Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where do We Stand Now?


Linking Landscape, Environment, Agriculture and Food Research Centre (LEAF), Instituto Superior de Agronomia, University of Lisbon, Tapada da Ajuda, 1349-017 Lisboa, Portugal
Beverages 2018, 4(2), 32; https://doi.org/10.3390/beverages4020032
Submission received: 19 February 2018
/
Revised: 12 March 2018
/
Accepted: 4 April 2018
/
Published: 10 April 2018
(This article belongs to the Special Issue Beverage Sensory Modification)
Abstract
:The unwanted modification of wine sensory attributes by yeasts of the species Brettanomyces bruxellensis due to the production of volatile phenols is presently the main microbiological threat to red wine quality. The effects of ethylphenols and other metabolites on wine flavor is now recognized worldwide and the object of lively debate. The focus of this review is to provide an update of the present knowledge and practice on the prevention of this problem in the wine industry. Brettanomyces bruxellensis, or its teleomorph, Dekkera bruxellensis, are rarely found in the natural environment and, although frequently isolated from fermenting substrates, their numbers are relatively low when compared with other fermenting species. Despite this rarity, they have long been studied for their unusual metabolical features (e.g., the Custers effect). Rising interest over the last decades is mostly due to volatile phenol production affecting high quality red wines worldwide. The challenges have been dealt with together by researchers and winemakers in an effective way and this has enabled a state where, presently, knowledge and prevention of the problem at the winery level is readily accessible. Today, the main issues have shifted from technological to sensory science concerning the effects of metabolites other than ethylphenols and the over estimation of the detrimental impact by ethylphenols on flavor. Hopefully, these questions will continue to be tackled together by science and industry for the benefit of wine enjoyment.
1. Introduction
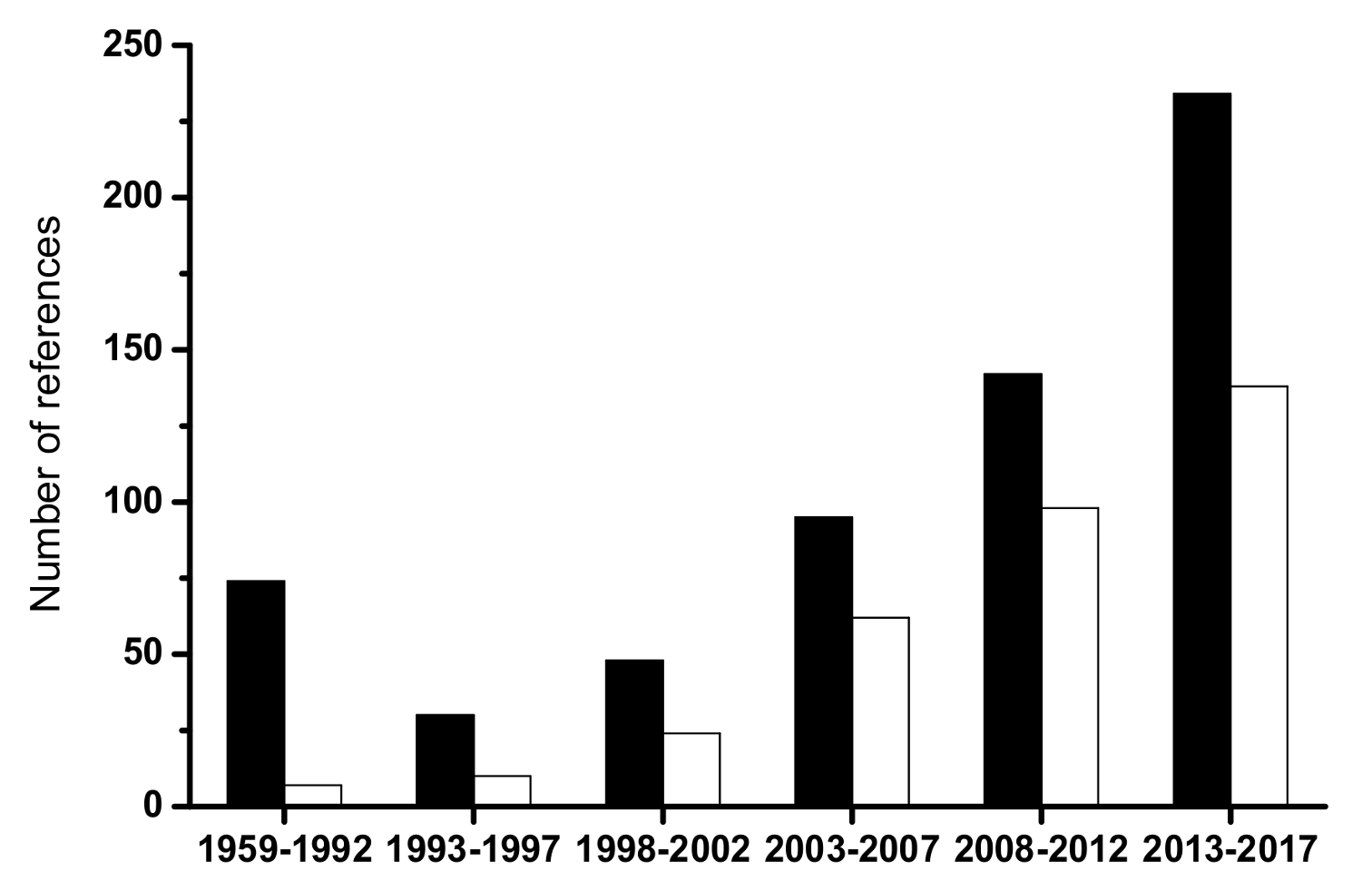
The yeast genus Brettanomyces has been related with the production and characteristics of English beers since the beginning of the XX century [1]. It has been under the attention of early yeast physiologists due to its unusual fermentation stimulation by oxygen, coined as the Custer’s effect [2]. In wines, the first isolates were obtained by Krumholz and Tauschanoff in 1933 without any particular technological concern [1], and the investigation on the genus was relatively scarce until the mid-nineties of last century. To understand the impact of these findings on Brett research, Figure 1 shows the number of peer-reviewed references retrieved from the Scopus database (www.scopus.com, assessed on the 29 January 2018) using “brettanomyces or dekkera and wine” as search words in article titles, abstracts, and keywords from 1959 until 2017. The increase in the numbers coincided with the publication of seminal research by scholars at Bordeaux University [3]. In addition, the total number was 339, which is more than half of the references that use only “brettanomyces or dekkera” (a total of 498). Although involved in other fermented foods, beverages, and ethanol production, most publications recognize that the role played by the species Brettanomyces bruxellensis (anamorph of Dekkera bruxellensis) in red wine spoilage is due to the production of “horse sweat” taint in bulk or bottled wines. Moreover, its effects are particularly notorious in high quality red wines aged in costly oak barrels, which considerably increase the economic losses provoked by spoilage yeasts in the wine industry. Presently, this species is regarded as the main threat posed by yeasts to wine quality [4], surpassing the research interest on the Zygosaccharomyces genus, which is another dangerous wine spoilage yeast. To understand the significance of the relative value of these figures, when the Scopus search executed using the key-words “zygosaccharomyces and wine” and “zygosaccharomyces”, the numbers were 128 and 1122, respectively.
The research of Brettanomyces gathers practically all fields of microbial research (physiology, metabolism, genomics, evolution, taxonomy, etc.) and has been regularly reviewed [1,5,6,7,8,9,10,11,12,13]. The aim of this review was to provide an additional approach that mainly concerns the most relevant scientific achievements and technological impacts, providing an update of the present knowledge and best practices on the prevention of this problem in the wine industry. Future challenges motivated by the sensory perception of volatile phenols will also be addressed because this issue is the subject of extensive debate among wine professionals and the general public.
2. Volatile Phenols (VPs): Their Incidence and Origin
The incidence of VPs in red wines is one of the most frequent wine defects alongside corkiness, reduced, and oxidized off-flavors [14]. The phenolic off-odors have been described as “medicinal”, “phenolic”, “rancid”, “sweaty”, “smoke”, “Band-aid®”, “barnyard” or “horse sweat”. The most important VP is 4-ethylphenol (4-EP), followed by 4-ethylguaiacol (4-EG), and 4-ethylcathecol (4-EC). Earlier works by the team of Pascal Chatonnet during the 1990s provided values for sensory thresholds that are still a reference today. For instance, in Bordeaux red wines, the preference threshold for 4-EP is 620 µg/L, and for the mixture (10:1) of 4-EP and 4-ethylguaiacol is 426 µg/L [3]. However, other thresholds have been published that reflect the influence of other factors among which the wine matrix is essential [15]. For instance, wine body and oak flavor are like buffers of the tainting effect of volatile phenols. In addition, other products of Brett metabolism like isovaleric and isobutyric acids [11] also influence the perception of “horse sweat”, which may explain why the smell of wines spiked with VPs is more objectionable than the smell of wines with the same concentrations but produced naturally.
The precursors of 4-EP, 4-EG, and 4-EC are hydroxycinnamic acids (p-coumaric, ferulic and caffeic acids, respectively), which are enzymatically decarboxylated by a cinnamate decarboxylase, leading to vinyl derivatives and reduced by a vinylphenol reductase, originating in the ethyl derivatives [11].
