
Genome Studies on Nematophagous and Entomogenous Fungi in China

Abstract
:1. Introduction
2. Nematophagous Fungi


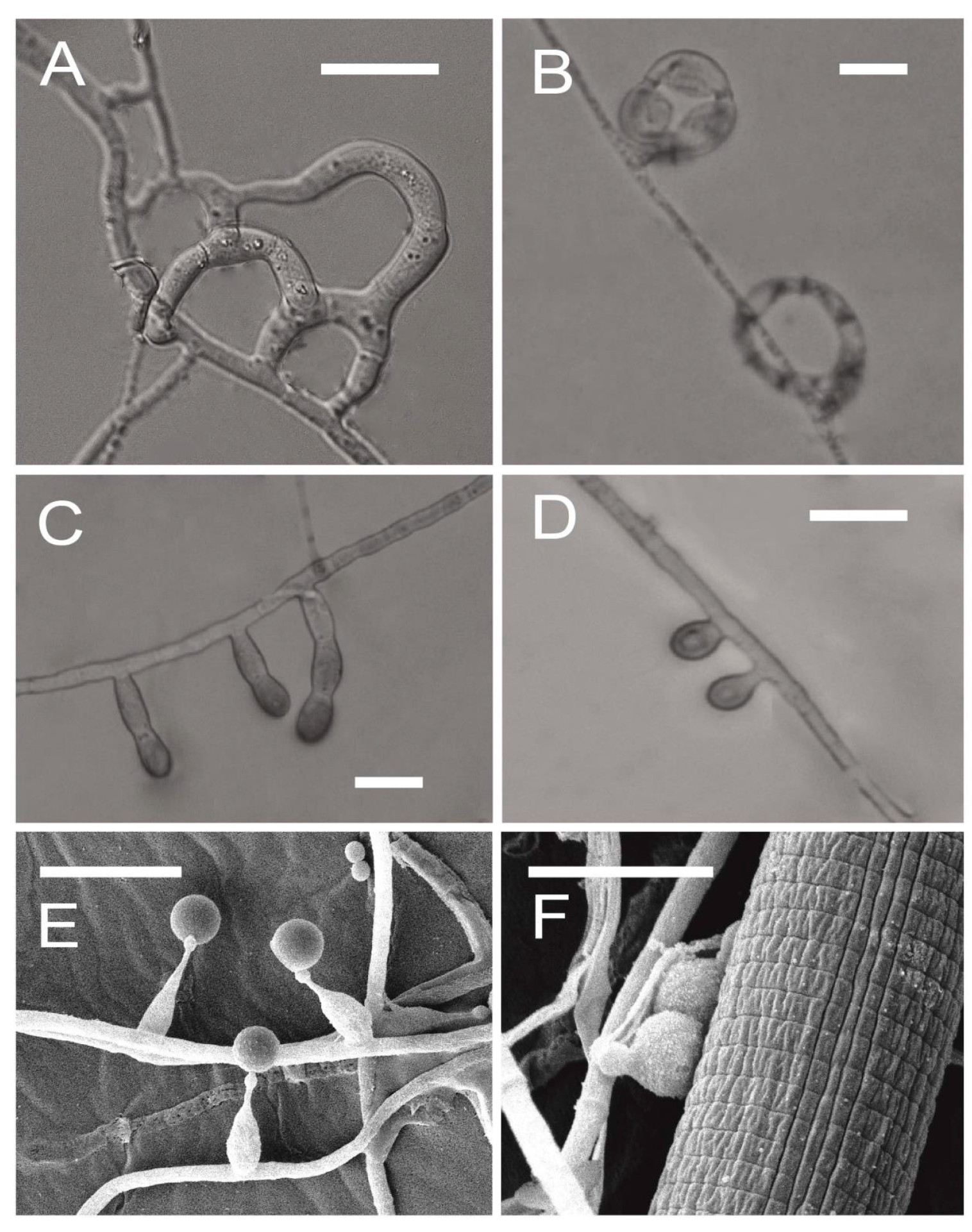{kind=link}
{kind=link}
| Features | Nematode-Trapping Fungi | Nematode Endoparasitic Fungi | Entomogenous Fungi | |||||
|---|---|---|---|---|---|---|---|---|
| Drechslerella stenobrocha | Arthrobotrys oligospora | Hirsutella minnesotensis | Metarhizium robertsii | Metarhizium acridum | Beauveria bassiana | Cordyceps militaris | Ophiocordyceps sinensis | |
| Assembled size (Mb) | 29.02 | 40.02 | 51.4 | 39.04 | 38.05 | 33.7 | 32.2 | ~120 |
| Protein-coding genes | 7781 | 11,479 | 12,702 | 10,582 | 9849 | 10,366 | 9684 | 6792 |
| Coverage (fold) | 80× | - | 128 | 100 | 107 | 76.6 | 147 | 241 |
| Number of scaffolds | 142 | - | 967 | 174 | 241 | 242 | 147 | - |
| Scaffold N50 (kb) | 434.4 | - | 382.4 | 1960 | 330 | 730 | 4550 | - |
| G + C content (%) | 52.5 | 45.2 | 52.1 | 51.49 | 49.91 | 51.5 | 51.4 | 46.1 |
| Simple repeat rate (%) | 0.92 | - | 1.33 | 0.98 | 1.52 | 2.03 | 3.04 | 37.98 |
| TEs (%) | - | - | 34.67 | - | - | - | - | - |
| Gene density (genes per Mb) | 268 | 271 | 247.1 | 271.1 | 258.8 | 308 | 257 | 87 |
