
Microsporidiosis in Vertebrate Companion Exotic Animals

1
Zoological medicine service, Faculté de médecine vétérinaire, Université de Montréal, 3200 Sicotte, Saint-Hyacinthe, QC J2S2M2, Canada
2
Clinique Vétérinaire Benjamin Franklin, 38 rue du Danemark, ZA Porte Océane, 56400 Brech, France
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
J. Fungi 2016, 2(1), 3; https://doi.org/10.3390/jof2010003
Submission received: 13 October 2015
/
Revised: 7 December 2015
/
Accepted: 18 December 2015
/
Published: 24 December 2015
Abstract
:Veterinarians caring for companion animals may encounter microsporidia in various host species, and diagnosis and treatment of these fungal organisms can be particularly challenging. Fourteen microsporidial species have been reported to infect humans and some of them are zoonotic; however, to date, direct zoonotic transmission is difficult to document versus transit through the digestive tract. In this context, summarizing information available about microsporidiosis of companion exotic animals is relevant due to the proximity of these animals to their owners. Diagnostic modalities and therapeutic challenges are reviewed by taxa. Further studies are needed to better assess risks associated with animal microsporidia for immunosuppressed owners and to improve detection and treatment of infected companion animals.
1. Introduction
Microsporidia are eukaryotic organisms with the smallest known genome [1]. Microsporidia had been classified as amitochondriate due to their lack of visible mitochondria, but sequences homologous to genes coding for mitochondria have since been discovered in their genome and remnants of mitochondria have been visualized in their cytoplasm [2]; therefore, they have been reclassified as fungi based on phylogenic analysis of multiple proteins in their genome, clustering preferentially with fungal proteins [2,3]. Currently, microsporidia are considered atypical fungi without mitochrondria [2,4,5,6]. More than 1200 species are described, some of which are able to infect multiple host species [7,8]. A comparative review is relevant for microsporidia, as diagnostic tests and potential treatments are common to many hosts, and veterinarians caring for companion animals may encounter these fungi in various host species.
Microsporidia were first reported in the middle of the 19th century when an outbreak threatened the silkworm industry [9,10]. The majority of microsporidia species infect arthropods [11]. Most species infecting vertebrate hosts are found in fish [11]; however, companion animals that are commonly seen by exotic veterinary practitioners, such as rabbits, rodents, birds and reptiles, can also be infected by microsporidia. Most microsporidia are obligate intracellular fungi [8,12]. Members of this phylum exclusively reproduce asexually [13]. Some microsporidia form xenomas, which are single hypertrophied host cells containing multiple microsporidia. Xenomas can reach 14 mm in diameter [14], making them macroscopically visible. These large xenomas have been described as “pseudotumors” [8,12]. Microsporidial spores are smaller than their protozoan counterparts and differ from them by being Gram-positive or Gram-variable [12]. Microsporidia often represent a diagnostic challenge because many microsporidial genera do not form xenoma and infection may be subclinical.
A typical microsporidian cycle involves three phases: the infective phase, which is characterized by polar tube extrusion and injection of the sporoplasm into the cytoplasm of a host cell; the proliferation phase or merogony; and, finally, the spore-forming phase or sporogony [12,15]. Spores can survive for over a year in the environment [16] and disinfection is necessary to inactivate them. Ultraviolet-sterilization of water at 6 mJ/cm2 has been shown to eliminate some microsporidial spores of concern to human health [16]. In contrast, some microsporidia are highly resistant to chlorine disinfection [17]. Spores measure 3 to 5 μm in diameter and are difficult to identify by light microscopy. Microsporidian genera can be classified based upon their ability to form xenomas [8] and upon whether or not spores develop in membrane-bound packets called sporophorous vesicles [12]. Identification of the precise species of microsporidia is based on the target tissue infected, the size and morphology of the spores, including the number of nuclei [18], the presence or absence of sporophorous vesicles and diplokarya or paired nuclei [14], the number of spores in sporophorous vesicles, if present [13], and the structure of xenomas, if present [14]. Currently, molecular diagnostic tests using polymerase chain reaction (PCR) with primers targeting the small subunit ribosomal RNA gene are required to confirm identification [19].
Diagnosis can be reached via direct visualization of typical spores by light or electronic microscopy, via cell culture, tissue histology, immunohistochemistry, or via serologic [20] and molecular techniques [21]. Microsporidian spores are Gram-positive or Gram-variable [14], birefringent and some of them are acid-fast-positive [12]. Chitin-binding fluorochromes [12,22,23], chromotrope stains [21], modified trichrome stain, and Giemsa stain [21] can also be used to stain microsporidial spores on smears and stool samples, which has been demonstrated in humans [22]. Notably, fluorescent stains, such as the Luna calcofluor stain, can be used since microsporidian spores contain chitin, but these stains require immunofluorescence microscopy and may lead to false positives since other fungal spores also fluoresce with these stains [24]. Molecular diagnostic techniques include PCR of the small subunit rRNA gene using PMP1 and PMP2 primers [25,26]. It is challenging to determine the actual prevalence of microsporidia infection in a population as shedding is typically intermittent [24] and transit of the fungi through the feces cannot be differentiated from digestive infection based on fecal detection. Of note, the intensity of fecal shedding, and thus the sensibility of fecal screening, has been correlated with severity of infection by intestinal microsporida in humans but has not been correlated with the occurrence of diarrhea [27]. These challenges are illustrated by the high variability of prevalence rates, even from studies derived from the same geographic area [28].
Fourteen microsporidial species have been shown to infect humans [28,29], with 90% of human cases being caused by Enterocytozoon bieneusi [30]. Suspected risk factors for microsporidiosis in human include young age [31,32], immunosuppression (e.g., acquired immune deficiency syndrome with low CD4 T lymphocytes [33]), treatment with immunosuppressive drugs [31], especially after organ transplantation [34], wearing contact lenses [35], and exposure to contaminated environments [36]. Among HIV-positive patients, reported risk factors also include poor sanitation [33], living in a rural area [33], consumption of watermelon [33], use of injectable drugs [37], exposure to contaminated water in hot tubs and spas, and occupational contact with water [37]; some of these factors are still debated. More information is needed regarding risk factors of developing microsporidiosis in animals, but immunomodulation by cyclophosphamide [38] or dexamethasone [39] has been shown to be a risk factor in rabbits.
A recent study involving more than 200 patients investigated fecal detection of microsporidia in healthy humans and immunocompetent patients presenting with diarrhea: interestingly, the prevalence of microsporidia was higher in the healthy group with 45% of healthy patients shedding microsporidia [24]. This suggests that digestive microsporidia are often asymptomatic in humans [20]. Among HIV-positive patients, infection with microsporidia did not significantly shorten survival in a case-control study [33].
The zoonotic potential of microsporidia was first suggested in 1995 [40]. However, the clinical importance of this potential risk remains to be elucidated, as there is no formal proof of zoonotic transmission of microsporidia [27]. A zoonotic disease requires: (1) infection of an animal by the microorganism, with or without disease; (2) transmission of the microorganism from the animal to a human, and (3) infection of a human and development of a disease [41]. Numerous reports show that human-infecting microsporida can be detected in the feces of animals [42,43,44,45]. However, demonstrating infection of animals requires tissue histopathology and confirmation of a human-infecting strain of microsporidia in the tissue by molecular techniques; microsporidia can indeed travel through the digestive tract of animals without an actual host infection. In this case, animals do not amplify microsporidia. It would, therefore, be misleading to claim a zoonotic risk; in fact, the risk of human infection may be the same in the presence of animal as it is through environmental exposure. A nested cohort study conducted in HIV-positive patients in Peru showed a statistical association between infection by Enterocytozoon bieneusi genotype 1 and contact with rabbits, ducks, sheep and some domestic animals [33]. However, the same study also highlighted a statistical association between poor sanitation at home and microsporidiosis [33]. This could be an important confounding factor as the limited availability of running water and contact with farm animals are both more likely to occur in rural areas [33]. In addition, the authors of the papers acknowledged that E. bieneusi genotype 1 infection has not been reported in animals [33]. Another genotype of E. bieneusi, named Peru 16, has been isolated from a diarrheic child and seven subclinical guinea pigs of the same household [46]. Owing to the growing interest about zoonotic microsporidia and to their pathogenicity in veterinary species, veterinarians should be familiar with the challenges associated with microsporidial infection diagnosis and treatment in companion animals. This review summarizes the last advances in the diagnosis and treatment of companion exotic animal microsporidiosis.
2. Ornamental Fish Microsporidiosis
2.1. Agents and Disease in Fish
More than 14 genera of microsporidia infect freshwater and marine fish [8], teleosts, and elasmobranchs [47]. Cases in elasmobranchs are very uncommon [48]: microsporidial infection in the free-ranging common stingray (Dasyatis pastinaca) in Turkey has been reported to cause nodules around the disc [49], while a systemic and fatal infection has been reported in the captive leopard shark [48]. Reported microsporidian genera in fish include Dasyatispora, Glugea, Heterosporis, Ichthyosporidium, Kabatana, Loma, Nucleospora, Pleistophora, Spraguea [1], Tetramicra [45], and Pseudoloma genera, among others [12]. Most genera are specific to a fish species [8], while genera that are able to form xenomas, such as Glugea, can infect a broad range of fish species [50]. Zoonotic Pleistophora spp. have been suspected in fishermen [51].
Microsporidiosis has been reported in captive and wild fish [52]. In ornamental fish, microsporidiosis is particularly common in zebrafish (Danio rerio) infected by Pseudoloma neurophilia [53] and Pleistophora spp. [54] and in tetra infected with Pleistophora hyphessobryconis, which causes the disease known as “neon tetra disease” [12]. Infection by P. neurophila has been reported in 74% of the laboratory facilities examined through the Zebrafish International Resource Center pathology service in 2010 [55]. Most fish microsporidia have a preferential target site of infection, including striated muscles [49], coelomic organs, gills, the digestive tract, ovaries, and the central nervous system [55].
2.2. Diagnosis of Fish Microsporidiosis
For xenoma-forming species, such as Glugea spp., external masses can be detected macroscopically. Skin scrapes may facilitate detection of the typical spores in tetra, although xenomas are absent with Pleistophora spp. infections [21,22]. Spores can develop within three to four weeks after infection in some microsporidia species [56] and shedding may occur through the feces, urine and sex products during spawning depending on the infected organs [50]. In addition, spores may be released from lesions on the body surfaces ante-mortem or post-mortem [13,48]. Chitin-binding fluorochromes are the most sensitive stains to highlight fish microsporidial spores in histologic sections [22,23]. In vitro cell culture of fish microsporidia has been historically more difficult to achieve than for human- or insect-infecting microsporidia [57].
