
Blood Serum Stimulates the Virulence Potential of Mucorales through Enhancement in Mitochondrial Oxidative Metabolism and Rhizoferrin Production
 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Spore Germination
2.3. Total RNA and DNA Isolation from M. lusitanicus
2.4. Oligonucleotide Design and Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.5. Determination of Mitochondrial DNA Abundance Using qPCR
2.6. Determination of Glucose and Ethanol Levels
2.7. Caenorhabditis Elegans and Galleria Mellonella Killing Assays
2.8. Relative Quantitation of Rhizoferrin
2.9. Respiration Measurements
2.10. Mitochondrial Membrane Potential and Hydroxyl Radical Quantification
2.11. Statistical Analysis
3. Results
3.1. Blood Serum Increases the Mitochondrial Activity, ROS Generation, and Virulence of M. lusitanicus
3.2. M. lusitanicus Spores Cultured in Blood Serum Demonstrate Improved Hyphal Development under Low Oxygen Levels
3.3. Blood Serum Increases the Mitochondrial Metabolism and Virulence of Clinically Relevant Mucorales
3.4. Blood Serum Increases the Rhizoferrin Levels in M. lusitanicus and other Clinically Relevant Mucorales
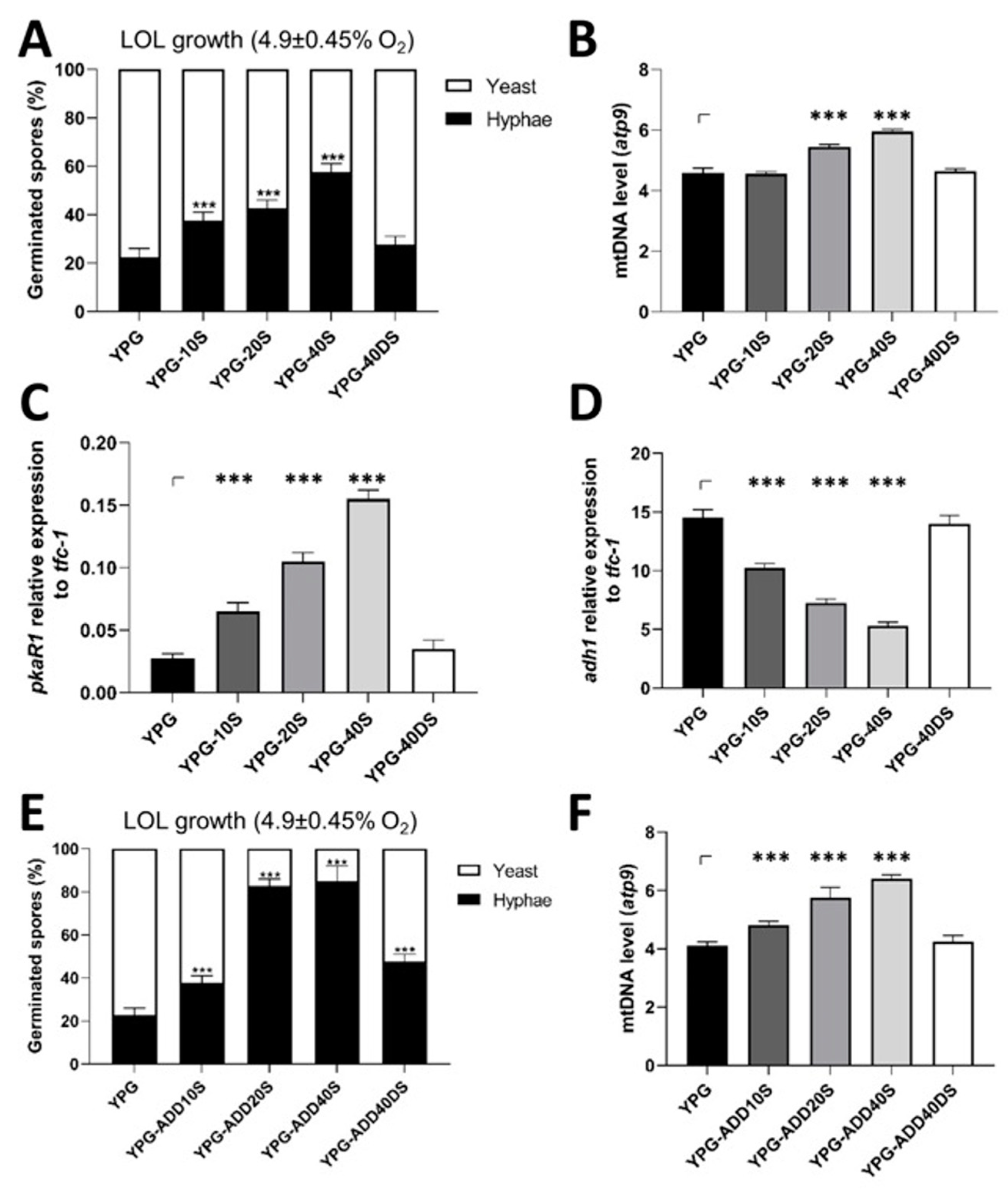
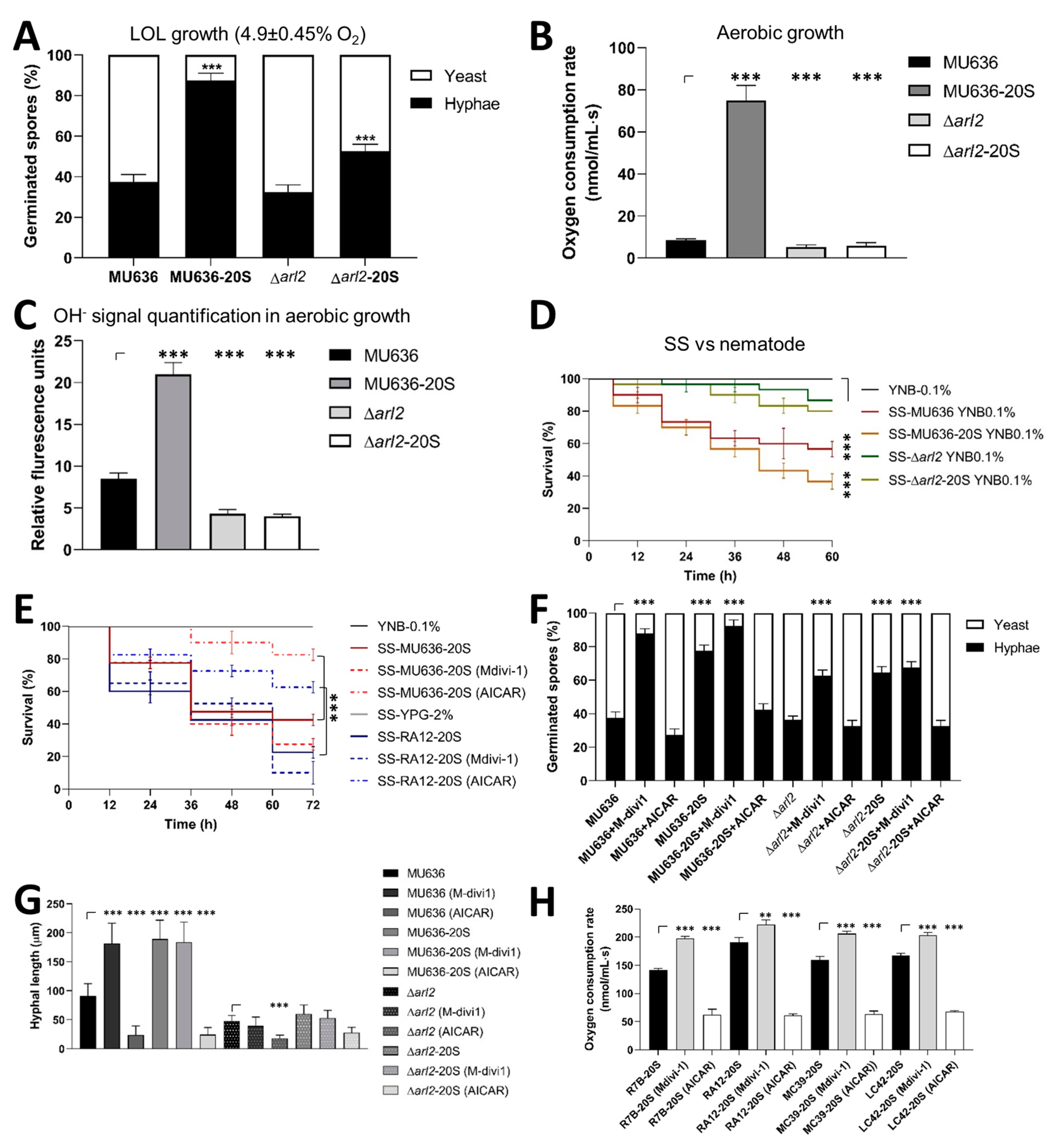
3.5. Blood Serum Requires Increased Mitochondrial Content for a Virulent Phenotype and Hyphal Growth under Low Oxygen Levels
3.6. Effect of N-acetylglucosamine on the Toxicity of Mucorales-Derived SS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lax, C.; Pérez-Arques, C.; Navarro-Mendoza, M.I.; Cánovas-Márquez, J.T.; Tahiri, G.; Pérez-Ruiz, J.A.; Osorio-Concepción, M.; Murcia-Flores, L.; Navarro, E.; Garre, V.; et