In grape juices those hydroxycinnamic acids are esterified, mainly to tartaric acid, in concentrations higher than 10 mg/L. In wines they may be present in the free or esterified form, either with tartaric acid [16,17], ethanol [18], hexoses [16] or polyphenols [7]. The release of hydroxycinnamic acids from anthocyanin esters during wine maturation may be only due to chemical reactions [7] but conversions of hydroxycinnamic acid precursors to VPs are typically dependent on enzyme or microbial activity. In grapes or grape juices, the tartaric esters may be hydrolyzed by enzymes from contaminant fungi or from commercial pectolytic preparations, both with cinnamoyl decarboxylase activity, which releases free hydroxycinnamic acid forms [11]. Ethyl and glucose esters of hydroxycinnamic acids may be metabolized by Brettanomyces [16,18] contrarily to tartaric acid esters [16,17]. Most of these tartaric esters are hydrolyzed only after malolactic fermentation, and it has been hypothesized that the hydrolytic activity of the lactic acid bacteria follows the completion of malic conversion to lactic acid [19,20]. Thus, the pool of hydroxycinnamic acids’ precursors provides the substrate for the production of VPs that are higher than the preference thresholds depending on the presence of active B. bruxellensis populations [20,21]. S. cerevisiae may produce vinyl derivatives that may be later reduced by B. bruxellensis. This species is highly efficient in the conversion of p-coumaric acid with molar rates higher than 90% [22]. Then, the natural pool of hydroxycinnamic acids (>10 mg/L) in wines are not likely to be a limitation for the production of VPs higher than the preference thresholds. Therefore, all red wines may be affected by the taint, given that the yeasts are able to grow. The belief that there are some grape varieties more susceptible to Brett growth has no scientific evidence [23].
The absence of “horse sweat” in white wines is probably due to the easier Brett inactivation, given that hydroxycinnamic acids are also present in concentrations similar to those of red wines [24]. The yeast species Pichia guilliermondii and lactic acid bacteria (Lactobacillus spp., Pediococcus spp.) also have the ability to produce 4-EP, but their spoiling potential in wines is not comparable to that of B. bruxellensis [25,26,27].
In conclusion, the natural concentrations of hydroxycinnamic acids in wines are high enough to provide substrate for the production of volatile phenols much above the preference thresholds. Therefore, the key for the prevention of the problem is to understand the ecology and the behavior of Brettanomyces in wines in order to apply the adequate control measures and avoid, or reduce, the conversion of hydroxycinnamic acids to VPs.
3. Brett Ecology: Infection Routes in the Winery
The Brettanomyces yeasts are mostly associated with fermented products, particularly with the post-fermentation or aging period of alcoholic beverages, like wine, beer, cider, kombucha, and tequila. Some reports refer their isolation from cheeses or fermented milks. Besides the food industries, Dekkera/Brettanomyces spp. have also been reported in industrial ethanol fermentations. These yeasts are scarcely mentioned outside of fermentation environments. Occasional reports include isolation from olives, flours, carbonated beverages, and bees and air at ground level in fruit orchards, honeys, and tree exudates [5]. Being a fermenting species, Brett probably shares the same natural habitat as S. cerevisiae (e.g., soil, oak bark, decaying vegetal tissues, and tree exudates) but it is even rarer. Despite the preference for fermented products, Dekkera/Brettanomyces spp. are usually not dominant and are reported in a low percentage of analyzed samples. Their occasional detection in sparkling wines may be related with their resistance to carbon dioxide, which is higher than that of S. cerevisiae and Z. bailli [28]. Thus, Brett can be regarded as sporadic contamination yeast that appears in high numbers mainly when other microorganisms have been inhibited.
The dissemination of B. bruxellensis in wine related environments is hard to evidence from sources contaminated by other yeasts due to its low growth rate. As a consequence, the use of selective media and long incubation periods are essential to its recovery [29], although even when using selective media, it has been rarely isolated from grapes [30] and clean winery environments. Rotten grapes appear to bear higher populations, which most likely explains the higher incidence of phenolic wines in vintages affected by this problem [31]. The frequency of isolation from cellar equipment increases in the presence of juice, wine residues, and leftovers (husks and pomaces). Insects and air dusts are probable vehicles of dissemination among infecting spots [32]. In contrast, it is a common contaminant easily recovered from red wines where numbers may attain high levels (>104 cells/mL) if preventive measures are not taken [33].
The frequency of isolation in red wines explains why careful vigilance must be given to wines purchased from other sources or sold as bulk wines to other wineries. Once established in a winery, Brett is very difficult to eradicate, especially when it has contaminated the wooden vats.
Old Wooden Vats: A Well-Known Ecological Niche
The traditional wooden vats used for red wine, sherry, lambic beer or cider production are typical niches of B. bruxellensis. In wines, the onset of problems related with its activity coincided with the worldwide increase in the utilization of oak barrels during the last 25 years. However, wines matured in stainless steel tanks are also affected by these yeasts. The survival in barriques is facilitated by the diffusion of oxygen either by stimulating yeast growth [34] or by reducing the levels of molecular sulfite active against yeasts. In addition, it is believed that cell immobilization in the wood structure contributes to the protection against preservatives. The ability to form pseudomycelium might as well favor the colonization of the porous structure of the wood and of the spaces between the staves and the grooves. The wood of new barriques is not the source of Brett but it is readily contaminated if filled with infected wine. On the contrary, high ethanol wines (>17% v/v) are not susceptible to these yeasts even if aged in old wooden vats, such as in the case of sherry or port-style wines.
Overall, the incidence of Brett increases during grape processing, from fermentation to red wine ageing. The routes of contamination may not be perfectly known but this yeast easily finds the way to infect stored red wines and initiates spoilage if not stopped in due time.
4. Brett Behavior and Tolerance in Wines
The practical absence of Brettanomyces from the early stages of grape processing is no longer observed after the end of malolactic fermentation. In certain cases its activity may already begin in the period between alcoholic and malolactic fermentations. It is not as tolerant to ethanol or sulfite as S. cerevisiae or Zygosaccharomyces bailii [35] but it has the ability to remain viable for long periods. The increase in their predominance appears to be the result of an exceptional resistance to minimal nutrient conditions, which is seen as the determinant in their survival during the production and storage steps and in their development once the environment becomes favorable (e.g., reduction in free sulfite during ageing) [11,30]. For instance, viable Brettanomyces have been reported in red wine bottled for more than 50 years [33].
4.1. Attention to the Unnoticed Presence of Brett
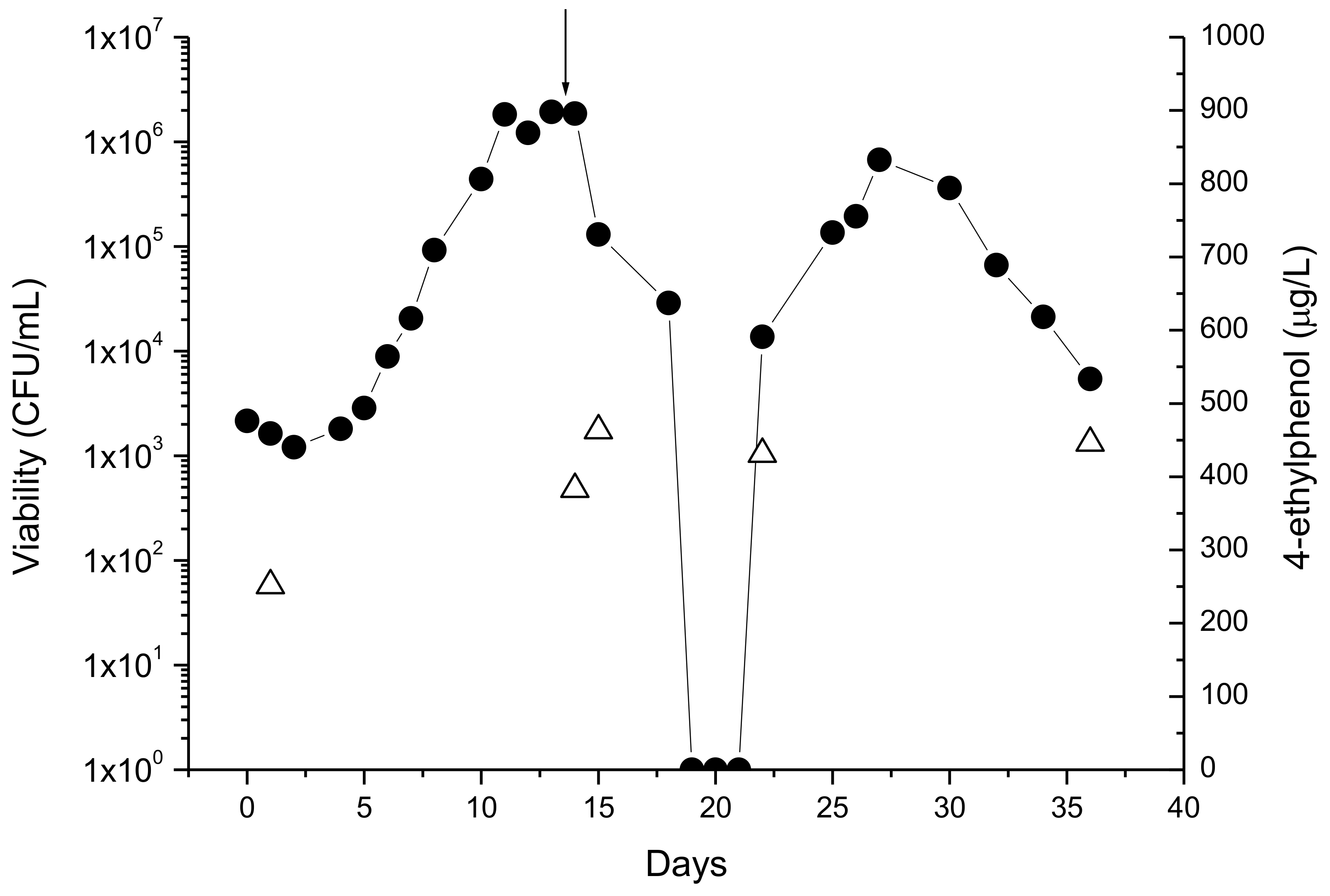
The behavior of D. bruxellensis in co-culture with S. cerevisiae (molasses, grape juice, and synthetic medium) is characterized by showing null or slow growth until about the end of fermentation. In post-fermentation it may grow, attaining levels as high as those observed with S. cerevisiae [22]. After inoculation in wines, D. bruxellensis growth shows a typical bell-shaped curve, with exponential growth followed by cell death. When the stress imposed to cells is high, a sharp decrease in viability is frequently observed and the wine seems to be Brett-free [24]. In fact, cells may be in a viable but non-culturable state (VBNC cells) [36]. However, these observations may well result from the fact the sample volume analyzed was too low to recover viable cells. Whatever the explanation, the technological significance of these observations is that after an apparent Brett-free period the growth may be re-initiated either by surviving or VBNC cells, as shown in Figure 2.