2.3. Treatment of Piscine Microsporidiosis
Disinfection of a fish tank or exhibit after an outbreak of microsporidiosis may be challenging [17]. While 5 ppm of chlorine, a typical concentration applied to tap water, is sufficient to inactivate Encephalitozoon cuniculi after 10 min, 1500 ppm of chlorine is needed at neutral pH to achieve more than 95% Glugea anomala spore inactivation, and higher pH decreases the efficacy of this disinfectant [17]. Ultraviolet sterilizers used in aquarium life support systems are an important means of preventing the spread of microsporidiosis in a facility through water sterilization [55,58]. In addition, surface disinfection techniques applied to fish eggs, such as iodine treatments in salmon and sodium hypochlorite bath treatments in zebrafish, have been unsuccessful to date [54]. This is expected because some microsporidia are thought to be transmitted vertically and can be present within the eggs [54].
Microsporidia may also be transmitted orally. Prevention of infection in piscivorous captive fish may require inactivating food-borne microsporidial spores. Inactivation of spores of Spraguea lophii in anglerfish can be achieved by freezing at −20 °C for 48 h, treatment at 60 °C for 15 min and microwaving at 750 watts for more than 60 s [59].
Treatment of infected fish has only been partially effective. Albendazole and fumagillin have been shown to decrease both the number of xenoma and the proportion of fish with xenomas in a school of rainbow trout (Oncorhynchus mykiss) infected by Loma salmonae, contrary to pyrimethamine, sulphaquinoxaline, amprolium and metronidazole [60], which have all been shown to be ineffective in treating active infections. However, fumagillin has also shown severe adverse effects in rainbow trout at efficacious dosages [61]. Fumagillin was initially used for the treatment of bee nosemosis [62]. It is a molecule produced by Aspergillus fumigatus [18]. Fumagillin is not approved for human or veterinary use and should not be used for food-animal species including fish [61]. In humans, fumagillin has been shown to cause reversible thrombocytopenia [18]. Quinine hydrochloride and decreasing the water temperature can delay the formation of xenomas in fish [63]. In addition, monensin has been shown to prevent infection by L. salmonae [64]. Although approved for some food-animal species [65], monensin is not approved for use in fish destined for human consumption in the United States [65], Europe [66] and Canada [67].
3. Rabbit and Rodents Encephalitozoonosis
3.1. Agent and Disease in Rabbits and Rodents
Encephalitozoon cuniculi is the most commonly reported microsporidia in non-human mammals [18]. To date, three strains of E. cuniculi have been officially described based on the number of repeated elements 5′-GTTT-3′ in the internal transcribed spacer (ITS) [7,40]. Strain I is typically found in rabbits, strain II has been described in rodents, and strain III has been reported in dogs and cotton-top tamarins [7,70,71]. Strains I and III have been shown to cause infection in humans [7,72], while strain II has also been detected in the stools of an HIV-infected patient [73]. In addition, novel strains have been reported in a human patient with a renal transplant [74], in cats [75], and in bearded dragons [76]. E. cuniculi is transmitted either vertically or horizontally in rabbits [7], including transmission via ingestion or inhalation of spores excreted in the urine of infected rabbits [77]. Spores can survive outside of the host for six weeks at 22 °C and less than a week at 4 °C [77]. Other microsporidia, such as E. bieneusi, have also been isolated from the feces of healthy rodents [46].
Most infections by E. cuniculi are subclinical [77] and some studies have reported seroprevalence ranging from 45% [78] to 89% in healthy rabbits [79]. T-cell immunity is thought to play a critical role in disease manifestation in rabbits and rodents, as in humans [80]. Alternatively, infection of target organs including the central nervous system, lungs and kidneys in rabbits and, less commonly, guinea pigs [77] can lead to cell rupture, inflammation, and clinical signs. Neurological signs are commonly reported and have been observed in some animals following experimental infection [81]. Interestingly, although vestibular disease is reported as a common neurological sign in rabbits infected with E. cuniculi [82], the lesions seem to affect the neocortex more often than the brainstem where the vestibular nuclei are located [83]. More precise histologic evaluation of the vestibular nuclei as well as systematic evaluation of the ear would be warranted to refine the post-mortem diagnosis of vestibular disease etiology, since the presence of E. cuniculi in the central nervous system may be incidental. Renal signs are also suspected but one study failed to demonstrate proteinuria in seropositive rabbits [84]. In mice, typical lesions include granulomatous hepatitis, interstitial nephritis, and meningoencephalitis. In immunocompromised mice, ascites is a common clinical sign of encephalitozoonosis [84]. Lens infection likely results from vertical transmission and can ultimately lead to uni- or bilateral phacoclastic uveitis [15,77,78,85,86]. Abortion and neonatal death have also been associated with E. cuniculi [77].
3.2. Diagnostic Challenge in Rabbits
Diagnosing encephalitozoonosis in rabbits is particularly challenging, due to a short excretion period and because neither histopathologic findings [87] nor serologic titers [81] correlate well with the severity of clinical signs. Since encephalitozoonosis is considered one of the most common causes of neurologic clinical signs in companion rabbits [82], veterinarians should be aware of these diagnostic challenges.
3.2.1. Antibody Detection, Electrophoresis and Acute Phase Proteins
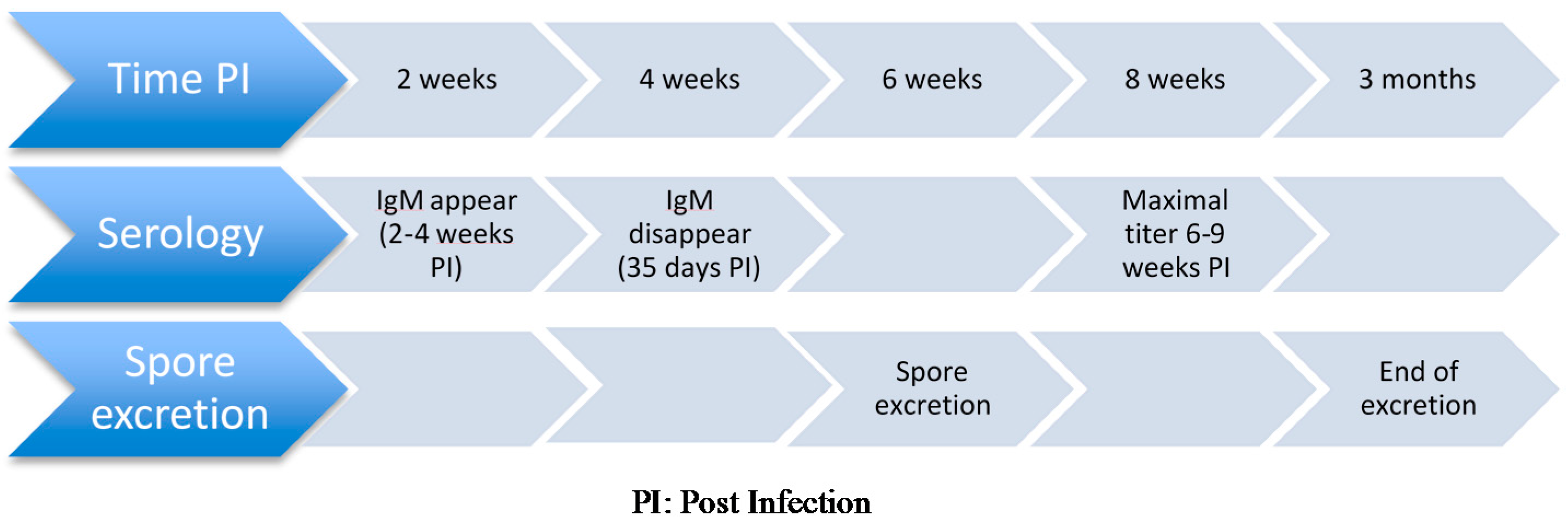
Available serologic tests for detection of antibodies against E. cuniculi in rabbits include carbon immunoassay [81,88], enzyme-linked immunoassay (ELISA) [80], and indirect immunofluorescent assay [89,90]. Antibodies become detectable two to four weeks after infection [81], which is up to two weeks before spores are detected in urine and tissues [91]. Maximal titers are detected six to nine weeks after infection and immunoglobulin M (IgM) decreases to 0 by 35 days post-infection [87]. Immunoglobulin G (IgG) becomes the dominant immunoglobulin around 17 days post-infection and can continue to be produced during chronic infection [87]. Sustained antibody titer has been reported to last more than seven years after experimental subcutaneous administration of inactivated spores [90]. Conversely, short antibody responses have also been documented [81]. Of note, offspring from seropositive dams may display detectable colostral antibodies for four weeks before titer becomes undetectable.
E. cuniculi-negative rabbit colonies have been established for research purposes, but many companion rabbits are seropositive to E. cuniculi [7], with a seroprevalence ranging from 29.5% to 73% depending on the study area and on the serologic assay used [78,87,92,93,94]. Seropositivity is very frequent in apparently healthy rabbits [95]. Some studies report significantly higher seroprevalence rates in symptomatic rabbit populations than in subclinical rabbits [78,93], whereas no statistically significant difference in seroprevalence between healthy and symptomatic rabbits was present in one study [95]. However, serologic titers correlated strongly with lesion intensity in the same study [95].
