HIV-Associated Cryptococcal Immune Reconstitution Inflammatory Syndrome Is Associated with Aberrant T Cell Function and Increased Cytokine Responses

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects and Procedures
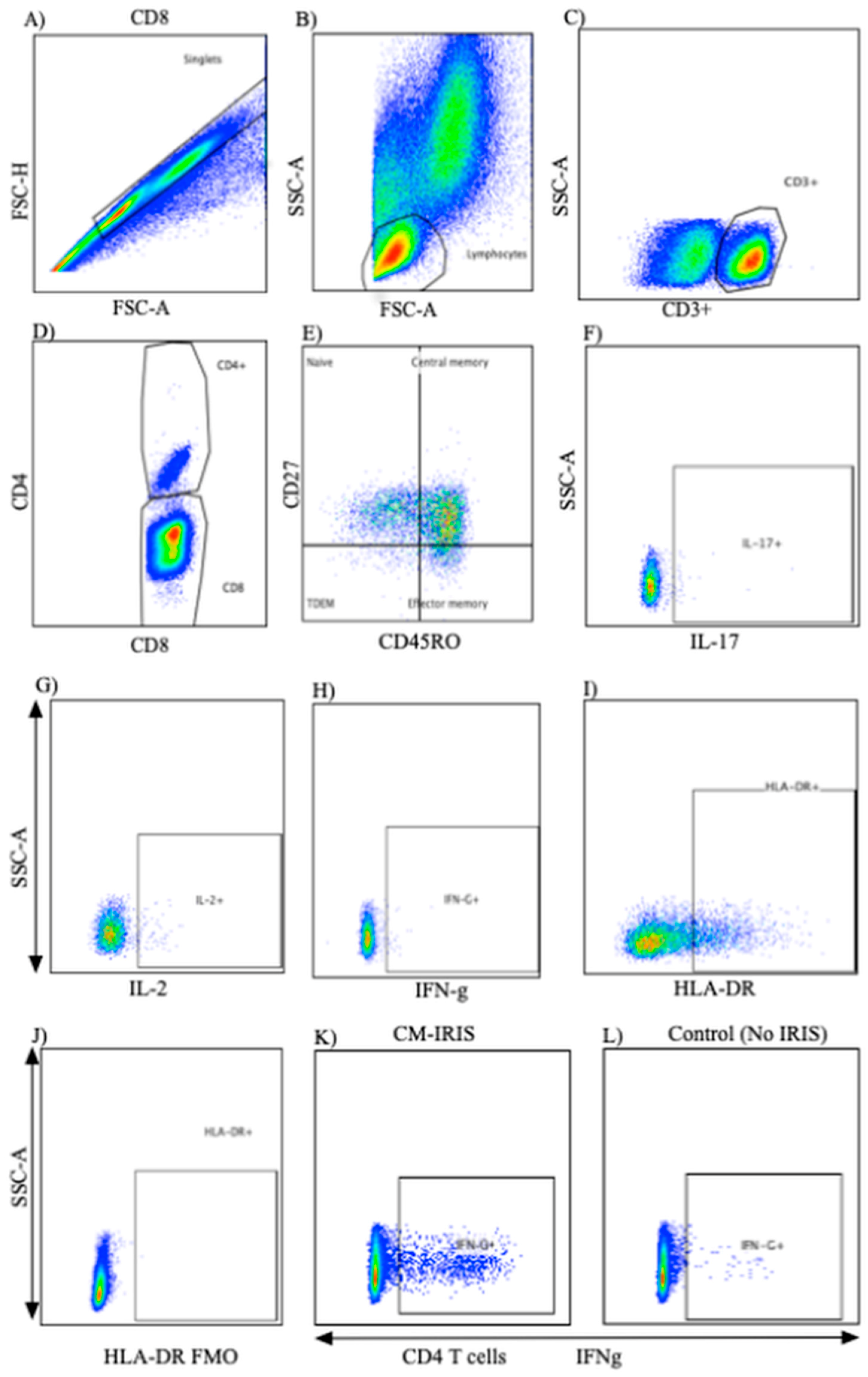
2.2. PBMC Stimulation and Surface Flow Cytometric Staining
2.3. Intracellular Cytokine Staining
2.4. Statistical Analysis
3. Results
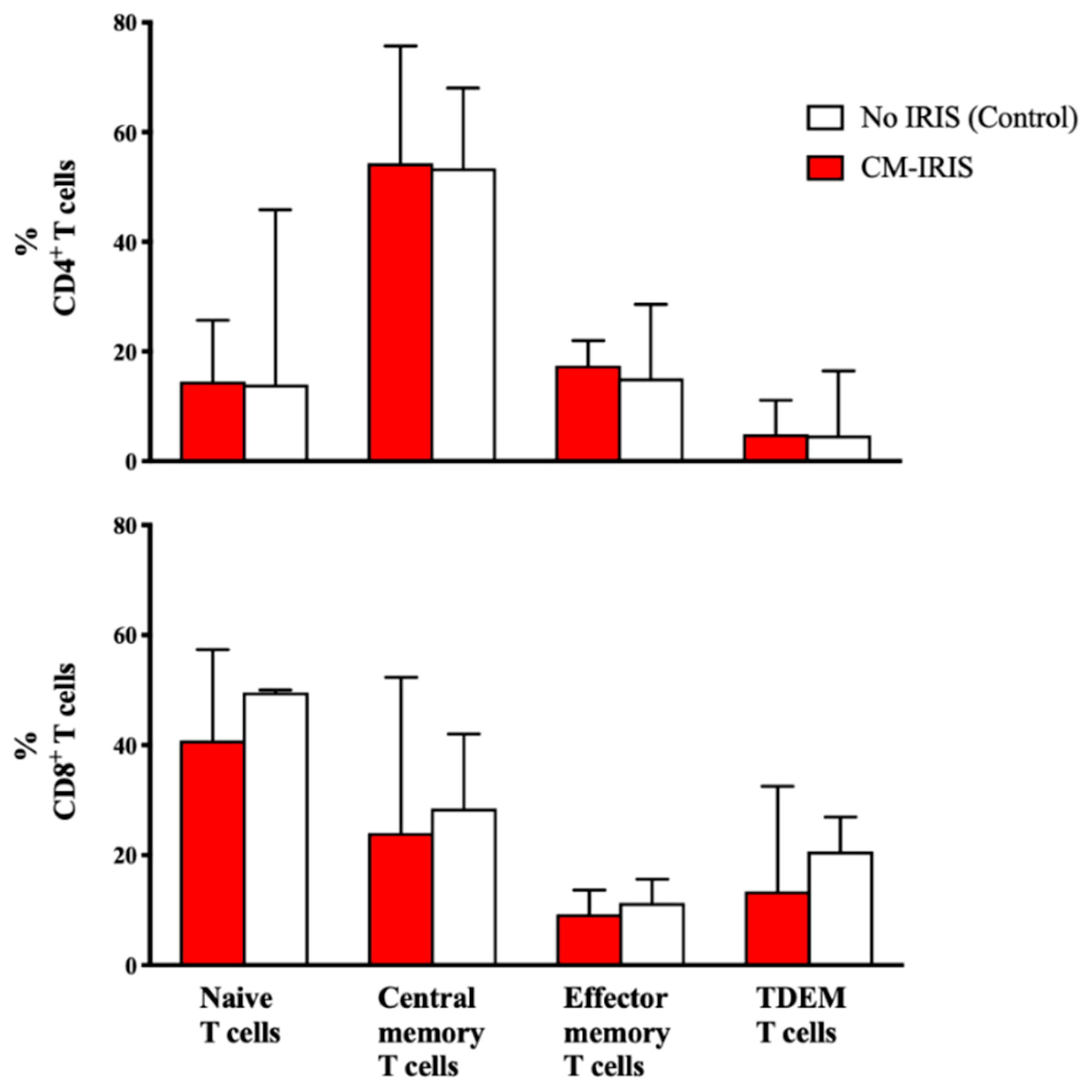
3.1. T Cell Phenotype and Activation at CM Diagnosis vs. Controls
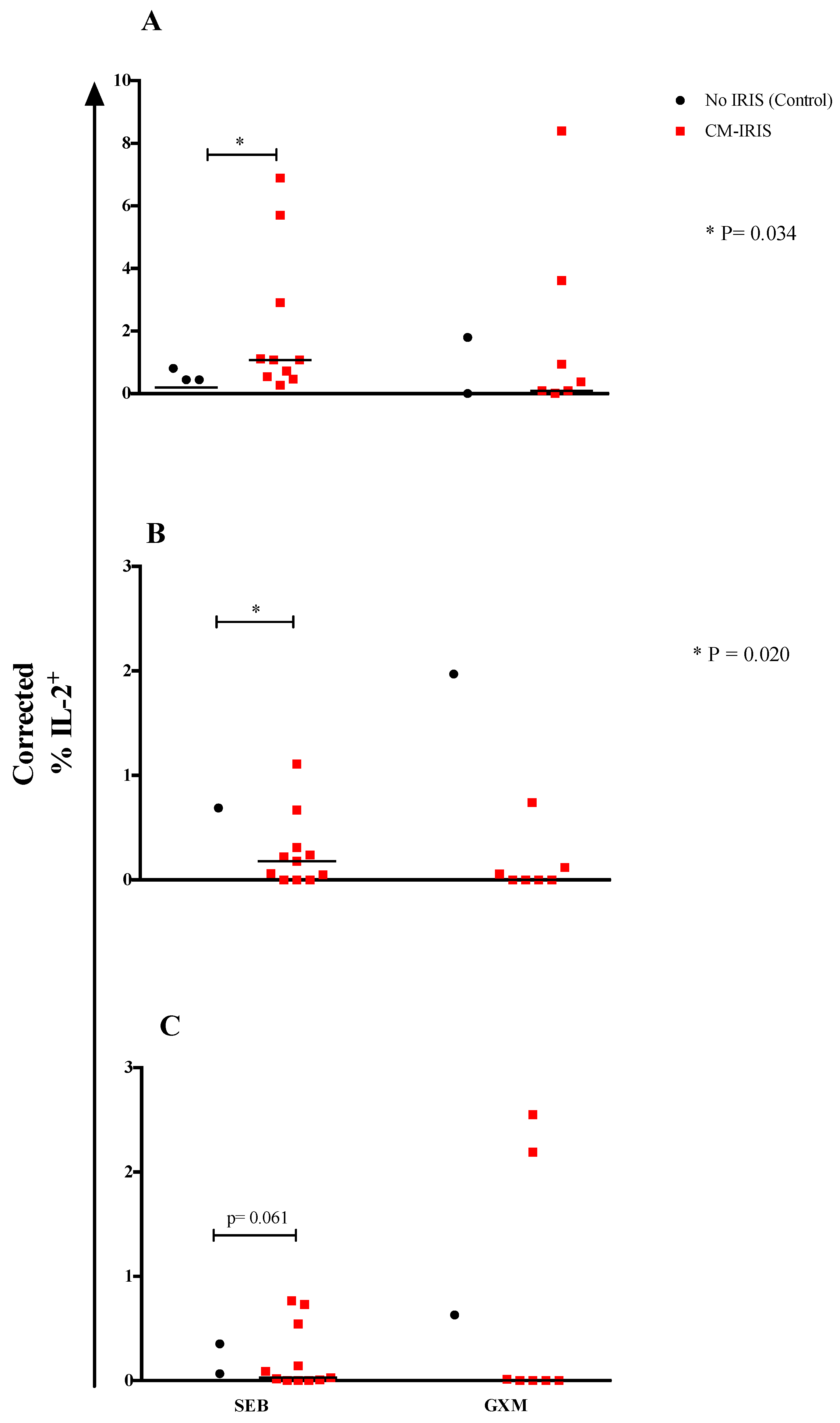
3.2. T Cell Cytokine Responses at Baseline
3.3. Cytokine Responses at CM-IRIS vs. Controls
3.4. Phenotype and Cytokine Responses among Subjects with Paired Samples at CM Diagnosis vs. CM-IRIS
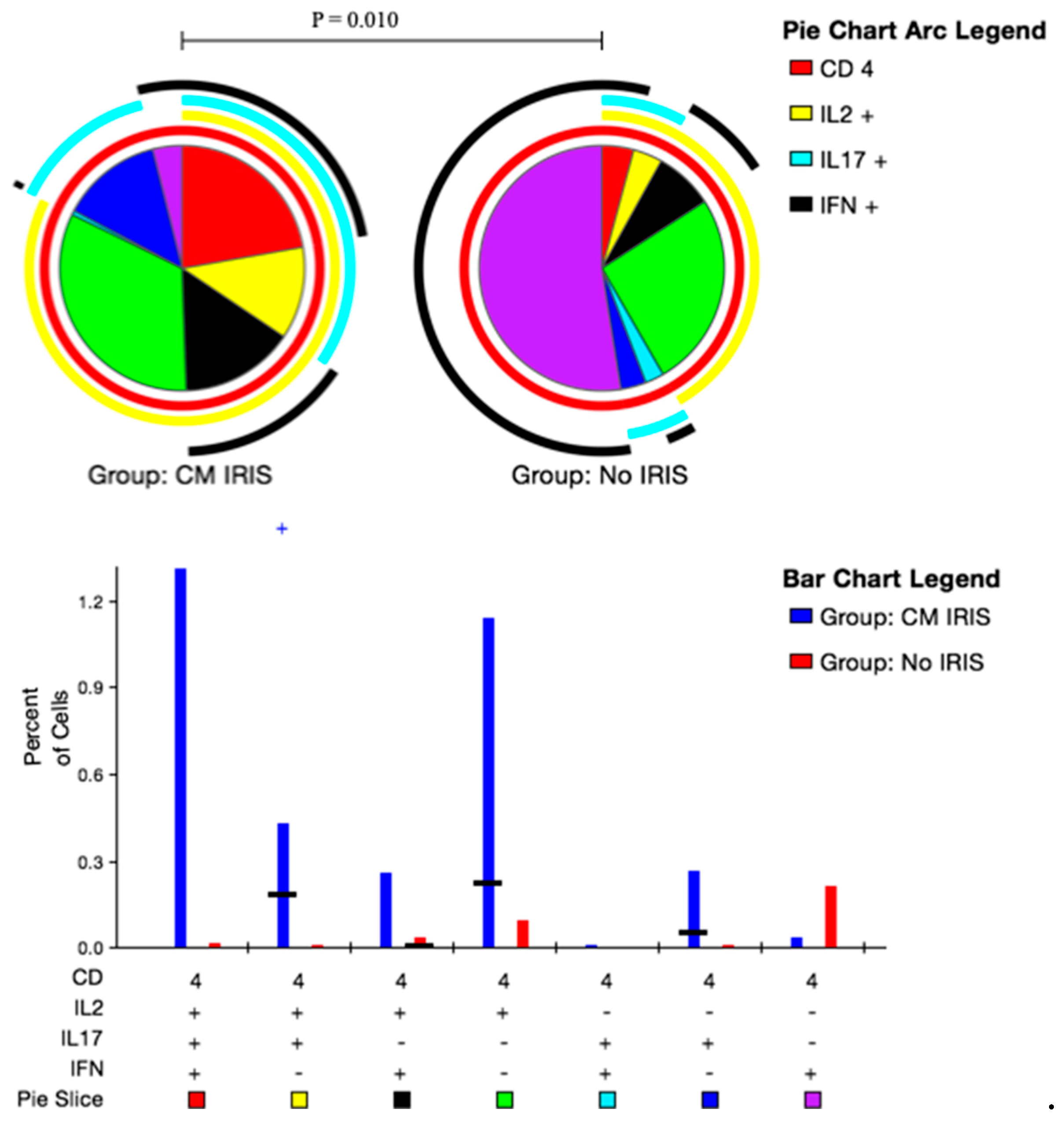
3.5. CD4+ and CD8+ T Cell Polyfunctional Cytokine Responses at CM-IRIS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Gordon, S.B.; Walsh, A.L.; Chaponda, M.; Gordon, M.A.; Soko, D.; Mbwvinji, M.; Molyneux, M.E.; Read, R.C. Bacterial meningitis in Malawian adults: Pneumococcal disease is common, severe, and seasonal. Clin. Infect. Dis. 2000, 31, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Hakim, J.G.; Gangaidzo, I.T.; Heyderman, R.S.; Mielke, J.; Mushangi, E.; Taziwa, A.; Robertson, V.J.; Musvaire, P.; Mason, P.R. Impact of HIV infection on meningitis in Harare, Zimbabwe: A prospective study of 406 predominantly adult patients. AIDS 2000, 14, 1401–1407. [Google Scholar]
- Jarvis, J.N.; Meintjes, G.; Williams, A.; Brown, Y.; Crede, T.; Harrison, T.S. Adult meningitis in a setting of high HIV and TB prevalence: Findings from 4961 suspected cases. BMC Infect. Dis. 2010, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Durski, K.N.; Kuntz, K.M.; Yasukawa, K.; Virnig, B.A.; Meya, D.B.; Boulware, D.R. Cost-effective diagnostic checklists for meningitis in resource-limited settings. J. Acquir. Immune Defic. Syndr. 2013, 63, e101–e108. [Google Scholar] [CrossRef]