| Exons per gene | 3.5 | 3.3 | 2.5 | 2.8 | 2.7 | 2.7 | 3.0 | 2.6 |
| tRNA genes | 82 | 149 | 145 | 141 | 122 | 113 | 136 | - |
| References | Liu et al. 2014 [10] | Yang et al. 2011 [9] | Lai et al. 2014 [13] | Gao et al. 2011 [18] | Gao et al. 2011 [18] | Xiao et al. 2012 [17] | Zheng et al. 2011 [12] | Hu et al. 2013 [15] |
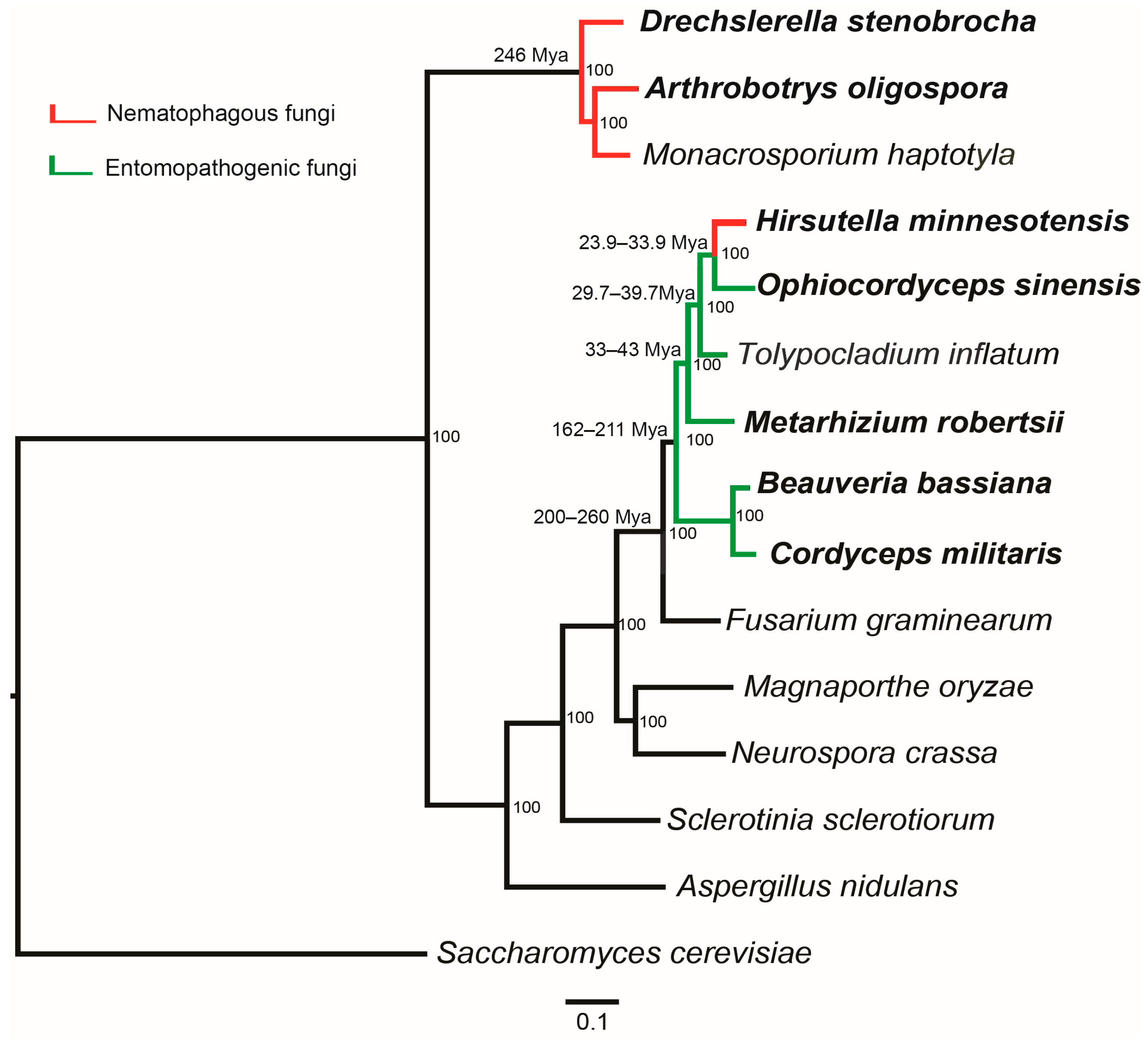
2.1. Different Origin of Nematode-Trapping and Endoparasitic Fungi
2.2. Lectins and Other Adhesive Proteins Involved in Adhering to Nematode

2.3. Formation of Traps and Infection Pegs
2.4. Virulence Factors Associated with Nematode Infection
2.4.1. Extracellular Enzymes
2.4.2. Secondary Metabolites
3. Entomopathogenic Fungi
3.1. Divergence and Origin of Insect Pathogens
3.2. Host Recognition and Signal Transduction
3.3. Secreted Enzymes Involved in Penetration of Insect Cuticles
3.4. Secondary Metabolites
3.5. Mechanisms of Fungal Pathogen Speciation and Host Adaptation
4. Conclusions
- Nematophagous and entomogenous fungi include many important species that impact ecology and human life. Origination and differentiation of some species such as O. sinensis or H. minnesotensis have been studied based on multigene analysis [83,84]. Availability of de novo genome sequences of these important species has made it possible for population genomics analysis that could provide a more comprehensive understanding of their origination, differentiation, and speciation.
- Phylogeny based on single gene or multigenes has been extensively applied in the systematics and evolution of fungi. Origination and evolution of trapping devices of nematode-trapping fungi have been deduced to result from the mass extinct events based on the multigene analysis [25,30]. Since a number of fungal genomes have been sequenced, phylogenomics has become a new strategy for the systematic and evolutionary study.
- Many nematophagous and entomopathogenic fungi have been developed/are being developed as biocontrol agents. The molecular mechanisms of fungi interacting with nematodes and insects are essential for utilization of these fungi in agricultural pest management. “-Omics” should be more efficient for finding and identifying the functional genes and pathways involved in the interaction between fungi and host pests that will help in the development of biocontrol agents.