A panel including an ELISA serologic test associated with the acute phase proteins C-reactive protein is offered by the University of Miami laboratory [89,96,97]. This panel has been evaluated based on clinical suspicion of encephalitozoonosis in rabbits, rather than actual confirmation of infection [89,96,97]. Titers above the cut-off for IgM have been found in 24% of clinically normal rabbits [89]. It is expected that some rabbits previously infected by E. cuniculi could be seropositive and have concomitantly high C-reactive proteins due to their current disease and the reported specificity of 100% is therefore surprising in these cases [97]. A study has shown a 10-fold mean increase of the C-reactive protein concentration in rabbits suspected to be infected by E. cuniculi, based on the description of those clinical signs which were not explained by another causative agent [89]. Histologic diagnosis was available for only three rabbits suspected to be infected by E. cuniculi in this study [89]. Due to the study design, it is unknown whether the clinical signs of these rabbits were actually associated with E. cuniculi infection. Conversely, haptoglobin and serum amyloid A were not increased in the rabbit suspect group [89]. The significance of seropositivity at the individual level is controversial in rabbits. Experimental infection of rabbits has shown that antibody titer intensity is not correlated with recovery of the organism from brain tissue [81]. A study comparing companion rabbits suspected to be infected by E. cuniculi with non-suspect rabbits has shown that suspect rabbits had antibody titers 1.7 times higher than rabbits from the non-suspect group with a mean titer of 1:1324 in the suspect group [96]. However, only 79% of the rabbits included in the “suspect” group were seropositive, which could be due to a lack of seroconversion of some infected rabbits or to misclassification of some rabbits that were not, in fact, infected by E. cuniculi [96]. In addition, the inter-assay coefficient of variation of the serologic test was not provided, and this difference is usually below one dilution and could therefore represent inherent test variation. In another study evaluating the same serologic panel, the presence of concomitantly increased IgG, IgM, and C-reactive protein concentrations was reported to have a specificity of 100% in detecting encephalitozoonosis in rabbits [97]. This result, almost too perfect to be trusted, raises some concerns regarding the validity of the study design. It seems logical that this perfect specificity would be an illusion: for example, it is expected that many individuals from a population of naïve juvenile rabbits will display high IgM and IgG concentrations, since E. cuniculi is a common fungus in their environment. If they are exposed concomitantly to other pathogens causing neurologic signs in rabbit kits, such as Pasteurella multocida causing suppurative encephalitis, C-reactive protein will also increase. They will therefore be presented with the perfect picture of the E. cuniculi positive rabbit based upon the serologic panel, yet their true pathologic condition would be pasteurellosis and their therapeutic plan may suffer from this mistaken result. Practically, most veterinarians will formulate a basic treatment plan before receiving serologic test results because of the delay in result reporting. However, veterinarians should be aware of the imperfect specificity and limited sensitivity [97] of this serologic panel. Post-mortem confirmation of infection using PCR was not available to evaluate if inclusion criteria were valid in this study. Overall, more studies are needed to confirm the value of serologic and acute phase protein panels: these studies should include cases with a definitive diagnosis rather than strong anecdotal information, as acknowledged by the authors of the study about the serologic panel [97].
Serum and plasma protein electrophoresis has been suggested as an adjunctive diagnostic test for encephalitozoonosis [96]. Rabbits classified as E. cuniculi suspects and other sick rabbits had a significantly lower albumin-to-globulin ratio, total protein, and α2-globulin fractions with a higher γ-globulin fraction compared to clinically normal groups [96]. However, no specific electrophoretic changes enabled differentiation of E. cuniculi-suspect rabbits from other sick rabbits [96] and this test was therefore not clinically relevant to orient the diagnosis toward encephalitozoonosis in sick rabbits.
Detection of antibodies against E. cuniculi in rabbit urine has been investigated [98]. Immunoglobulin G and fragments of immunoglobulin G against the polar tube and spore wall can be measured separately [98]. In a colony of seropositive rabbits, only some seropositive individuals had detectable urinary immunoglobulins [98]. The authors interpreted this test as being possibly more specific than blood serology to detect renal disease associated with E. cuniculi; detection of urinary antibodies may indicate impaired glomerular filtration associated with multiplication of E. cuniculi in the kidney and resulting in excretion of antibodies [98]. However, this test requires further validation [98], especially since histologic lesions are more commonly tubular than glomerular with E. cuniculi [83].
Cerebrospinal fluid protein evaluation has been compared between 23 laboratory seronegative rabbits and 20 E. cuniculi-seropositive and Toxoplasma-negative companion rabbits with neurologic clinical signs including 12 with confirmed postmortem infection [99]. Protein concentration and cerebrospinal fluid leukocyte counts were significantly higher in infected rabbits than in healthy rabbits [99]. It would be interesting to evaluate the results of cerebrospinal cytologic evaluation in rabbits affected by other neurologic disease to compare them to those of E. cuniculi-infected rabbits. However, this procedure is rarely performed in companion rabbits due to its inherent risks, which limits the applications of this technique.
3.2.2. Antemortem Antigenic Tests
Antemortem antigenic tests include detection of E. cuniculi spores in the urine via real-time PCR. The main limitation of this technique is the short temporal period of spore excretion in rabbits, which limits the test sensitivity. Seroconversion precedes renal shedding and is still detectable afterwards [77] (Figure 1). Whether or not shedding increases in immunocompromised rabbits is debatable: some authors have not shown any effect of dexamethasone administration on spore excretions [39] while others suggest the contrary [100], as dexamethasone has been reported to increase spore excretion in mice [101,102]. Urinary spore excretion typically occurs between four weeks and 12 weeks post-infection [77], which may be earlier than the development of clinical signs.
In the case of phacoclastic uveitis, detection of E. cuniculi spores in the lenses of rabbits is also performed by conventional PCR or immunohistochemistry after enucleation or lens phacoemulsification [78,103]. In a study performed in companion rabbits suspected to be infected by E. cuniculi, a PCR test successfully detected E. cuniculi in the lenses of four out of five patients while 32 urine samples and 12 cerebrospinal fluid samples were negative [78].
Figure 1.
Sequence of events used for diagnosis of Encephalitozoon cuniculi in an immunocompetent companion rabbits.
Figure 1.
Sequence of events used for diagnosis of Encephalitozoon cuniculi in an immunocompetent companion rabbits.

3.2.3. PostMortem Diagnosis
E. cuniculi can also be challenging to diagnose postmortem [104]. The use of histopathology as the sole means of spore detection is at once insensitive and non-specific [83]. Histologic lesions may include nonsuppurative granulomatous encephalitis [15], spinal root inflammation, chronic lymphoplasmocytic to granulomatous interstitial nephritis [82], pneumonitis, myocarditis, hepatitis, splenitis, and uveitis [77]. Renal lesions have been detected as early as one month after experimental infection while cerebral lesions are thought to develop later on, after about two to three months [15,81]. In a study comparing patients’ neurologic signs to their postmortem lesions, no correlation was found between the severity of clinical signs and histopathologic lesions [87]. The Ziehl-Neelsen stain enabled detection of a higher number of spores than the trichrome stain, but the difference in sensitivity between the two staining techniques was not statistically evaluated [87]. Spores also stain with Goodpasture’s carbol fuchsin [77].
The proposed gold standard for diagnosing E. cuniculi infection is real-time PCR on tissue samples, which is currently the best technique of diagnosis. One study compared immunohistochemistry (IHC), real-time PCR, and histopathology of kidney, brain and lung tissue [83]. Histopathology was found to be less sensitive than immunohistochemistry, which was less sensitive than real-time PCR [83]. Brain and kidney were the most sensitive tissues for E. cuniculi detection. Within the brain, lesions were most commonly localized in the gray matter of the neocortex [83]. Within the kidney, acute lesions were mainly localized in the medulla while chronic lesions were mostly localized in the renal cortex [83]. In another study comparing the sensitivity of IHC and in-situ hybridization in infected rabbit tissue with electron microscopy, confirmation IHC was shown to be more sensitive than in-situ hybridization [104]. However IHC sensitivity was only 37% on kidney and brain samples and 60% on lens samples in this study, which included naturally infected companion rabbits [104].
3.3. Therapeutic Challenges
Due to the challenges associated with E. cuniculi diagnosis, especially antemortem, evaluating treatment efficacy is particularly difficult. Similar to diagnostic test evaluation, many authors have attempted studies in rabbits based on a presumptive infected status.
Proposed treatments for encephalitozoonosis in rabbits include benzimidazoles such as fenbendazole [105], oxibendazole, and albendazole [106]. Some fluoroquinolones have been suggested to have an inhibitory effect in vitro and in vivo in mice against Encephalitozoon intestinalis and some other microsporidia [107]. However, it should be noted that ciprofloxacin, a metabolite of enrofloxacin, did not have a significant effect when compared to norfloxacin and ofloxacin. The effect of fluoroquinolones on E. cuniculi has not been investigated to the authors’ knowledge and in vivo studies in rabbits using dosages previously established by pharmacokinetic studies [108] would be needed to investigate the effect of ofloxacin in this circumstance.
Fenbendazole has been shown to have a prophylactic and therapeutic effect against E. cuniculi in rabbits [105]. Fenbendazole metabolism has been investigated in rabbits [109,110]. Fenbendazole has been suggested as a superior treatment option compared to oxytetracycline in rabbits naturally infected by E. cuniculi based on survival rate and clinical improvement of their neurological score at 10 days [100]. However, postmortem diagnosis in this study was based exclusively on histopathology; therefore, some rabbits in this study could have been infected with other agents responsible for causing multifocal granulomatous encephalitis [100]. In addition, this study did not include a control group and the treatment attribution was neither randomized nor double-blinded for fenbendazole [100]. Although previously thought to be innocuous [105], albendazole, fenbendazole, and oxibendazole have all been reported to cause aplastic anemia and pancytopenia in rabbits [111]. In a case series of 13 suspected cases of benzimidazole intoxication, only one rabbit survived after receiving intensive supportive care including blood transfusions and erythropoietin administration [111]. Therefore, complete blood count rechecks or serial hematocrits are recommended as part of the follow up to benzimidazole treatment in rabbits. Albendazole should not be used in pregnant rabbits as it has been reported to be teratogenic and embryotoxic [112]. In addition, treatment with albendazole failed to clear microsporidia from experimentally infected rodents and further immunosuppression resulted in reactivation of E. cuniculi in an experimental model [113], showing the ineffectiveness of this approach [113].
Clinically, the use of corticosteroids in the treatment of encephalitozoonosis in rabbits seems contraindicated, as rabbits are particularly susceptible to the immunosuppressive effects of corticosteroids, which could amplify the parasitic infection [39]. In addition, a prospective study in a rabbit cohort did not show any impact of dexamethasone on clinical signs, as dexamethasone neither increased rabbit survival rate nor prolonged duration of survival [100]. Again, this study did not thoroughly confirm infection by E. cuniculi postmortem as no molecular testing was attempted on tissue and central nervous system lesions consistent with encephalitozoonosis were attributed to E. cuniculi even in seronegative rabbits [100]: hence, the treated group could have included rabbits with toxoplasmosis, among others. It is unknown whether the addition of dexamethasone can decrease albendazole elimination rate in rabbits as observed in humans [114,115]. Currently, dexamethasone is not recommended as an adjunctive treatment of encephalitozoonosis in rabbits.
Future therapeutic tools for use in rabbits may include drugs that have been tested in rodents and fish. New drugs under investigation include p38 mitogen-activated protein kinase (MAPK) inhibitors [116]. This class of drugs has been shown to prevent clinical signs in mice experimentally infected two days prior to administration of these drugs [116]. This class of drug is currently experimental and cannot be used clinically by veterinarians. Fumagillin-related compounds have been shown to inhibit E. cuniculi in vitro [106,117], but are not used currently in rabbits to the authors’ knowledge, although they have been suggested [112].
