
Stephanoascus ciferrii Complex: The Current State of Infections and Drug Resistance in Humans

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Collection
3. Results
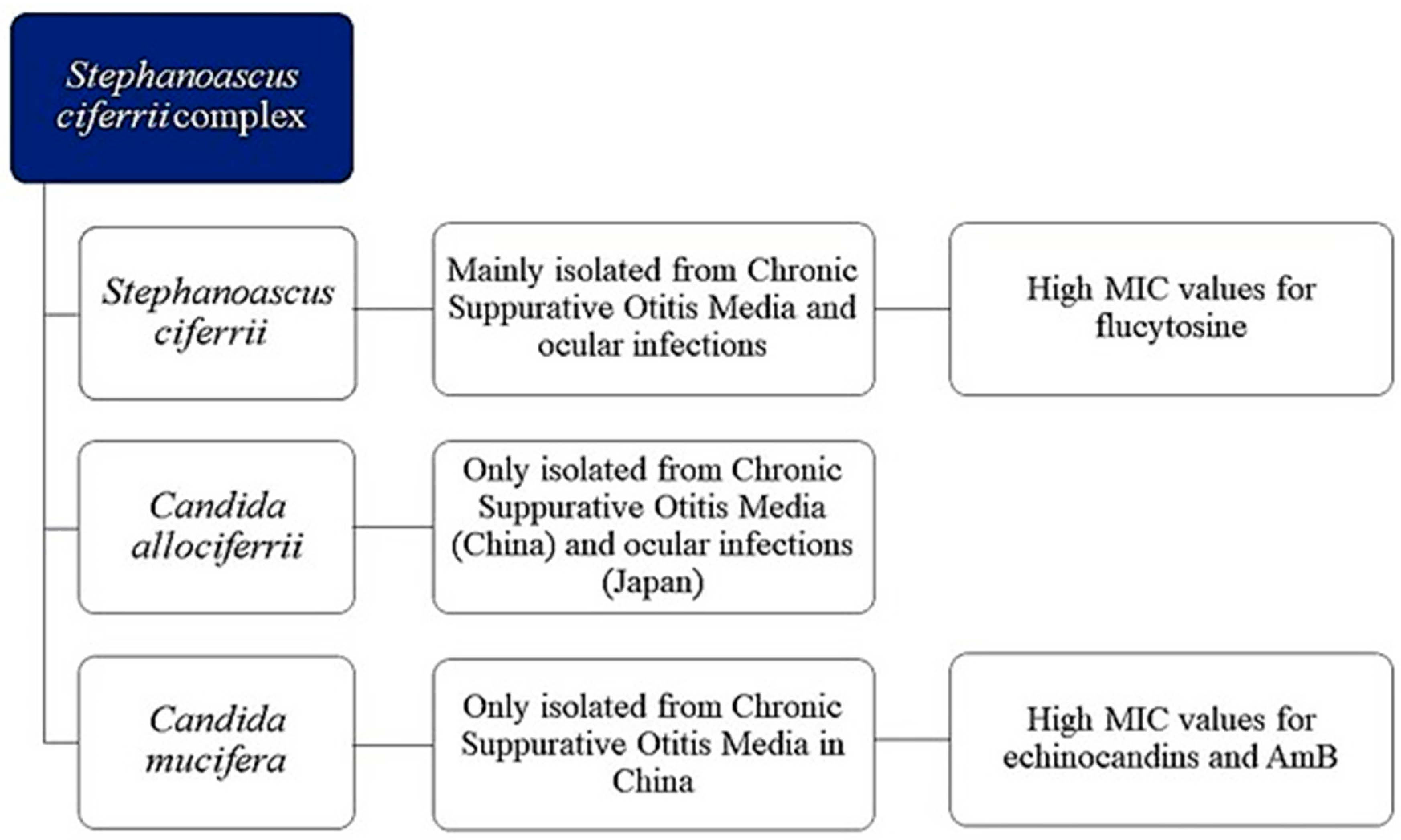
3.1. S. ciferrii Complex: A Real Fungal Pathogen
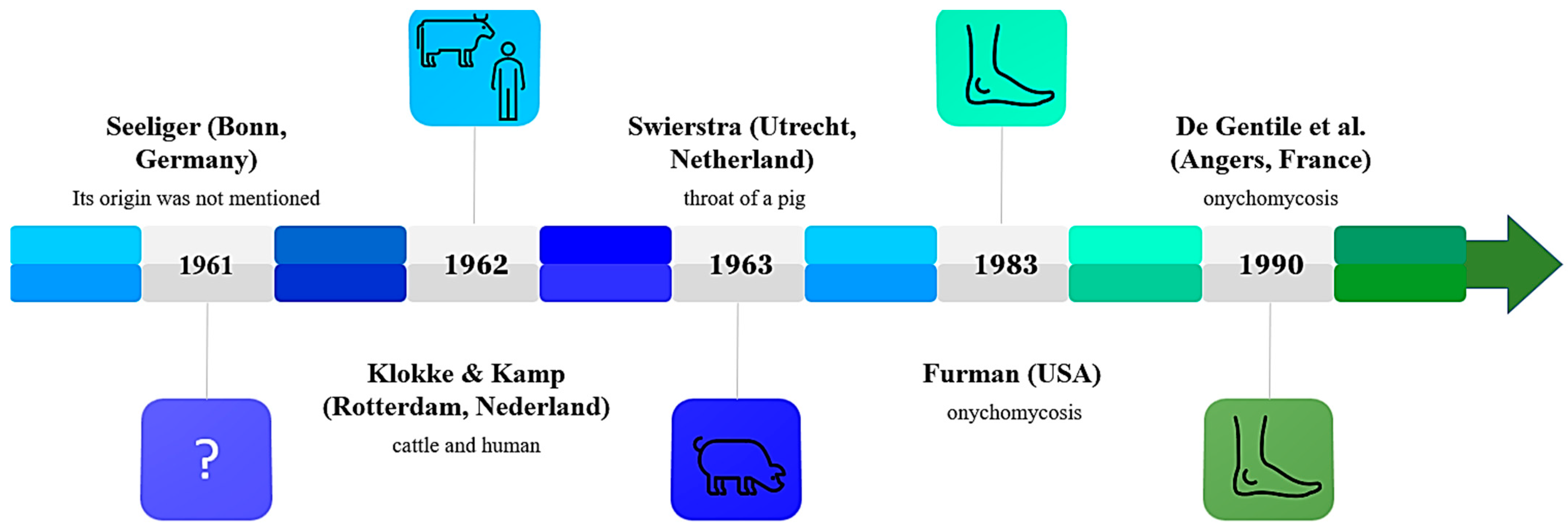
3.2. History
3.3. Morphology and Identification of S. ciferrii Complex
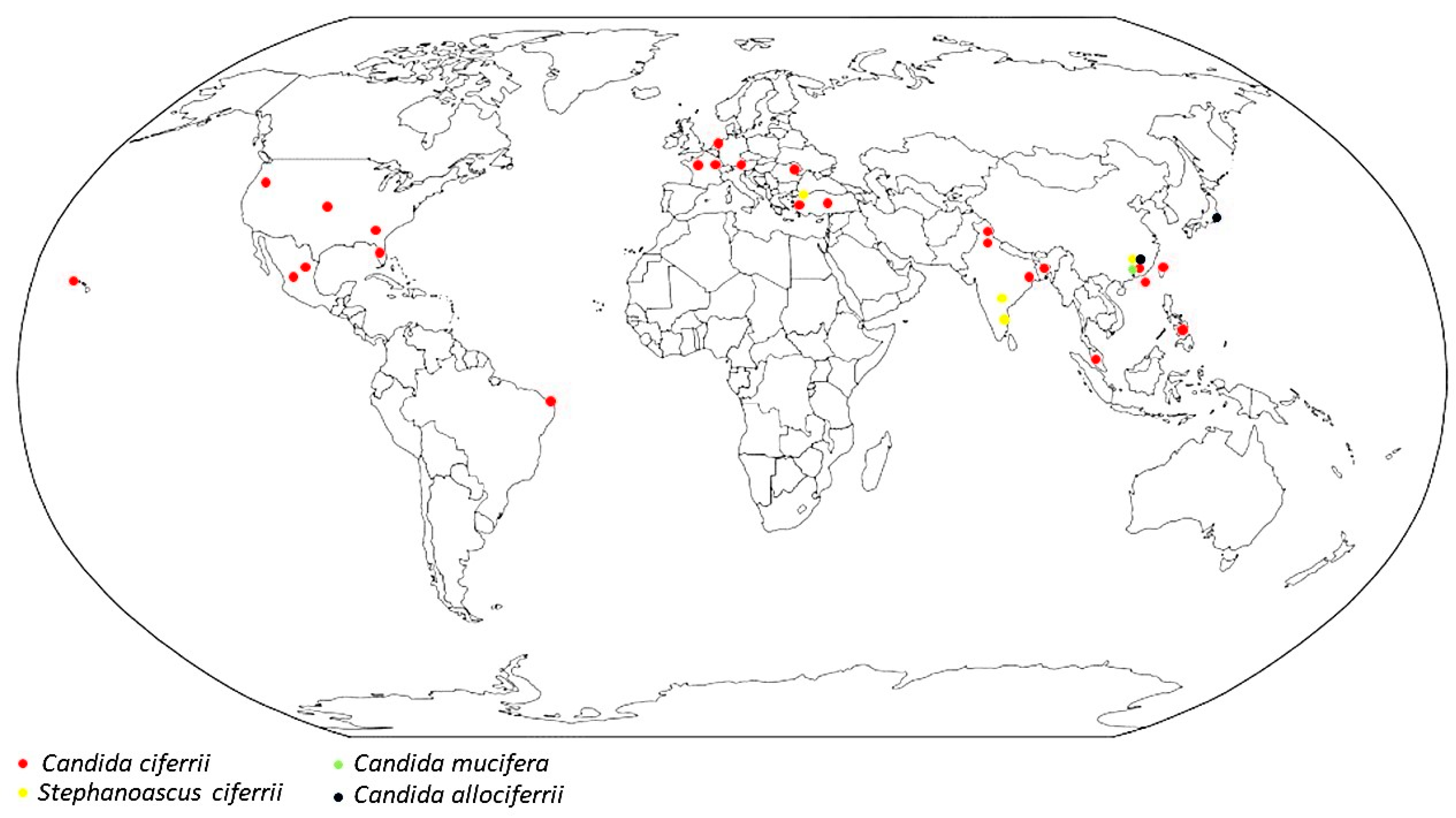
3.4. Epidemiology
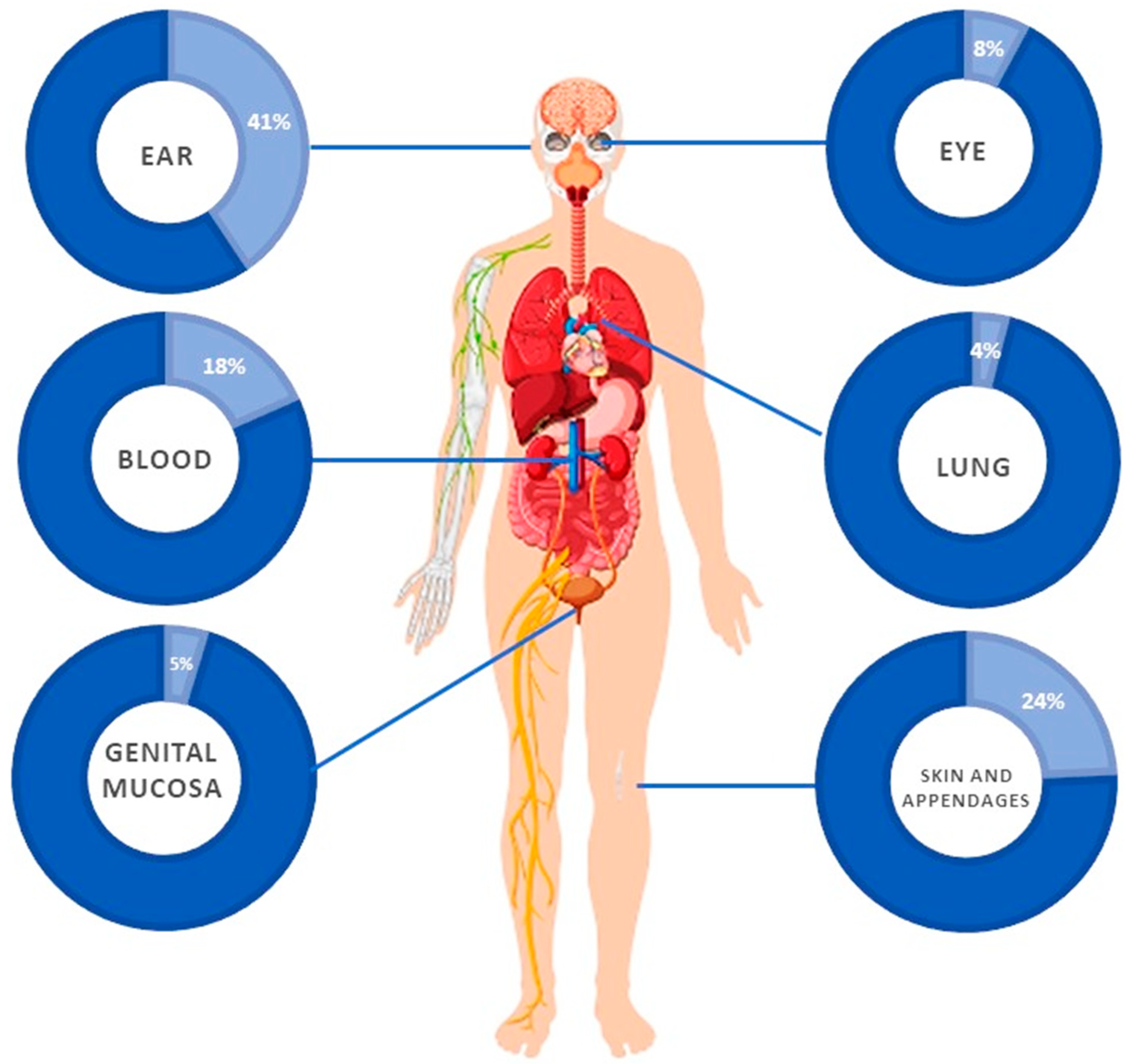
3.5. Clinical Features
3.5.1. Auricular Infections
3.5.2. Infections of the Skin and Its Appendages
3.5.3. Ocular Infections
3.5.4. Candidemia
3.5.5. Vaginal Infections
3.5.6. Pulmonary Infections
3.6. Treatment and Resistance
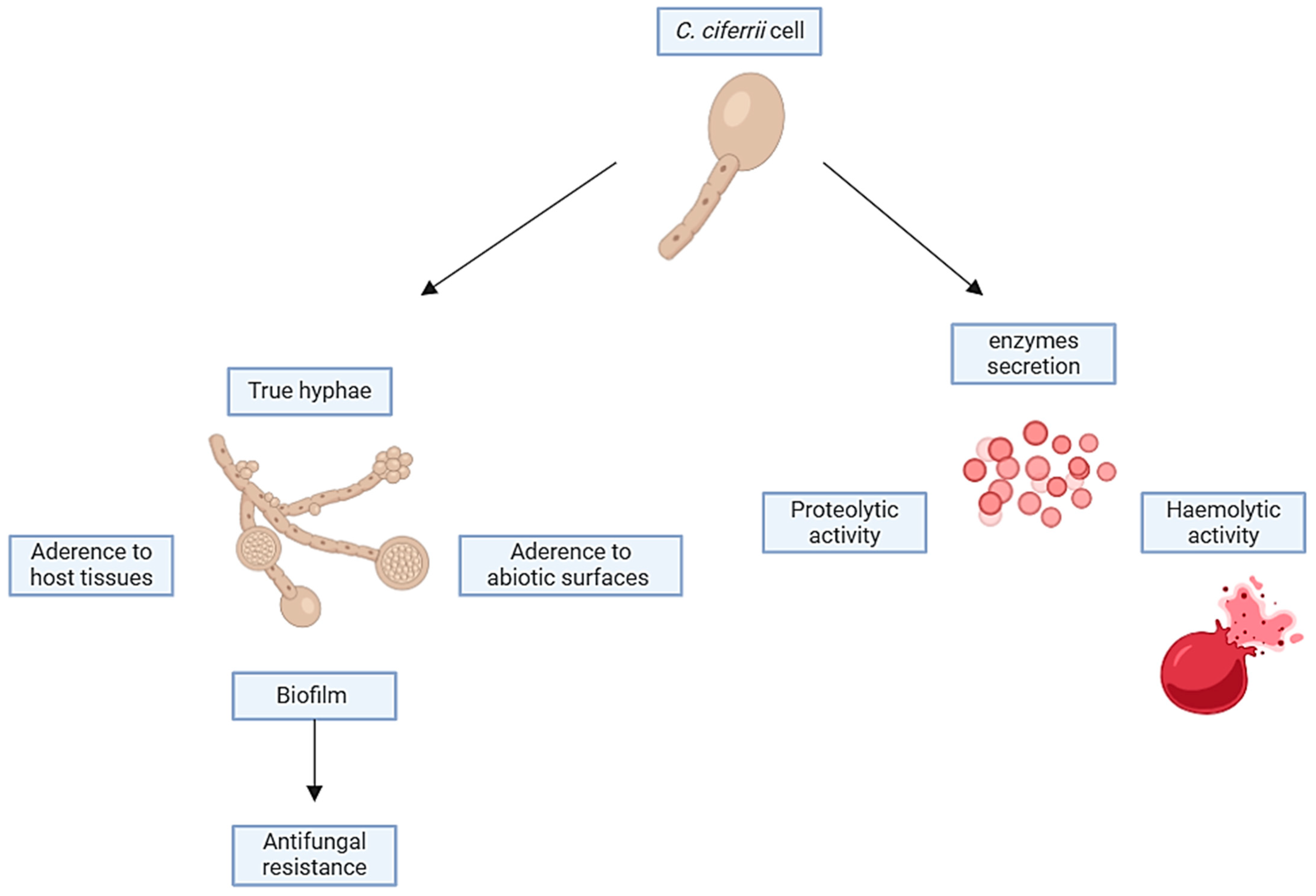
3.7. Virulence Factors
3.8. S. ciferrii Complex as Colonizer in Humans
3.9. S. ciferrii Complex in the One Health Perspective
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dellière, S.; Guery, R.; Candon, S.; Rammaert, B.; Aguilar, C.; Lanternier, F.; Chatenoud, L.; Lortholary, O. Understanding Pathogenesis and Care Challenges of Immune Reconstitution Inflammatory Syndrome in Fungal Infections. J. Fungi 2018, 4, 139. [Google Scholar] [CrossRef]
- Campione, E.; Cosio, T.; Lanna, C.; Mazzilli, S.; Ventura, A.; Dika, E.; Gaziano, R.; Dattola, A.; Candi, E.; Bianchi, L. Predictive role of vitamin A serum concentration in psoriatic patients treated with IL-17 inhibitors to prevent skin and systemic fungal infections. J. Pharmacol. Sci. 2020, 144, 52–56. [Google Scholar] [CrossRef]
- Kidd, S.E.; Chen, S.C.; Meyer, W.; Halliday, C.L. A New Age in Molecular Diagnostics for Invasive Fungal Disease: Are We Ready? Front. Microbiol. 2020, 10, 2903. [Google Scholar] [CrossRef]
- Banerjee, S.; Denning, D.W.; Chakrabarti, A. One Health aspects & priority roadmap for fungal diseases: A mini-review. Indian J. Med. Res. 2021, 153, 311–319. [Google Scholar] [CrossRef]
- Hofer, U. Candida auris’ potential link to climate change. Nat. Rev. Microbiol. 2019, 17, 588. [Google Scholar] [CrossRef]
- Simões, D.; de Andrade, E.; Sabino, R. Fungi in a One Health Perspective. Encyclopedia 2023, 3, 900–918. [Google Scholar] [CrossRef]
- Denning, D.W. Calling upon all public health mycologists: To accompany the country’s burden papers from 14 countries. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 923–924. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.J.; Ferraro, J.P.; Rea, S.; Kaufusi, S.; Goodman, B.E.; Spalding, J. Epidemiology and Clinical Features of Invasive Fungal Infection in a US Health Care Network. Open Forum Infect. Dis. 2018, 5, ofy187. [Google Scholar] [CrossRef] [PubMed]
- Kotey, F.C.; Dayie, N.T.; Tetteh-Uarcoo, P.B.; Donkor, E.S. Candida Bloodstream Infections: Changes in Epidemiology and Increase in Drug Resistance. Infect. Dis. 2021, 14, 11786337211026927. [Google Scholar] [CrossRef]
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control 2016, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Chrenkova, V.; Vadkertiova, R.; Vlachova, K.; Babjuk, M.; Lischke, R.; Bebrova, E.; Hubacek, P. Candida sojae: First report of a human infection. J. Mycol. Med. 2022, 32, 101309. [Google Scholar] [CrossRef] [PubMed]