- Secondary metabolites are not only involved in the interaction between fungi and their host pests, but also the precursors to develop nematicides, insecticides and drugs. Destruxins were identified in 1990s while their biosynthetic puzzle was not solved until the genomes of Metarhizium spp. were sequenced [75,76]. “-Omics” have provided an efficient strategy to decipher and identify gene clusters encoding secondary metabolites.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barron, G.L. The Nematode-Destroying Fungi; Canadian Biological Publications Ltd.: Guelph, ON, Canada, 1977. [Google Scholar]
- Liu, X.; Xiang, M.; Che, Y. The living strategy of nematophagous fungi. Mycoscience 2009, 50, 20–25. [Google Scholar] [CrossRef]
- Roberts, D.W.; St Leger, R.J. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar] [PubMed]
- Dong, C.; Guo, S.; Wang, W.; Liu, X. Cordyceps industry in China. Mycology 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. Biocontrol 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Cobb, N.A. The monochs (Mononchus Bastian, 1866) a genus of free living predatory nematodes. Soil Sci. 1917, 3, 431–486. [Google Scholar] [CrossRef]
- Krassilstschik, I.M. La production industrielle des parasites vegetaux pour la destruction des insects nuisibles. Bull. Sci. Fr. Belg. 1988, 19, 461–472. (in French). [Google Scholar]
- Liu, X.Z.; Li, S. Fungal secondary metabolites in biological control of crop pests. In Handbook of Industrial Mycology; An, Z., Ed.; Marcel Dekker Inc.: New York, NY, USA, 2004; pp. 723–747. [Google Scholar]
- Yang, J.; Wang, L.; Ji, X.; Feng, Y.; Li, X.; Zou, C.; Xu, J.; Ren, Y.; Mi, Q.; Wu, J.; et al. Genomic and proteomic analyses of the fungus Arthrobotrys oligospora provide insights into nematode-trap formation. PLoS Pathog. 2011, 7, e1002179. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, W.; Lai, Y.; Xiang, M.; Wang, X.; Zhang, X.; Xiang, M.; Liu, X. Drechslerella stenobrocha genome illustrates the mechanism of constricting rings and the origin of nematode predation in fungi. BMC Genom. 2014, 15, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Pramer, D. Nematode-trapping fungi. Science 1964, 144, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, F.; Liu, X. Hirsutella minnesotensis sp nov., a new pathogen of the soybean cyst nematode. Mycologia 2000, 92, 819–824. [Google Scholar] [CrossRef]
- Lai, Y.; Liu, K.; Zhang, X.; Zhang, X.; Li, K.; Wang, N.; Shu, C.; Wu, Y.; Wang, C.; Bushly, K.E.; et al. Comparative genomics and transcriptomics analyses reveal divergent lifestyle features of nematode endoparasitic fungus Hirsutella minnesotensis. Genome Biol. Evol. 2014, 6, 3077–3093. [Google Scholar] [CrossRef] [PubMed]
- Borneman, J.; Becker, J.O. Identifying microorganisms involved in specific pathogen suppression in soil. Annu. Rev. Phytopathol. 2007, 45, 153–172. [Google Scholar] [PubMed]
- Hu, X.; Zhang, Y.J.; Xiao, G.H.; Peng, Z.; Xia, Y.L.; Zhang, X.Y.; St Leger, R.J.; Liu, X.; Wang, C. Genome survey uncovers the secrets of sex and lifestyle in caterpillar fungus. Chin. Sci. Bull. 2013, 58, 2846–2854. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional chinese medicine. Genome Biol. 2011, 12, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; St Leger, R.J.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jin, K.; Ying, S.H.; Zhang, Y.; Xiao, G.; Shang, Y.; Duan, Z.; Hu, X.; Xie, X.; Zhou, G.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; St Leger, R.J. Genomics of entomopathogenic fungi. In The Ecological Genomics of Fungi; Martin, F., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 243–260. [Google Scholar]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Nowrousian, M.; Stajich, J.E.; Chu, M.; Engh, I.; Espagne, E.; Halliday, K.; Kamerewerd, J.; Kempken, F.; Knab, B.; Kuo, H.C.; et al. De novo assembly of a 40 Mb eukaryotic genome from short sequence reads: Sordaria macrospora, a model organism for fungal morphogenesis. PLoS Genet. 2010, 6, e1000891. [Google Scholar] [CrossRef] [PubMed]
- Ahrén, D.; Tholander, M.; Fekete, C.; Rajashekar, B.; Friman, E.; Johansson, T.; Tunlid, A. Comparison of gene expression in trap cells and vegetative hyphae of the nematophagous fungus Monacrosporium haptotylum. Microbiology 2005, 151, 789–803. [Google Scholar]