Many drugs have been cited for potential treatment of rabbit encephalitozoonosis but should not be used. Although used to treat microsporidiosis in fish and anecdotally cited in rabbits, toltrazuril and its metabolite ponazuril have not been tested in vivo in rabbits for the treatment of encephalitozoonosis [118]. In vitro, toltrazuril and ronidazole did not show any effect against E. cuniculi [106]. Sulfonamides and paromomycin have not shown in vitro efficacy against E. cuniculi [119], although they are occasionally cited as treatment options [100]. Fluoroquinolones have been suggested as a treatment for human microsporidiosis but applications in rabbits have not been investigated [107]. Tetracycline derivates have been shown to be only partially effective and clinical experience does not support their use in rabbits [100].
Regarding environmental disinfection, E. cuniculi has been shown to be susceptible to exposure for 10 minutes to common disinfectants [120,121], including bleach 0.1%–10%, quaternary ammonia, 70% alcohol [121], phenolic derivates, sodium hydroxide, chloramine, iodophores, hydrogen peroxide, and amphoteric surfactants [112].
Experimental vaccination of New Zealand White rabbits with inactivated Encephalitozoon spp. spores induced a seroconversion and antibodies were predicted to last for about seven years [90]. However, efficacy of this vaccine against microsporidial infection has not been tested as the main goal of the study was to develop a better source of anti-microsporidial serum for immunocompromised persons [90].
4. Avian, Reptile and Amphibian Microsporidiosis
4.1. Microsporidia in Amphibians
Microsporidia have been uncommonly reported in amphibians [122] both in anura [123,124,125] and urodela [126]. Pleistophora spp. has been associated with myositis in toads [123] and with kyphosis and epaxial myositis in San Marcos salamanders (Eurycea nana) [126]. Digestive xenomas have been described in Bufo marinus tadpoles infected with Alloglugea sp. [124]. Necrotizing hepatitis was described with unidentified microsporidia in tadpoles of the endangered mountain yellow-legged frogs (Rana muscosa) [127] and intestinal xenoma have been reported in subclinical cane toad tadpoles (Rhinella marina) [124]. In the cases of digestive lesions, typical spores may be detected on fecal examination [122]. Spores may also be visualized on blood smear in case of septicemia [115]. Diagnosis can be made via histology with visualization of Gram-positive spores in tissues, electron microscopy, and molecular techniques including PCR followed by sequencing [128]. Chloramphenicol sodium succinate and topical oxytetracycline hydrochloride, and polymyxin B sulfate have been suspected to inhibit microsporidial spore production in amphibians in a single case series; however, supportive care and other treatments were also administered in these American giant tree frogs and the report did not include a control group [125].
4.2. Microsporidia in Reptiles
Microsporidia are rarely reported in squamates and rhynchocephalia [8,16,76,128,129] but have not been reported in chelonians to the authors’ knowledge. Some microsporidia species have only been described in reptiles, such as Glugea danilewskyi [130], Pleistophora atretii [131] or Encephalitozoon lacertae [132], while others, such as Heterosporis anguillarum, are suspected to be transmissible between fish and reptiles [8]. A strain of E. cuniculi distinct from mammalian E. cuniculi has been described in bearded dragons (Pogona vitticeps) [76] but it is unknown whether this species was the same as those in earlier reports as molecular techniques were not available [129]. Similarly, reported Encephalitozoon sp. in African skinks (Mabuya perrotetii) [133] could be the same species as microsporidia reported in Podarcis muralis based on morphology [132]. Transmission of microsporidia in reptiles has been poorly studied and infection through ingestion of infected prey items and vertical transmission are suspected [13].
Systemic infections have been reported in bearded dragons with non-specific clinical signs including lethargy, anorexia, and severe polydipsia [76,129]. Granulomatous lesions were detected in the kidneys, liver, spleen, lungs, adrenal, gonads, fat bodies, and digestive tract mucosa [76]. Some microsporidia have also been found in endothelial cells of the central nervous system capillaries [129]. Unlike bearded dragons, a localized form of microsporidiosis characterized by myositis and thickening of the coelomic wall cranial to the cloaca have been reported in a gartersnake (Thamnophis sirtalis) [8] and in tuataras (Sphenodon punctatus) [128]. Interestingly, while microsporidia are thought to be intracellular fungi, some extracellular forms of Heterosporis anguillarum were noted in the gartersnake [8]. Microsporidia have been detected in the feces of 19 subclinical snakes and a subclinical lizard from a zoological collection, which died of other causes; this finding has led to the suspicion that microsporidia could be opportunistic pathogens in squamates [16]. Alternatively, the spores observed could originate from the reptiles’ food and represent a passage through the digestive tract rather than a true infection.
4.3. Microsporidia in Birds
4.3.1. Agents and Disease in Birds
Microsporidia carried by birds have been studied to evaluate zoonotic risks associated with wild [134,135] and captive birds [136,137,138,139]. When investigating feral urban pigeons and exotic birds seized from illegal trade in Brazil, shedding prevalence was found to be higher in pigeons, with 31% of pigeon stools being positive for microsporidia, although some birds from this study could have been sampled repeatedly [140]. Similarly, 29% of 124 individually sampled pigeons in Spain were found to shed microsporidia [141]. However, many studies have shown only excretion of microsporidial spores in bird stools, which could represent a simple transit through the digestive tract rather than an actual infection as demonstrated in naturally exposed budgerigars [142]. Hence, the detection of the most common microsporidia isolated from humans, E. bieneusi, in birds living in close contact with people, such as urban pigeons [141] and domestic chickens [143], may in fact reflect a transit rather than a true zoonotic risk. The distinction between mechanical vector and infected bird is especially important when very sensitive molecular screening tools such as PCR are being used [141].
Actual infection of birds associated with disease and pathologic changes has been confirmed in lovebirds infected by Encephalitozoon sp. [144,145,146,147], budgerigars [148], eclectus parrots [149], double-yellow headed Amazon parrots [150,151], an umbrella cockatoo [62], a yellow-streaked lory [152], an ostrich [25,153], and Anna’s hummingbirds [26] infected by Encephalitozoon hellem (Table 1). E. hellem is often implicated in human microsporidiosis but, so far, only genotype 1 has been found both in humans and birds, while genotypes 2, 3, and an unnamed genotype have been isolated in only one host group [18]. However, in cases reported before molecular diagnostic tests became available, the genus of microsporidia was assigned based exclusively upon electron microscopy and could be controversial [150]. Fatalities have been associated with microsporidial infections in birds [144,148]. Described clinical signs include diarrhea, hyporexia, respiratory distress [150], and keratoconjunctivitis [62,151]. Differential diagnoses for microsporidiosis should include psittacosis in Psittacidae. In gyrfalcon (Falco rusticolus), E. cuniculi genotype II has been suspected to cause clinical signs but causality was not demonstrated [154]. Many authors consider avian species to be a possible source of zoonotic microsporidiosis, either by acting as mechanical vectors or by replicating microsporidia [140,151,152,155]. However, mouse inoculation with microsporidia originating from birds has not resulted in infection [144] and no confirmed zoonotic cases have been reported to date.

{kind=link}
Table 1.
Avian species with reported microsporidial lesions and characterization technique used to determine the microsporidial agent.
| Avian Host Species | Host Latin Name | Microsporidia Species | Characterization Technique Used | Reference |
|---|---|---|---|---|
| Anna’s hummingbird | Calypte anna | Encephalitozoon hellem | PCR | [26] |
| Budgerigars | Melopsittacus undulatus | Encephalitozoon hellem and Encephalitozoon cuniculi | PCR | [142,148] |
| Double yellow-headed amazon | Amazona ochrocephala orathrix | Encephalitozoon sp. | TEM (morphologic) | [150] |
| Eclectus parrots | Eclectus roratus | Encephalitozoon hellem | PCR | [149] |
| Ostrich | Struthio camelus | Encephalitozoon hellem | PCR | [25] |
| Peach-faced lovebird | Agapornis roseicollis | Encephalitozoon sp. | TEM (morphologic) | [145,147] |
| Umbrella cockatoo | Cacatua alba | Encephalitozoon hellem | PCR | [62] |
| Yellow-streaked lori | Chalcopsitta scintillata | Encephalitozoon hellem | PCR | [152] |
4.3.2. Diagnosis of Avian Microsporidiosis
As for other species, diagnostic tests include immunofluorescence and classical stains [26,152] on tissues or body fluids including feces [152], electron microscopy [25,26,154], and molecular tests. In budgerigars, PCR has shown a higher sensitivity for detection in feces than microscopic examination [154]. In addition, the same study demonstrated that PCR was more sensitive than histology and tissue electron microscopy [154]. Fecal shedding has been shown to be intermittent and repeated testing may be required for detection [154]. In a case report, microsporidia were detected on fecal samples via Gram stain but not via PCR, although this was possibly due to intermittent shedding [151]. In this case report, a serologic test developed for rodents to screen for E. cuniculi yielded a positive result in an Amazon parrot [151]. No control was available and this test would need to be validated for use in avian species.
4.3.3. Treatment of Avian Microsporidiosis
An umbrella cockatoo has been treated with oral albendazole 25 mg/kg q24h for 90 days and topical neomycin, polymyxin, and bacitracin ointment for keratoconjunctivitis associated with microsporidia [62]. The clinical signs resolved, but three months after the end of the treatment, the contralateral eye developed keratoconjunctivitis. Albendazole treatment was resumed at the same dose for four months and the bird remained free of clinical signs for the following two years [62]. In another case of keratoconjunctivitis in a Psittacidae, oral albendazole at 50 mg/kg q24h for three months, oral itraconazole 10 mg/kg q24h, and fumagillin topical administration every 2 h during the day successfully resolved clinical signs, and post-treatment corneal swab and liver biopsy were negative via PCR and special stains [151]. No adverse effects from albendazole were reported in these two patients.
Of note, fenbendazole, albendazole and other benzimidazoles are toxic in some avian species [156] and can cause medullary aplasia, digestive signs, and death. Toxicity has been reported in some columbiformes [157,158], some vultures [159], storks [159,160], pink-backed pelicans (Pelecanus rufescens) [161], a leukemic great-horned owl (Bubo virginianus) [162], and, anecdotally, in cockatiels (Nymphicus hollandicus) [156] and keas (Nestor notabilis) [151]. Because columbiformes are suspected to excrete microsporidia more frequently than other birds [142], finding a viable treatment for this order would be important.