- Cohen, D.B.; Zijlstra, E.E.; Mukaka, M.; Reiss, M.; Kamphambale, S.; Scholing, M.; Waitt, P.I.; Neuhann, F. Diagnosis of cryptococcal and tuberculous meningitis in a resource-limited African setting. Tropical Medicine & International Health 2010, 15, 910–917. [Google Scholar] [CrossRef]
- IeDEA and ART Cohort Collaborations; Avila, D.; Althoff, K.N.; Mugglin, C.; Wools-Kaloustian, K.; Koller, M.; Dabis, F.; Nash, D.; Gsponer, T.; Sungkanuparph, S.; et al. Immunodeficiency at the start of combination antiretroviral therapy in low-, middle-, and high-income countries. J. Acquir. Immune Defic. Syndr. 2014, 65, e8–e16. [Google Scholar] [CrossRef] [PubMed]
- Haddow, L.J.; Colebunders, R.; Meintjes, G.; Lawn, S.D.; Elliott, J.H.; Manabe, Y.C.; Bohjanen, P.R.; Sungkanuparph, S.; Easterbrook, P.J.; French, M.A.; et al. Cryptococcal immune reconstitution inflammatory syndrome in HIV-1-infected individuals: Proposed clinical case definitions. Lancet Infect. Dis. 2010, 10, 791–802. [Google Scholar] [CrossRef]
- DeSimone, J.A.; Pomerantz, R.J.; Babinchak, T.J. Inflammatory reactions in HIV-1-infected persons after initiation of highly active antiretroviral therapy. Ann. Intern. Med. 2000, 133, 447–454. [Google Scholar] [CrossRef]
- Lederman, M.M.; Valdez, H. Immune restoration with antiretroviral therapies: Implications for clinical management. JAMA 2000, 284, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Post, M.J.; Thurnher, M.M.; Clifford, D.B.; Nath, A.; Gonzalez, R.G.; Gupta, R.K.; Post, K.K. CNS-immune reconstitution inflammatory syndrome in the setting of HIV infection, part 2: Discussion of neuro-immune reconstitution inflammatory syndrome with and without other pathogens. AJNR 2013, 34, 1308–1318. [Google Scholar] [CrossRef]
- Jenny-Avital, E.R.; Abadi, M. Immune reconstitution cryptococcosis after initiation of successful highly active antiretroviral therapy. Clin. Infect. Dis. 2002, 35, e128–e133. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, Y.D.; Greenwald, J.H.; DerSimonian, R.; Roby, G.; Antonelli, L.R.; Sher, A.; Roederer, M.; Sereti, I. Selective expansion of polyfunctional pathogen-specific CD4(+) T cells in HIV-1-infected patients with immune reconstitution inflammatory syndrome. Blood 2012, 119, 3105–3112. [Google Scholar] [CrossRef]