4.2. Resistance to Antimicrobials
Sulphur dioxide is the most common and effective preservative utilized in wineries. However, levels may be as high as 40 mg/L of free sulfite at pH 3.5, to control D. bruxellensis in wines aged in barrels [24]. The active form is the molecular one, and so the lower the pH the higher the proportion of molecular sulfite. The effective free sulfite values in red wines may look somewhat high but probably reflect the proportion of sulfite bound to the anthocyanins that are counted as free sulfite by the current titration methods. The strategy should be directed to increase the ratio of free to bound sulfite so that inhibitory levels may be reached under a lower concentration of total sulfur dioxide.
Attempts to reduce sulfite utilization boosted the search for commercial alternatives. Sorbic acid has long been known to act against fermenting yeasts in bottled wines but D. bruxellensis is resistant to the maximum legal concentration of 200 mg/L in wines. Presently, the focus has been turned to dimethyldicarbonate (DMDC) and chitosan, which are now commercially available. Other antimicrobials, like killer proteins and peptides of microbial origin (e.g., zymocins) are reported as effective in research articles but are not yet spread in the industry [37,38].
DMDC is an effective agent against Brett growth, depending on the initial cell concentration [39] (Table 1). In wines matured in barrels it is an efficient tool to prevent blooms, being used in regular additions up to the maximum permitted level of 200 mg/L. In the EU it is only authorized just before bottling, in wines with more than 5 g/L of sugar, and it is claimed to be efficient if yeast counts that are less than 500 CFU/mL. The efficiency also depends on adequate DMDC homogenization, which requires the use of specific equipment that makes the treatment cost high (>0.05 €/bottle). In addition, DMDC hydrolysis releases methanol that should be monitored if excessive concentrations are suspected.
Chitosan is a natural derivative of chitin and prevents Brett growth in wines with variable efficiency [40]. As with sulfite or DMDC, the important is to ensure that cells do not recover viability after the death phase as shown in Figure 2. The legal limit (0.1 g/L to inactivate Brett) is enough when contaminations are low but it is not as effective when cells are present in high numbers and growing. The product is not cheap (>0.05 €/L) and careful examination of the balance between costs and benefits should be done before application.
4.3. The Hurdle Concept in Food and Wine Preservation
The judicious utilization of sulfite is the key to preventing the building up of D. bruxellensis contaminations, however, sometimes the edge of its OIV guideline legal limit of utilization in red wines is reached (150 mg/L in red wines with less than 4 g/L reducing sugar, 300 mg/L in wines with more than 4 g/L reducing sugar) (http://www.oiv.int, accessed on the 11 February 2018). Then, DMDC or chitosan may be used to minimize the utilization of sulfur dioxide. These chemical preservatives are not the only options. The winemaker may also choose to control Brett using physical methods. The overall concept of spoilage prevention is known as the hurdle concept in food microbiology. The idea is to weaken microbial populations by making them to “jump” several hurdles. The more hurdles to jump the easier it would be to prevent microbial growth. In wineries, these hurdles include environmental factors (e.g., storage temperature, dissolved oxygen) and processing factors (e.g., fining, filtration, heat treatments, high pressure, pulse electric fields, preservatives) [41,42,43] that when applied correctly contribute to decreasing the utilization of sulfur dioxide.
In conclusion, Brett inactivation may be achieved by several alternative processes. The effectiveness of the different options must be ascertained under each real condition by appropriate monitoring strategies.
5. Brett Prevention: How to Monitor Contaminations
Wine technologists have two different attitudes when facing the threat of D. bruxellensis. One, which we can call “optimistic”, results from the absence of wines spoiled by the yeasts and the thought that it only happens to the others. The other, which we can call “pessimistic”, results from traumatic experiences in the past. Naturally, neither are the correct attitudes. The first runs under high risks that will, eventually, bring disastrous consequences. The second leads to exaggerated precautions and to high costs, either economically or in wine quality. Assuming that these yeasts are always present, although not detected, it is necessary to learn how to live with them in the winery and apply the best preventive measures.
5.1. To Know Where They Are and How Fit They Are
The first step to live with Brett is to know where it is. The proper prevention of Brett activity depends on its detection by microbiological analysis and to decide the most adequate treatment. Today, several microbiological methods are available to monitor B. bruxellensis periodically, including highly specific molecular methods [44,45,46]. The most common and suitable technique is plate counting. However, many wineries do not have regular microbiological control. The small dimension of wine enterprises is a serious limitation to the development of a routine microbial control. The costs of equipment from the simplest for plate counting to advanced instruments (e.g., fluorescence microscope, real-time polymerase chain reaction (PCR), flow cytometer) and the requirement for skilled labor are still a burden for most small and medium enterprises. Therefore, the economic losses associated with D. bruxellensis activity have moved many companies to ask for external support, which may be easily found today.
In our lab routine, we currently apply a simplified detection technique based on growth in a selective solid or liquid medium. Serial dilutions of wine samples are inoculated in plates or test tubes, incubated at room temperature and the results checked after 4 to 15 days. These microbial determinations are accompanied by 4-EP determination and gas chromatography as a measure of the spoiling activity.
The microbiological analysis should be regarded at two levels: (i) when the wines are stored in bulk, in oak barrels or in tanks; and (ii) when the wine is to be bottled. In the first situation, the main purpose is to avoid the production of VPs in levels high enough to produce off-flavors and off-tastes. Thus, it is not mandatory to eliminate Brett completely, but to assure that the level of contamination or of activity is low enough to keep 4-EP levels constant. In the second situation, the main purpose is to have bottled wine free from these yeasts. Only one viable cell per bottle may be the cause for spoilage much later.
5.2. Microbial Guidelines
In bulk stored wines, it is satisfactory to detect B. bruxellensis monthly, bimonthly or even every three months, depending on the contamination history. The sample volumes are from 100, 10, 1, to 0.1 mL, from a blend composed by wine from the interface air/liquid and from different depths of the container. In case the result is positive for 1 mL, or less, and the level of 4-EP is higher than 150 μg/L, it is recommended to reduce the microbial populations by fine filtration (<1 µm) immediately, accompanied by sulfite addition [35]. Negative results in 1 mL mean that contamination is low and only fining followed by sulfite should be enough. Chitosan, when contamination is low, or thermal treatments, when numbers are high, are two of the alternatives during wine ageing. The effectiveness of the treatments must be ascertained by microbiological analysis. When 4-EP levels are stable there is no need to reduce contamination because cells may be present but are not active, thus reducing the additions of sulfite during storage.
For wines before bottling, the criteria are more stringent, and detection should be made on 100, 10, and 1 mL of wine, sampled as described above. In case the result is positive in 1 or 10 mL, it is recommended a very fine or sterilizing filtration. If positive detection is only obtained for 100 mL, it is admissible to control viable cells only by the addition of preservatives (e.g., 40 mg/L of free sulfite, at pH 3.50). In this case, bottling must be technically correct and dissolved oxygen should be lowered to practically zero. Otherwise, it may be recommended to use DMDC or, as an alternative, a thermal treatment to destroy viable cells [35].
5.3. The Question of Real Time PCR
Given the slow growth of these yeasts, cultured media can only give results after more than 4–5 days and so early detection depends on the use of direct techniques. Presently, there are several real time PCR protocols that provide results in about 4–6 h and have a high sensitivity (<10 cells/mL) [45,46]. They have two main drawbacks. One is the cost, which is very high for a routine analysis (>60 €/sample). The second is related with false positive responses given by the DNA of dead cells. In this case, protocols must be adapted to remove this DNA from the samples [47].
6. Brett Prevention: How to Control, Kill, and Cure
Knowing the monitoring and treatment options available, we will describe below our main observations and decisions during our empirical experience in wineries. The first idea is always to reduce cell numbers and growth so that treatments may be limited as much as possible. If these measures are not taken in due time, VPs may attain high levels and curative measures may be used.
