
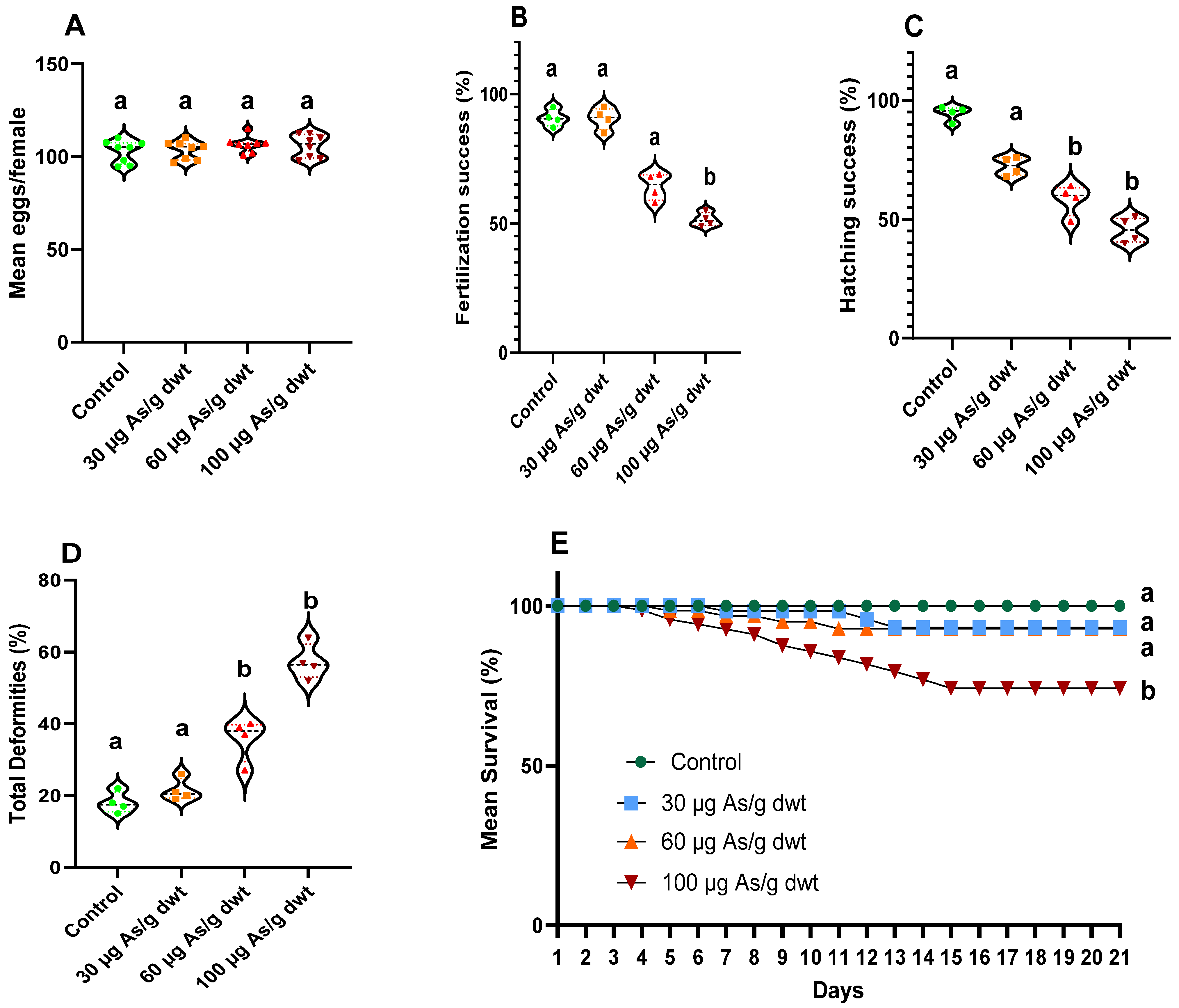
3.2. Reproductive and Developmental Effects of Chronic Arsenic Exposure in Females
3.3. Reproductive and Developmental Effects of Chronic Arsenic Exposure in Males
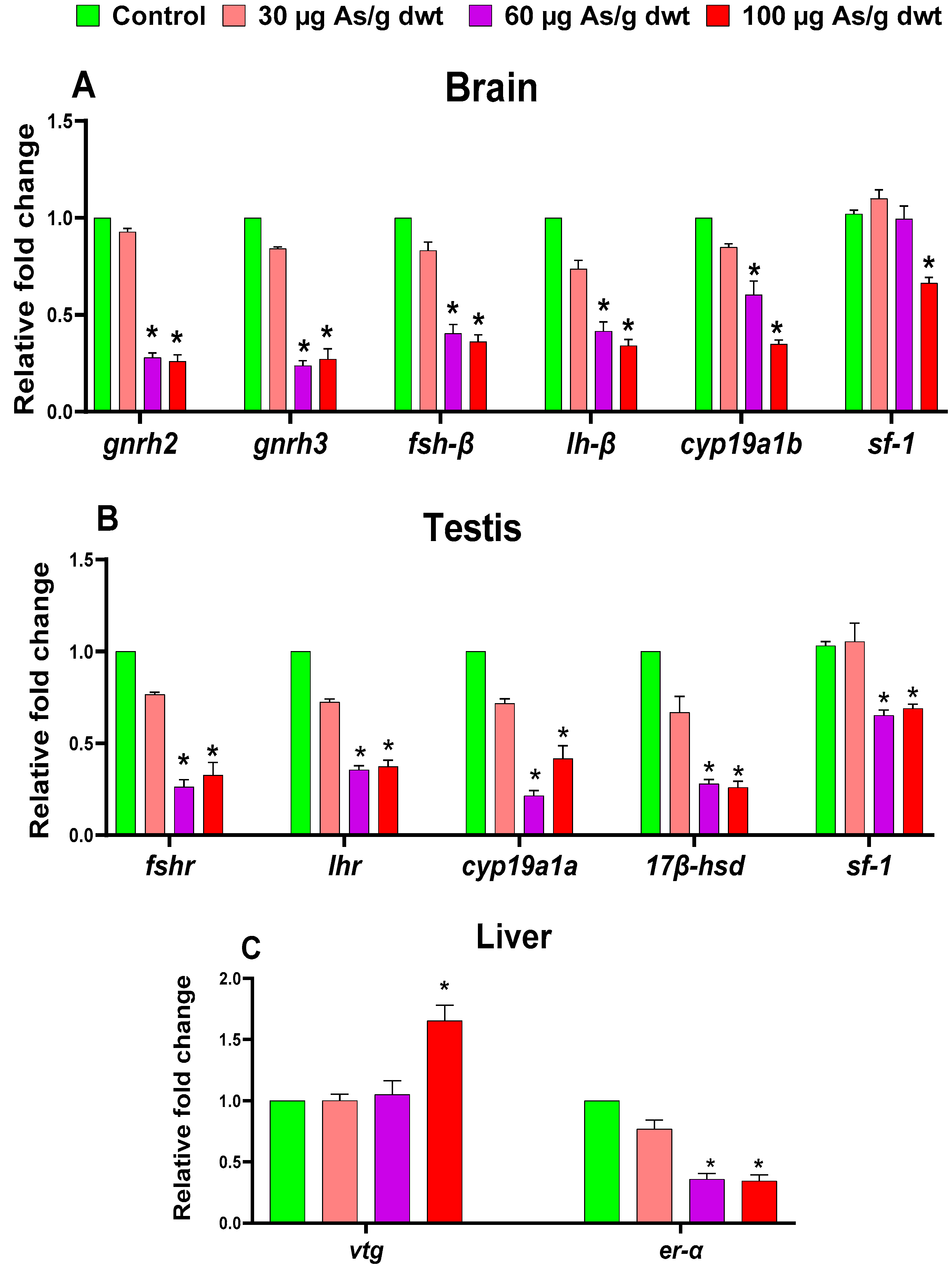
3.4. Sex-Specific Effects of Chronic Dietary Arsenic Exposure on the Expression of HPG-L Axis Genes in Zebrafish
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hughes, M.F.; Beck, B.D.; Chen, Y.; Lewis, A.S.; Thomas, D.J. Arsenic Exposure and Toxicology: A Historical Perspective. Toxicol. Sci. 2011, 123, 305–332. [Google Scholar] [CrossRef] [PubMed]
- ATSDR. Toxicological Profile for Arsenic. 2007. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp2.pdf (accessed on 24 March 2024).