- de Gentile, L.; Bouchara, J.P.; Cimon, B.; Chabasse, D. Candida ciferrii: Clinical and microbiological features of an emerging pathogen. Mycoses 1991, 34, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.A.; Franco-Palacios, D.; King, C.S.; Shlobin, O.A.; Nathan, S.D.; Katugaha, S.B.; Mani, H.; Brown, A.W. A 24-Year-Old Woman with Precipitous Respiratory Failure After Lung Transplantation. Chest 2018, 153, e53–e56. [Google Scholar] [CrossRef] [PubMed]
- Kreger-Van Rij, N.J. Candida ciferrii, a new yeast species. Mycopathol. Mycol. Appl. 1965, 26, 49–52. [Google Scholar] [CrossRef]
- Furman, R.M.; Ahearn, D.G. Candida ciferrii and Candida chiropterorum isolated from clinical specimens. J. Clin. Microbiol. 1983, 18, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Soki, H.; Abo, K.; Yamazaki, K.; Kojima, T.; Oda, T.; Uzawa, Y.; Kikuchi, K. First Report of Intraorbital Abscess Caused by Candida allociferrii and Specific PCR for Differentiating Stephanoascus ciferrii Complex Species. Med. Mycol. J. 2015, 56, E9–E14. (In Japanese) [Google Scholar] [CrossRef] [PubMed]
- Soki, H.; Nagase, Y.; Yamazaki, K.; Oda, T.; Kikuchi, K. Isolation of the yeast-like fungus Stephanoascus ciferrii by culturing the aural discharge of a patient with intractable otitis media. Case report. J. Jpn. Assoc. Infect. Dis. 2010, 84, 210–212. (In Japanese) [Google Scholar] [CrossRef]
- Agın, H.; Ayhan, Y.; Devrim, I.; Gülfidan, G.; Tulumoglu, S.; Kayserili, E. Fluconazole-, amphotericin-B-, caspofungin-, and anidulafungin-resistant Candida ciferrii: An unknown cause of systemic mycosis in a child. Mycopathologia 2011, 172, 237–239. [Google Scholar] [CrossRef]
- Danielescu, C.; Cantemir, A.; Chiselita, D. Successful treatment of fungal endophthalmitis using intravitreal caspofungin. Arq. Bras. Oftalmol. 2017, 80, 196–198. [Google Scholar] [CrossRef]
- Baquero, F.; Moya, A. Intelligibility in microbial complex systems: Wittgenstein and the score of life. Front. Cell. Infect. Microbiol. 2012, 2, 88. [Google Scholar] [CrossRef] [PubMed]
- Mixão, V.; Saus, E.; Hansen, A.P.; Lass-Florl, C.; Gabaldón, T. Genome Assemblies of Two Rare Opportunistic Yeast Pathogens: Diutina rugosa (syn. Candida rugosa) and Trichomonascus ciferrii (syn. Candida ciferrii). G3 2019, 9, 3921–3927. [Google Scholar] [CrossRef] [PubMed]
- Ueda-Nishimura, K.; Kozaburo, M. Species distinction of the ascomycetous heterothallic yeast-like fungus Stephanoascus ciferrii complex: Description of Candida allociferrii sp. nov. and reinstatement of Candida mucifera Kocková-Kratochvílová et Sláviková. Int. J. Syst. Evol. Microbiol. 2002, 52, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Middelhoven, W.J.; De Kievit, H.; Biesbroek, A.L. Yeast species utilizing uric acid, adenine, n-alkylamines or diamines as sole source of carbon and energy. Antonie Van Leeuwenhoek 1985, 51, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wu, Z.; Liu, P.; Chen, Y.; Liao, K.; Peng, Y.; He, Y. Identification and Antifungal Susceptibility Analysis of Stephanoascus ciferrii Complex Species Isolated from Patients with Chronic Suppurative Otitis Media. Front. Microbiol. 2021, 12, 680060. [Google Scholar] [CrossRef] [PubMed]