6.1. Reduction of Dissemination
The infections come into the winery through the grapes, wines, insects, and used barriques. In the case of grapes, it is not efficient to analyze and separate the infected grapes. Assuming that the prevalence of D. bruxellensis is higher in vintages of poor sanitary quality grapes, care should be taken to (i) minimize the time between alcoholic and malolactic fermentation and (ii) avoid cross contaminations that may jeopardize other finished wines. Outsourced red wines must be monitored before blending and treated adequately (see below). Insect dissemination through winery atmosphere, during harvest or during bottling, must be minimized. New barriques do not contain Brettanomyces, but used barriques, when recovered, are a most probable infection source for the barreled wines. However, new barriques may be even more suitable to support Brett growth because of higher oxygen permeability and nutrient release from the wood [6].
Once inside the winery, the most important factor to reduce dissemination is proper hygiene to avoid cross contaminations when moving wine. Common disinfectants used in the food industry are effective against yeast species and also against Brettanomyces [48]. The most efficient products are alkaline detergents, iodophors, and peracetic acid based sanitizers. Thus, the main concern is how to sanitize, properly, points of complex geometry or difficult access, particularly in bottling machines, like dead ends of filters, valves, gauges, or hoses. Pumps and hoses for wine transfer between tanks is another concern. Less efficient, or virtually impossible, is the sterilization of wooden vats common in traditional fermentation processes and in modern fashionable wine ageing. Sanitation with hot water or steam is essential, although not completely efficient. Even after steaming, Brett may be recovered from wood layers up to 4–6 mm below the surface [49]. Whatever the treatment adopted in wineries, one should bear in mind that, in barrels, the critical factor is the inability of the disinfecting agent (e.g., hot water, steam, ozone, microwaves, UV light) to reach the deeper layers of the wood.
6.2. Prevention of D. bruxellensis Growth and 4-EP Production
The prevention first depends on careful D. bruxellensis monitoring during all wine storage time. It should begin after malolactic fermentation or even before, when extended periods occur between the end of wine fermentation and the onset of malolactic fermentation. During this period wines are left unprotected by sulfite and kept at higher temperatures to promote the bioconversion of malic acid, which stimulate premature Brett growth. After malolactic fermentation, the frequency of analysis depends on its detection, but even in the absence of contaminations at least three analyses per year (after winter, before harvest, after harvest) should be done to avoid unnoticed growth [35]. This frequently happens during the harvest period, when temperatures are higher and attention is directed to pick grapes and ferment wines.
The main concern when a contaminated sample appears is to avoid any cross contamination. This is especially important when processing products from external sources, such as purchased wines of unknown origin. The best solution, at first, is to detect the presence of D. bruxellensis to determine the measures to be taken. In case wines are highly contaminated, the most efficient measure would be to filter through pores tighter than 1.0 µm, or even 0.8 µm, given that sulfur dioxide reduces cell size [50]. Even knowing the difficulty to sterilize by filtration young wines and the controversy of such measures, our opinion is that it should be considered, particularly when wines are to be matured in costly oak barrels.
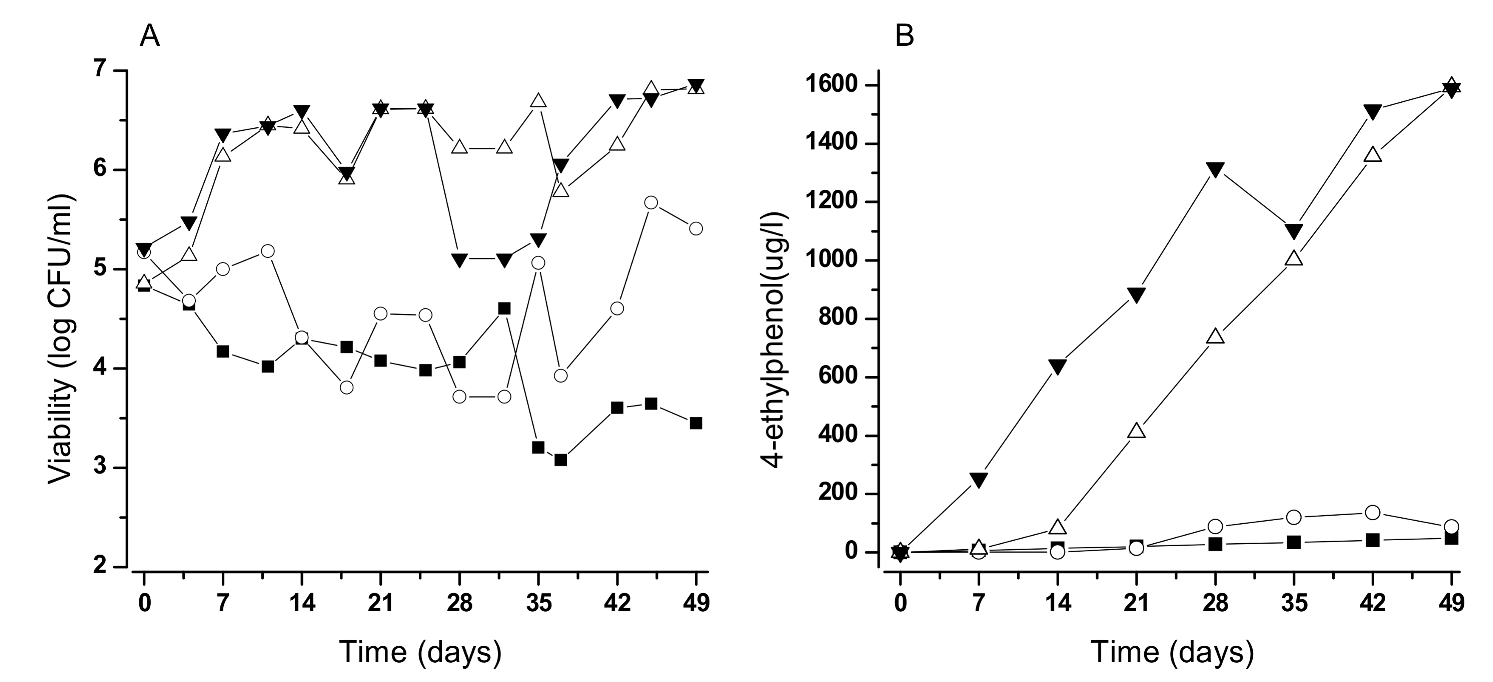
Concerning factors promoting wine colonization, special attention should be paid to the levels of free sulfite, levels of dissolved oxygen, presence of residual sugars, and the storage temperature. Ideally, if wines could be stored at less than 10 °C there would be no yeast growth and concurrently no noticeable 4-EP production (Figure 3).
Oxygen is essential to wine ageing but it also stimulates growth and the production of volatile phenols [34]. Even under the practical absence of oxygen D. bruxellensis grows and produces 4-ethylphenol but at lower rates that are enough to affect wine quality. Therefore, all operations contributing to oxygen diffusion must be minimized, or carefully monitored, like rackings, pumpings, toppings, bottling and, particularly, micro-oxygenation—a fashionable process to accelerate red wine ageing. Oxygen also contributes to a quicker loss of free sulfites during storage, opening a window of opportunity for Brett growth. In our experience, the most frequent reason for unexpected growth is the drop of free sulfites below 20 mg/L (at pH 3.5).
Another aspect to keep in mind is the level of nutrients in the wine. These yeasts may grow well under very low levels of residual sugar (<2 g/L and in wines where no nutrients were added). However, reducing nitrogen additions to the minimum required to finish fermentations is always a wise strategy to minimize Brett growth during storage. The idea is to create a “nutrient desert” that turns the wine less susceptible to yeast growth. Most likely this is the mechanism underlying inhibition by Metschnikowia pulcherrima, when this species, by producing pulcherriminic acid, depletes the iron present in the medium, making it unavailable to the other yeasts [51]. During bottling, a sugar addition to smooth mouth-feel up to 10 g/L does not increase susceptibility to spoilage in wines with high ethanol, and so extra doses of sulfur dioxide to compensate higher sugar are not required, which is contrary to common knowledge [52]. However, if cells have the ability to grow, higher residual sugar stimulates proliferation [53].
The option to kill actively growing populations through heat treatments using mild temperatures in low volume tanks (35 °C overnight) or pasteurization regimes at bottling can be successful, because Brett is not heat resistant. A solution for contaminated bottled wine is to keep the bottles at 35 °C and check until viable counts fall to 0/bottle.
6.3. Curative Measures
When the wine is off-tainted, there are no effective curative measures without depreciating it. In this situation, we always weigh the possibility of blending tainted wine with “clean” wine. Although this measure may attenuate the defect of the tainted wine by dilution, it cannot be seen as a curative measure. In fact, mixtures of wines free of 4-EP are only effective for small proportions of tainted wines, because large volumes of “clean” wine must be used to obtain a blend with 4-ethylphenol levels lower than the preference threshold.
The effective reduction of 4-EP levels may be obtained by adsorbents (e.g., yeast lees, fining agents, activated charcoal) [54] but wine favorable aroma compounds or color are also removed and a balance must be drawn between the benefits and losses of wine attributes. Our experience with activated charcoal reveals that it may be an efficient option when preventive measures have failed (Figure 4). In this case, besides a reduction in VPs, the wine was also reduced in color and flavor intensity but it was perfectly adequate for blending with un-tainted wines. The use of nanofiltration together with activated charcoal (process accepted by the OIV) and reverse osmosis are other alternatives claimed to be effective in VP reduction. It is also possible that esterified cellulose may be used in filtration sheets to decrease volatile phenols [55].