- Kumar, A.; Raj, V.; Srivastava, A.; Ali, M.; Ghosh, A.K.; Rachamalla, M.; Kumar, D. Autophagy in Arsenic Exposed Population and Cancer Patients. In Autophagy and Metabolism: Potential Target for Cancer Therapy; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Kumar, A.; Ali, M.; Raj, V.; Kumari, A.; Rachamalla, M.; Niyogi, S.; Kumar, D.; Sharma, A.; Saxena, A.; Panjawani, G.; et al. Arsenic Causing Gallbladder Cancer Disease in Bihar. Sci. Rep. 2023, 13, 4259. [Google Scholar] [CrossRef]
- Thakur, M.; Rachamalla, M.; Niyogi, S.; Datusalia, A.K.; Flora, S.J.S. Molecular Mechanism of Arsenic-Induced Neurotoxicity Including Neuronal Dysfunctions. Int. J. Mol. Sci. 2021, 22, 10077. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Rachamalla, M.; Kumar, P.S.; Dua, K. Assessment and Impact of Metal Toxicity on Wildlife and Human Health. In Metals in Water; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Sharma, R.; Abubakar, M.D.; Bisht, P.; Rachamalla, M.; Kumar, A.; Murti, K.; Ravichandiran, V.; Kumar, N. Arsenic Exposure and Amyloid Precursor Protein Processing: A Focus on Alzheimer’s Disease. Curr. Mol. Pharmacol. 2023, 17, 1–9. [Google Scholar] [CrossRef]
- Jamwal, A.; Rachamalla, M.; Niyogi, S. Environmental Toxicology of Arsenic to Wildlife (Nonhuman Species): Exposure, Accumulation, Toxicity, and Regulations. In Handbook of Arsenic Toxicology; Academic Press: Cambridge, MA, USA, 2023. [Google Scholar]
- Kumari, B.; Kumar, V.; Sinha, A.K.; Ahsan, J.; Ghosh, A.K.; Wang, H.; DeBoeck, G. Toxicology of Arsenic in Fish and Aquatic Systems. Environ. Chem. Lett. 2017, 15, 43–64. [Google Scholar] [CrossRef]
- Rachamalla, M.; Chinthada, J.; Kushwaha, S.; Putnala, S.K.; Sahu, C.; Jena, G.; Niyogi, S. Contemporary Comprehensive Review on Arsenic-Induced Male Reproductive Toxicity and Mechanisms of Phytonutrient Intervention. Toxics 2022, 10, 744. [Google Scholar] [CrossRef]
- Cottingham, K.L.; Karimi, R.; Gruber, J.F.; Zens, M.S.; Sayarath, V.; Folt, C.L.; Punshon, T.; Morris, J.S.; Karagas, M.R. Diet and Toenail Arsenic Concentrations in a New Hampshire Population with Arsenic-Containing Water. Nutr. J. 2013, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Erickson, R.J.; Mount, D.R.; Highland, T.L.; Hockett, J.R.; Hoff, D.J.; Jenson, C.T.; Lahren, T.J. The Effects of Arsenic Speciation on Accumulation and Toxicity of Dietborne Arsenic Exposures to Rainbow Trout. Aquat. Toxicol. 2019, 210, 227–241. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, A.J.; Wang, N.X.; Li, C.; Sha, J.; Jia, J.; Alessi, D.S.; Yan, B.; Ok, Y.S. Arsenic Bioaccumulation and Biotransformation in Aquatic Organisms. Environ. Int. 2022, 163, 107221. [Google Scholar] [CrossRef]
- Geiszinger, A.E.; Goessler, W.; Francesconi, K.A. The Marine Polychaete Arenicola Marina: Its Unusual Arsenic Compound Pattern and Its Uptake of Arsenate from Seawater. Mar. Env. Res. 2002, 53, 37–50. [Google Scholar] [CrossRef]
- Fattorini, D.; Regoli, F. Arsenic Speciation in Tissues of the Mediterranean Polychaete Sabella spallanzanii. Env. Toxicol. Chem. 2004, 23, 1881–1887. [Google Scholar] [CrossRef]
- Boyle, D.; Brix, K.V.; Amlund, H.; Lundebye, A.K.; Hogstrand, C.; Bury, N.R. Natural Arsenic Contaminated Diets Perturb Reproduction in Fish. Env. Sci. Technol. 2008, 42, 5354–5360. [Google Scholar] [CrossRef]