- De Gentile, L.; Bouchara, J.P.; Le Clec’h, C.; Cimon, B.; Symoens, F.; Chabasse, D. Prevalence of Candida ciferrii in elderly patients with trophic disorders of the legs. Mycopathologia 1995, 131, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Gunsilius, E.; Lass-Flörl, C.; Kähler, C.M.; Gastl, G.; Petzer, A.L. Candida ciferrii, a new fluconazole-resistant yeast causing systemic mycosis in immunocompromised patients. Ann. Hematol. 2001, 80, 178–179. [Google Scholar] [CrossRef]
- Cheng, M.F.; Yu, K.W.; Tang, R.B.; Fan, Y.H.; Yang, Y.L.; Hsieh, K.S.; Ho, M.; Lo, H.J. Distribution and antifungal susceptibility of Candida species causing candidemia from 1996 to 1999. Diagn. Microbiol. Infect Dis. 2004, 48, 33–37. [Google Scholar] [CrossRef]
- Saha, K.; Sit, N.K.; Maji, A.; Jash, D. Recovery of fluconazole sensitive Candida ciferrii in a diabetic chronic obstructive pulmonary disease patient presenting with pneumonia. Lung India 2013, 30, 338–340. [Google Scholar] [CrossRef]
- Villanueva-Lozano, H.; Treviño-Rangel, R.J.; Hernández-Balboa, C.L.; González, G.M.; Martínez-Reséndez, M.F. An unusual case of Candida ciferrii fungemia in an immunocompromised patient with Crohn’s and Mycobacterium bovis disease. J. Infect. Dev. Ctries 2016, 10, 1156–1158. [Google Scholar] [CrossRef]
- Dave, V.P.; Sharma, S.; Dave, P.J.; Joseph, J.; Pappuru, R.R. Clinical presentations, diagnostic dilemma, and management outcomes of chronic postoperative endophthalmitis caused by Stephanoascus ciferrii. Retin. Cases Brief Rep. 2021, 15, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Thakar, M.; Kumar, R.; Loomba, P. Postpartum Endogenous Endophthalmitis in a Young Immunocompetent Female by a Rare Fungus Candida ciferrii. Ocul. Immunol. Inflamm. 2022, 30, 2043–2046. [Google Scholar] [CrossRef] [PubMed]
- Robles-Tenorio, A.; Serrano-Ríos, F.E.; Tarango-Martínez, V.M. Onychomycosis by Candida ciferrii caused fatal multisystemic dissemination in a patient with diabetes mellitus type 2. J. Eur. Acad. Dermatol. Venereol. 2022, 36, e77–e79. [Google Scholar] [CrossRef] [PubMed]
- Sathi, F.A.; Paul, S.K.; Ahmed, S.; Alam, M.M.; Nasreen, S.A.; Haque, N.; Islam, A.; Nila, S.S.; Afrin, S.Z.; Aung, M.S.; et al. Prevalence and Antifungal Susceptibility of Clinically Relevant Candida Species, Identification of Candida auris and Kodamaea ohmeri in Bangladesh. Trop. Med. Infect. Dis. 2022, 7, 211. [Google Scholar] [CrossRef]
- Ng, K.P.; Kuan, C.S.; Kaur, H.; Na, S.L.; Atiya, N.; Velayuthan, R.D. Candida species epidemiology 2000–2013: A laboratory-based report. Trop. Med. Int. Health 2015, 20, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Romald, P.N.; Sridharan, K.S.; Mohanty, S.; Kindo, A.J. Rare Isolate of Stephanoascus ciferrii from the Aural Discharge of Post-mastoidectomy Patient—A Case Report. J. Clin. Diagn. Res. 2019, 13, DD01–DD03. [Google Scholar] [CrossRef]
- Demiray, T.; Hafizoglu, T.; Koroglu, M.; Ozbek, A.; Altindis, M. The First Case of Stephanoascus ciferrii Infection in a Newborn and Review of Literature. Nobel Med. 2015, 11, 100–197. [Google Scholar]
- Papìla, Ç.; Özguler, M.; Albayrak, S.; Devecì, Y.; Alataso, O. A Case of Septic Pulmonary Embolism due to Candida ciferrii. F.Ü. Sağ. Bil. Tıp Derg. 2016, 30, 25–29. Available online: http://www.fusabil.org (accessed on 23 December 2023).