- Chang, C.C.; Lim, A.; Omarjee, S.; Levitz, S.M.; Gosnell, B.I.; Spelman, T.; Elliott, J.H.; Carr, W.H.; Moosa, M.Y.; Ndung’u, T.; et al. Cryptococcosis-IRIS is associated with lower cryptococcus-specific IFN-gamma responses before antiretroviral therapy but not higher T-cell responses during therapy. J. Infect. Dis. 2013, 208, 898–906. [Google Scholar] [CrossRef]
- Boulware, D.R.; Bonham, S.C.; Meya, D.B.; Wiesner, D.L.; Park, G.S.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Paucity of initial cerebrospinal fluid inflammation in cryptococcal meningitis is associated with subsequent immune reconstitution inflammatory syndrome. J. Infect. Dis. 2010, 202, 962–970. [Google Scholar] [CrossRef]
- Meya, D.B.; Okurut, S.; Zziwa, G.; Cose, S.; Bohjanen, P.R.; Mayanja-Kizza, H.; Joloba, M.; Boulware, D.R.; Yukari Manabe, C.; Wahl, S.; et al. Monocyte Phenotype and IFN-gamma-Inducible Cytokine Responses Are Associated with Cryptococcal Immune Reconstitution Inflammatory Syndrome. J. Fungi (Basel) 2017, 3, 28. [Google Scholar] [CrossRef]
- Boulware, D.R.; Rolfes, M.A.; Rajasingham, R.; von Hohenberg, M.; Qin, Z.; Taseera, K.; Schutz, C.; Kwizera, R.; Butler, E.K.; Meintjes, G.; et al. Multisite validation of cryptococcal antigen lateral flow assay and quantification by laser thermal contrast. Emerg. Infect. Dis. 2014, 20, 45–53. [Google Scholar] [CrossRef]
- Rajasingham, R.; Rhein, J.; Klammer, K.; Musubire, A.; Nabeta, H.; Akampurira, A.; Mossel, E.C.; Williams, D.A.; Boxrud, D.J.; Crabtree, M.B.; et al. Epidemiology of meningitis in an HIV-infected Ugandan cohort. Am. J. Trop. Med. Hyg. 2015, 92, 274–279. [Google Scholar] [CrossRef]
- Boulware, D.R.; Meya, D.B.; Muzoora, C.; Rolfes, M.A.; Huppler Hullsiek, K.; Musubire, A.; Taseera, K.; Nabeta, H.W.; Schutz, C.; Williams, D.A.; et al. Timing of antiretroviral therapy after diagnosis of cryptococcal meningitis. N. Engl. J. Med. 2014, 370, 2487–2498. [Google Scholar] [CrossRef] [PubMed]
