
The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas

Abstract
:1. Introduction
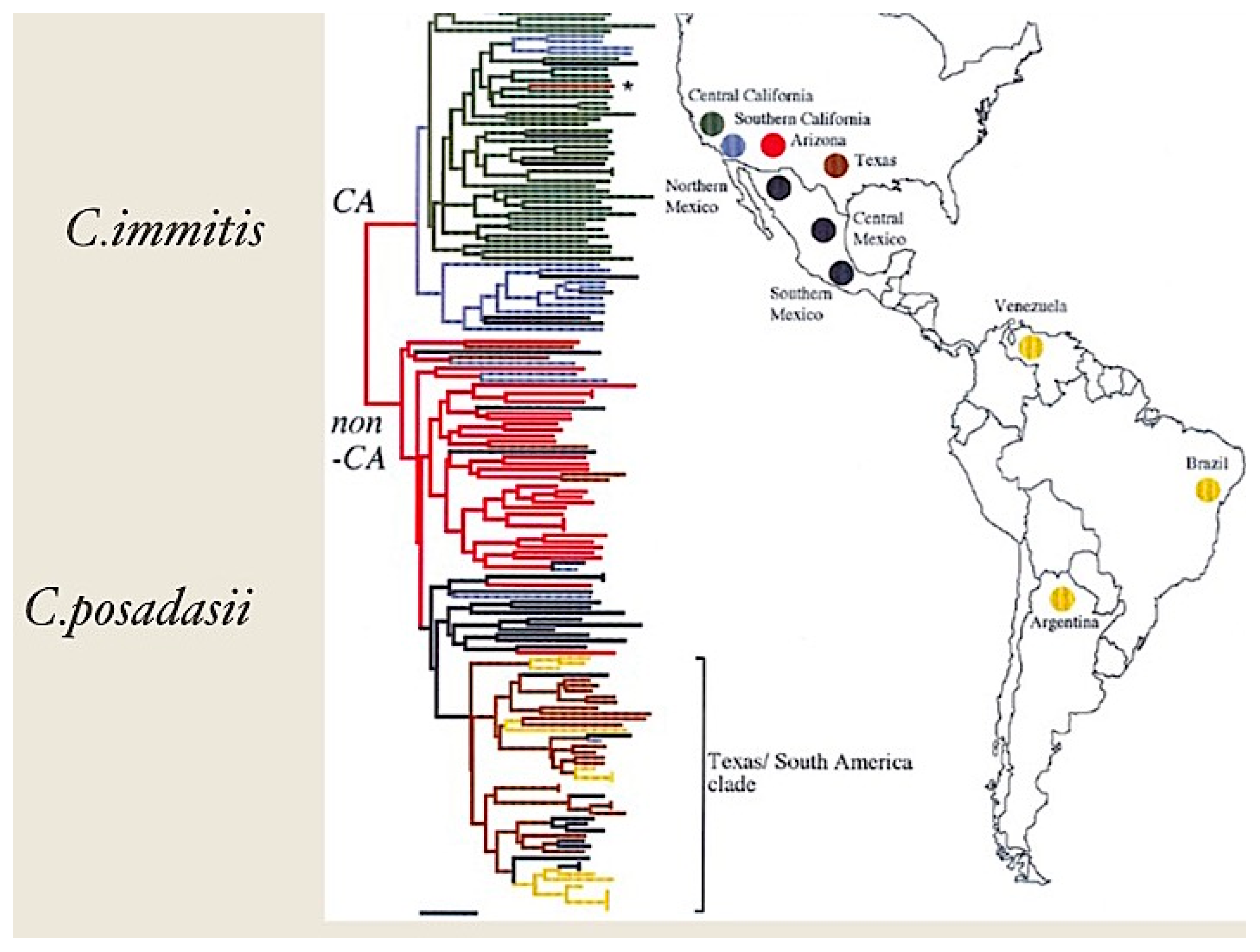
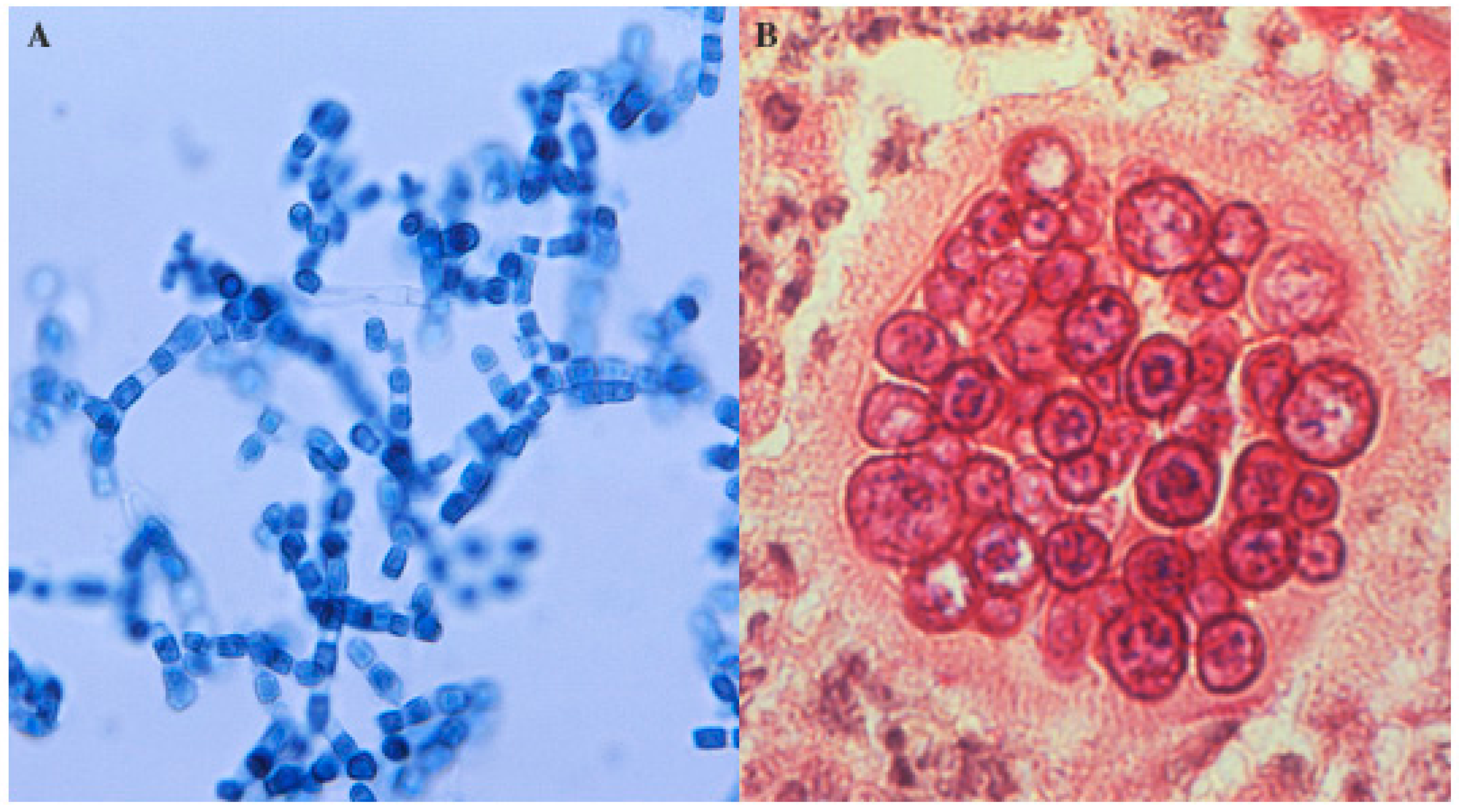
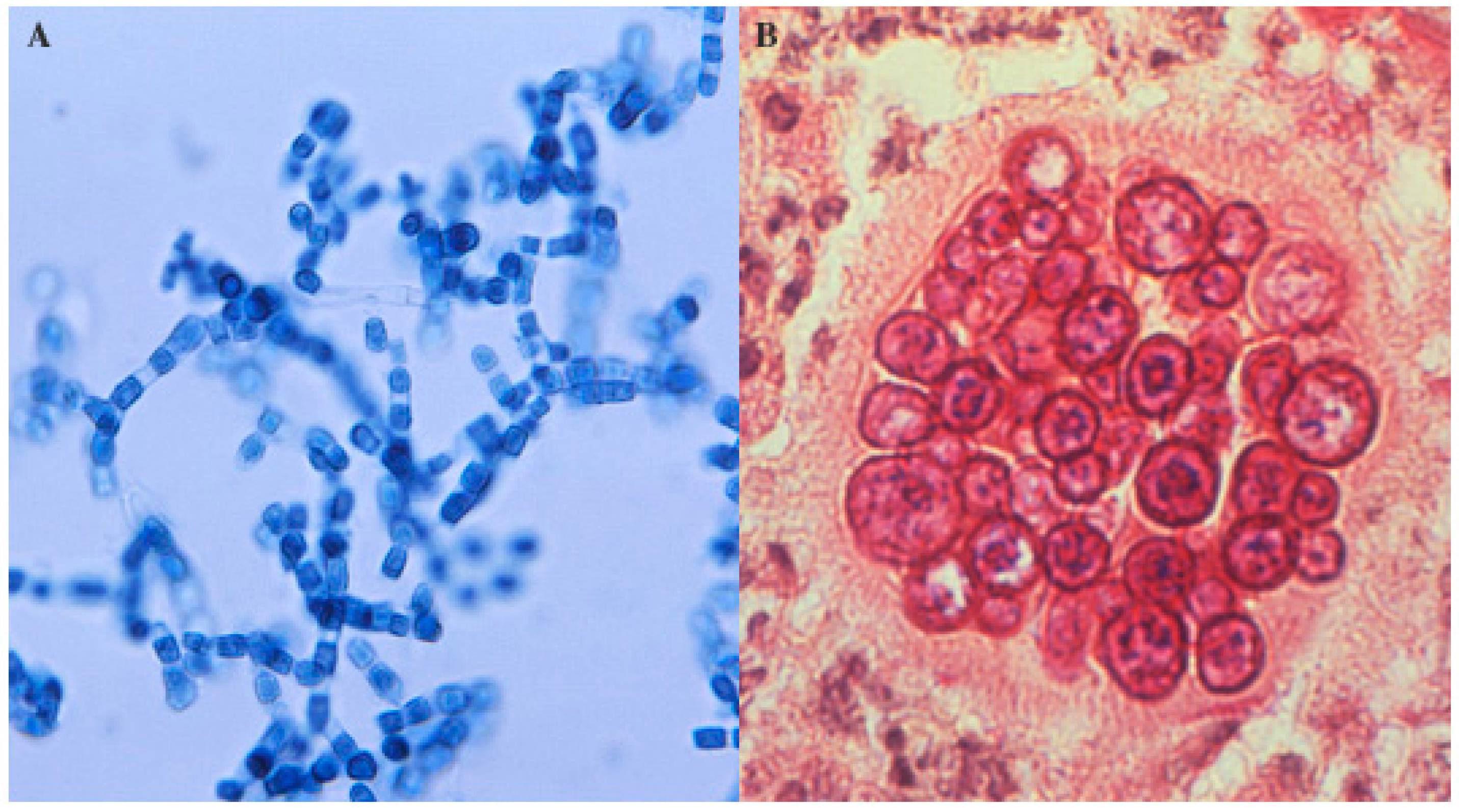
2. Microbiology
3. Immunology
4. Vaccination
4.1. Overview
4.2. Live Attenuated Mutants
4.3. Vaccine Candidate Proteins
5. Other Issues
6. Human Trial
7. A Path to a Human Vaccine
Conflicts of Interest
References
- Fisher, M.C.; Koenig, G.L.; White, T.J.; San-Blas, G.; Negroni, R.; Alvarez, I.G.; Wanke, B.; Taylor, J.W. Biogeographic range expansion into South America by Coccidioides immitis mirrors new world patterns of human migration. Proc. Natl. Acad. Sci. USA 2001, 98, 4558–4562. [Google Scholar] [CrossRef] [PubMed]
- Luna-Isaac, J.A.; Muniz-Salazar, R.; Baptista-Rosas, R.C.; Enriquez-Paredes, L.M.; Castanon-Olivares, L.R.; Contreras-Perez, C.; Bazan-Mora, E.; Gonzalez, G.M.; Gonzalez-Martinez, M.R. Genetic analysis of the endemic fungal pathogens Coccidioides posadasii and Coccidioides immitis in Mexico. Med. Mycol. 2014, 52, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Barker, B.M.; Hoover, S.; Nix, D.E.; Ampel, N.M.; Frelinger, J.A.; Orbach, M.J.; Galgiani, J.N. Recent advances in our understanding of the environmental, epidemiological, immunological, and clinical dimensions of Coccidioidomycosis. Clin. Microbiol. Rev. 2013, 26, 505–525. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Hajjeh, R.; Jibson, R.; Spiegel, R.; Harp, E.; Maeshall, G.; Gunn, R.; McNeil, M.; Pinner, R.; Burger, R.; et al. A Coccidioidomycosis outbreak following the Northridge, Calif, Earthquake. JAMA 1997, 277, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Ampel, N.M.; Blair, J.E.; Catanzaro, A.; Johnson, R.H.; Stevens, D.A.; Williams, P.L. Coccidioidomycosis. Clin. Infect. Dis. 2005, 41, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- CDC. Increase in reported Coccidioidomycosis—United States, 1998–2011. MMWR 2013, 62, 1–20. [Google Scholar]