5. Conclusions
Microsporidiosis is a rare disease in most companion exotic animals. However, subclinical carriage and shedding have been reported in all vertebrate classes, with or without associated disease. Antemortem diagnosis is complicated by the polymorphism of clinical signs, including masses, myositis, neurologic signs, and any vague clinical signs from systemic disease, which encompasses a wide array of cases seen by veterinarians. Due to the suspected zoonotic potential of some of these fungi, especially in immunocompromised owners, and due to the close contact between companion animals and their owners, clinicians should keep microsporidiosis in differential diagnoses lists. More research would be beneficial to assess treatment efficacy in companion exotic animals.
Acknowledgments
The authors would like to thank Christine Parker for her help with the manuscript revision.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Campbell, S.E.; Williams, T.A.; Yousuf, A.; Soanes, D.M.; Paszkiewicz, K.H.; Williams, B.A. The genome of Spraguea lophii and the basis of host-microsporidian interactions. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Thomarat, F.; Vivarès, C.P.; Gouy, M. Phylogenetic analysis of the complete genome sequence of Encephalitozoon cuniculi supports the fungal origin of microsporidia and reveals a high frequency of fast-evolving genes. J. Mol. Evol. 2004, 59, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Gill, E.E.; Fast, N.M. Assessing the microsporidia-fungi relationship: Combined phylogenetic analysis of eight genes. Gene 2006, 375, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Heinz, E.; Williams, T.A.; Nakjang, S.; Noël, C.J.; Swan, D.C.; Goldberg, A.V.; Harris, S.R.; Weinmaier, T.; Markert, S.; Becher, D.; et al. The genome of the obligate intracellular parasite Trachipleistophora hominis: New insights into microsporidian genome dynamics and reductive evolution. PLoS Pathog. 2012, 8, e1002979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lucking, R.; et al. A higher-level phylogenetic classification of the fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Zhang, R.; de Koeyer, D.; Pan, G.; Li, T.; Liu, T.; Zhou, Z. New evidence on the relationship between microsporidia and fungi: A genome-wide analysis by darkhorse software. Can. J. Microbiol. 2014, 60, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Mathis, A.; Weber, R.; Deplazes, P. Zoonotic potential of the microsporidia. Clin. Microbiol. Rev. 2005, 18, 423–445. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.; Graner, I.; Csokai, J. Heterosporis anguillarum infection in a garter snake (Thamnophis sirtalis). J. Comp. Pathol. 2014, 150, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, L. Etudes sur la Maladie des vers a Soie, Moyen Pratique Assuré de la Combattre et d’en Prévenir le Retour; Gauthier-Villars: Paris, France, 1870; p. 434. (In France) [Google Scholar]
- Didier, E.S. Microsporidiosis: An emerging and opportunistic infection in humans and animals. Acta Trop. 2005, 94, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.C.; Nai, Y.S.; Wang, C.Y.; Solter, L.F.; Hsu, H.C.; Wang, C.H.; Lo, C.F. A new microsporidium, Triwangia caridinae gen. nov., sp. nov. Parasitizing fresh water shrimp, Caridina formosae (decapoda: Atyidae) in Taiwan. J. Invertebr. Pathol. 2013, 112, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Noga, E. Problem 70: Microsporidian infection. In Fish Disease Diagnosis and Treatment; Wiley-Blackwell: Ames, IA, USA, 2006; pp. 247–253. [Google Scholar]
- Jacobson, E.R. Chapter 12: Parasites and parasitic diseases of reptiles. In Infectious Diseases and Pathology of Reptiles; CRC Press: Boca Raton, FL, USA, 2007; pp. 571–665. [Google Scholar]
- Bruno, D.W.; Nowak, B.; Elliott, D.G. Guide to the identification of fish protozoan and metazoan parasites in stained tissue sections. Dis. Aquat. Organ. 2006, 70, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Künzel, F.; Joachim, A. Encephalitozoonosis in rabbits. Parasitol. Res. 2010, 106, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, T.K.; Cranfield, M.R. Cryptosporidium serpentis oocysts and microsporidian spores in feces of captive snakes. J. Parasitol. 2000, 86, 413–414. [Google Scholar] [CrossRef]
- Ferguson, J.A.; Watral, V.; Schwindt, A.R.; Kent, M.L. Spores of two fish microsporidia (Pseudoloma neurophilia and Glugea anomala) are highly resistant to chlorine. Dis. Aquat. Organ. 2007, 76, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Didier, E.S.; Didier, P.J.; Snowden, K.F.; Shadduck, J.A. Microsporidiosis in mammals. Microbes Infect. 2000, 2, 709–720. [Google Scholar] [CrossRef]
- Sanders, J.L.; Lawrence, C.; Nichols, D.K.; Brubaker, J.F.; Peterson, T.S.; Murray, K.N.; Kent, M.L. Pleistophora hyphessobryconis (microsporidia) infecting zebrafish Danio rerio in research facilities. Dis. Aquat. Organ. 2010, 91, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sak, B.; Kváč, M.; Kučerová, Z.; Květoňová, D.; Saková, K. Latent microsporidial infection in immunocompetent individuals—A longitudinal study. PLoS Negl. Trop. Dis. 2011, 5, e1162. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L. Laboratory identification of the microsporidia. J. Clin. Microbiol. 2002, 40, 1892–1901. [Google Scholar] [CrossRef] [PubMed]
- Luna, V.A.; Stewart, B.K.; Bergeron, D.L.; Clausen, C.R.; Plorde, J.J.; Fritsche, T.R. Use of the fluorochrome calcofluor white in the screening of stool specimens for spores of microsporidia. Am. J. Clin. Pathol. 1995, 103, 656–659. [Google Scholar] [PubMed]
- Peterson, T.S.; Spitsbergen, J.M.; Feist, S.W.; Kent, M.L. Luna stain, an improved selective stain for detection of microsporidian spores in histologic sections. Dis. Aquat. Organ. 2011, 95, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, I.; Cetin, F.; AL Dogruman, F.; Oguz, I.; Aksu, N. Prevalence of microsporidia in healthy individuals and immunocompetent patients with acute and chronic diarrhea. Infect. Dis. 2016, 48, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.; Logan, K. Molecular identification of Encephalitozoon hellem in an ostrich. Avian Dis. 1999, 43, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.; Daft, B.; Nordhausen, R.W. Morphological and molecular characterization of Encephalitozoon hellem in hummingbirds. Avian Pathol. 2001, 30, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.R.; Karkhanis, S.; Rabeneck, L.; Marino, B.; Foote, L.W. Quantitative light microscopic detection of Enterocytozoon bieneusi in stool specimens: A longitudinal study of human immunodeficiency virus-infected microsporidiosis patients. J. Clin. Microbiol. 1996, 34, 520–523. [Google Scholar] [PubMed]
- Anane, S.; Attouchi, H. Microsporidiosis: Epidemiology, clinical data and therapy. Gastroenterol. Clin. Biol. 2010, 34, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Didier, E.S.; Weiss, L.M. Microsporidiosis: Current status. Curr. Opin. Infect. Dis. 2006, 19, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, M.; Wan, Q.; Li, Y.; Lu, Y.; Jiang, Y.; Tao, W.; Li, W. Enterocytozoon bieneusi genotypes in children in Northeast China and assessment of risk of zoonotic transmission. J. Clin. Microbiol. 2014, 52, 4363–4367. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, U.; Khanduja, S.; Pant, P.; Prasad, K.N.; Dhole, T.N.; Sharma, R.K.; Ghoshal, U.C. Intestinal microsporidiosis in renal transplant recipients: Prevalence, predictors of occurrence and genetic characterization. Indian J. Med. Microbiol. 2015, 33, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Lobo, M.L.; Xiao, L.; Antunes, F.; Matos, O. Microsporidia as emerging pathogens and the implication for public health: A 10-year study on HIV-positive and -negative patients. Int. J. Parasitol. 2012, 42, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Kawai, V.; Vargas, D.; Rabke-Verani, J.; Williamson, J.; Chavez-Valdez, R.; Xiao, L.; Sulaiman, I.; Vivar, A.; Ticona, E.; et al. The epidemiology of intestinal microsporidiosis in patients with HIV/AIDS in Lima, Peru. J. Infect. Dis. 2005, 191, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Hocevar, S.N.; Paddock, C.D.; Spak, C.W.; Rosenblatt, R.; Diaz-Luna, H.; Castillo, I.; Luna, S.; Friedman, G.C.; Antony, S.; Stoddard, R.A.; et al. Microsporidiosis acquired through solid organ transplantation: A public health investigation. Ann. Intern. Med. 2014, 160, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Theng, J.; Chang, C.; Ling, M.L.; Tan, D. Microsporidial keratoconjunctivitis in a healthy contact lens wearer without human immunodeficiency virus infection. Ophthalmology 2001, 108, 976–978. [Google Scholar] [CrossRef]
- Matos, O.; Lobo, M.; Xiao, L. Epidemiology of Enterocytozoon bieneusi infection in humans. J. Parasitol. Res. 2012, 2012, 981424. [Google Scholar] [CrossRef] [PubMed]
- Dascomb, K.; Frazer, T.; Clark, R.A.; Kissinger, P.; Didier, E. Microsporidiosis and HIV. J. Acquir. Immune Defic. Syndr. 2000, 24, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Horváth, M.; Leng, L.; Stefkovic, M.; Révajová, V.; Halanová, M. Lethal encephalitozoonosis in cyclophosphamide-treated rabbits. Acta Vet. Hung. 1999, 47, 85–93. [Google Scholar] [PubMed]
- Jeklova, E.; Leva, L.; Kovarcik, K.; Matiasovic, J.; Kummer, V.; Maskova, J.; Skoric, M.; Faldyna, M. Experimental oral and ocular Encephalitozoon cuniculi infection in rabbits. Parasitology 2010, 137, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Didier, E.; Vossbrinck, C.R.; Baker, M.D.; Rogers, L.B.; Bertucci, D.C.; Shadduck, J.A. Identification and characterization of three Encephalitozoon cuniculi strains. Parasitology 1995, 111, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, P.; Conti, L. Human-animal Medicine: Clinical Approachto Zoonoses, Toxicants and Other Shared Health Risks; Saunders Elsevier: Maryland Heights, MO, USA, 2010. [Google Scholar]