- Chan, J.F.W.; Tsan, C.C.; Yang, J.; Xing, F.; Leung, S.M.; Ngan, A.H.Y.; Lau, S.K.P.; Woo, P.C.Y.; Yuen, K.Y. Clinical characteristics, laboratory identification, and in vitro antifungal susceptibility of Trichomonascus (Candida) ciferrii isolates associated with granular myringitis. In Proceedings of the The 26th European Congress of Clinical Microbiology and Infectious Diseases (ECCMID 2016), Amsterdam, The Netherlands, 9–12 April 2016. [Google Scholar]
- Vieira de Melo, A.P.; Zuza-Alves, D.L.; da Silva-Rocha, W.P.; Ferreira Canário de Souza, L.B.; Francisco, E.C.; Salles de Azevedo Melo, A.; Maranhão Chaves, G. Virulence factors of Candida spp. obtained from blood cultures of patients with candidemia attended at tertiary hospitals in Northeast Brazil. J. Mycol. Med. 2019, 29, 132–139. [Google Scholar] [CrossRef]
- Capoor, M.R.; Gupta, D.K.; Verma, P.K.; Sachdeva, H.C. Rare yeasts causing fungemia in immunocompromised and haematology patients: Case series from Delhi. Indian. J. Med. Microbiol. 2015, 33, 576–579. [Google Scholar] [CrossRef]
- Kano, R.; Makimura, K.; Kushida, T.; Nomura, M.; Yamaguchi, H.; Hasegawa, A. First isolation of Stephanoascus ciferrii from a cat. Microbiol. Immunol. 2000, 44, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.N.; Ortiz, S.O.; Mello, M.M.; de Mattos Oliveira, F.O.; Severo, L.C.; Goebel, C.S. Comparison between four usual methods of identification of Candida species. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 281–287. [Google Scholar] [CrossRef]
- Kumar, A.; Babu, R.; Bijulal, S.; Abraham, M.; Sasidharan, P.; Kathuria, S.; Sharma, C.; Meis, J.F.; Chowdhary, A. Invasive mycosis due to species of Blastobotrys in immunocompromised patients with reduced susceptibility to antifungals. J. Clin. Microbiol. 2014, 52, 4094–4099. [Google Scholar] [CrossRef] [PubMed]
- Al-Haqqan, A.; Al-Sweih, N.; Ahmad, S.; Khan, S.; Joseph, L.; Varghese, S.; Khan, Z. Azole-resistant Candida blankii as a newly recognized cause of bloodstream infection. New Microbes New Infect. 2018, 26, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Y.; Chiang-Ni, C.; Teng, S.H. Current status of MALDI-TOF mass spectrometry in clinical microbiology. J. Food Drug Anal. 2019, 27, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Marklein, G.; Josten, M.; Klanke, U.; Müller, E.; Horré, R.; Maier, T.; Wenzel, T.; Kostrzewa, M.; Bierbaum, G.; Hoerauf, A.; et al. Matrix-assisted laser desorption ionization-time of flight mass spectrometry for fast and reliable identification of clinical yeast isolates. J. Clin. Microbiol. 2009, 47, 2912–2917. [Google Scholar] [CrossRef] [PubMed]
- Roehmel, J.F.; Tintelnot, K.; Bernhardt, A.; Seibold, M.; Staab, D.; Schwarz, C. Arxula adeninivorans causing invasive pulmonary mycosis and fungaemia in cystic fibrosis. Lancet 2015, 385, 1476. [Google Scholar] [CrossRef] [PubMed]
- Balada-Llasat, J.M.; LaRue, H.; Kamboj, K.; Rigali, L.; Smith, D.; Thomas, K.; Pancholi, P. Detection of yeasts in blood cultures by the Luminex xTAG fungal assay. J. Clin. Microbiol. 2012, 50, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Ropars, J.; Maufrais, C.; Diogo, D.; Marcet-Houben, M.; Perin, A.; Sertour, N.; Mosca, K.; Permal, M.; Laval, G.; Bouchier, C.; et al. Gene flow contributes to diversification of the major fungal pathogen Candida albicans. Nat. Commun. 2018, 9, 2253. [Google Scholar] [CrossRef]
- Schröder, M.S.; Martinez de San Vicente, K.; Prandini, T.H.R.; Hammel, S.; Higgins, D.G.; Bagagli, E.; Wolfe, K.H.; Butler, G. Multiple Origins of the Pathogenic Yeast Candida orthopsilosis by Separate Hybridizations between Two Parental Species. PLoS Genet. 2016, 12, e1006404. [Google Scholar] [CrossRef]
- Gabaldón, T. Large-scale assignment of orthology: Back to phylogenetics? Genome Biol. 2008, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Butler, G.; Kenny, C.; Fagan, A.; Kurischko, C.; Gaillardin, C.; Wolfe, K.H. Evolution of the MAT locus and its Ho endonuclease in yeast species. Proc. Natl. Acad. Sci. USA 2004, 101, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Biplob, J.A.; Sathi, F.A.; Nila, S.S.; Paul, A.; Khanam, J.; Chowdhury, C.S.; Khan, M.S. Ear Infections by Non albicans Candida Species with Isolation of Rare Drug Resistant Species in a Tertiary Care Hospital of Bangladesh. Mymensingh Med. J. 2023, 32, 644–648. [Google Scholar]
- Punia, R.S.; Singhal, S.K.; Kundu, R.; Das, A.; Chander, J. Fungal Suppurative Otitis Media (Histopathology) Among Patients in North India. Head Neck Pathol. 2019, 13, 149–153. [Google Scholar] [CrossRef]
- Kasinathan, S.; Kondamudi, N.P. Bullous Myringitis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Sharma, B.; Nonzom, S. Superficial mycoses, a matter of concern: Global and Indian scenario-an updated analysis. Mycoses 2021, 64, 890–908. [Google Scholar] [CrossRef] [PubMed]
- Rather, S.; Keen, A.; Shah, F.Y.; Yaseen, A.; Farooq, S.; Bakhshi, A. Candidal Onychomycosis: Clinico Epidemiological Profile, Prevailing Strains, and Antifungal Susceptibility Pattern—A Study from a Tertiary Care Hospital. Indian J. Dermatol. 2021, 66, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Tomic-Canic, M.; Burgess, J.L.; O’Neill, K.E.; Strbo, N.; Pastar, I. Skin Microbiota and its Interplay with Wound Healing. Am. J. Clin. Dermatol. 2020, 21, 36–43. [Google Scholar] [CrossRef]
- Petrillo, F.; Sinoca, M.; Fea, A.M.; Galdiero, M.; Maione, A.; Galdiero, E.; Guida, M.; Reibaldi, M. Candida Biofilm Eye Infection: Main Aspects and Advance in Novel Agents as Potential Source of Treatment. Antibiotics 2023, 12, 1277. [Google Scholar] [CrossRef]
- Kalkanci, A.; Ozdek, S. Ocular fungal infections. Curr. Eye Res. 2011, 36, 179–189. [Google Scholar] [CrossRef]
- Parke, D.W.; Jones, D.B.; Gentry, L.O. Endophthalmitis is present in remarkable percentages of patients with Candida albicans fungemia. Ophthalmology 1982, 89, 789–796. [Google Scholar] [CrossRef]
- Cuervo, G.; Garcia-Vidal, C.; Puig-Asensio, M.; Merino, P.; Vena, A.; Martin-Pena, A.; Montejo, J.M.; Ruiz, A.; Lazaro-Perona, F.; Fortun, J.; et al. Usefulness of guideline recommendations for prognosis in patients with candidemia. Med. Mycol. 2019, 57, 659–667. [Google Scholar] [CrossRef]
- Pappas, P.; Lionakis, M.; Arendrup, M.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Koehler, P.; Stecher, M.; Cornely, O.A.; Koehler, D.; Vehreschild, M.; Bohlius, J.; Wisplinghoff, H.; Vehreschild, J.J. Morbidity and mortality of candidemia in Europe: An epidemiologic meta-analysis. Clin. Microbiol. Infect. 2019, 25, 1200–1212. [Google Scholar] [CrossRef]
- Fournier, P.; Schwebel, C.; Maubon, D.; Vesin, A.; Lebeau, B.; Foroni, L.; Hamidfar-Roy, R.; Cornet, M.; Timsit, J.F.; Pelloux, H. Antifungal use influences Candida species distribution and susceptibility in the intensive care unit. J. Antimicrob. Chemother. 2011, 66, 2880–2886. [Google Scholar] [CrossRef]
- Shah, D.N.; Yau, R.; Lasco, T.M.; Weston, J.; Salazar, M.; Palmer, H.R.; Garey, K.W. Impact of prior inappropriate fluconazole dosing on isolation of fluconazole-nonsusceptible Candida species in hospitalized patients with candidemia. Antimicrob. Agents Chemother. 2012, 56, 3239–3243. [Google Scholar] [CrossRef]