7. The Brett Sequel: from “Terroir” to “Terror”
The flavors responsible for the Brett character were certainly known for a long time. It is feasible that it contributed to the so-called “gout du terroir”, used to describe certain tainted wines in France by the mid 20th century. Bulk Californian red wines by 1960–1970 were also frequently affected by this taint (Ralph Kunkee, personal communication). In our case, the awareness of the problem begun in the last decade of the last century, when some problematic red wines were tasted by Prof. Dennis Dubourdieu in a workshop organized by our faculty. Coincidently, wine became a fashionable product, worthy of global exposure in the USA. It was a time when noticeably tainted phenolic Bordeaux wines were taken as the expression of terroir by leading wine critics. In addition, barrique utilization increased worldwide to cope with consumer preferences and increasing Brett infection risks. Before winemakers could deal with the problem it was already the object of lively discussions in specialized journals and consumer related websites.
A complex Brett aroma wheel was recently published [56], describing quite variable descriptors, not all unpleasant, such as earthy, leather, savory, spicy or woody, that may be found in high quality red wines. Despite this complexity, the result of generalized “horse sweat” awareness is that, today, every nose seems to be particularly appropriate to find an off-flavor. In fact, the recognition of volatile phenols seems to be equivalent in individuals from very different origins or backgrounds [15]. However, the question is not the recognition of the flavor but the fact that at the least perception of animal notes, the wine may be readily incriminated as an excessively barnyard tainted wine. Particularly in fine wines, bottled for decades, the aging bouquet may have notes resembling volatile phenols, albeit being present in low levels. In a world of uniform globalized taste, dominated by the pleasantness of intense fruity-oak flavors and a full-sweet mouthfeel, there seems to be little tolerance to wines with leather-game scents. As a result, winemakers are haunted by the terror of having the slightest note imparted by volatile phenols and become easy victims of technical alternatives that are frequently ineffective to calm their nightmares.
The Issue of Chemical Limits
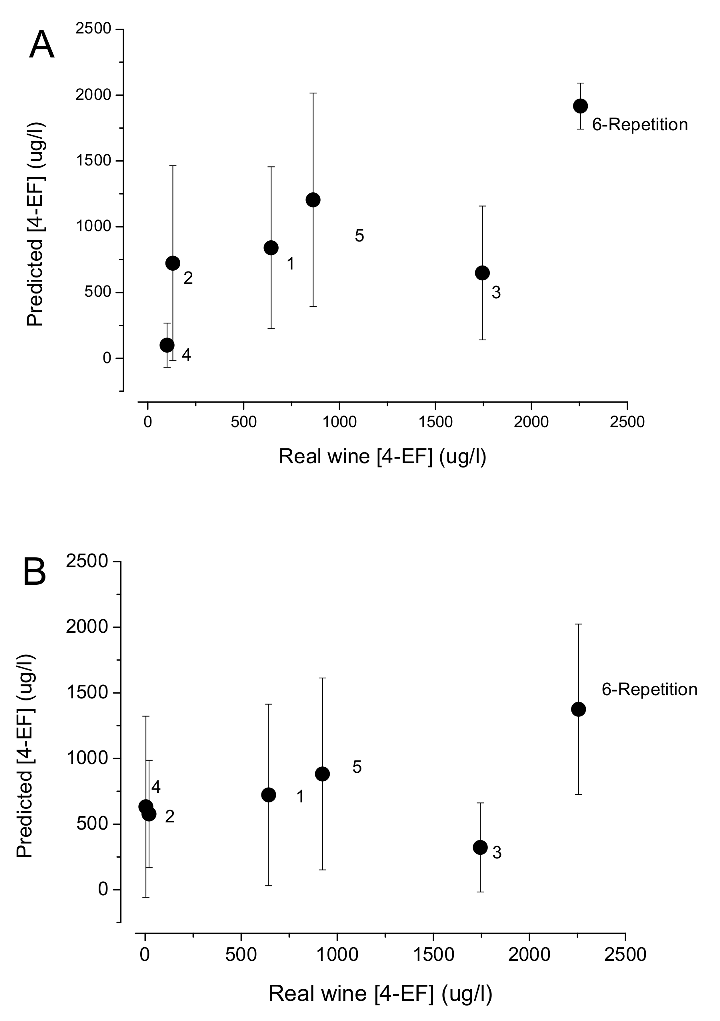
Given that volatile phenols, where 4-ethylphenol is by far the most relevant, are regarded as spoilage molecules, it is expected that the wine regulatory bodies and large retail companies will try to establish limits for the acceptability of wines based on the average sensory detection threshold of 500 µg/L of 4-ethylphenol. This can be compared with the classic limits for volatile acidity, which is a chemical indicator of spoilage by acetic acid bacteria. However, threshold values for volatile phenols must be regarded only as indicative of horse-sweat taint. Given the different aromatic integration of 4-ethylphenol smell in the overall wine flavor, well-known by wine professionals and a concept explained by Clark Smith in his book Postmodern Winemaking [57], high concentrations are frequently not detected by trained tasters (Figure 5). Therefore, clearly tainted wines can only be determined by sensory analysis. The odorant chemical concentration can only be directly related with Brett activity, as described before.
8. Final Remarks
According to our experience, the control of B. bruxellensis spoilage is, presently, the most serious microbial problem in red wine quality and poses serious constraints to sulfite reduction in its production. There are several technical alternatives that prevent or kill contaminant populations, which contribute to a decrease in its utilization, but they are not fully effective when used alone. Winemakers should learn how to live with these yeasts by monitoring their activity, applying appropriate inactivation measures only when necessary, and weighing the cost-benefit of each option. Besides their technical expertise, winemakers should also communicate to consumers and other wine professionals the difference between volatile phenol sensory detection and wine depreciation, enabling them to understand the diversity and aptitudes of wines with animal-leather flavors.
Acknowledgments
Research was funded by the Fundação para a Ciêcnia e Tecnologia (FCT) trough project UID/AGR/04129/2013.
Conflicts of Interest
The author has no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Schifferdecker, A.J.; Dashko, S.; Ishchuk, O.P.; Piškur, J. The wine and beer yeast Dekkera bruxellensis. Yeast 2014, 31, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, W.A.; Wikén, T.O. The Custers effect (negative Pasteur effect) as a diagnostic criterion for the genus Brettanomyces. Antonie Leeuwenhoek 1969, 35, A31–A32. [Google Scholar]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.-N.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Curtin, C.; Varela, C.; Borneman, A. Harnessing improved understanding of Brettanomyces bruxellensis biology to mitigate the risk of wine spoilage. Aust. J. Grape Wine Res. 2015, 21, 680–692. [Google Scholar] [CrossRef]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage activities of Dekkera/Brettanomyces spp. In Food Spoilage Microorganisms; Blackburn, C., Ed.; Woodhead Publishers: Cambridge, UK, 2006; Chapter 13; pp. 354–398. [Google Scholar]
- Renouf, V.; Lonvaud-Funel, A.; Coulon, J. The origin of Brettanomyces bruxellensis in wines: A review. J. Int. Sci. Vigne Vin 2007, 41, 161–173. [Google Scholar] [CrossRef]
- Suárez, R.; Suárez-Lepe, J.A.; Morata, A.; Calderón, F. The production of ethylphenols in wine by yeasts of the genera Brettanomyces and Dekkera: A review. Food Chem. 2007, 102, 10–21. [Google Scholar] [CrossRef]
- Oelofse, A.; Pretorius, I.S.; du Toit, M. Significance of Brettanomyces and Dekkera during Winemaking: A Synoptic Review. S. Afr. J. Enol. Vitic. 2008, 29, 128–144. [Google Scholar] [CrossRef]
- Wedral, D.; Shewfelt, R.; Frank, J. The challenge of Brettanomyces in wine. LWT Food Sci. Technol. 2010, 43, 1474–1479. [Google Scholar] [CrossRef]
- Zuehlke, J.M.; Petrova, B.; Edwards, C.G. Advances in the control of wine spoilage by Zygosaccharomyces and Dekkera/Brettanomyces. Annu. Rev. Food Sci. Technol. 2013, 4, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K. Brettanomyces yeasts—From spoilage organisms to valuable contributors to industrial fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.D.; Divol, B. Brettanomyces bruxellensis, a survivalist prepared for the wine apocalypse and other beverages. Food Microbiol. 2016, 59, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Agnolucci, M.; Tirelli, A.; Cocolin, L.; Toffanin, A. Brettanomyces bruxellensis yeasts: Impact on wine and winemaking. World J. Microbiol. Biotechnol. 2017, 33, 180. [Google Scholar] [CrossRef] [PubMed]
- Goode, J.; Harrop, S. Wine Faults and Their Prevalence: Data from the World’s Largest Blind Tasting. In Proceedings of the 20th Entretiens Scientifiques Lallemand, Horsens, Denmark, 15 May 2008. [Google Scholar]
- Schumaker, M.; Chandra, M.; Malfeito-Ferreira, M.; Ross, C. Influence of Brettanomyces ethylphenols on red wine aroma evaluated by consumers in the United States and Portugal. Food Res. Int. 2017, 100, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Hixson, J.; Hayasaka, Y.; Curtin, C.; Sefton, M.; Taylor, D. Hydroxycinnamoyl Glucose and Tartrate Esters and Their Role in the Formation of Ethylphenols in Wine. J. Agric. Food Chem. 2016, 64, 9401–9411. [Google Scholar] [CrossRef] [PubMed]
- Schopp, L.; Lee, J.; Osborne, J.; Chescheir, S.; Edwards, C. Metabolism of Nonesterified and Esterified Hydroxycinnamic Acids in Red Wines by Brettanomyces bruxellensis. J. Agric. Food Chem. 2013, 61, 11610–11617. [Google Scholar] [CrossRef] [PubMed]
- Hixson, J.L.; Sleep, N.R.; Capone, D.L.; Elsey, G.M.; Curtin, C.D.; Sefton, M.A.; Taylor, D.K. Hydroxycinnamic Acid Ethyl Esters as Precursors to Ethylphenols in Wine. J. Agric. Food Chem. 2012, 60, 2293–2298. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, M.J.; Torres, M.; Palma, V.; Alves, E.; Patão, R.; Costa Freitas, A.M. Impact of malolactic fermentation on low molecular weight phenolic compounds. Talanta 2008, 74, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Chescheir, S.; Philbin, D.; Osborne, J.P. Impact of Oenococcus oeni on wine hydroxycinnamic acids and volatile phenol production by Brettanomyces bruxellensis. Am. J. Enol. Vitic. 2015, 66, 357–362. [Google Scholar] [CrossRef]