- Słodkowicz-Kowalska, A. Animal reservoirs of human virulent microsporidian species. Wiad. Parazytol. 2009, 55, 63–65. [Google Scholar] [PubMed]
- Zhao, W.; Zhang, W.; Wang, R.; Liu, W.; Liu, A.; Yang, D.; Yang, F.; Karim, M.R.; Zhang, L. Enterocytozoon bieneusi in sika deer (Cervus nippon) and red deer (Cervus elaphus): Deer specificity and zoonotic potential of its genotypes. Parasitol. Res. 2014, 113, 4243–4250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, J.; Karim, M.R.; Zhao, J.; Dong, H.; Ai, W.; Li, F.; Zhang, L.; Wang, R. Zoonotic Enterocytozoon bieneusi genotypes in Pere David’s deer (Elaphurus davidianus) in Henan, C-China. Exp. Parasitol. 2015, 155, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.R.; Wang, R.; Dong, H.; Zhang, L.; Li, J.; Zhang, S.; Rume, F.I.; Qi, M.; Jian, F.; Sun, M.; et al. Genetic polymorphism and zoonotic potential of Enterocytozoon bieneusi from nonhuman primates in China. Appl. Environ. Microbiol. 2014, 80, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Cama, V.A.; Pearson, J.; Cabrera, L.; Pacheco, L.; Gilman, R.; Meyer, S.; Ortega, Y.; Xiao, L. Transmission of Enterocytozoon bieneusi between a child and guinea pigs. J. Clin. Microbiol. 2007, 45, 2708–2710. [Google Scholar] [CrossRef] [PubMed]
- Lom, J. A catalogue of described genera and species of microsporidians parasitic in fish. Syst. Parasitol. 2002, 53, 81–99. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.M. A retrospective study of disease in elasmobranchs. Vet. Pathol. 2013, 50, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Diamant, A.; Goren, M.; Yokes, M.B.; Galil, B.S.; Klopman, Y.; Huchon, D.; Szitenberg, A.; Karhan, S.U. Dasyatispora levantinae gen. et sp. nov., a new microsporidian parasite from the common stingray Dasyatis pastinaca in the eastern mediterranean. Dis. Aquat. Organ. 2010, 91, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Lom, J.; Dykova, I. Microsporidian xenomas in fish seen in wider perspective. Folia Parasitol. 2005, 52, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Weber, R.; Bryan, R.T.; Schwartz, D.A.; Owen, R.L. Human microsporidial infections. Clin. Microbiol. Rev. 1994, 7, 426–461. [Google Scholar] [PubMed]
- Post, G. Chapter VII: Animal parasites of fishes. In Textbook of Fish Health, 2nd ed.; Post, G., Ed.; T.F.H. Publications, Inc.: Neptune City, NJ, USA, 1987; pp. 172–174. [Google Scholar]
- Sanders, J.L.; Peterson, T.S.; Kent, M.L. Early development and tissue distribution of Pseudoloma neurophilia in the zebrafish, Danio rerio. J. Eukaryot. Microbiol. 2014, 61, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.L.; Watral, V.; Kent, M.L. Microsporidiosis in zebrafish research facilities. ILAR J. 2012, 53, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.; Dreska, M.; Nasiadka, A.; Rinne, M.; Matthews, J.L.; Carmichael, C.; Bauer, J.; Varga, Z.M.; Westerfield, M. Comparative medicine, transmission, diagnosis, and recommendations for control of Pseudoloma neurophilia infections in laboratory zebrafish (Danio rerio) facilities. Comp. Med. 2011, 61, 322–329. [Google Scholar] [PubMed]
- Kent, M.L.; Speare, D.J. Review of the sequential development of Loma salmonae (microsporidia) based on experimental infections of rainbow trout (Oncorhynchus mykiss) and chinook salmon (O. tshawytscha). Folia Parasitol. 2005, 52, 63–68. [Google Scholar] [CrossRef] [PubMed]
- McConnachie, S.; Sheppard, J.; Wright, G.M.; Speare, D.J. Development of the microsporidian parasite, Loma salmonae, in a rainbow trout gill epithelial cell line (RTG-1): Evidence of xenoma development in vitro. Parasitology 2015, 142, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Speare, D. Ultraviolet light control of horizontal transmission of Loma salmonae. J. Fish Dis. 2004, 27, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Leiro, J.M.; Piazzon, C.; Dominguez, B.; Mallo, N.; Lamas, J. Evaluation of some physical and chemical treatments for inactivating microsporidian spores isolated from fish. Int. J. Food Microbiol. 2012, 156, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Speare, D.J.; Athanassopoulou, F.; Daley, J.; Sanchez, J.G. A preliminary investigation of alternatives to fumagillin for the treatment of Loma salmonae infection in rainbow trout. J. Comp. Pathol. 1999, 121, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Lauren, D.J.; Wishkovsky, A.; Groff, J.M.; Hedrick, R.P.; Hinton, D.E. Toxicity and pharmacokinetics of the antibiotic fumagillin in yearling rainbow trout (Salmo gairdneri). Toxicol. Appl. Pharmacol. 1989, 98, 444–453. [Google Scholar] [CrossRef]
- Phalen, D.N.; Logan, K.S.; Snowden, K.F. Encephalitozoon hellem infection as the cause of a unilateral chronic keratoconjunctivitis in an umbrella cockatoo (Cacatua alba). Vet. Ophthalmol. 2006, 9, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Speare, D.J.; Ritter, G.; Schmidt, H. Quinine hydrochloride treatment delays xenoma formation and dissolution in rainbow trout challenged with Loma salmonae. J. Comp. Pathol. 1998, 119, 459–465. [Google Scholar] [CrossRef]
- Becker, J.A.; Speare, D.J.; Daley, J.; Dick, P. Effects of monensin dose and treatment time on xenoma reduction in microsporidial gill disease in rainbow trout, Oncorhynchus mykiss (walbaum). J. Fish Dis. 2002, 25, 673–680. [Google Scholar] [CrossRef]
- Food Animal Residue Avoidance Databank, Veterinarian’s Guide to Residue Avoidance Management. 2015. Available online: http://www.farad.org/ (accessed on 5 December 2015).
- Daniel, P. Available chemotherapy in mediterranean fish farming: Use and needs. In The Use of Veterinary Drugs and Vaccines in Mediterranean Aquaculture; Rogers, C., Basurco, B., Eds.; CIHEAM: Zaragoza, Spain, 2009; Volume 86, pp. 197–205. [Google Scholar]
- Health Canada. List of Veterinary Drugs That are Authorized for Sale by Health Canada for Use in Food-producing Aquatic Animals. 2010. Available online: http://www.hc-sc.gc.ca/dhp-mps/vet/legislation/pol/aquaculture_anim-eng.php (accessed on 5 July 2015).
- Rodríguez-Tovar, L.; Becker, J.A.; Markham, R.J.; Speare, D. Induction time for resistance to microsporidial gill disease caused by loma salmonae following vaccination of rainbow trout (Oncorhynchus mykiss) with a spore-based vaccine. Fish Shellfish Immunol. 2006, 21, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Speare, D.; Markham, R.; Guselle, N. Development of an effective whole-spore vaccine to protect against microsporidial gill disease in rainbow trout (Oncorhynchus mykiss) by using a low-virulence strain of Loma salmonae. Clin. Vaccine Immunol. 2007, 14, 1652–1654. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.; Logan, K.; Didier, E.S. Encephalitozoon cuniculi strain III is a cause of encephalitozoonosis in both humans and dogs. J. Infect. Dis. 1999, 180, 2086–2088. [Google Scholar] [CrossRef] [PubMed]
- Reetz, J.; Wiedemann, M.; Aue, A.; Wittstatt, U.; Ochs, A.; Thomschke, A.; Manke, H.; Schwebs, M.; Rinder, H. Disseminated lethal Encephalitozoon cuniculi (genotype III) infections in cotton-top tamarins (Oedipomidas oedipus)-a case report. Parasitol. Int. 2004, 53, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Didier, E.S.; Visvesvara, G.S.; Baker, M.D.; Rogers, L.B.; Bertucci, D.C.; de Groote, M.A.; Vossbrinck, C.R. A microsporidian isolated from an aids patient corresponds to Encephalitozoon cuniculi III, originally isolated from domestic dogs. J. Clin. Microbiol. 1996, 34, 2835–2837. [Google Scholar] [PubMed]
- Sokolova, O.I.; Demyanov, A.V.; Bowers, L.C.; Didier, E.S.; Yakovlev, A.V.; Skarlato, S.O.; Sokolova, Y.Y. Emerging microsporidian infections in russian HIV-infected patients. J. Clin. Microbiol. 2011, 49, 2102–2108. [Google Scholar] [CrossRef] [PubMed]
- Talabani, H.; Sarfati, C.; Pillebout, E.; van Gool, T.; Derouin, F.; Menotti, J. Disseminated infection with a new genovar of Encephalitozoon cuniculi in a renal transplant recipient. J. Clin. Microbiol. 2010, 48, 2651–2653. [Google Scholar] [CrossRef] [PubMed]
- Benz, P.; Maass, G.; Csokai, J.; Fuchs-Baumgartinger, A.; Schwendenwein, I.; Tichy, A.; Nell, B. Detection of Encephalitozoon cuniculi in the feline cataractous lens. Vet. Ophthalmol. 2011, 14, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.; Csokai, J.; Graner, I.; Eisenberg, T.; Pantchev, N.; Eskens, H.U.; Nedorost, N. Encephalitozoonosis in two inland bearded dragons (Pogona vitticeps). J. Comp. Pathol. 2013, 148, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Percy, D.; Barthold, S. Chapter 6: Rabbit, parasitic diseases. In Pathology of Laboratory Rodents and Rabbits, 3rd ed.; Percy, D., Barthold, S., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 287–294. [Google Scholar]
- Künzel, F.; Gruber, A.; Tichy, A.; Edelhofer, R.; Nell, B.; Hassan, J.; Leschnik, M.; Thalhammer, J.G.; Joachim, A. Clinical symptoms and diagnosis of encephalitozoonosis in pet rabbits. Vet. Parasitol. 2008, 151, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Berger Baldotto, S.; Cray, C.; Turner Giannico, A.; Reifer, L.; Montiani-Ferreira, F. Seroprevalence of Encephalitozoon cuniculi infection in pet rabbits in Brazil. J. Exot. Pet Med. 2015, 24, 435–440. [Google Scholar] [CrossRef]