- Sondermeyer, G.; Lee, L.; Gilliss, D.; Tabnak, F.; Vugia, D. Coccidioidomycosis-associated hospitalizations, California, USA, 2000–2011. Emerg. Infect. Dis 2013, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, L.; Nix, D.; Wright, M.; Lindberg, E.; Fagan, T.; Lieberman, D.; Stoffer, T.; Ampel, N.M.; Galgiani, J.N. Coccidioidomycosis as a common cause of community-acquired pneumonia. Emerg. Infect. Dis 2006, 12, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.; Brown, J.; Benedict, K.; Park, B. Coccidioidomycosis: Epidemiology. CLEP 2013, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.A.; Magee, D.M. Coccidioidomycosis: Host response and vaccine development. Clin. Microbiol. Rev. 2004, 17, 804–839. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Coccidioidomycosis: a Reemerging infectious disease. Emerg. Infect. Dis 1996, 2, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.; Laniado-Laborin, R. Coccidioidomycosis—A fungal disease of the Americas. PLoS Negl. Trop. Dis. 2005, 2, 0015–0019. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.A.; Anderson, S.M.; Imholte, S.B.; Erhart, L.M.; Chen, S.; Park, B.J.; Christ, C.; Komatsu, K.K.; Chiller, T.; Sunenshine, R.H. Enhanced surveillance of Coccidioidomycosis, Arizona, USA, 2007–2008. Emerg. Infect. Dis 2010, 16, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Pappagianis, D. Epidemiology of Coccidioidomycosis. Curr. Top. Med. Mycol. 1988, 2, 199–238. [Google Scholar] [PubMed]
- Dewsnup, D.H.; Galgiani, J.N.; Graybill, J.R.; Diaz, M.; Rendon, A.; Cloud, G.A.; Stevens, D.A. Is It Ever Safe to Stop Azole Therapy for Coccidioides Immitis meningitis? Ann. Intern. Med. 1996, 124, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Whiston, E.; Taylor, J.W. Comparative phylogenomics of pathogenic and nonpathogenic species. G3 2016, 6, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; Singhania, A.; Fierer, J.; Goldberg, J.; Kirkland, T.N.; Woelk, C.H. Gene expression in human fungal pathogen Coccidioides Immitis changes as arthroconidia differentiate into spherules and mature. BMC Microbiol. 2013, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Whiston, E.; Zhang Wise, H.; Sharpton, T.J.; Jui, G.; Cole, G.T.; Taylor, J.W. Comparative transcriptomics of the saprobic and parasitic growth phases in Coccidioides spp. PLoS ONE 2012, 7, e41034. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E.; Whiting, E.G. The use of Coccidioidin. Am. Rev. Tuberc. 1948, 57, 330–360. [Google Scholar] [PubMed]