- Choudhury, B.P.; Roychoudhury, S.; Sengupta, P.; Toman, R.; Dutta, S.; Kesari, K.K. Arsenic-Induced Sex Hormone Disruption: An Insight into Male Infertility. In Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2022; Volume 1391. [Google Scholar]
- Flora, S.J.S.; Agrawal, S. Arsenic, Cadmium, and Lead. In Reproductive and Developmental Toxicology; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Navarro, P.A.A.S.; Liu, L.; Keefe, D.L. In Vivo Effects of Arsenite on Meiosis, Preimplantation Development, and Apoptosis in the Mouse. Biol. Reprod. 2004, 70, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Navarro, P.A.A.S.; Liu, L.; Ferriani, R.A.; Keefe, D.L. Arsenite Induces Aberrations in Meiosis That Can Be Prevented by Coadministration of N-Acetylcysteine in Mice. Fertil. Steril. 2006, 85, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.J.; Kollus, K.M.; Propper, C.R. Environmentally Relevant Arsenic Exposure Affects Morphological and Molecular Endpoints Associated with Reproduction in the Western Mosquitofish, Gambusia affinis. Sci. Total Environ. 2022, 830, 154448. [Google Scholar] [CrossRef] [PubMed]
- Erkan, M.; Aydin, Y.; Yilmaz, B.O.; Yildizbayrak, N. Arsenic-Induced Oxidative Stress in Reproductive Systems. In Toxicology: Oxidative Stress and Dietary Antioxidants; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Raeeszadeh, M.; Karimfar, B.; Amiri, A.A.; Akbari, A. Protective Effect of Nano-Vitamin C on Infertility Due to Oxidative Stress Induced by Lead and Arsenic in Male Rats. J. Chem. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Zargari, F.; Rahaman, M.S.; KazemPour, R.; Hajirostamlou, M. Arsenic, Oxidative Stress and Reproductive System. J. Xenobiot. 2022, 12, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Rachamalla, M.; Salahinejad, A.; Khan, M.; Datusalia, A.K.; Niyogi, S. Chronic Dietary Exposure to Arsenic at Environmentally Relevant Concentrations Impairs Cognitive Performance in Adult Zebrafish (Danio rerio) via Oxidative Stress and Dopaminergic Dysfunction. Sci. Total Environ. 2023, 886, 163771. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.U.; Haider, S.; Tahir, A.; Afsar, T.; Almajwal, A.; Amor, H.; Razak, S. Mechanistic Insight into the Protective Effects of Fisetin against Arsenic-Induced Reproductive Toxicity in Male Rats. Sci. Rep. 2023, 13, 1–13. [Google Scholar] [CrossRef]
- Celino, F.T.; Yamaguchi, S.; Miura, C.; Miura, T. Arsenic Inhibits in Vitro Spermatogenesis and Induces Germ Cell Apoptosis in Japanese Eel (Anguilla japonica). Reproduction 2009, 138, 279–287. [Google Scholar] [CrossRef]
- Tramunt, B.; Montagner, A.; Tan, N.S.; Gourdy, P.; Rémignon, H.; Wahli, W. Roles of Estrogens in the Healthy and Diseased Oviparous Vertebrate Liver. Metabolites 2021, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Hiramatsu, N.; Fujita, T. Vitellogenesis and Choriogenesis in Fishes. Fish. Sci. 2016, 82, 187–202. [Google Scholar] [CrossRef]