- Schwartz, I.S.; Patterson, T.F. The Emerging Threat of Antifungal Resistance in Transplant Infectious Diseases. Curr. Infect. Dis. Rep. 2018, 20, 2. [Google Scholar] [CrossRef]
- Galia, L.; Pezzani, M.D.; Compri, M.; Callegari, A.; Rajendran, N.B.; Carrara, E.; Tacconelli, E.; The Combacte Magnet Epi-Net Network. Surveillance of Antifungal Resistance in Candidemia Fails to Inform Antifungal Stewardship in European Countries. J. Fungi 2022, 8, 249. [Google Scholar] [CrossRef]
- Gaziano, R.; Sabbatini, S.; Monari, C. The Interplay between Candida albicans, Vaginal Mucosa, Host Immunity and Resident Microbiota in Health and Disease: An Overview and Future Perspectives. Microorganisms 2023, 11, 1211. [Google Scholar] [CrossRef]
- Miravitlles, M.; Anzueto, A. Chronic Respiratory Infection in Patients with Chronic Obstructive Pulmonary Disease: What Is the Role of Antibiotics? Int. J. Mol. Sci. 2017, 18, 1344. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Zhao, L.; Wang, C.; Wu, X.; Yan, H. Clinical characteristics of septic pulmonary embolism in adults: A systematic review. Respir. Med. 2014, 108, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Berkow, E.L.; Lockhart, S.R. Fluconazole resistance in Candida species: A current perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- Hassanmoghadam, F.; Shokohi, T.; Hedayati, M.T.; Aslani, N.; Haghani, I.; Nabili, M.; Lotfali, E.; Davari, A.; Moazeni, M. High prevalence of itraconazole resistance among Candida parapsilosis isolated from Iran. Curr. Med. Mycol. 2019, 5, 43–46. [Google Scholar] [CrossRef]
- Chandrasekar, P.H.; Sobel, J.D. Micafungin: A new echinocandin. Clin. Infect. Dis. 2006, 42, 1171–1178. [Google Scholar] [CrossRef]
- Syed, Y.Y. Rezafungin: First Approval. Drugs 2023, 83, 833–840. [Google Scholar] [CrossRef]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Cumbie, R.; Driscoll, E.; Pasculle, A.W.; Clancy, C.J. Rate of FKS Mutations among Consecutive Candida Isolates Causing Bloodstream Infection. Antimicrob. Agents Chemother. 2015, 59, 7465–7470. [Google Scholar] [CrossRef]
- Coste, A.T.; Kritikos, A.; Li, J.; Khanna, N.; Goldenberger, D.; Garzoni, C.; Zehnder, C.; Boggian, K.; Neofytos, D.; Riat, A.; et al. Fungal Infection Network of Switzerland (FUNGINOS). Emerging echinocandin-resistant Candida albicans and glabrata in Switzerland. Infection 2020, 48, 761–766. [Google Scholar] [CrossRef]
- Carolus, H.; Pierson, S.; Lagrou, K.; Van Dijck, P. Amphotericin B and Other Polyenes—Discovery, Clinical Use, Mode of Action and Drug Resistance. J. Fungi 2020, 6, 321. [Google Scholar] [CrossRef]
- Chandrasekar, P. Management of invasive fungal infections: A role for polyenes. J. Antimicrob. Chemother. 2011, 66, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Posch, W.; Blatzer, M.M.; Wilflingseder, D.; Lass-Flörl, C. Aspergillus terreus: Novel lessons learned on amphotericin B resistance. Med. Mycol. 2018, 56, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Escandon, P.; Chow, N.A.; Caceres, D.H.; Gade, L.; Berkow, E.L.; Armstrong, P.; Rivera, S.; Misas, E.; Duarte, C.; Moulton-Meissner, H.; et al. Molecular epidemiology of Candida auris in Colombia reveals a highly related, countrywide colonization with regional patterns in amphotericin B resistance. Clin. Infect. Dis. 2019, 68, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guideline for the diagnosis and management of mucormycosis: An initiative of the European Confederation of Medical Mycology in cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef] [PubMed]
- Sigera, L.S.M.; Denning, D.W. Flucytosine and its clinical usage. Ther. Adv. Infect. Dis. 2023, 10, 20499361231161387. [Google Scholar] [CrossRef]
- Vermes, A.; Guchelaar, H.J.; Dankert, J. Flucytosine: A review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 2000, 46, 17–19. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hansen, A.; Lass-Flörl, C.; Lackner, M.; Rare Yeast Study Group. Antifungal susceptibility profiles of rare ascomycetous yeasts. J. Antimicrob. Chemother. 2019, 74, 2649–2656. [Google Scholar] [CrossRef]
- Álvarez-Uría, P.; Muñoz, A.; Vena, J.; Guinea, L.J.; Marcos-Zambrano, P.; Escribano, C.; Sánchez-Carrillo, E.; Bouza, Collaborative Study Group of Mycology (COMIC). Fungaemia caused by rare yeasts: Incidence, clinical characteristics and outcome over 10 years. J. Antimicrob. Chemother. 2018, 73, 823–825. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Letscher-Bru, V.; Neuvéglise, C.; Dromer, F. Yarrowia lipolytica causes sporadic cases and local outbreaks of infections and colonisation. Mycoses 2020, 63, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, A.A.; Pérez-Hansen, A.; Lackner, M.; Lass-Flörl, C.; Boekhout, T. Elevated minimum inhibitory concentrations to antifungal drugs prevail in 14 rare species of candidemia-causing Saccharomycotina yeasts. Med. Mycol. 2020, 58, 987–995. [Google Scholar] [CrossRef]
- Lavergne, R.A.; Barbier, P.; Mobuchon, L.; Banerjee, A.; Prasad, R.; Morio, F. Draft Genome Sequence of the Fluconazole-Resistant Yarrowia lipolytica Clinical Isolate CBS 18115. Microbiol. Resour. Announc. 2023, 12, e0126022. [Google Scholar] [CrossRef]
- Yu, J.; Liu, X.; Guo, D.; Yang, W.; Chen, X.; Zou, G.; Wang, T.; Pang, S.; Zhang, G.; Dong, J.; et al. Antifungal susceptibility profile and local epidemiological cut-off values of Yarrowia (Candida) lipolytica: An emergent and rare opportunistic yeast. Microbiol. Spectr. 2024, 12, e0320323. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S612–S617. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G. Rezafungin—Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins. J. Fungi 2020, 6, 262. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Escribano, P.; Sánchez, C.; Muñoz, P.; Bouza, E.; Guinea, J. Antifungal resistance to fluconazole and echinocandins is not emerging in yeast isolates causing fungemia in a Spanish tertiary care center. Antimicrob. Agents Chemother. 2014, 58, 4565–4572. [Google Scholar] [CrossRef] [PubMed]
- Díaz-García, J.; Mesquida, A.; Sánchez-Carrillo, C.; Reigadas, E.; Muñoz, P.; Escribano, P.; Guinea, J. Monitoring the Epidemiology and Antifungal Resistance of Yeasts Causing Fungemia in a Tertiary Care Hospital in Madrid, Spain: Any Relevant Changes in the Last 13 Years? Antimicrob. Agents Chemother. 2021, 65, e01827-20. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Zerbato, V.; Sincovich, S.; Cosimi, L.; Zorat, F.; Costantino, V.; Di Santolo, M.; Busetti, M.; Di Bella, S.; Principe, L.; et al. Candida lipolytica Bloodstream Infection in an Adult Patient with COVID-19 and Alcohol Use Disorder: A Unique Case and a Systematic Review of the Literature. Antibiotics 2023, 12, 691. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.D.; Robbins, N.; Zaas, A.K.; Schell, W.A.; Perfect, J.R.; Cowen, L.E. Hsp90 Governs Echinocandin Resistance in the Pathogenic Yeast Candida albicans via Calcineurin. PLoS Pathog. 2009, 5, e1000532. [Google Scholar] [CrossRef] [PubMed]
- Sardi, J.C.O.; Scorzoni, L.; Bernardi, T.; Fusco-Almeida, A.M.; Mendes Giannini, M.J. Candida species: Current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J. Med. Microbiol. 2013, 62, 10–24. [Google Scholar] [CrossRef]