- Smith, C.E.; Saito, M.T.; Simons, S.A. Pattern of 39,500 serologic tests in Coccidioidomycosis. J. Am. Med. Assoc. 1956, 160, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-J.; Kirkland, T.N.; Hall, L.K.; Wopschall, J.; Smith, R.C.; Hung, C.-Y.; Chen, X.; Tarcha, E.; Thomas, P.W.; Cole, G.T. Characterization of a serodiagnostic complement fixation antigen of Coccidioides posadasii expressed in the nonpathogenic fungus Uncinocarpus reesii. J. Clin. Microbiol. 2005, 43, 5462–5469. [Google Scholar] [CrossRef] [PubMed]
- Ampel, N.M.; Kramer, L.A.; Li, L.; Carroll, D.S.; Kerekes, K.M.; Johnson, S.M.; Pappagianis, D. In vitro whole-blood analysis of cellular immunity in patients with active Coccidioidomycosis by using the antigen preparation T27K. Clin. Diagn. Lab. Immunol. 2002, 9, 1039–1043. [Google Scholar] [CrossRef] [PubMed]
- Nesbit, L.A.; Knox, K.S.; Nguyen, C.T.; Roesch, J.; Wheat, L.J.; Johnson, S.M.; Pappagianis, D.; Chavez, S.; Ampel, N.M. Immunological characterization of bronchoalveolar lavage fluid in patients with acute pulmonary Coccidioidomycosis. J. Infect. Dis. 2013, 208, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, M.; Gern, B.; Hung, C. Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice. J. Clin. Investig. 2011, 121, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.A.; Viriyakosol, S.; Fierer, J.; Kirkland, T.N. The Role of reactive oxygen intermediates in experimental Coccidioidomycois in mice. BMC Microbiol. 2011, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Nitric oxide synthase activity has limited influence on the control of Coccidioides infection in mice. Microb. Pathog. 2011, 51, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.; Fierer, J. Inbred mouse strains differ in resistance to lethal Coccidioides Immitis infection. Infect. Immun. 1983, 40, 912–916. [Google Scholar] [PubMed]
- Kong, Y.C.; Levine, H.B. Experimentally induced immunity in the mycoses. Bacteriol. Rev. 1967, 31, 35–53. [Google Scholar]
- Zimmermann, C.R.; Johnson, S.M.; Martens, G.W.; White, A.G.; Zimmer, B.L.; Pappagianis, D. Protection against lethal murine Coccidioidomycosis by a soluble vaccine from spherules. Infect. Immun. 1998, 66, 2342–2345. [Google Scholar] [PubMed]