- Roederer, M.; Nozzi, J.L.; Nason, M.C. SPICE: Exploration and analysis of post-cytometric complex multivariate datasets. Cytometry 2011, 79, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, J.N.; Casazza, J.P.; Stone, H.H.; Meintjes, G.; Lawn, S.D.; Levitz, S.M.; Harrison, T.S.; Koup, R.A. The phenotype of the Cryptococcus-specific CD4+ memory T-cell response is associated with disease severity and outcome in HIV-associated cryptococcal meningitis. J. Infect. Dis. 2013, 207, 1817–1828. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Alviano, C.S.; Travassos, L.R. Pathogenicity of Cryptococcus neoformans: Virulence factors and immunological mechanisms. Microbes Infect. 1999, 1, 293–301. [Google Scholar] [CrossRef]
- Buchanan, K.L.; Murphy, J.W. What makes Cryptococcus neoformans a pathogen? Emerg. Infect. Dis. 1998, 4, 71–83. [Google Scholar] [CrossRef]
- Feldmesser, M.; Casadevall, A. Mechanism of action of antibody to capsular polysaccharide in Cryptococcus neoformans infection. Front. Biosci. 1998, 3, d136–d151. [Google Scholar] [CrossRef]
- Yauch, L.E.; Lam, J.S.; Levitz, S.M. Direct inhibition of T-cell responses by the Cryptococcus capsular polysaccharide glucuronoxylomannan. PLoS Pathog. 2006, 2, e120. [Google Scholar] [CrossRef]
- Barber, D.L.; Andrade, B.B.; Sereti, I.; Sher, A. Immune reconstitution inflammatory syndrome: The trouble with immunity when you had none. Nat. Rev. Microbiol. 2012, 10, 150–156. [Google Scholar] [CrossRef]
- Lortholary, O.; Fontanet, A.; Memain, N.; Martin, A.; Sitbon, K.; Dromer, F.; French Cryptococcosis Study Group. Incidence and risk factors of immune reconstitution inflammatory syndrome complicating HIV-associated cryptococcosis in France. AIDS 2005, 19, 1043–1049. [Google Scholar] [CrossRef]
- Siddiqui, A.A.; Brouwer, A.E.; Wuthiekanun, V.; Jaffar, S.; Shattock, R.; Irving, D.; Sheldon, J.; Chierakul, W.; Peacock, S.; Day, N.; et al. IFN-gamma at the site of infection determines rate of clearance of infection in cryptococcal meningitis. J. Immunol. 2005, 174, 1746–1750. [Google Scholar] [CrossRef]