- Nordbring-Hertz, B.; Jansson, H.B.; Tunlid, A. Nematophagous fungi. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd: Alicante, Spain, 2006. [Google Scholar]
- Yang, Y.; Yang, E.; An, Z.; Liu, X. Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B. Nematode-induced morphogenesis in the predacious fungus Arthrobotrys oligospora. Nematologica 1977, 23, 443–451. [Google Scholar] [CrossRef]
- Liu, K.; Tian, J.; Xiang, M.; Liu, X. How carnivorous fungi use three-celled constricting rings to trap nematodes. Protein Cell 2012, 3, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Zhang, K. Arthrobotrys oligospora: A model organism for understanding the interaction between fungi and nematodes. Mycology 2011, 2, 59–78. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Zehr, E.I. Parasitism of the Nematode Criconemella xenoplax by the Fungus Hirsutella rhossiliensis. Phytopathology 1982, 72, 1378–1381. [Google Scholar] [CrossRef]
- Yang, E.; Xu, L.; Yang, Y.; Zhang, X.; Xiang, M.; Wang, C.; An, Z.; Liu, X. Origin and evolution of carnivorism in the Ascomycota (fungi). Proc. Natl Acad Sci USA 2012, 109, 10960–10965. [Google Scholar] [CrossRef] [PubMed]
- Barron, G.L. Predatory fungi, wood decay, and the carbon cycle. Biodivers 2003, 4, 3–9. [Google Scholar] [CrossRef]
- Jin, Y.G.; Wang, Y.; Wang, W.; Shang, Q.H.; Cao, C.Q.; Erwin, D.H. Pattern of marine mass extinction near the Permian-Triassic boundary in South China. Science 2000, 289, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Espagne, E.; Lespinet, O.; Malagnac, F.; Silva, C.D.; Jaillon, O.; Porcel, B.M.; Couloux, A.; Aury, J.; Ségurens, B.; Poulain, J.; et al. The genome sequence of the model ascomycete fungus Podospora anserina. Genome Biol. 2008, 9, R77. [Google Scholar] [CrossRef] [PubMed]
- Floudas, D.; Hibbett, D.S. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordbring-Hertz, B.; Mattiasson, B. Action of a nematode-trapping fungus shows lectin-mediated host–microorganism interaction. Nature 1979, 5731, 477–479. [Google Scholar] [CrossRef]
- Rosenzweig, W.D.; Ackroyd, D. Binding characteristics of lectins involved in the trapping of nematodes by fungi. Appl. Environ. Microbiol. 1983, 46, 1093–1096. [Google Scholar] [PubMed]
- Balogh, J.; Tunlid, A.; Rosén, S. Deletion of a lectin gene does not affect the phenotype of the nematode-trapping fungus Arthrobotrys oligospora. Fungal Genet. Biol. 2003, 39, 128–135. [Google Scholar] [CrossRef]
- Wang, B.; Wu, W.; Liu, X. Purification and characterization of a neutral serine protease with nematicidal activity from Hirsutella rhossiliensis. Mycopathologia 2007, 163, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, X.; Wu, W.; Liu, X.; Li, S. Purification, characterization, and gene cloning of an alkaline serine protease from a highly virulent strain of the nematode-endoparasitic fungus Hirsutella rhossiliensis. Microbiol. Res. 2009, 164, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Åhman, J.; Johansson, T.; Olsson, M.; Punt, P.J.; van den Hondel, J.; Tunlid, A. Improving the pathogenicity of a nematode-trapping fungus by genetic engineering of a subtilisin with nematotoxic activity. Appl. Environ. Microbiol. 2002, 68, 3408–3415. [Google Scholar]
- Lopez-Llorca, L.V.; Maciá-Vicente, J.G.; Jansson, H.B. Mode of action and interactions of nematophagous fungi. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Springer Netherlands: Dordrecht, The Netherlands, 2008; pp. 51–76. [Google Scholar]
- Anke, H.; Stadler, M.A.; Sterner, O. Linoleic acid—The nematicidal principle of several nematophagous fungi and its production in trap-forming submerged cultures. Arch. Microbiol. 1993, 160, 401–405. [Google Scholar]
- Stadler, M.; Sterner, O.; Anke, H. New biologically active compounds from the nematode-trappmg fungus Arthrobotrys oligospora fresen. Z. Naturforschung 2014, 48, 843–850. [Google Scholar]
- Lai, Y. Molecular mechanism of Hirsutella spp. parasitizing on soybean cyst nematodes & a novel high-throughput nematicidal assay using embryo cells and larvae of Caenorhabditis elegans. In preparation.