- Madsen, M.G.; Edwards, N.K.; Petersen, M.A.; Mokwena, L.; Swiegers, J.H.; Arneborg, N. Influence of Oenococcus oeni and Brettanomyces bruxellensis on hydroxycinnamic acids and volatile phenols of aged wine. Am. J. Enol. Vitic. 2017, 68, 23–29. [Google Scholar] [CrossRef]
- Dias, L.; Pereira-da-Silva, S.; Tavares, M.; Malfeito-Ferreira, M.; Loureiro, V. Factors affecting the production of 4-ethylphenol by the yeast Dekkera bruxellensis in enological conditions. Food Microbiol. 2003, 20, 377–384. [Google Scholar] [CrossRef]
- Chandra, M.; Madeira, I.; Coutinho, A.; Albergaria, M.; Malfeito-Ferreira, M. Growth and volatile phenol production by Brettanomyces bruxellensis in different grapevine varieties during fermentation and in finished wine. Eur. Food Res. Technol. 2015, 242, 487–494. [Google Scholar] [CrossRef]
- Barata, A.; Caldeira, J.; Botellheiro, R.; Pagliara, D.; Malfeito-Ferreira, M.; Loureiro, V. Survival patterns of Dekkera bruxellensis in wines and inhibitory effect of sulphur dioxide. Int. J. Food Microbiol. 2008, 121, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Correia, P.; Nobre, A.; Malfeito-Ferreira, M.; Loureiro, V. Growth and 4-ethylphenol production by the yeast Pichia guilliermondii in grape juices. Am. J. Enol. Vitic. 2006, 57, 133–138. [Google Scholar]
- Dias, L.; Dias, S.; Sancho, T.; Stender, H.; Querol, A.; Malfeito-Ferreira, M.; Loureiro, V. Identification of yeasts isolated from wine related environments and capable of producing 4-ethylphenol. Food Microbiol. 2003, 20, 567–574. [Google Scholar] [CrossRef]
- Martorell, P.; Barata, A.; Malfeito-Ferreira, M.; Fernández-Espinar, M.; Loureiro, V.; Querol, A. Molecular typing of the yeast species Dekkera bruxellensis and Pichia guilliermondii recovered from wine related sources. Int. J. Food Microbiol. 2006, 106, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Ison, R.; Gutteridge, C. Determination of the carbonation tolerance of yeasts. Lett. Appl. Microbiol. 1987, 5, 11–13. [Google Scholar] [CrossRef]
- Rodrigues, N.; Gonçalves, G.; Pereira-da-Silva, S.; Malfeito-Ferreira, M.; Loureiro, V. Development and use of a new medium to detect yeasts of the genera Dekkera/Brettanomyces spp. J. Appl. Microbiol. 2001, 90, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Renouf, V.; Lonvaud-Funel, A. Development of an enrichment medium to detect Dekkera/Brettanomyces bruxellensis a spoilage yeast, on the surface of grape berries. Microbiol. Res. 2007, 162, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Guerzoni, M.E.; Marchetti, R. Analysis of yeast flora associated with grape sour rot and of the chemical disease markers. Appl. Environ. Microbiol. 1987, 53, 571–576. [Google Scholar] [PubMed]
- Connell, L.; Stender, H.; Edwards, C.G. Rapid detection and identification of Brettanomyces from winery air samples based on peptide nucleic acid analysis. Am. J. Enol. Vitic. 2002, 53, 322–324. [Google Scholar]
- Renouf, V.; Perello, M.-C.; De Revel, G.; Lonvaud-Funel, A. Survival of wine microorganisms in the bottle during storage. Am. J. Enol. Vitic. 2007, 58, 379–386. [Google Scholar]
- Malfeito-Ferreira, M.; Rodrigues, N.; Loureiro, V. The influence of oxygen on the “horse sweat taint” in red wines. Ital. Food Beverage Technol. 2001, 24, 34–38. [Google Scholar]
- Malfeito-Ferreira, M. Yeasts and wine off-flavours: A technological perspective. Ann. Microbiol. 2011, 61, 95–102. [Google Scholar] [CrossRef]
- Capozzi, V.; Di Toro, M.R.; Grieco, F.; Michelotti, V.; Salma, M.; Lamontanara, A.; Russo, P.; Orrù, L.; Alexandre, H.; Spano, G. Viable But Not Culturable (VBNC) state of Brettanomyces bruxellensis in wine: New insights on molecular basis of VBNC behaviour using a transcriptomic approach. Food Microbiol. 2016, 59, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Bizzaro, D.; Comitini, F. Evaluation of damage induced by Kwkt and Pikt zymocins against Brettanomyces/Dekkera spoilage yeast, as compared to sulphur dioxide. J. Appl. Microbiol. 2016, 121, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. Evaluation of the inhibitory effect of dimethyl dicarbonate (DMDC) against wine microorganisms. Food Microbiol. 2008, 25, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Petrova, B.; Cartwright, Z.M.; Edwards, C.G. Effectiveness of chitosan preparations against Brettanomyces bruxellensis grown in culture media and red wines. J. Int. Sci. Vigne Vin 2016, 50, 49–56. [Google Scholar] [CrossRef]
- Fabrizio, V.; Vigentini, I.; Parisi, N.; Picozzi, C.; Compagno, C.; Foschino, R. Heat inactivation of wine spoilage yeast Dekkera bruxellensis by hot water treatment. Lett. Appl. Microbiol. 2015, 61, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Delsart, C.; Grimi, N.; Boussetta, N.; Miot-Sertier, C.; Ghidossi, R.; Vorobiev, E.; Mietton-Peuchot, M. Impact of pulsed-electric field and high-voltage electrical discharges on red wine microbial stabilization and quality characteristics. J. Appl. Microbiol. 2016, 120, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, S.; Silva, F.V. High pressure processing inactivation of Brettanomyces bruxellensis in seven different table wines. Food Control 2017, 81, 1–8. [Google Scholar] [CrossRef]
- Röder, C.; König, H.; Fröhlich, J. Species-specific identification of Dekkera/Brettanomyces yeasts by fluorescently labeled DNA probes targeting the 26S rRNA. FEMS Yeast Res. 2007, 7, 1013–1026. [Google Scholar]
- Agnolucci, M.; Scarano, S.; Rea, F.; Toffanin, A.; Nuti, M. Detection of Dekkera/Brettanomyces bruxellensis in pressed Sangiovese grapes by real time PCR. Ital. J. Food Sci. 2007, 19, 153–164. [Google Scholar]
- Longin, C.; Julliat, F.; Serpaggi, V.; Maupeu, J.; Bourbon, G.; Rousseaux, S.; Guilloux-Benatier, M.; Alexandre, H. Evaluation of three Brettanomyces qPCR commercial kits: Results from an interlaboratory study. J. Int. Sci. Vigne Vin 2016, 50, 223–230. [Google Scholar] [CrossRef]
- Vendrame, M.; Manzano, M.; Comi, G.; Bertrand, J.; Iacumin, L. Use of propidium monoazide for the enumeration of viable Brettanomyces bruxellensis in wine and beer by quantitative PCR. Food Microbiol. 2014, 42, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Lourenço, A.; Barata, A.; Brito, L.; Malfeito-Ferreira, M.; Loureiro, V. Susceptibility of wine spoilage yeasts and bacteria in the planktonic state and in biofilms to disinfectants. Ann. Microbiol. 2010, 60, 549–556. [Google Scholar] [CrossRef]
- Barata, A.; Laureano, P.; D’Antuono, I.; Martorell, P.; Stender, H.; Malfeito-Ferreira, M.; Querol, A.; Loureiro, V. Enumeration and identification of 4-ethylphenol producing yeasts recovered from the wood of wine ageing barriques after different sanitation treatments. J. Food Res. 2013, 2, 140–149. [Google Scholar] [CrossRef]
- Umiker, N.L.; Descenzo, R.A.; Lee, J.; Edwards, C.G. Removal of Brettanomyces bruxellensis from red wine using membrane filtration. J. Food Process. Preserv. 2013, 37, 799–805. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Chandra, M.; Barata, A.; Ferreira-Dias, S.; Malfeito-Ferreira, M.; Loureiro, V. A response surface methodology study on the role of factors affecting growth and volatile phenol production by Brettanomyces bruxellensis ISA 2211 in wine. Food Microbiol. 2014, 42, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Pagliara, D.; Piccininno, T.; Tarantino, F.; Ciardulli, W.; Malfeito-Ferreira, M.; Loureiro, V. The effect of sugar concentration and temperature on growth and volatile phenol production by Dekkera bruxellensis in wine. FEMS Yeast Res. 2008, 8, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Milheiro, J.; Filipe-Ribeiro, L.; Cosme, F.; Nunes, F. A simple, cheap and reliable method for control of 4-ethylphenol and4-ethylguaiacol in red wines. Screening of fining agents for reducing volatile phenols levels in red wines. J. Chromatogr. B 2017, 1041–1042, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Larcher, R.; Puecher, C.; Rohregger, S.; Malacarne, M.; Nicolini, G. 4-Ethylphenol and 4-ethylguaiacol depletion in wine using esterified cellulose. Food Chem. 2012, 132, 2126–2130. [Google Scholar] [CrossRef]
- Joseph, L.; Albino, E.; Bisson, L. Creation and Use of a Brettanomyces Aroma Wheel. Catalyst 2017, 1, 12–30. [Google Scholar]
- Smith, C. Postmodern Winemaking, Rethinking the Modern Science of an Ancient Craft; University California Press: Berkeley, CA, USA, 2013; ISBN 9780520282599. [Google Scholar]
Figure 1.