- Murdock, B.J.; Huffnagle, G.B.; Olszewski, M.A.; Osterholzer, J.J. Interleukin-17A enhances host defense against cryptococcal lung infection through effects mediated by leukocyte recruitment, activation, and gamma interferon production. Infect. Immu. 2014, 82, 937–948. [Google Scholar] [CrossRef]
- Boulware, D.R.; Meya, D.B.; Bergemann, T.L.; Wiesner, D.L.; Rhein, J.; Musubire, A.; Lee, S.J.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Clinical features and serum biomarkers in HIV immune reconstitution inflammatory syndrome after cryptococcal meningitis: A prospective cohort study. PLoS Med. 2010, 7, e1000384. [Google Scholar] [CrossRef]
- Chang, C.C.; Dorasamy, A.A.; Gosnell, B.I.; Elliott, J.H.; Spelman, T.; Omarjee, S.; Naranbhai, V.; Coovadia, Y.; Ndung’u, T.; Moosa, M.Y.; et al. Clinical and mycological predictors of cryptococcosis-associated immune reconstitution inflammatory syndrome. AIDS 2013, 27, 2089–2099. [Google Scholar] [CrossRef]
- Jarvis, J.N.; Meintjes, G.; Rebe, K.; Williams, G.N.; Bicanic, T.; Williams, A.; Schutz, C.; Bekker, L.G.; Wood, R.; Harrison, T.S. Adjunctive interferon-gamma immunotherapy for the treatment of HIV-associated cryptococcal meningitis: A randomized controlled trial. AIDS 2012, 26, 1105–1113. [Google Scholar] [CrossRef]
- Boulware, D.R.M.D.; Bergemann, T.L.; Wiesner, D.; Rhein, J.; Williams, D.; Vlasova, I.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Gene Expression Biomarkers in Blood Predict Cryptococcal Immune Reconstitution Inflammatory Syndrome (IRIS). In Proceedings of the 5th IAS Conference on HIV Pathogenesis, Treatment & Prevention, Abstract no. TUPEB163, Cape Town, South Africa, 19–23 July 2009. [Google Scholar]
- Vlasova-St Louis, I.; Chang, C.C.; Shahid, S.; French, M.A.; Bohjanen, P.R. Transcriptomic Predictors of Paradoxical Cryptococcosis-Associated Immune Reconstitution Inflammatory Syndrome. Open Forum Infect. Dis. 2018, 5, ofy157. [Google Scholar] [CrossRef]
- Yoon, H.A.; Nakouzi, A.; Chang, C.C.; Kuniholm, M.H.; Carreno, L.J.; Wang, T.; Ndung’u, T.; Lewin, S.R.; French, M.A.; Pirofski, L.A. Association between plasma antibody responses and risk for Cryptococcus-associated immune reconstitution inflammatory syndrome. J. Infect. Dis. 2018, 219, 420–428. [Google Scholar] [CrossRef]