- Roberts, D.W.; Humber, R.A. Entomogenous fungi. In Biology of Conidial Fungi; Academic Press: New York, NY, USA, 1981; pp. 201–236. [Google Scholar]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Sung, G.H.; Ban, S.; Nakagiri, A.; Chen, M.J.; Huang, B.; Li, Z.; Spatafora, J.W. New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 2012, 104, 182–197. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; St Leger, R.J. The Metarhizium anisopliae perilipin homolog MPL1 regulates lipid metabolism, appressorial turgor pressure, and virulence. J. Biol. Chem. 2007, 282, 21110–21115. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; St Leger, R.J. The MAD1 adhesin of Metarhizium anisopliae links adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryot. Cell 2007, 6, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Pava-Ripoll, M.; Wang, S.; St Leger, R.J. Protein kinase a regulates production of virulence determinants by the entomopathogenic fungus, Metarhizium anisopliae. Fungal Genet. Biol. 2009, 46, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Jiang, X.; Wang, G.; Luo, Z.; Fan, Y.; Wu, Z.; Pei, Y. Requirement of a mitogen-activated protein kinase for appressorium formation and penetration of insect cuticle by the entomopathogenic fungus Beauveria bassiana. Appl. Microbiol. Biotechnol. 2010, 76, 2262–2270. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Borovsky, D.; Hawking, C.; Ortiz-Urquiza, A.; Keyhani, N. Exploiting host molecules to augment mycoinsecticide virulence. Nat. Biotechnol. 2012, 30, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Keyhani, N.O.; Yu, X.; He, Z.; Luo, Z. The MAP kinase Bbslt2 controls growth, conidiation, cell wall integrity, and virulence in the insect pathogenic fungus Beauveria bassiana. Fungal Genet. Biol. 2012, 49, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Kittakoop, P.; Kirtikara, K.; Hywel-Jones, N.L.; Thebtaranonth, Y. Bioactive substances from insect pathogenic fungi. Acc. Chem. Res. 2005, 38, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.L.; Noronha, E.F.; Miller, R.N.G.; Franco, O.L. Novel insights in the use of hydrolytic enzymes secreted by fungi with biotechnological potential. Lett. Appl. Microbiol. 2007, 44, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.O.B.; Santi, L.; Berger, M.; Pinto, A.F.M.; Guimarães, J.A.; Schrank, A. Characterization of a spore surface lipase from the biocontrol agent Metarhizium anisopliae. Process Biochem. 2009, 44, 829–834. [Google Scholar] [CrossRef]
- Kim, H.G.; Song, H.; Yoon, D.H.; Song, B.W.; Park, S.M. Cordyceps pruinosa extracts induce apoptosis of HeLa cells by a caspase dependent pathway. J. Ethnopharmacol. 2010, 128, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.; Poinar, G.; Spatafora, J. The oldest fossil evidence of animal parasitism by fungi supports a cretaceous diversification of fungal-arthropod symbioses. Mol. Phylogenet. Evol. 2008, 49, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, C.A.; Güldener, U.; Xu, J.; Trail, F.; Turgeon, B.G.; Pietro, A.D.; Walton, J.D.; Ma, L.; Baker, S.E.; Rep, M.; et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 2008, 317, 1400–1402. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.; Sung, G.; Sung, J.; Hywel-Jones, N.; White, J. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; St Leger, R.J. Field studies using a recombinant mycoinsecticide (Metarhizium anisopliae) reveal that it is rhizosphere competent. Appl. Environ. Microbiol. 2002, 68, 6383–6387. [Google Scholar] [CrossRef] [PubMed]
- Milner, R.J.; Lim, R.P.; Hunter, D.M. Risks to the aquatic ecosystem from the application of Metarhizium anisopliae for locust control in Australia. Pest Manag. Sci. 2002, 58, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; St Leger, R.J. Mrt, a gene unique to fungi, encodes an oligosaccharide transporter and facilitates rhizosphere competency in Metarhizium robertsii. Plant Physiol. 2010, 154, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequencing and analysis of the biomassdegrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.D.; Liu, X.Z. Occurrence and diversity of insect-associated fungi in natural soils in china. Appl. Soil Ecol. 2008, 39, 100–108. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Ericsson, T.D. Environmental and behavioral constraints on the infection of wireworms by Metarhizium anisopliae. Environ. Entomol. 2007, 36, 1415–1420. [Google Scholar] [CrossRef]