Number of references related with “Brettanomyces” (black bars) and with “Brettanomyces or Dekkera and wine” (white bars) retrieved from Scopus search engine (www.scopus.com).
Figure 1.
Number of references related with “Brettanomyces” (black bars) and with “Brettanomyces or Dekkera and wine” (white bars) retrieved from Scopus search engine (www.scopus.com).
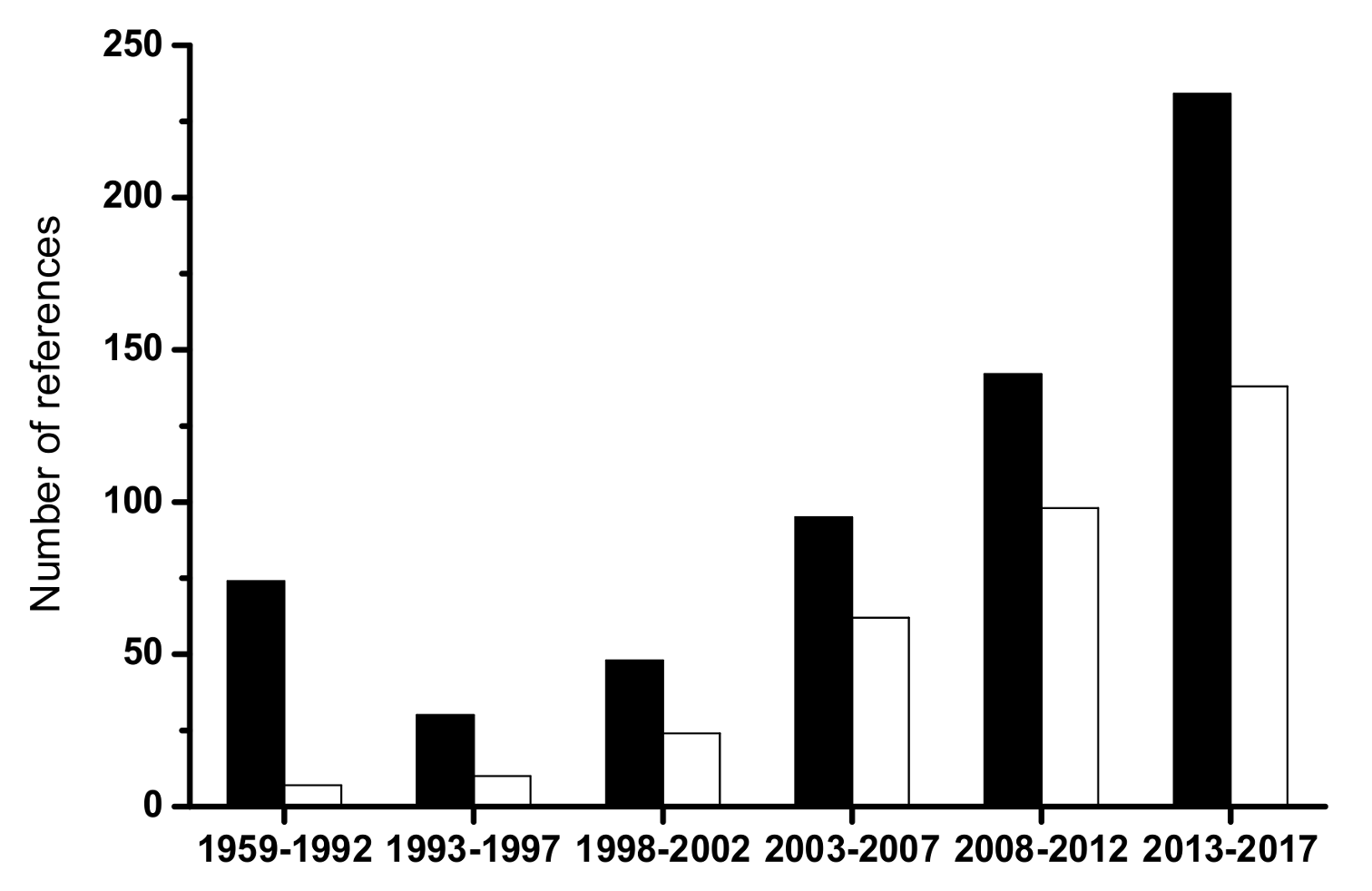
Figure 2.
Effect of chitosan on the viability of growing cells D. bruxellensis (•) and on 4-ethylphenol (∆) production. Arrow: moment of chitosan addition. Absence of viable cells (CFU <1/mL) is indicated as 1 CFU/mL due to the logarithmic scale of the y-axis.
Figure 2.
Effect of chitosan on the viability of growing cells D. bruxellensis (•) and on 4-ethylphenol (∆) production. Arrow: moment of chitosan addition. Absence of viable cells (CFU <1/mL) is indicated as 1 CFU/mL due to the logarithmic scale of the y-axis.
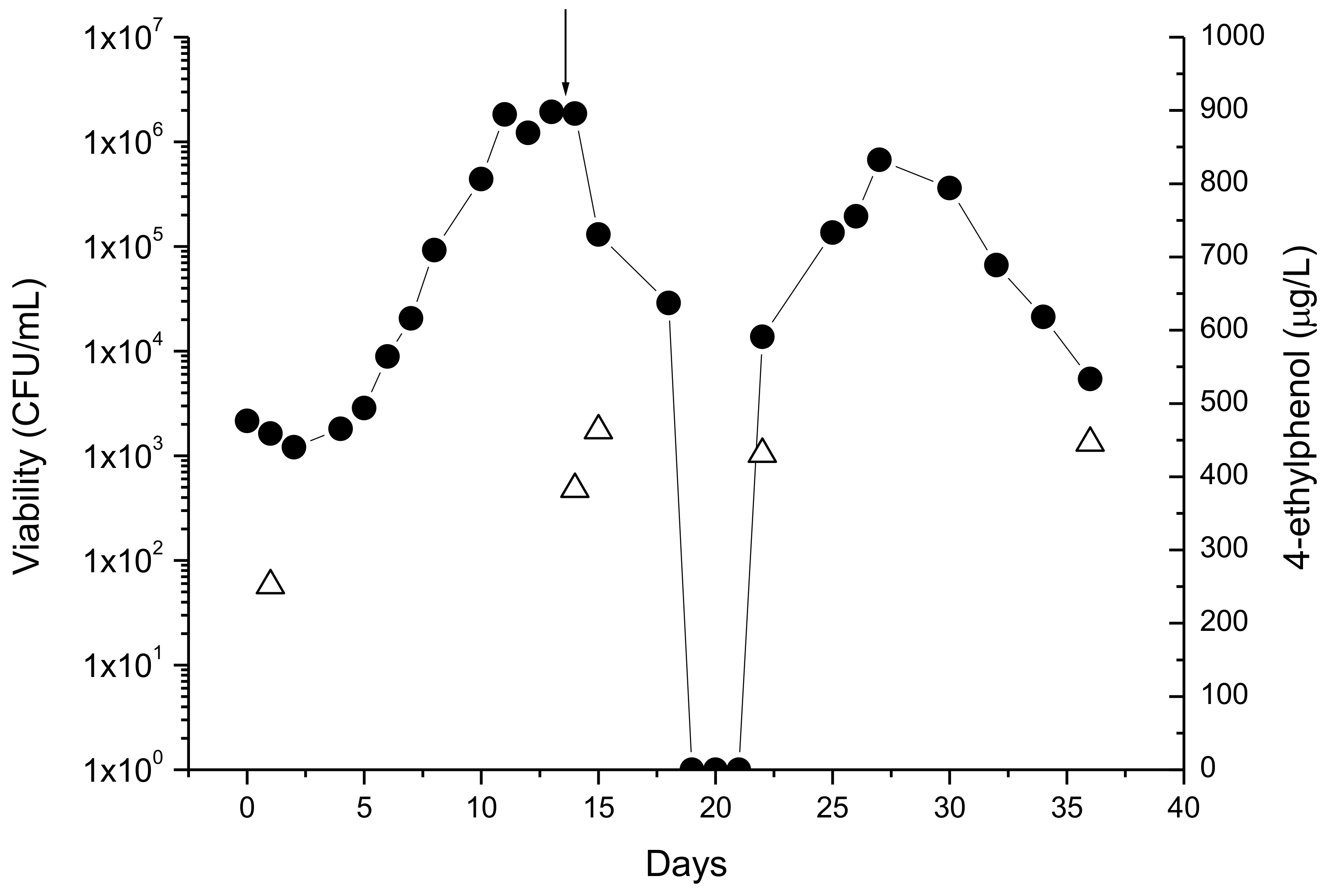
Figure 3.