- Hoyer, K.K.; Dooms, H.; Barron, L.; Abbas, A.K. Interleukin-2 in the development and control of inflammatory disease. Immunol. Rev. 2008, 226, 19–28. [Google Scholar] [CrossRef]
- Zhou, Q.; Gault, R.A.; Kozel, T.R.; Murphy, W.J. Immunomodulation with CD40 stimulation and interleukin-2 protects mice from disseminated cryptococcosis. Infect. Immu. 2006, 74, 2161–2168. [Google Scholar] [CrossRef]
- Barber, D.L.; Andrade, B.B.; McBerry, C.; Sereti, I.; Sher, A. Role of IL-6 in Mycobacterium avium--associated immune reconstitution inflammatory syndrome. J. Immunol. 2014, 192, 676–682. [Google Scholar] [CrossRef]
- Goovaerts, O.; Jennes, W.; Massinga-Loembe, M.; Ceulemans, A.; Worodria, W.; Mayanja-Kizza, H.; Colebunders, R.; Kestens, L.; Group, T.-I.S. LPS-binding protein and IL-6 mark paradoxical tuberculosis immune reconstitution inflammatory syndrome in HIV patients. PLoS ONE 2013, 8, e81856. [Google Scholar] [CrossRef]
- Narendran, G.; Andrade, B.B.; Porter, B.O.; Chandrasekhar, C.; Venkatesan, P.; Menon, P.A.; Subramanian, S.; Anbalagan, S.; Bhavani, K.P.; Sekar, S.; et al. Paradoxical tuberculosis immune reconstitution inflammatory syndrome (TB-IRIS) in HIV patients with culture confirmed pulmonary tuberculosis in India and the potential role of IL-6 in prediction. PLoS ONE 2013, 8, e63541. [Google Scholar] [CrossRef]
- Bettelli, E.; Korn, T.; Oukka, M.; Kuchroo, V.K. Induction and effector functions of T(H)17 cells. Nature 2008, 453, 1051–1057. [Google Scholar] [CrossRef]
- Tajima, M.; Wakita, D.; Noguchi, D.; Chamoto, K.; Yue, Z.; Fugo, K.; Ishigame, H.; Iwakura, Y.; Kitamura, H.; Nishimura, T. IL-6-dependent spontaneous proliferation is required for the induction of colitogenic IL-17-producing CD8+ T cells. J. Exp. Med. 2008, 205, 1019–1027. [Google Scholar] [CrossRef]