- Anete Lallo, M.; Porta Miche Hirschfeld, M. Encephalitozoonosis in pharmacologically immunosuppressed mice. Exp. Parasitol. 2012, 131, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Kunstyr, I.; Lev, L.; Naumann, S. Humoral antibody response of rabbits to experimental infection with Encephalitozoon cuniculi. Vet. Parasitol. 1986, 21, 223–232. [Google Scholar] [CrossRef]
- Grüber, A.; Pakozdy, A.; Weissenböck, H.; Csokai, J.; Künzel, F. A retrospective study of neurological disease in 118 rabbits. J. Comp. Pathol. 2009, 140, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Leipig, M.; Matiasek, K.; Rinder, H.; Janik, D.; Emrich, D.; Baiker, K.; Hermanns, W. Value of histopathology, immunohistochemistry, and real-time polymerase chain reaction in the confirmatory diagnosis of Encephalitozoon cuniculi infection in rabbits. J. Vet. Diagn. Investig. 2012, 25, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Reusch, B.; Murray, J.K.; Papasouliotis, K.; Redrobe, S.P. Urinary protein:creatinine ratio in rabbits in relation to their serological status to Encephalitozoon cuniculi. Vet. Rec. 2009, 164, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Percy, D.; Barthold, S. Chapter 1: Mouse. In Pathology of Laboratory Rodents and Rabbits, 3rd ed.; Percy, D., Barthold, S., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 3–124. [Google Scholar]
- Jeklova, E.; Jekl, V.; Kovarcik, K.; Hauptman, K.; Koudela, B.; Neumayerova, H.; Knotek, Z.; Faldyna, M. Usefulness of detection of specific IgM and IgG antibodies for diagnosis of clinical encephalitozoonosis in pet rabbits. Vet. Parasitol. 2010, 170, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Csokai, J.; Gruber, A.; Kunzel, F.; Tichy, A.; Joachim, A. Encephalitozoonosis in pet rabbits (Oryctolagus cuniculus): Pathohistological findings in animals with latent infection versus clinical manifestation. Parasitol. Res. 2009, 104, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Waller, T.; Morein, B.; Fabiansson, E. Humoral immune response to infection with Encephalitozoon cuniculi in rabbits. Lab. Anim. 1978, 12, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; Rodriguez, M.; Fernandez, Y. Acute phase protein levels in rabbits with suspected Encephalitozoon cuniculi infection. J. Exot. Pet Med. 2013, 22, 280–286. [Google Scholar] [CrossRef]
- Sobottka, I.; Iglauer, F.; Schuler, T.; Schmetz, C.; Visvesvara, G.S.; Albrecht, H.; Schwartz, D.A.; Pieniazek, N.J.; Bartscht, K.; Laufs, R.; et al. Acute and long-term humoral immunity following active immunization of rabbits with inactivated spores of various Encephalitozoon species. Parasitol. Res. 2001, 87, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.C.; Gallichio, H.A. Serological and histological studies on adult rabbits with recent, naturally acquired encephalitozoonosis. Res. Vet. Sci. 1978, 24, 260–261. [Google Scholar] [PubMed]
- Lyngset, A. A survey of serum antibodies to Encephalitozoon cuniculi in breeding rabbits and their young. Lab. Anim. Sci. 1980, 30, 558–561. [Google Scholar] [PubMed]
- Harcourt-Brown, F.M.; Holloway, H.K. Encephalitozoon cuniculi in pet rabbits. Vet. Rec. 2003, 152, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Csokai, J.; Joachim, A.; Gruber, A.; Tichy, A.; Pakozdy, A.; Kunzel, F. Diagnostic markers for encephalitozoonosis in pet rabbits. Vet. Parasitol. 2009, 163, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Oohashi, E.; Dautu, G.; Ueno, A.; Kariya, T.; Furuya, K. High seroprevalence of Encephalitozoon cuniculi in pet rabbits in Japan. J. Vet. Med. Sci. 2008, 70, 1301–1304. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; Arcia, G.; Schneider, R.; Kelleher, S.A.; Arheart, K.L. Evaluation of the usefulness of an ELISA and protein electrophoresis in the diagnosis of Encephalitozoon cuniculi infection in rabbits. Am. J. Vet. Res. 2009, 70, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; McKenny, S.; Perritt, E.; Arheart, K.L. Utility of IgM titers with IgG and C-reactive protein quantification in the diagnosis of suspected Encephalitozoon cuniculi infection in rabbits. J. Exot. Pet Med. 2015, 24, 356–360. [Google Scholar] [CrossRef]
- Furuya, K.; Asakura, T.; Igarashi, M.; Morita, T. Microsporidian Encephalitozoon cuniculi antibodies in rabbit urine samples. Vet. Rec. 2009, 165, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Jass, A.; Matiasek, K.; Henke, J.; Küchenhoff, H.; Hartmann, K.; Fischer, A. Analysis of cerebrospinal fluid in healthy rabbits and rabbits with clinically suspected encephalitozoonosis. Vet. Rec. 2008, 162, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Sieg, J.; Hein, J.; Jass, A.; Sauter-Louis, C.; Hartmann, K.; Fischer, A. Clinical evaluation of therapeutic success in rabbits with suspected encephalitozoonosis. Vet. Parasitol. 2012, 187, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Lallo, M.A.; Dos Santos, M.J.; Bondan, E.F. Experiental Encephalitozoon cuniculi infection in dexamethasone-immunosuppressed mice. Rev. Saude Publica 2006, 36, 621–626. [Google Scholar]
- Herich, R.; Levkutova, M.; Kokincakova, T.; Reiterova, K.; Hipikova, V.; Levkut, M. Diagnosis and manifestation of encephalitozoonosis in mice after experimental infection with different species and application of dexamethasone. J. Vet. Med. Ser. A 2009, 53, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Felchle, L.M.; Sigler, R.L. Phacoemulsification for the management of Encephalitozoon cuniculi-induced phacoclastic uveitis in a rabbit. Vet. Ophthalmol. 2002, 5, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Habenbacher, B.; Klang, A.; Fragner, K.; Dinhopl, N.; Kunzel, F.; Weissenbock, H. Comparative evaluation of specific methods for labeling of Encephalitozoon cuniculi in paraffin wax-embedded tissue samples. J. Vet. Diagn. Investig. 2012, 24, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Suter, C.; Müller-Doblies, U.U.; Hatt, J.M.; Deplazes, P. Prevention and treatment of Encephalitozoon cuniculi infection in rabbits with fenbendazole. Vet. Rec. 2001, 148, 478–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franssen, F.F.; Lumeij, J.T.; van Knapen, F. Susceptibility of Encephalitozoon cuniculi to several drugs in vitro. Antimicrob. Agents Chemother. 1995, 39, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Didier, E.S.; Bowers, L.; Stovall, M.E.; Kuebler, D.; Mittleider, D.; Brindley, P.J.; Didier, P.J. Antimicrosporidial activity of (fluoro)quinolones in vitro and in vivo. Folia Parasitol. 2005, 52, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Marangos, M.N.; Zhu, Z.; Nicolau, D.P.; Klepser, M.E.; Nightingale, C.H. Disposition of ofloxacin in female New Zealand white rabbits. J. Vet. Pharmacol. Ther. 1997, 20, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Short, C.R.; Barker, S.A.; Hsieh, L.C.; Ou, S.P.; McDowell, T. Disposition of fenbendazole in the rabbit. Res. Vet. Sci. 1988, 44, 215–219. [Google Scholar] [PubMed]
- Short, C.R.; Flory, W.; Hsieh, L.C.; Barker, S.A. The oxidative metabolism of fenbendazole: A comparative study. J. Vet. Pharmacol. Ther. 1988, 11, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.; Garner, M.; Reavill, D. Benzimidazole toxicosis in rabbits: 13 cases (2003 to 2011). J. Exot. Pet Med. 2014, 23, 188–195. [Google Scholar] [CrossRef]
- Fischer, P.; Carpenter, J.W. Chapter 19: Neurologic and musculoskeletal diseases. In Ferrets, Rabbits and Rodents Medicine and Surgery, 3rd ed.; Elsevier: St. Louis, MO, USA, 2013; pp. 245–256. [Google Scholar]
- Kotkova, M.; Sak, B.; Kvetonova, D.; Kvac, M. Latent microsporidiosis caused by Encephalitozoon cuniculi in immunocompetent hosts: A murine model demonstrating the ineffectiveness of the immune system and treatment with albendazole. PLoS ONE 2013, 8, e60941. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Hurtado, M.; Medina, M.T.; Sanchez, M.; Sotelo, J. Dexamethasone increases plasma levels of albendazole. J. Neurol. 1990, 237, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Takayanagui, O.M.; Lanchote, V.L.; Marques, M.P.; Bonato, P.S. Therapy for neurocysticercosis: Pharmacokinetic interaction of albendazole sulfoxide with dexamethasone. Ther. Drug Monit. 1997, 19, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Daniel, B.J.; Brumlik, M.J.; Burow, M.E.; Zou, W.; Khan, I.A.; Wadsworth, S.; Siekierka, J.; Curiel, T.J. Drugs designed to inhibit human p38 mitogen-activated protein kinase activation treat Toxoplasma gondii and Encephalitozoon cuniculi infection. Antimicrob. Agents Chemother. 2007, 51, 4324–4328. [Google Scholar] [CrossRef] [PubMed]
- Shadduck, J.A. Effect of fumagillin on in vitro multiplication of Encephalitozoon cuniculi. J. Protozool. 1980, 27, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M. Therapeutic review: Ponazuril. J. Exot. Pet Med. 2008, 17, 228–229. [Google Scholar] [CrossRef]
- Beauvais, B.; Sarfati, C.; Challier, S.; Derouin, F. In vitro model to assess effect of antimicrobial agents on Encephalitozoon cuniculi. Antimicrob. Agents Chemother. 1994, 38, 2440–2448. [Google Scholar] [CrossRef] [PubMed]
- Waller, T. Sensitivity of Encephalitozoon cuniculi to various temperatures, disinfectants and drugs. Lab. Anim. 1979, 13, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.N.; Dicristina, J.A.; Lindsay, D.S. Activity of bleach, ethanol and two commercial disinfectants against spores of Encephalitozoon cuniculi. Vet. Parasitol. 2006, 136, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Poynton, S.; Whitaker, B.R. Protozoa and metazoa infecting amphibians. In Amphibian Medicine and Captive Husbandry; Wright, G.M., Whitaker, B.R., Eds.; Krieger Publishing Co: Malabar, FL, USA, 2001; pp. 401–495. [Google Scholar]