- Desesso, J.M. Teratogen Update: Inorganic Arsenic. Teratology 2001, 64, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Ghosh, D. The Involvement of Hypophyseal-Gonadal and Hypophyseal-Adrenal Axes in Arsenic-Mediated Ovarian and Uterine Toxicity: Modulation by HCG. J. Biochem. Mol. Toxicol. 2010, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Bhaumik, S.; Nag Chaudhury, A.; Das Gupta, S. Arsenic Induced Changes in Growth Development and Apoptosis in Neonatal and Adult Brain Cells in Vivo and in Tissue Culture. Toxicol. Lett. 2002, 128, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, J.A.; Da Cunha Martins, A.; Barbosa, F. Teratogenicity, Genotoxicity and Oxidative Stress in Zebrafish Embryos (Danio rerio) Co-Exposed to Arsenic and Atrazine. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 172–173, 7–12. [Google Scholar] [CrossRef]
- Li, X.; Ma, Y.; Li, D.; Gao, X.; Li, P.; Bai, N.; Luo, M.; Tan, X.; Lu, C.; Ma, X. Arsenic Impairs Embryo Development via Down-Regulating Dvr1 Expression in Zebrafish. Toxicol. Lett. 2012, 212, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Zhang, J.Y.; Wang, Q.; Zhu, E.; Chen, W.; Lin, H.; Chen, J.; Hong, H. Environmentally Relevant Concentrations of Arsenite Induces Developmental Toxicity and Oxidative Responses in the Early Life Stage of Zebrafish. Environ. Pollut. 2019, 254, 113022. [Google Scholar] [CrossRef] [PubMed]
- Kubota, R.; Kunito, T.; Tanabe, S.; Ogi, H.; Shibata, Y. Maternal Transfer of Arsenic to Eggs of Black-Tailed Gull (Larus crassirostris) from Rishiri Island, Japan. Appl. Organomet. Chem. 2002, 16, 463–468. [Google Scholar] [CrossRef]
- Ebisuda, K.I.; Kunito, T.; Kubota, R.; Tanabe, S. Arsenic Concentrations and Speciation in the Tissues of Ringed Seals (Phoca hispida) from Pangnirtung, Canada. Appl. Organomet. Chem. 2002, 16, 451–457. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ghosh, S.; Biswas, B.; Pramanik, S.; Nriagu, J.; Bhowmick, S. Epigenetic Modifications from Arsenic Exposure: A Comprehensive Review. Sci. Total Environ. 2022, 810, 151218. [Google Scholar] [CrossRef]
- Panzica-Kelly, J.M.; Zhang, C.X.; Danberry, T.L.; Flood, A.; DeLan, J.W.; Brannen, K.C.; Augustine-Rauch, K.A. Morphological Score Assignment Guidelines for the Dechorionated Zebrafish Teratogenicity Assay. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Brannen, K.C.; Panzica-Kelly, J.M.; Danberry, T.L.; Augustine-Rauch, K.A. Development of a Zebrafish Embryo Teratogenicity Assay and Quantitative Prediction Model. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Salahinejad, A.; Attaran, A.; Meuthen, D.; Rachamalla, M.; Chivers, D.P.; Niyogi, S. Maternal Exposure to Bisphenol S Induces Neuropeptide Signaling Dysfunction and Oxidative Stress in the Brain, and Abnormal Social Behaviors in Zebrafish (Danio rerio) Offspring. Sci. Total Environ. 2022, 830, 154794. [Google Scholar] [CrossRef] [PubMed]
- Kodzhahinchev, V.; Rachamalla, M.; Al-Dissi, A.; Niyogi, S.; Weber, L.P. Examining the Subchronic (28-Day) Effects of Aqueous Cd-BaP Co-Exposure on Detoxification Capacity and Cardiac Function in Adult Zebrafish (Danio rerio). Aquat. Toxicol. 2023, 263, 106672. [Google Scholar] [CrossRef] [PubMed]
- Pedlar, R.M.; Ptashynski, M.D.; Wautier, K.G.; Evans, R.E.; Baron, C.L.; Klaverkamp, J.F. The Accumulation, Distribution, and Toxicological Effects of Dietary Arsenic Exposure in Lake Whitefish (Coregonus clupeaformis) and Lake Trout (Salvelinus namaycush). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 131, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Kubota, R.; Kunito, T.; Tanabe, S. Chemical Speciation of Arsenic in the Livers of Higher Trophic Marine Animals. Proc. Mar. Pollut. Bull. 2002, 45, 218–223. [Google Scholar] [CrossRef]