- Vignesh, R.; Kumarasamy, N.; Lim, A.; Solomon, S.; Murugavel, K.G.; Balakrishnan, P.; Solomon, S.S.; Mayer, K.H.; Swathirajan, C.R.; Chandrasekaran, E.; et al. TB-IRIS after initiation of antiretroviral therapy is associated with expansion of preexistent Th1 responses against Mycobacterium tuberculosis antigens. J. Acquir. Immune Defic. Syndr. 2013, 64, 241–248. [Google Scholar] [CrossRef]
- Grant, P.M.; Komarow, L.; Lederman, M.M.; Pahwa, S.; Zolopa, A.R.; Andersen, J.; Asmuth, D.M.; Devaraj, S.; Pollard, R.B.; Richterman, A.; et al. Elevated interleukin 8 and T-helper 1 and T-helper 17 cytokine levels prior to antiretroviral therapy in participants who developed immune reconstitution inflammatory syndrome during ACTG A5164. J. Infect. Dis. 2012, 206, 1715–1723. [Google Scholar] [CrossRef]
- Zaragoza, O.; Rodrigues, M.L.; De Jesus, M.; Frases, S.; Dadachova, E.; Casadevall, A. The capsule of the fungal pathogen Cryptococcus neoformans. Adv. Appl. Microbiol. 2009, 68, 133–216. [Google Scholar] [CrossRef]
- Delliere, S.; Guery, R.; Candon, S.; Rammaert, B.; Aguilar, C.; Lanternier, F.; Chatenoud, L.; Lortholary, O. Understanding Pathogenesis and Care Challenges of Immune Reconstitution Inflammatory Syndrome in Fungal Infections. J. Fungi (Basel) 2018, 4, 139. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls (n = 6) | CM-IRIS (n = 11) | p-Value | |
|---|---|---|---|
| Men, N (%) | 1 (17%) | 8 (73%) | 0.05 |
| Age, years | 35 (28, 40) | 35 (29, 42) | 0.937 |
| CD4+ T cells/μL-Diagnosis | 8 (5, 166) | 6.5 (4, 28) | 0.828 |
| - >3 month on ART | 156 (55, 309) | 68 (33, 79) | 0.256 |
| CD8+ T cells/μL-Diagnosis | 163 (97, 784) | 256 (140, 591) | 0.515 |
| - >3 month on ART | 1005 (615, 1086) | 831 (565, 997) | 0.463 |
| Plasma HIV RNA (log10 copies/mL) | 5.1 (4.6, 5.2) | 5.3 (4.8, 5.6) | 0.260 |
| CSF Cryptococcus (log10 CFU/mL) | 3.97 (2.45, 5.26) | 5.33 (4.96, 5.46) | 0.078 |
| CSF CRAG titer, 1:x | 4512 (528, 12192) | 7200 (4048, 16384) | 0.455 |
| CSF protein (mg/dL) | 60 (47, 68) | 53 (20, 70) | 0.471 |
| CSF WBC/μL | 20 (<5, 45) | <5 (<5, <5) | 0.169 |
| Duration from ART initiation (days) | 67 (48, 92) | 78 (43, 202) | 0.737 |
| Controls (n = 5) | CM-IRIS (n = 10) | p-Value | |
|---|---|---|---|