- Gómez-Gaviria, M.; Ramírez-Sotelo, U.; Mora-Montes, H.M. Non-albicans Candida Species: Immune Response, Evasion Mechanisms, and New Plant-Derived Alternative Therapies. J. Fungi 2022, 9, 11. [Google Scholar] [CrossRef]
- Singh, D.P.; Kumar Verma, R.; Sarswat, S.; Saraswat, S. Non-Candida albicans Candida species: Virulence factors and species identification in India. Curr. Med. Mycol. 2021, 7, 8–13. [Google Scholar] [CrossRef]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Pakshir, K.; Zomorodian, K.; Karamitalab, M.; Jafari, M.; Taraz, H.; Ebrahimi, H. Phospholipase, esterase and hemolytic activities of Candida spp. isolated from onychomycosis and oral lichen planus lesions. J. Mycol. Med. 2013, 23, 113–118. [Google Scholar] [CrossRef]
- Yigit, N.; Aktas, E.; Dagistan, S.; Ayyildiz, A. Investigating biofilm production, coagulase and hemolytic activity in Candida species isolated from denture stomatitis patients. Eurasian J. Med. 2011, 43, 27–32. [Google Scholar] [CrossRef] [PubMed]
- de Souza Ramos, L.; Barbedo, L.S.; Braga-Silva, L.A.; dos Santos, A.L.S.; Pinto, M.R.; da Graça Sgarbi, D.B. Protease and phospholipase activities of Candida spp. Isolated from cutaneous candidiasis. Rev. Iberoam Micol. 2015, 32, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Price, M.F.; Cawson, R.A. Phospholipase activity in Candida albicans. Sabouraudia 1977, 15, 179–185. [Google Scholar] [CrossRef]
- Nayak, A.P.; Green, B.J.; Beezhold, D.H. Fungal hemolysins. Med. Mycol. 2013, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [PubMed]
- Nouraei, H.; Jahromi, M.G.; Jahromi, L.R.; Zomorodian, K.; Pakshir, K. Potential Pathogenicity of Candida Species Isolated from Oral Cavity of Patients with Diabetes. Biomed. Res. Int. 2021, 2021, 9982744. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; Lopez-Ribot, J.L. Candida biofilms: An update. Eukaryot. Cell. 2005, 4, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Pistoia, E.S.; Cosio, T.; Campione, E.; Pica, F.; Volpe, A.; Marino, D.; Di Francesco, P.; Monari, C.; Fontana, C.; Favaro, M.; et al. All-Trans Retinoic Acid Effect on Candida albicans Growth and Biofilm Formation. J. Fungi 2022, 8, 1049. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Dzajic, E.; Jensen, R.H.; Johansen, H.K.; Kjældgaard, P.; Knudsen, J.D.; Kristensen, L.; Leitz, C.; Lemming, L.E.; Nielsen, L.; et al. Epidemiological changes with potential implication for antifungal prescription recommendations for fungaemia: Data from a nationwide fungaemia surveillance programme. Clin. Microbiol. Infect. 2013, 19, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Seker, E. Identification of Candida species isolated from bovine mastitic milk and their in vitro hemolytic activity in Western Turkey. Mycopathologia 2010, 169, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Leite de Andrade, M.C.; Soares de Oliveira, M.A.; Santos, F.A.G.D.; Ximenes Vilela, P.; da Silva, M.N.; Macêdo, D.P.C.; de Lima Neto, R.G.; Neves, H.J.P.; Brandão, I.S.L.; Chaves, G.M.; et al. A New approach by optical coherence tomography for elucidating biofilm formation by emergent Candida species. PLoS ONE 2017, 12, e0188020. [Google Scholar] [CrossRef] [PubMed]
- Kühbacher, A.; Burger-Kentischer, A.; Rupp, S. Interaction of Candida Species with the Skin. Microorganisms 2017, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Juayang, A.C.; Lim, J.P.T.; de los Reyes, Z.C.B.; Tuante, M.D.B.; Batiles, Z.I.P.; Guino-o, J.F.N.V.; Villanueva, F.R.A.; de los Reyes, B.B. Antifungal Resistance of Candida Species in Bacolod City, Philippines. J. Infect. Dis. Epidemiol. 2019, 5, 076. [Google Scholar]
- Pendleton, K.M.; Huffnagle, G.B.; Dickson, R.P. The significance of Candida in the human respiratory tract: Our evolving understanding. Pathog. Dis. 2017, 75, ftx029. [Google Scholar] [CrossRef]
- Foroozan, M.; Contet-Audonneau, N.; Granel-Brocard, F.; Barbaud, A.; Luc Schmutz, J. Prevalence analysis of fungi in chronic lower extremity ulcers. Wounds 2011, 23, 68–75. [Google Scholar]
- Shivanna, K.R. Climate change and its impact on biodiversity and human welfare. Proc. Indian Natl. Sci. Acad. 2022, 88, 160–171. [Google Scholar] [CrossRef]
- van Rhijn, N.; Bromley, M. The Consequences of Our Changing Environment on Life Threatening and Debilitating Fungal Diseases in Humans. J. Fungi 2021, 7, 367. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Sood, P.; Rudramurthy, S.M.; Chen, S.; Kaur, H.; Capoor, M.; Chhina, D.; Rao, R.; Eshwara, V.K.; Xess, I.; et al. Incidence, characteristics and outcome of ICU-acquired candidemia in India. Intensive Care Med. 2015, 41, 285–295. [Google Scholar] [CrossRef]
- One Health High-Level Expert Panel (OHHLEP); Hayman, D.T.S.; Adisasmito, W.B.; Almuhairi, S.; Behravesh, C.B.; Bilivogui, P.; Bukachi, S.A.; Casas, N.; Becerra, N.C.; Charron, D.F.; et al. Developing One Health surveillance systems. One Health 2023, 17, 100617. [Google Scholar] [CrossRef]
- Litwinowicz, A.; Blaszkowska, J. Hirudo verbana is a source of fungal isolates potentially pathogenic to humans. Afr. J. Microbiol. Res. 2013, 7, 5358–5363. [Google Scholar]
- Gomes Ados, R.; Cabana, Â.; Osório Lda, G.; Santin, R.; Schuch, I.D.; Serra, E.F.; Nascente, P.S.; Meireles, M.C. First isolation of the Stephanoascus ciferrii in feline otitis in Brazil. Braz. J. Microbiol. 2014, 45, 11011–11103. [Google Scholar] [CrossRef]
- Ulloa, M.; Lappe, P.; Aguilar, S.; Park, H.; Pérez-Mejía, A.; Toriello, C.; Taylor, M.L. Contribution to the study of the mycobiota present in the natural habitats of Histoplasma capsulatum: An integrative study in Guerrero, Mexico. Rev. Mex. Biodivers. 2006, 77, 153–168. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W. The Yeasts, A Taxonomic Study, 4th ed.; Elsevier: Amsterdam, Netherlands, 1998; 1055p. [Google Scholar]
- Różański, P.; Slaska, B.; Różańska, D. Prevalence of yeasts in English full blood mares. Mycopathologia 2013, 175, 339–344. [Google Scholar] [CrossRef]
- Pławińska-Czarnak, J.; Wódz, K.; Strzałkowska, Z.; Żychska, M.; Nowak, T.; Kwieciński, A.; Kwieciński, P.; Bielecki, W.; Rodo, A.; Rzewuska, M.; et al. Comparison of automatic methods MALDI-TOF, VITEK2 and manual methods for the identification of intestinal microbial communities on the example of samples from alpacas (Vicugna pacos). J. Vet. Res. 2023, 67, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Větrovský, T.; Morais, D.; Kohout, P.; Lepinay, C.; Algora Gallardo, C.; Awokunle Hollá, S.; Doreen Bahnmann, B.; Bílohnědá, K.; Brabcová, V.; D’Alò, F.; et al. GlobalFungi, a global database of fungal occurrences from high-throughput-sequencing metabarcoding studies. Sci. Data 2020, 7, 228. [Google Scholar] [CrossRef] [PubMed]
- Baublis, J.A.; Wharton, R.A.; Volz, P.A. Diversity of micro-fungi in an Antarctic dry valley. J. Basic Microbiol. 1991, 31, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.M.; Spencer, J.F.T.; de Figueroa, L.I.; Garro, O.; Fengler, E. Yeasts associated with pods and exudates of algarrobo trees (Prosopis spp.) and species of columnar cacti in northwest Argentina. Appl. Microbiol. Biotechnol. 1996, 44, 736–739. [Google Scholar]
- Schroeder, J.W.; Martin, J.T.; Angulo, D.F.; Barbosa, J.M.; Perea, R.; Arias-Del Razo, I.; Sebastián-González, E.; Dirzo, R. Community composition and diversity of Neotropical root-associated fungi in common and rare trees. Biotropica 2018, 50, 694–703. [Google Scholar] [CrossRef]