- Xue, J.; Chen, X.; Selby, D.; Hung, C.Y.; Yu, J.J.; Cole, G.T. A Genetically engineered live attenuated vaccine of Coccidioides Posadasii protects BALB/C mice against Coccidioidomycosis. Infect. Immun. 2009, 77, 3196–3208. [Google Scholar] [CrossRef] [PubMed]
- Narra, H.P.; Shubitz, L.F.; Mandel, M.A.; Trinh, H.T.; Griffin, K.; Buntzman, A.S.; Frelinger, J.A.; Galgiani, J.N.; Orbach, M.J. A Coccidioides CPS1 deletion mutant is avirulent and protects mice from lethal infection. Infect. Immun. 2016, 84, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- Pappagianis, D. Evaluation of the protective efficacy of the killed Coccidioides immitis spherule vaccine in humans. Am. J. Respir. Cell Mol. Biol. 1993, 148, 656–660. [Google Scholar]
- Abuodeh, R.O.; Shubitz, L.F.; Siegel, E.; Snyder, S.; Peng, T.; Orsborn, K.I.; Brummer, E.; Stevens, D.A.; Galgiani, J.N. Resistance to Coccidioides immitis in mice after immunization with recombinant protein or a DNA vaccine of a proline-rich antigen. Infect. Immun. 1999, 67, 2935–2940. [Google Scholar] [PubMed]
- Delgado, N.; Xue, J.; Yu, J.J.; Hung, C.Y.; Cole, G.T. A Recombinant β-1,3-glucanosyltransferase homolog of Coccidioides posadasii protects mice against Coccidioidomycosis. Infect. Immun. 2003, 71, 3010–3019. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, M.; Brandhorst, T.T.; Sullivan, T.D.; Filutowicz, H.; Sterkel, A.; Stewart, D.; Li, M.; Lerksuthirat, T.; LeBert, V.; Shen, Z.T.; et al. Calnexin induces expansion of antigen-specific CD4+ T cells that confer immunity to fungal ascomycetes via conserved epitopes. Cell Host Microbe 2015, 17, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Basrur, V.; Hung, C.Y.; Gardner, M.J.; Cole, G.T. A Recombinant aspartyl protease of coccidioides Posadasii induces protection against pulmonary Coccidioidomycosis in mice. Infect. Immun. 2006, 74, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L.F.; Yu, J.J.; Hung, C.Y.; Kirkland, T.N.; Peng, T.; Perrill, R.; Simons, J.; Xue, J.; Herr, R.A.; Cole, G.T.; et al. Improved protection of mice against lethal respiratory infection with Coccidioides posadasii using two recombinant antigens expressed as a single protein. Vaccine 2006, 24, 5904–5911. [Google Scholar] [CrossRef] [PubMed]