- Dufour, S.; Sebert, M.E.; Weltzien, F.A.; Rousseau, K.; Pasqualini, C. Neuroendocrine Control by Dopamine of Teleost Reproduction. J. Fish. Biol. 2010, 76, 129–160. [Google Scholar] [CrossRef]
- Simonneaux, V.; Ancel, C.; Poirel, V.J.; Gauer, F. Kisspeptins and RFRP-3 Act in Concert to Synchronize Rodent Reproduction with Seasons. Front. Neurosci. 2013, 7, 22. [Google Scholar] [CrossRef]
- Pierce, J.G.; Parsons, T.F. Glycoprotein Hormones: Structure and Function. Annu. Rev. Biochem. 1981, 50, 465–495. [Google Scholar] [CrossRef] [PubMed]
- Levavi-Sivan, B.; Bogerd, J.; Mañanós, E.L.; Gómez, A.; Lareyre, J.J. Perspectives on Fish Gonadotropins and Their Receptors. Gen. Comp. Endocrinol. 2010, 165, 412–437. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L.; Rice, D.A.; Lala, D.S.; Ikeda, Y.; Luo, X.; Wong, M.; Bakke, M.; Zhao, L.; Frigeri, C.; Hanley, N.A.; et al. Steroidogenic Factor 1: An Essential Mediator of Endocrine Development. Recent. Prog. Horm. Res. 2002, 57, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Hoivik, E.A.; Lewis, A.E.; Aumo, L.; Bakke, M. Molecular Aspects of Steroidogenic Factor 1 (SF-1). Mol. Cell Endocrinol. 2010, 315, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Trant, J.M.; Gavasso, S.; Ackers, J.; Chung, B.C.; Place, A.R. Developmental Expression of Cytochrome P450 Aromatase Genes (CPY19a and CYP19b) in Zebrafish Fry (Danio rerio). Proc. J. Exp. Zool. 2001, 290, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Polzonetti-Magni, A.M.; Mosconi, G.; Soverchia, L.; Kikuyama, S.; Carnevali, O. Multihormonal Control of Vitellogenesis in Lower Vertebrates. Int. Rev. Cytol. 2004, 239, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, R.; Affaticati, P.; Yamamoto, K.; Jolly, C.; Bureau, C.; Baloche, S.; Gonnet, F.; Vernier, P.; Dufour, S.; Pasqualini, C. Dopamine Inhibits Reproduction in Female Zebrafish (Danio rerio) via Three Pituitary D2 Receptor Subtypes. Endocrinology 2013, 154, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Barsagade, V.G. Dopamine System in the Fish Brain: A Review on Current Knowledge. J. Entomol. Zool. Stud. 2020, 8, 2549–2555. [Google Scholar] [CrossRef]
- Zubair, M.; Ahmad, M.; Qureshi, Z.I. Review on Arsenic-Induced Toxicity in Male Reproductive System and Its Amelioration. Andrologia 2017, 49, e12791. [Google Scholar] [CrossRef] [PubMed]
- Akram, Z.; Jalali, S.; Shami, S.A.; Ahmad, L.; Batool, S.; Kalsoom, O. Adverse Effects of Arsenic Exposure on Uterine Function and Structure in Female Rat. Exp. Toxicol. Pathol. 2010, 62, 451–459. [Google Scholar] [CrossRef]
- Chatterjee, A.; Chatterji, U. Arsenic Abrogates the Estrogen-Signaling Pathway in the Rat Uterus. Reprod. Biol. Endocrinol. 2010, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Val, P.; Lefrançois-Martinez, A.M.; Veyssière, G.; Martinez, A. SF-1 a Key Player in the Development and Differentiation of Steroidogenic Tissues. Nucl. Recept. 2003, 1, 8. [Google Scholar] [CrossRef] [PubMed]
- Roy Moulik, S.; Pal, P.; Majumder, S.; Mallick, B.; Gupta, S.; Guha, P.; Roy, S.; Mukherjee, D. Gonadotropin and Sf-1 Regulation of Cyp19a1a Gene and Aromatase Activity during Oocyte Development in the Rohu, L. rohita. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, Y.; Zhao, A.; Cai, X.; Yu, A.; Xu, Q.; Wang, P.; Yao, J.; Wang, Q.; Wang, W. Arsenic Exposure Diminishes Ovarian Follicular Reserve and Induces Abnormal Steroidogenesis by DNA Methylation. Ecotoxicol. Env. Saf. 2022, 241, 113816. [Google Scholar] [CrossRef] [PubMed]