- Schappe, T.; Albornoz, F.E.; Turner, B.L.; Neat, A.; Condit, R.; Jones, F.A. The role of soil chemistry and plant neighbourhoods in structuring fungal communities in three Panamanian rainforests. J. Ecol. 2017, 105, 569–579. [Google Scholar] [CrossRef]
- Wainwright, B.J.; Zahn, G.L.; Arlyza, I.S.; Amend, A.S. Seagrass-associated fungal communities follow Wallace’s line, but host genotype does not structure fungal community. J. Biogeogr. 2018, 45, 762–770. [Google Scholar] [CrossRef]
- Kueffer, C.; Drake, D.R.; Fernández-Palacios, J.M. Island biology: Looking towards the future. Biol. Lett. 2014, 10, 20140719. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Maitra, P.; Gan, H.Y.; Chen, L.; Li, S.; Tu, T.; Chen, L.; Mi, X.; Gao, C.; Zhang, D.; et al. Soil fungal diversity and community assembly: Affected by island size or type? FEMS Microbiol. Ecol. 2021, 97, fiab062. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, C.L.; Vann, L.E.; Eisen, J.A. Global Diversity and Biogeography of the Zostera marina Mycobiome. Appl. Environ. Microbiol. 2021, 87, e0279520. [Google Scholar] [CrossRef] [PubMed]
- Kocková-Kratochvílová, A.; Sláviková, E. Candida mucifera n. sp. J. Basic Microbiol. 1998, 28, 613–618. [Google Scholar] [CrossRef]
- Shoubao, Y.; Jie, Y.; TingTing, S.; Jiaquan, G.; Cuie, S. Yeast diversity in pit mud and related volatile compounds in fermented grains of chinese strong-flavour liquor. AMB Express 2023, 13, 56. [Google Scholar] [CrossRef]
- Brilhante, R.S.; Paiva, M.A.; Sampaio, C.M.; Castelo-Branco, D.S.; Teixeira, C.E.; de Alencar, L.P.; Bandeira, T.J.; Monteiro, A.J.; Cordeiro, R.A.; Pereira-Neto, W.A.; et al. Azole resistance in Candida spp. isolated from Catú Lake, Ceará, Brazil: An efflux-pump-mediated mechanism. Braz. J. Microbiol. 2016, 47, 33–38. [Google Scholar] [CrossRef]
- Castelo-Branco, D.S.C.M.; Paiva, M.A.N.; Teixeira, C.E.C.; Caetano, E.P.; Guedes, G.M.M.; Cordeiro, R.A.; Brilhante, R.S.N.; Rocha, M.F.G.; Sidrim, J.J.C. Azole resistance in Candida from animals calls for the One Health approach to tackle the emergence of antimicrobial resistance. Med. Mycol. 2020, 58, 896–905. [Google Scholar] [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Fungal Priority Pathogens List to guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022; ISBN 978-92-4-006024-1.
- Vitiello, A.; Ferrara, F.; Boccellino, M.; Ponzo, A.; Cimmino, C.; Comberiati, E.; Zovi, A.; Clemente, S.; Sabbatucci, M. Antifungal Drug Resistance: An Emergent Health Threat. Biomedicines 2023, 11, 1063. [Google Scholar] [CrossRef]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Jackson, B.R.; Chow, N.; Forsberg, K.; Litvintseva, A.P.; Lockhart, S.R.; Welsh, R.; Vallabhaneni, S.; Chiller, T. On the Origins of a Species: What Might Explain the Rise of Candida auris? J. Fungi 2019, 5, 58. [Google Scholar] [CrossRef]
- Jia, X.; Liang, Q.; Chi, F.; Cao, W. Otomycosis in Shanghai: Aetiology, clinical features and therapy. Mycoses 2012, 55, 404–409. [Google Scholar] [CrossRef]
- Shweihat, Y.; Perry, J., 3rd; Shah, D. Isolated Candida infection of the lung. Respir. Med. Case Rep. 2015, 16, 18–19. [Google Scholar] [CrossRef]
- Farooq, H.; Monowar, T.; Latt, S.S.; Chinni, S.V.; Zainol, N.H.; Sabesan, G.S. A Review of Studies on Candida Species in Peninsular Malaysia. Adv. Biomed. Res. 2022, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Nouri, H.; Moghimi, H.; Geranpayeh Vaghei, M.; Nasr, S. Blastobotrys persicus sp. nov., an ascomycetous yeast species isolated from cave soil. Antonie Van Leeuwenhoek 2018, 111, 517–524. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Years | Clinical Presentation | Sex | Ethnicity | Age (Years) | Immunity State | Underlying Conditions | Initial Symptoms/Signs | Outcomes | Treatment | Resistance (MIC) | Identification | Country |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Furman and Ahearn [16] | 1983 | Otomycosis | Male | NA | 25 | NA | NA | Oesophageal pain. Headaches. Ear pressure | Recovery | 1% hydrocortisone; 2.0% acetic acid solution | NA | Morphological and physiological identification | USA (Kansas) |
| Onychomycosis | Female | NA | NA | NA | NA | NA | NA | NA | NA | USA (Hawaii) | |||
| NA | Male | NA | NA | NA | NA | NA | NA | NA | NA | USA (Oregon) | |||
| Gangrenous foot | Female | NA | 68 | NA | NA | NA | NA | NA | NA | USA (Geogia) | |||
| Tinea pedis | NA | NA | NA | NA | NA | NA | NA | NA | NA | USA (Florida) | |||
| De Gentile et al. [13] | 1991 | Toenail onyxis | NA | NA | 52 | NA | NA | NA | NA | NA | NA | ATB 32C (API system) | France |
| 1991 | Toenail onyxis | NA | NA | 69 | NA | NA | NA | NA | NA | NA | France | ||
| 1991 | Toenail onyxis | Male | NA | 88 | NA | Auricular fibrillation. Extensive cutaneous ulceration | Ingrowing toenails | NA | NA | Flu; Flucy (microdilution) | France | ||
| 1991 | Onychomycosis | Male | NA | 70 | NA | Arteriopathy. Alcoholic. Smoker. Venous insufficiency with large, perforated ulcers | Onychopathy | NA | NA | NA | France | ||
| 1991 | Toenail onyxis | NA | NA | 84 | NA | Trophic disorders. Nummular eczema. Farmer | NA | NA | NA | NA | France | ||
| 1991 | Toenail onyxis | Male | NA | 75 | Immunocompromised | Diabetes. Rheumatism. Valvulopathy. Ulceration legs. Tinea pedis | Tinea pedis. Onyxis. Tinea cruris | NA | NA | NA | France | ||
| De Gentile et al. [26] | 1995 | Onychomycosis | 12 patients | NA | 35–95 (mean: 74) | NA | Trophic disorders | NA | NA | NA | Iso; Flu; Itra | API 20C AUX; ID 32C | France |
| Gunsilius et al. [27] | 2001 | Invasive candidiasis | Male | Caucasian | 62 | Immunocompromised | Acute myeloid leukaemia (AML, FAB-M1) | Fever. Erythematous skin papules | Exitus | Liposomal AmB (5.3 g) | Flu (>64 µg/mL) | API 20C AUX | Austria |
| Cheng et al. [28] | 2004 | Candidemia | Male | NA | NA | Immunocompetent | NA | NA | Recovery | NA | Flu (16 µg/mL) | API-32C | China (Taiwan) |
| Agin et al. [19] | 2011 | Candidemia | Male | Turkey | 8 | Immunocompetent | Down Syndrome. Cerebral palsy | Respiratory distress. Cough. Fever. Decubitus ulcer on the right gluteal region | Exitus | Lipid Complex AmB | AmB (1 µg/mL); Flu (64 µg/mL); Caspo (32 µg/mL); Anidu (32 µg/mL) | Vitek2 | Turkey |
| Saha et al. [29] | 2013 | Pneumonia | Female | NA | 55 | Immunocompromised | Bidi smoker. COPD. Diabetes. | Dyspnoea. Mucopurulent cough | Recovery | Intravenous liposomal AmB (150 mg daily) was initiated and changed to oral Flu (150 mg daily) after 4 days | No | Vitek2 | India |
| Daielescu et al. [20] | 2014 | Endophthalmitis | Female | East Europe | 57 | Immunocompetent | Cataract | VA decreased after cataract surgery | Recovery | 50 µg/0.1 mL and then 250 µg/0.1 mL intravitreal Caspo | Flu Vori AmB | Vitek2 | Romania |
| Soki et al. [17] | 2015 | Infraorbital abscess | Male | NA | 79 | Immunocompromised | Enucleation of choroidal melanoma | Infraorbital abscess | Recovery | Topical AmB 0.1% | Flu (64 μg/mL) | 18S rDNA, ITS1-26S rDNA D1/D2 region sequence | Japan |
| Vilanueva-Lozano et al. [30] | 2016 | Candidemia | Female | Mexican | 30 | Immunocompromised | Crohn’s disease. Mycobacterium bovis disease | Headache. Fever. Disorientation | Recovery | Posa | Flu (32 µg/mL) | Vitek2 | Mexico |