- Hurtgen, B.J.; Hung, C.-Y.; Ostroff, G.R.; Levitz, S.M.; Cole, G.T. Construction and evaluation of a novel recombinant T cell epitope-based vaccine against Coccidioidomycosis. Infect. Immun. 2012, 80, 3960–3974. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, T.J.; Stajich, J.E.; Rounsley, S.D.; Gardner, M.J.; Wortman, J.R.; Jordar, V.S.; Maiti, R.; Kodira, C.D.; Neafsey, D.E.; Zeng, Q.; et al. Comparative genomic analyses of the human fungal pathogens Coccidioides and their relatives. Genome Res. 2009, 19, 1722–1731. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Ampel, N.M.; Christian, L.; Seshan, K.R.; Cole, G.T. A Major cell surface antigen of Coccidioides immitis which elicits both humoral and cellular immune responses. Infect. Immun. 2000, 68, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Huppert, M.; Spratt, N.S.; Vukovich, K.R.; Sun, S.H. Antigenic analysis of coccidioidin and spherulin determined by two-dimensional immunoelectrophoresis. Infect. Immun. 1978, 20, 541–551. [Google Scholar]
- Zhu, Y.; Yang, C.; Magee, D.M.; Cox, R.A. Molecular cloning and characterization of Coccidioides immitis antigen 2 cDNA. Infect. Immun. 1996, 64, 2695–2699. [Google Scholar] [PubMed]
- Dugger, K.E.A. Cloning and sequence analysis of the cDNA for a protein from Coccidioides immitis with immunogenic potential. Biochem. Biophys. Res. Commun. 1996, 218, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Raz, E.; Datta, S.K. Molecular and cellular mechanisms of protective immunity to Coccidioidomycosis. Vaccine 2006, 24, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Johannesson, H.; Vidal, P.; Guarro, J.; Herr, R.; Cole, G.; Taylor, J. Positive directional selection in the proline-rich antigen (PRA) gene among the human pathogenic fungi Coccidioides immitis, C. posadasii and their closest relatives. Mol. Biol. Evol. 2004, 21, 1131–1145. [Google Scholar]
- Tarcha, E.J.; Basrur, V.; Hung, C.Y.; Gardner, M.J.; Cole, G.T. Multivalent recombinant protein vaccine against Coccidioidomycosis. Infect. Immun. 2006, 74, 5802–5813. [Google Scholar] [CrossRef] [PubMed]