- Taşçi, T.; Eldem, V.; Erkan, M. Sodium Arsenic Alters the Gene Expression of Some Steroidogenic Genes in TM3 Leydig Cell. Celal Bayar Üniversitesi Fen. Bilim. Derg. 2019, 15, 265–270. [Google Scholar] [CrossRef]
- Seif, M.; Abd El-Aziz, T.; Sayed, M.; Wang, Z. Zingiber officinale Ethanolic Extract Attenuates Oxidative Stress, Steroidogenic Gene Expression Alterations, and Testicular Histopathology Induced by Sodium Arsenite in Male Rats. Environ. Sci. Pollut. Res. 2021, 28, 19783–19798. [Google Scholar] [CrossRef]
- Pandey, R.; Garg, A.; Gupta, K.; Shukla, P.; Mandrah, K.; Roy, S.; Chattopadhyay, N.; Bandyopadhyay, S. Arsenic Induces Differential Neurotoxicity in Male, Female, and E2-Deficient Females: Comparative Effects on Hippocampal Neurons and Cognition in Adult Rats. Mol. Neurobiol. 2022, 59, 2729–2744. [Google Scholar] [CrossRef]
- Kishi, H.; Kitahara, Y.; Imai, F.; Nakao, K.; Suwa, H. Expression of the Gonadotropin Receptors during Follicular Development. Reprod. Med. Biol. 2018, 17, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Jana, K.; Jana, S.; Samanta, P.K. Effects of Chronic Exposure to Sodium Arsenite on Hypothalamo-Pituitary-Testicular Activities in Adult Rats: Possible an Estrogenic Mode of Action. Reprod. Biol. Endocrinol. 2006, 4, 9. [Google Scholar] [CrossRef]
- Wang, A.; Holladay, S.D.; Wolf, D.C.; Ahmed, S.A.; Robertson, J.L. Reproductive and Developmental Toxicity of Arsenic in Rodents: A Review. Int. J. Toxicol. 2006, 25, 319–331. [Google Scholar] [CrossRef]
- Machado-Neves, M. Effect of Heavy Metals on Epididymal Morphology and Function: An Integrative Review. Chemosphere 2022, 291, 133020. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.F.; Machado-Neves, M.; Bastos, D.S.S.; Couto Santos, F.; Guimarães Ervilha, L.O.; Coimbra, J.L.d.P.; Araújo, L.d.S.; Oliveira, L.L.d.; Guimarães, S.E.F. Impact of Prenatal Arsenic Exposure on the Testes and Epididymides of Prepubertal Rats. Chem. Biol. Interact. 2021, 333, 109314. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Sato, M.; Suzuki, T.; Nohara, K. Inorganic Arsenic Exposure-Induced Premature Senescence and Senescence-Associated Secretory Phenotype (SASP) in Human Hepatic Stellate Cells. Toxicol. Appl. Pharmacol. 2022, 454, 116231. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rachamalla, M.; Salahinejad, A.; Kodzhahinchev, V.; Niyogi, S. Reproductive and Developmental Effects of Sex-Specific Chronic Exposure to Dietary Arsenic in Zebrafish (Danio rerio). Toxics 2024, 12, 302. https://doi.org/10.3390/toxics12040302
Rachamalla M, Salahinejad A, Kodzhahinchev V, Niyogi S. Reproductive and Developmental Effects of Sex-Specific Chronic Exposure to Dietary Arsenic in Zebrafish (Danio rerio). Toxics. 2024; 12(4):302. https://doi.org/10.3390/toxics12040302
Chicago/Turabian StyleRachamalla, Mahesh, Arash Salahinejad, Vladimir Kodzhahinchev, and Som Niyogi. 2024. "Reproductive and Developmental Effects of Sex-Specific Chronic Exposure to Dietary Arsenic in Zebrafish (Danio rerio)" Toxics 12, no. 4: 302. https://doi.org/10.3390/toxics12040302