- Kulkarni, R.D.; Thon, M.R.; Pan, H.; Dean, R.A. Novel G-protein-coupled receptor-like proteins in the plant pathogenic fungus Magnaporthe grisea. Genome Biol. 2005, 6, R24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, K.; Won, M.; Lee, S.; Jang, Y.; Hoe, K.; Kim, D.; Lee, J.; Kim, K.; Yoo, H. Isolation of a novel gene from schizosaccharomyces pombe: Stm1+ encoding a seven-transmembrane loop protein that may couple with the heterotrimeric Gα 2 protein, Gpa2. J. Biol. Chem. 2001, 276, 40190–40201. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.B.; Read, A.F. Can fungal biopesticides control malaria? Nat. Rev. Microbiol. 2007, 5, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Fang, W.; Guo, S.; Pei, X.; Zhang, Y.; Xiao, Y.; Li, D.; Jin, K.; Bidochka, M.J.; Pei, Y. Increased insect virulence in Beauveria bassiana strains overexpressing an engineered chitinase. Appl. Environ. Microbiol. 2007, 73, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Park, S.J.; Lee, S.G.; Shin, S.C.; Choi, D.H. Cordycepin: Selective growth inhibitor derived from liquid culture of Cordyceps militaris against Clostridium spp. J. Agric. Food Chem. 2000, 48, 2744–2748. [Google Scholar] [CrossRef] [PubMed]
- Vining, L.C.; Kelleher, W.J.; Schwarting, A.E. Oosporein production by a strain of Beauveria bassiana originally identified as amanita muscaria. Can. J. Microbiol. 1962, 8, 931–933. [Google Scholar] [CrossRef]
- Wat, C.T.; Mcinnes, A.G.; Smith, D.G.; Wright, J.; Vining, C.L. Yellow pigments of Beauveria species. Structures of tenellin and bassianin. Can. J. Chem. 1977, 55, 4090–4098. [Google Scholar] [CrossRef]
- Kagamizono, T.; Nishino, E.; Matsumoto, K.; Kawashima, A.; Kishimoto, M.; Sakai, N.; He, B.M.; Chen, Z.X.; Adachi, T.; Morimoto, S.; et al. Bassiatin, a new platelet aggregation inhibitor produced by Beauveria bassiana K-717. J. Antibiot. 1995, 48, 1407–1412. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Chen, T.H.; Liu, B.L.; Wu, L.C.; Chen, Y.C.; Tzeng, Y.M.; Hsu, S.L. Destruxin B isolated from entomopathogenic fungus Metarhizium anisopliae induces apoptosis via a Bcl-2 family-dependent mitochondrial pathway in human nonsmall cell lung cancer cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 548929. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, M.J.; Moorhouse, E.R.; Bateman, R.; Reynolds, S.E.; Charnley, A.K. The role of destruxins in the pathogenicity of Metarhizium anisopliae for three species of Insect. J. Invertebr. Pathol. 1999, 74, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the biosynthetic puzzle of destruxins in Metarhizium species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xiao, G.; Zheng, P.; Shang, Y.; Su, Y.; Zhang, X.; Liu, X.; Zhan, S.; St Leger, R.J.; Wang, C. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [PubMed]
- Driver, F.; Milner, R.J.; Trueman, J.W.H. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycol. Res. 2000, 104, 134–150. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Kamp, A.M.; Lavender, T.M.; Dekoning, J.; Croos, J.N.D. Habitat association in two genetic groups of the insect-pathogenic fungus Metarhizium anisopliae: Uncovering cryptic species? Appl. Environ. Microbiol. 2001, 67, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Jiang, X.; Cheng, X.; Wang, N.; Chen, S.; Xiang, M.; Liu, X. Genetic structure and parasitization-related ability divergence of a nematode fungal pathogen Hirsutella minnesotensis following founder effect in China. Fungal Genet. Biol. 2015, 97, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Li, Y.; Ma, S.; Wang, C.; Xiang, M. Phylogeography and evolution of a fungal-insect association on the Tibetan Plateau. Mol. Ecol. 2014, 23, 5337–5355. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Cheng, X.; Liu, X.; Xiang, M. Genome Studies on Nematophagous and Entomogenous Fungi in China. J. Fungi 2016, 2, 9. https://doi.org/10.3390/jof2010009
Zhang W, Cheng X, Liu X, Xiang M. Genome Studies on Nematophagous and Entomogenous Fungi in China. Journal of Fungi. 2016; 2(1):9. https://doi.org/10.3390/jof2010009
Chicago/Turabian StyleZhang, Weiwei, Xiaoli Cheng, Xingzhong Liu, and Meichun Xiang. 2016. "Genome Studies on Nematophagous and Entomogenous Fungi in China" Journal of Fungi 2, no. 1: 9. https://doi.org/10.3390/jof2010009