- Orsborn, K.I.; Shubitz, L.F.; Peng, T.; Kellner, E.M.; Orbach, M.J.; Haynes, P.A.; Galgiani, J.N. Protein expression profiling of Coccidioides posadasii by two-dimensional differential in-gel electrophoresis and evaluation of a newly recognized peroxisomal matrix protein as a recombinant vaccine candidate. Infect. Immun. 2006, 74, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Dowling, J.K.; Mansell, A. Toll-Like Receptors: The Swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 5, e85. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Lerche, N.W.; Pappagianis, D.; Yee, J.L.; Galgiani, J.N.; Hector, R.F. Safety, antigenicity, and efficacy of a recombinant Coccidioidomycosis vaccine in Cynomolgus Macaques (Macaca Fascicularis). Ann. N. Y. Acad. Sci. 2007, 1111, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L. Comparative aspects of Coccidioidomycosis in animals and human. Ann. N. Y. Acad. Sci. 2007, 1111, 395–403. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Antigen | Form | Adjuvant | Activity | Reference |
|---|---|---|---|---|
| Live attenuated mutants | N/A a | N/A | Active | [30,31] |
| Formalin-killed spherules | N/A | N/A | Active in mice but not humans | [28,32] |
| Spherule extract | N/A | Various | Active | [29] |
| Ag2/PRA | Protein, DNA | Various | Moderately active Inactive | [33,10] |
| β-glucanosyltransferase | Protein | CpG-ODN b | Moderately active | [34] |
| Calnexin | Protein | Glucan and Adjuplex | Modestly active | [35] |
| Aspartyl protease | Protein | CpG-ODN | Moderately active | [36] |
| CSA c | Protein | CpG-ODN and MPLA d | Modestly active | [37] |
| Ag2/PRA and CSA fusion protein | Protein | CpG-ODN and MPLA | Highly active | [37] |
| Phospholipase, α-mannosidase and aspartyl protease | Protein | CpG-ODN | Highly active | [36,38] |
© 2016 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirkland, T.N. The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas. J. Fungi 2016, 2, 34. https://doi.org/10.3390/jof2040034
Kirkland TN. The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas. Journal of Fungi. 2016; 2(4):34. https://doi.org/10.3390/jof2040034
Chicago/Turabian StyleKirkland, Theo N. 2016. "The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas" Journal of Fungi 2, no. 4: 34. https://doi.org/10.3390/jof2040034
APA StyleKirkland, T. N. (2016). The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas. Journal of Fungi, 2(4), 34. https://doi.org/10.3390/jof2040034