Growth (A) and 4-EP production (B) by B. bruxellensis under wines stored at different temperatures (symbols: ■, 3 °C; ○, 10 °C; ∆, 15 °C; ▼, 20 °C). Cells were inoculated in 1 L Schott flasks and incubated without agitation to mimic tank storage conditions.
Figure 3.
Growth (A) and 4-EP production (B) by B. bruxellensis under wines stored at different temperatures (symbols: ■, 3 °C; ○, 10 °C; ∆, 15 °C; ▼, 20 °C). Cells were inoculated in 1 L Schott flasks and incubated without agitation to mimic tank storage conditions.
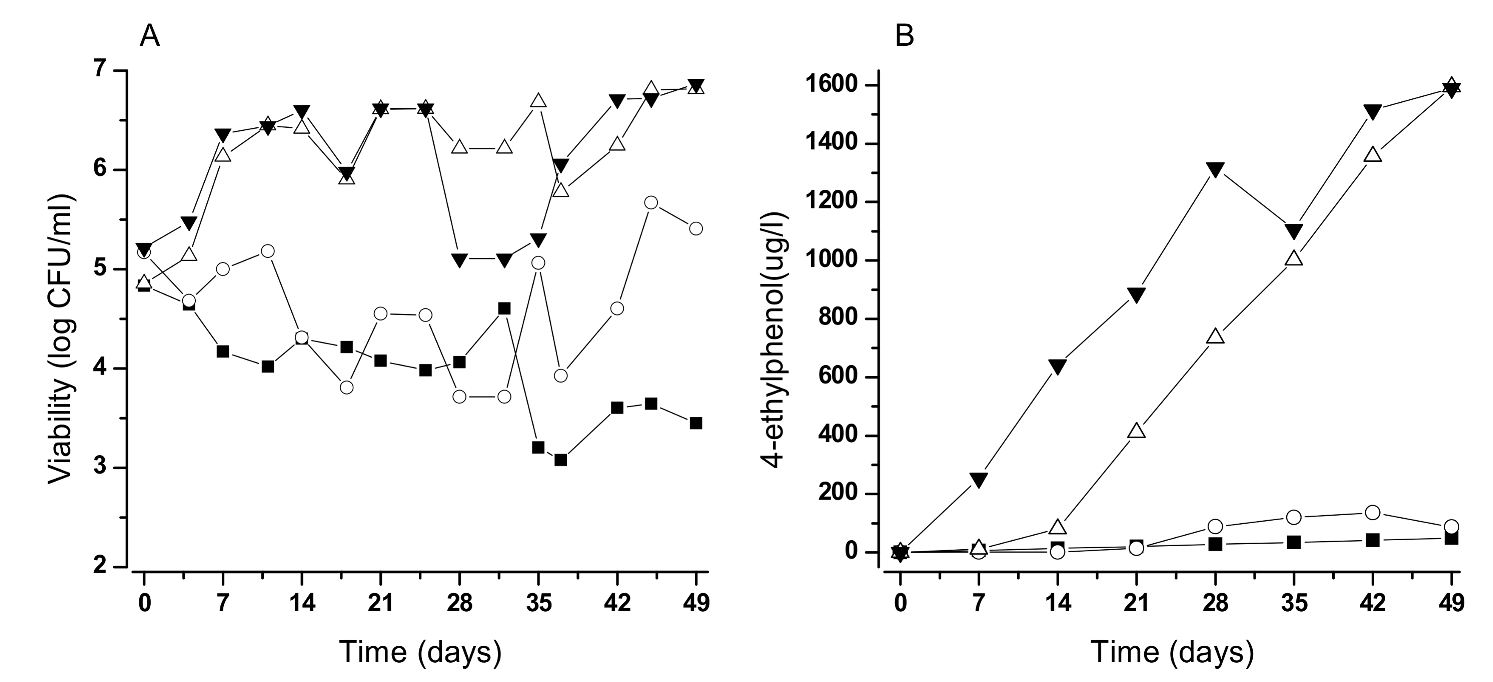
Figure 4.
Effect of activated charcoal on the reduction of 4-EP (µg/L) in two red different wines (A,B) after 3 (●), 10 (■) and 21 days (∆) of contact. Insertions: effect on color intensity (■) and hue (×10, ●) after 10 days of contact.
Figure 4.
Effect of activated charcoal on the reduction of 4-EP (µg/L) in two red different wines (A,B) after 3 (●), 10 (■) and 21 days (∆) of contact. Insertions: effect on color intensity (■) and hue (×10, ●) after 10 days of contact.
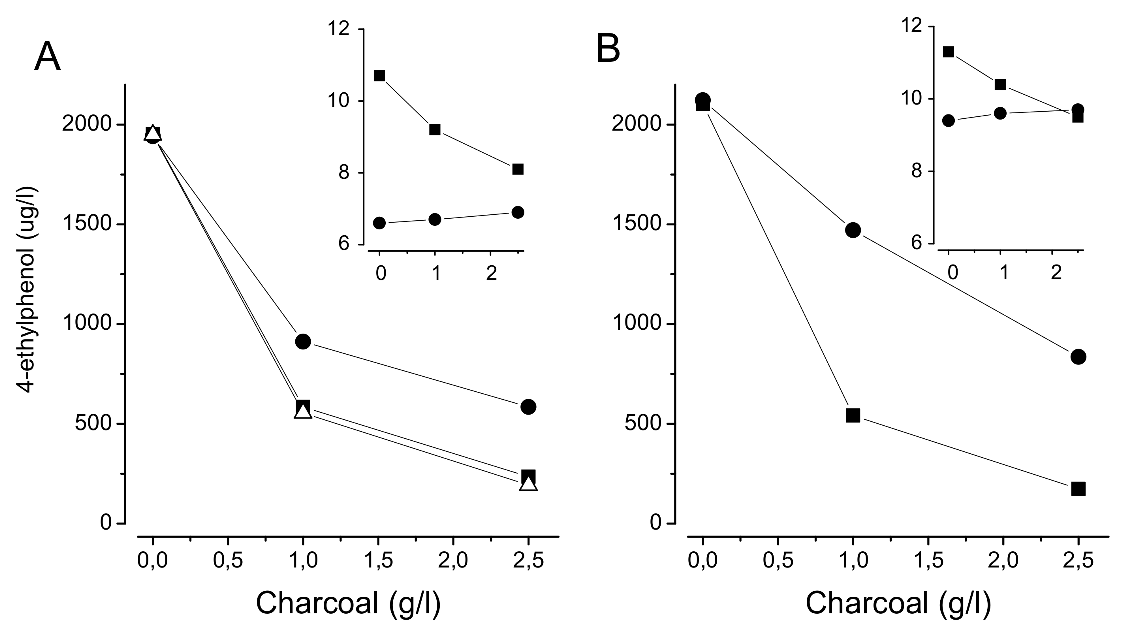
Figure 5.
Relation between the real chemical concentration of 4-ethylphenol and the respective predicted concentration given by two different tasting panels ((A) DOC certification office; (B) faculty enology students). Wines from 1 to 5 are commercial brands. Wine 6 is a blank sample spiked with of 4-ethylphenol (2000 µg/L) and 4-ethylguaiacol (250 µg/L). Vertical bars indicate standard deviation.
Figure 5.
Relation between the real chemical concentration of 4-ethylphenol and the respective predicted concentration given by two different tasting panels ((A) DOC certification office; (B) faculty enology students). Wines from 1 to 5 are commercial brands. Wine 6 is a blank sample spiked with of 4-ethylphenol (2000 µg/L) and 4-ethylguaiacol (250 µg/L). Vertical bars indicate standard deviation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Minimum lethal concentration of dimethyldicarbonate (DMDC) (mg/L) against several wine related microbial species as a function of initial cellular inoculum (adapted from [39]).
Table 1.
Minimum lethal concentration of dimethyldicarbonate (DMDC) (mg/L) against several wine related microbial species as a function of initial cellular inoculum (adapted from [39]).
| Species | 500 cells/mL | >104 cells/mL | Species | 500 cells/mL | >104 cells/mL |
|---|---|---|---|---|---|
| D. bruxellensis ISA 1791 | 100 | 300 | S. pombe ISA 1190 | 100 | >300 |
| P. guilliermondii ISA 2105 | 100 | 300 | Z. bailii ISA 1307 | 25 | 200 |
| S. cerevisiae ISA 1000 | 100 | 200 | Lactic acid bacteria | >300 | >300 |
| S. cerevisiae ISA 1026 | 100 | 200 | Acetic acid bacteria | >300 | >300 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Malfeito-Ferreira, M. Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where do We Stand Now? Beverages 2018, 4, 32. https://doi.org/10.3390/beverages4020032
AMA Style
Malfeito-Ferreira M. Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where do We Stand Now? Beverages. 2018; 4(2):32. https://doi.org/10.3390/beverages4020032
Chicago/Turabian StyleMalfeito-Ferreira, Manuel. 2018. "Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where do We Stand Now?" Beverages 4, no. 2: 32. https://doi.org/10.3390/beverages4020032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.