| CD4+ T cells | |||
| Central memory (TCM) | |||
| IFN-γ+ | 6 (3, 11) | 0.8 (0, 3) | 0.005 |
| IL-2+ | 5 (2, 22) | 1 (0.1, 3) | 0.012 |
| IL-17+ | 2 (1, 6) | 0.5 (0, 2) | 0.054 |
| Effector memory (TEM) | |||
| IFN-γ+ | 8 (4, 16) | 0.5 (0, 3) | 0.027 |
| IL-2+ | 3 (2, 13) | 0 (0, 0.1) | 0.004 |
| IL-17+ | 2 (1.9, 3) | 0 | <0.001 |
| Terminally differentiated effector memory (TTDEM) | |||
| IFN-γ+ | 3 (1, 16) | 0 (0, 0.4) | 0.005 |
| IL-2+ | 74 (12, 86) | 0 | <0.001 |
| IL-17+ | 0.1 (0, 1.1) | 0 (0, 2) | 0.624 |
| CD8+ T cells | |||
| Central memory (TCM) | |||
| IFN-γ+ | 2.4 (2.2, 3.1) | 0.4 (0.1, 0.5) | <0.001 |
| IL-2+ | 1.1 (0.5, 2.1) | 0.07 (0.03, 0.3) | 0.005 |
| IL-17+ | 1.2 (0.9, 1.4) | 0.08 (0.02, 0.2) | 0.003 |
| Effector memory (TEM) | |||
| IFN-γ+ | 6.2 (3.1, 9.9) | 0.7 (0.2, 1.5) | 0.005 |
| IL-2+ | 1.2 (0.5, 1.7) | 0.01 (0, 0.1) | <0.001 |
| IL-17+ | 1.2 (1.0, 1.3) | 0.06 (0, 0.2) | <0.001 |
| Terminally differentiated effector memory (TTDEM) | |||
| IFN-γ+ | 1.3 (0.6, 3.5) | 0.3 (0.2, 0.9) | 0.037 |
| IL-2+ | 1.1 (0.3, 1.2) | 0.03 (0, 0.6) | 0.068 |
| IL-17+ | 0.09 (0.08, 0.25) | 0.01 (0, 0.17) | 0.119 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meya, D.B.; Okurut, S.; Zziwa, G.; Cose, S.; Boulware, D.R.; Janoff, E.N. HIV-Associated Cryptococcal Immune Reconstitution Inflammatory Syndrome Is Associated with Aberrant T Cell Function and Increased Cytokine Responses. J. Fungi 2019, 5, 42. https://doi.org/10.3390/jof5020042
Meya DB, Okurut S, Zziwa G, Cose S, Boulware DR, Janoff EN. HIV-Associated Cryptococcal Immune Reconstitution Inflammatory Syndrome Is Associated with Aberrant T Cell Function and Increased Cytokine Responses. Journal of Fungi. 2019; 5(2):42. https://doi.org/10.3390/jof5020042
Chicago/Turabian StyleMeya, David B., Samuel Okurut, Godfrey Zziwa, Stephen Cose, David R. Boulware, and Edward N. Janoff. 2019. "HIV-Associated Cryptococcal Immune Reconstitution Inflammatory Syndrome Is Associated with Aberrant T Cell Function and Increased Cytokine Responses" Journal of Fungi 5, no. 2: 42. https://doi.org/10.3390/jof5020042