| Dave et al. [31] | 2018 | Endophthalmitis | Female | NA | 50 | Immunocompetent | Chronic post-cataract surgery endophthalmitis | IOL plaque | Recovery | Oral keto tablet (200 mg) twice/d with topical Nata 5% eye drops and intravitreal injection of AmB (5 mg/0.1 mL) | AmB | Vitek2 | India |
| 2018 | Endophthalmitis | Female | NA | 59 | NA | Chronic post-cataract surgery endophthalmitis | Mutton-fat keratic precipitates. Vitritis. AC membranes | Recovery | Intravitreal AmB injection (5 mg/0.1 mL) | NA | Vitek2 | India | |
| 2018 | Endophthalmitis | Female | NA | 79 | NA | Chronic post-cataract surgery endophthalmitis | Panuveitis | Recovery | Intravitreal AmB and oral keto (200 mg) 2 times/day | NA | Vitek2 | India | |
| 2018 | Endophthalmitis | Male | NA | 66 | NA | Chronic post-cataract surgery endophthalmitis | Hypopyon. Low-grade vitritis | Recovery | Oral Itra tablet (100 mg) twice/d with topical Nata 5% eye drops, and intravitreally, an injection of AmB (5 mg/0.1 mL) | NA | Vitek2 | India | |
| Bansal et al. [32] | 2021 | Endophthalmitis | Female | Asian Indian | 26 | Immunocompetent | Uncomplicated vaginal delivery | VA decreased | Recovery | Flu (3 months) | No | Vitek2 | India |
| Guo et al. [25] | 2021 | Otomycosis | 17 males; 15 females | NA | Mean: 36 | NA | NA | CSOM | NA | NA | 13 (40.6%) Flucy 32 (100%) Flu 32 and 256 μg/mL | ITS region rDNA sequence | China |
| Robles-Tenorio et al. [33] | 2022 | Onychomycosis and candidemia | Male | Mexican | 58 | Immunocompromised | Diabetes mellitus type 2 (DM2) | Cough. Headache. Dyspnoea | Exitus | AmB | Flu | Vitek2 | Mexico |
| Sathi et al. [34] | 2022 | 9 candidemia; 2 ocular diseases; 1 VCC | 12 patients | NA | NA | NA | NA | NA | NA | NA | Blood samples: 6 resistant to Flu, Itra; 1 resistant to Vori; 5 resistant to Amb. From eye: 1 resistant to Flu, Itra, Cotri. From HVG: 1 resistant to Flu, Itra, Vori, AmB | Bangladesh | |
| Ng et al. [35] | 2013 (cases from 2000) | Vaginal candidiasis | 3 females | NA | NA | NA | NA | NA | NA | NA | NA | ITS1-5.8S-ITS2 region | Malaysia |
| Romald et al. [36] | 2019 | CSOM | Female | Indian | 57 | Immunocompetent | Post-mastoidectomy | Ear pain, profuse ear discharge, and hard of hearing in left ear for past three months | Recovery | Oral Vori and topical Clotri | Flu > 32 μg/mL Itra ≥ 16 μg/mL Vori < 0.025 μg/mL AmB ≥ 16 μg/mL Posa ≤ 0.025 μg/mL Caspo ≤ 0.025 μg/mL | ITS1 4 | India |
| Demiray et al. [37] | 2015 | Candidemia | Female | NA | Newborn, 23 days | NA | Neonatal diaphragmic hernia | Systemic mycosis. Cutaneous lesions. Respiratory distress | Recovery | Flu (1 × 21 mg/d) | AmB ≤ 0.25 µg/mL Flucy ≤ 1 µg/mL Flu ≤ 1 µg/mL Vori < 0.12 µg/mL | API system | Turkey |
| Papìla et al. [38] | 2016 | Septic pulmonary embolism | Female | NA | 70 | NA | Patient with type 2 DM | Altered consciousness. Speech disorder. Vomiting. Loss of power in the left. Confusion. Pupillary isocoria. Deep tendon reflexes (DTRs). Direct light reflex (DLR) (++/++). Left hemiplegia | Recovery | Anidu (200 mg daily loading dose and 100 mg daily maintenance dose) for 4 weeks; treatment was discharged with 400 mg Flu (2 weeks) | NA | Vitek2 | Turkey |
| Chan et al. [39] | 2015 | Granular myringitis | 3 males; 1 female | NA | Mean: 38 Range:33–42 | 3 immunocompetent; 1 immunosuppressed | 3 no comorbidities. 1 nasopharyngeal carcinoma, diabetes mellitus, and hyperlipidaemia | Obstructive sensation and/or pruritis in ears. Pustular otorrhoea. Otalgia | Recovery | Responses to treatment with irrigation and topical antifungal cream are generally good | Flu; AmB; Flucy | Vitek2 YST ID Card system. API 20C AUX; MALDI-TOF MS; ITS | China (Hong Kong) |
| Vieira de Melo et al. [40] | 2019 (blood collection from 2011 to 2015) | Candidemia | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | Brazil |
| Capoor et al. [41] | 2015 | Fungemia | Female | NA | 30 | Immunosuppressed | AML Fluconazole prophylaxis | NA | Expired | NA | Resistant to AmB, Flu, Itra | API 20C AUX | India |
| Reference | Medium | Texture | Colour |
|---|---|---|---|
| Furman and Ahearn, 1983 [16]; Kano et al., 2000 [42] | SDA | Small; round; could be tough, raised, and wrinkled or smooth | Milky white or cream |
| Romald et al., 2019 [36] | TTZ | Dry, rough, wrinkled | Pink to white |
| PDA | Rough | Non-pigmented | |
| OMA | Wrinkled | White | |
| CMA | Dry, rough, wrinkled | ||
| HiCrome™ | Dry, wrinkled | Blue-white | |
| Guo et al., 2021 [25] | CHROMagar™ | Regular round | Blue centre and white edge |
| Misidentification | ||||
|---|---|---|---|---|
| Reference | Isolated Pathogen | Biochemical | Molecular | Proteomic |
| Balada-Llasat et al., 2012 [49] | Candida ciferrii | Cryptococcus neoformans | ||
| Kumar et al., 2014 [44] | Blastobotrys serpentis | Stephanoascus ciferrii | ||
| Blastobotrys proliferans | Stephanoascus ciferrii | |||
| Roehmel et al., 2015 [48] | Arxula adeninivorans | Stephanoascus ciferrii | ||
| Warren et al., 2017 [14] | Cryptococcus laurentii | Candida ciferrii | ||
| A. Al-Haqqan et al., 2018 [45] | Candida blankii | Candida ciferrii | ||
| N° of Resistant Isolates of S. ciferrii Complex | Flucy | Iso | Flu | Itra | Vori | AmB | Caspo | Anidu | |
|---|---|---|---|---|---|---|---|---|---|
| Skin and appendages | 22 | 1 (4.5%) | 12 (54%) | 14 (64%) | 12 (54%) | 0 | 0 | 0 | 0 |
| Blood | 17 | 0 | 0 | 12 (71%) | 7 (41%) | 1 (6%) | 7 (41%) | 1 (6%) | 1 (6%) |
| Ear | 41 | 17 (41%) | 0 | 41 (100%) | 1 (2%) | 0 | 4 (8%) | 0 | 0 |
| Eye | 7 | 0 | 0 | 2 (29%) | 0 | 1 (14%) | 3 (43%) | 0 | 0 |
| Lung | 2 | NA | NA | NA | NA | NA | NA | NA | NA |
| Genital mucosa | 4 | 0 | 0 | 1 (25%) | 1 (25%) | 1 (25%) | 1 (25%) | 0 | 0 |
| Reference | Years | Clinical Presentation | Sex | Ethnicity | Age (Years) | Immunity State | Underlying Conditions | Resistance (MIC) | Identification | Country |
|---|---|---|---|---|---|---|---|---|---|---|
| Foroozan et al. [105] | 2011 | NA | NA | NA | NA | NA | Low-extremity cutaneous ulcers | NA | NA | France |
| Juayang et al. [103] | 2017–2018 (published 2019) | 19 patients | NA | NA | 17 cases > 55 years old; 2 cases < 55 years old | NA | NA | 42.1% Fluco; 38.9% Voria | VITEK 2 | Philippines |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cosio, T.; Pica, F.; Fontana, C.; Pistoia, E.S.; Favaro, M.; Valsecchi, I.; Zarabian, N.; Campione, E.; Botterel, F.; Gaziano, R. Stephanoascus ciferrii Complex: The Current State of Infections and Drug Resistance in Humans. J. Fungi 2024, 10, 294. https://doi.org/10.3390/jof10040294
Cosio T, Pica F, Fontana C, Pistoia ES, Favaro M, Valsecchi I, Zarabian N, Campione E, Botterel F, Gaziano R. Stephanoascus ciferrii Complex: The Current State of Infections and Drug Resistance in Humans. Journal of Fungi. 2024; 10(4):294. https://doi.org/10.3390/jof10040294
Chicago/Turabian StyleCosio, Terenzio, Francesca Pica, Carla Fontana, Enrico Salvatore Pistoia, Marco Favaro, Isabel Valsecchi, Nikkia Zarabian, Elena Campione, Françoise Botterel, and Roberta Gaziano. 2024. "Stephanoascus ciferrii Complex: The Current State of Infections and Drug Resistance in Humans" Journal of Fungi 10, no. 4: 294. https://doi.org/10.3390/jof10040294