- Canning, E.U.; Elkan, E.; Trigg, P.I. Plistophora myotrophica spec. nov., causing high mortality in the common toad Bufo bufo L., with notes on the maintenance of Bufo and Xenopus in the laboratory. J. Protozool. 1964, 11, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Paperna, I.; Lainson, R. Alloglugea bufonis nov. gen., nov. sp. (microsporidia: Glugeidae), a microsporidian of Bufo marinus tadpoles and metamorphosing toads (amphibia: Anura) from Amazon Brazil. Dis. Aquat. Organ. 1995, 23, 7–16. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Cranfield, M.R.; Bicknese, E.J.; Wisnieski, A.P. Progressive ulcerative dermatitis in captive, wild-caught, south-american giant tree frog (Phyllomedusa bicolor) with microsporidial septicemia. J. Zoo Wildl. Med. 1996, 27, 522–527. [Google Scholar]
- Gamble, K.; Garner, M.M.; West, E.S.; West, G.; Didier, E.S.; Cali, A.; Alvarado, T.P. Kyphosis associated with microsporidial myositis in San Marcos salamanders. J. Herpetol. Med. Surg. 2005, 15, 14–17. [Google Scholar]
- Pessier, A.P. Infectious diseases of amphibians: It isn’t just redleg anymore. In Current Therapy in Reptile Medicine and Surgery; Divers, S., Mader, D.R., Eds.; Elsevier: St. Louis, MO, USA, 2014; pp. 247–254. [Google Scholar]
- Liu, S.K.; King, F.W. Microsporidiosis in the tuatara. J. Am. Vet. Med. Assoc. 1971, 159, 1578–1582. [Google Scholar] [PubMed]
- Jacobson, E.R.; Green, D.E.; Undeen, A.H.; Cranfield, M.; Vaughn, K.L. Systemic microsporidiosis in inland bearded dragons (Pogona vitticeps). J. Zoo Wildl. Med. 1998, 29, 315–323. [Google Scholar] [PubMed]
- Guyénot, E.; Naville, A. Recherches sur le parasitisme et l’evolution d’une microsporidie Glugea danilewskyi l. Pfr. Revue Suisse Zool. 1922, 30, 1–61. [Google Scholar]
- Narasimhamurti, C.; Kalavati, C.; Ahamed, S. A new microsporidian, Plistophora atretii sp. n. From the freshwater snake Atretium schistosum Gunther. Acta Protozool. 1982, 21, 127–137. [Google Scholar]
- Canning, E.U. Encephalitozoon lacertae n. sp., a microsporidian parasite of the lizard Podarcis muralis. In Society of Protozoologists Special Publication, No 1; Canning, E.U., Ed.; Society of Protozoologists: Utica, NY, USA, 1981; pp. 57–64. [Google Scholar]
- Koudela, B.; Didier, E.S.; Rogers, L.B.; Modry, D.; Kucerova, S. Intestinal microsporidiosis in african skink Mabuya perrotetii. Folia Parasitol. 1998, 45, 149–155. [Google Scholar] [PubMed]
- Malcekova, B.; Valencakova, A.; Molnar, L.; Kocisova, A. First detection and genotyping of human-associated microsporidia in wild waterfowl of Slovakia. Acta Parasitol. 2013, 58, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Pirestani, M.; Sadraei, J.; Forouzandeh, M. Molecular characterization and genotyping of human related microsporidia in free-ranging and captive pigeons of Tehran, Iran. Infect. Genet. Evol. 2013, 20, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Kasickova, D.; Sak, B.; Kvac, M.; Ditrich, O. Detection of Encephalitozoon cuniculi in a new host—Cockateel (Nymphicus hollandicus) using molecular methods. Parasitol. Res. 2007, 101, 1685–1688. [Google Scholar] [CrossRef] [PubMed]
- Kasickova, D.; Sak, B.; Kvac, M.; Ditrich, O. Sources of potentially infectious human microsporidia: Molecular characterisation of microsporidia isolates from exotic birds in the Czech republic, prevalence study and importance of birds in epidemiology of the human microsporidial infections. Vet. Parasitol. 2009, 165, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lee, S.S.; Lyoo, Y.S.; Park, H.M. DNA detection and genotypic identification of potentially human-pathogenic microsporidia from asymptomatic pet parrots in South Korea as a risk factor for zoonotic emergence. Appl. Environ. Microbiol. 2011, 77, 8442–8444. [Google Scholar] [CrossRef] [PubMed]
- Slodkowicz-Kowalska, A.; Graczyk, T.K.; Nowosad, A.; Majewska, A.C. First detection of microsporidia in raised pigeons in poland. Ann. Agric. Environ. Med. 2013, 20, 13–15. [Google Scholar] [PubMed]
- Lallo, M.A.; Calabria, P.; Milanelo, L. Encephalitozoon and Enterocytozoon (microsporidia) spores in stool from pigeons and exotic birds: Microsporidia spores in birds. Vet. Parasitol. 2012, 190, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Haro, M.; Izquierdo, F.; Henriques-Gil, N.; Andres, I.; Alonso, F.; Fenoy, S.; del Aguila, C. First detection and genotyping of human-associated microsporidia in pigeons from urban parks. Appl. Environ. Microbiol. 2005, 71, 3153–3157. [Google Scholar] [CrossRef] [PubMed]
- Sak, B.; Kasickova, D.; Kvac, M.; Kvetonova, D.; Ditrich, O. Microsporidia in exotic birds: Intermittent spore excretion of Encephalitozoon spp. In naturally infected budgerigars (Melopsittacus undulatus). Vet. Parasitol. 2010, 168, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Reetz, J.; Rinder, H.; Thomschke, A.; Manke, H.; Schwebs, M.; Bruderek, A. First detection of the microsporidium Enterocytozoon bieneusi in non-mammalian hosts (chickens). Int. J. Parasitol. 2002, 32, 785–787. [Google Scholar] [CrossRef]
- Kemp, R.L.; Kluge, J.P. Encephalitozoon sp. in the blue-masked lovebird, Agapornis personata (Reichenow): First confirmed report of microsporidan infection in birds. J. Protozool. 1975, 22, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Novilla, M.N. Microsporidian infection in the pied peach-faced lovebird (Agapornis roseicollis). Avian Dis. 1978, 22, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.J.; Lees, S.; Higgins, R.J.; Harcourt-Brown, N.H. Microsporidian infection in lovebirds (Agapornis spp.). Avian Pathol. 1986, 15, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.H.; Prior, H.C. Microsporidiosis in a peach-faced lovebird (Agapornis roseicollis). Aust. Vet. J. 1994, 71, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Black, S.S.; Steinohrt, L.A.; Bertucci, D.C.; Rogers, L.B.; Didier, E.S. Encephalitozoon hellem in budgerigars (Melopsittacus undulatus). Vet. Pathol. 1997, 34, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Pulparampil, N.; Graham, D.; Phalen, D.; Snowden, K. Encephalitozoon hellem in two eclectus parrots (Eclectus roratus): Identification from archival tissues. J. Eukaryot. Microbiol. 1998, 45, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Poonacha, K.B.; William, P.D.; Stamper, R.D. Encephalitozoonosis in a parrot. J. Am. Vet. Med. Assoc. 1985, 186, 700–702. [Google Scholar] [PubMed]
- Canny, C.; Ward, D.; Patton, S. Microsporidian keratoconjunctivitis in a double yellow-headed amazon parrot (Amazona ochrocephala orathrix). J. Avian Med. Surg. 1999, 13, 279–286. [Google Scholar]
- Suter, C.; Mathis, A.; Hoop, R.; Deplazes, P. Encephalitozoon hellem infection in a yellow-streaked lory (Chalcopsitta scintillata) imported from indonesia. Vet. Rec. 1998, 143, 694–695. [Google Scholar] [PubMed]
- Gray, M.L.; Puette, M.; Latimer, K.S. Microsporidiosis in a young ostrich (Struthio camelus). Avian Dis. 1998, 42, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Malcekova, B.; Valencakova, A.; Luptakova, L.; Molnar, L.; Ravaszova, P.; Novotny, F. First detection and genotyping of Encephalitozoon cuniculi in a new host species, gyrfalcon (Falco rusticolus). Parasitol. Res. 2011, 108, 1479–1482. [Google Scholar] [CrossRef] [PubMed]
- Slodkowicz-Kowalska, A.; Graczyk, T.K.; Tamang, L.; Jedrzejewski, S.; Nowosad, A.; Zduniak, P.; Solarczyk, P.; Girouard, A.S.; Majewska, A.C. Microsporidian species known to infect humans are present in aquatic birds: Implications for transmission via water? Appl. Environ. Microbiol. 2006, 72, 4540–4544. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, T.; Yeager, J. Pet bird toxicity and related environmental concerns. Vet. Clin. N. Am. 2008, 11, 229–259. [Google Scholar] [CrossRef] [PubMed]
- Howard, L.L.; Papendick, R.; Stalis, I.H.; Allen, J.L.; Sutherland-Smith, M.; Zuba, J.R.; Ward, D.L.; Rideout, B.A. Fenbendazole and albendazole toxicity in pigeons and doves. J. Avian Med. Surg. 2002, 16, 203–210. [Google Scholar] [CrossRef]
- Gozalo, A.S.; Schwiebert, R.S.; Lawson, G.W. Mortality associated with fenbendazole administration in pigeons (Columba livia). J. Am. Assoc. Lab. Anim. Sci. 2006, 45, 63–66. [Google Scholar] [PubMed]
- Bonar, C.; Lewandowski, A.; Schaul, J. Suspected fenbendazole toxicosis in 2 vulture species (Gyps africanus, Torgos tracheliotus) and marabou storks (Leptoptilos crumeniferus). J. Avian Med. Surg. 2003, 17, 16–19. [Google Scholar] [CrossRef]
- Weber, M.A.; Terrell, S.P.; Neiffer, D.L.; Miller, M.A.; Mangold, B.J. Bone marrow hypoplasia and intestinal crypt cell necrosis associated with fenbendazole administration in five painted storks. J. Am. Vet. Med. Assoc. 2002, 221, 417–419, 369. [Google Scholar] [CrossRef] [PubMed]
- Weber, M. Chapter 13: Pelicaniformes. In Zoo and Wildlife Medicine, 5th ed.; Fowler, M.E., Miller, R., Eds.; Sauders: St. Louis, MO, USA, 2003; pp. 118–122. [Google Scholar]
- Wiley, J.L.; Whittington, J.K.; Wilmes, C.M.; Messick, J.B. Chronic myelogenous leukemia in a great horned owl (Bubo virginianus). J. Avian Med. Surg. 2009, 23, 36–43. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vergneau-Grosset, C.; Larrat, S. Microsporidiosis in Vertebrate Companion Exotic Animals. J. Fungi 2016, 2, 3. https://doi.org/10.3390/jof2010003
AMA Style
Vergneau-Grosset C, Larrat S. Microsporidiosis in Vertebrate Companion Exotic Animals. Journal of Fungi. 2016; 2(1):3. https://doi.org/10.3390/jof2010003
Chicago/Turabian StyleVergneau-Grosset, Claire, and Sylvain Larrat. 2016. "Microsporidiosis in Vertebrate Companion Exotic Animals" Journal of Fungi 2, no. 1: 3. https://doi.org/10.3390/jof2010003
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.