al. Pathways, and Mechanisms Involved in the Virulence of Mucorales. Genes 2020, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Alqarihi, A.; Ibrahim, A.S. Mucormycosis. Ref. Modul. Life Sci. 2020, 1, 600–612. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, J.B.; de Hoog, G.S.; Bensch, K.; Schwartze, V.U.; Voigt, K.; Alastruey-Izquierdo, A.; Kurzai, O.; Walther, G. A new species concept for the clinically relevant Mucor circinelloides complex. Persoonia 2020, 44, 67–97. [Google Scholar] [CrossRef] [PubMed]
- Lax, C.; Cánovas-Márquez, J.T.; Tahiri, G.; Navarro, E.; Garre, V.; Nicolás, F.E. Genetic Manipulation in Mucorales and New Developments to Study Mucormycosis. Int. J. Mol. Sci. 2022, 23, 3454. [Google Scholar] [CrossRef] [PubMed]
- Petrikkos, G.; Tsioutis, C. Recent Advances in the Pathogenesis of Mucormycosis. Clin. Ther. 2018, 40, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Hoenigl, M.; Seidel, D.; Sprute, R.; Cunha, C.; Oliverio, M.; Goldman, G.H.; Ibrahim, A.S.; Carvalho, A. COVID-19-associated fungal infections. Nat. Microbiol. 2022, 7, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Choksi, T.; Agrawal, A.; Date, P.; Rathod, D.; Gharat, A.; Ingole, A.; Chaudhari, B.; Pawar, N. Cumulative Mortality and Factors Associated With Outcomes of Mucormycosis After COVID-19 at a Multispecialty Tertiary Care Center in India. JAMA Ophthalmol. 2022, 140, 66–72. [Google Scholar] [CrossRef]
- Kumar, A. Mucormycosis in COVID-19 recovered patients. J. Med. Virol. 2022, 94, 1272–1273. [Google Scholar] [CrossRef]
- Orlowski, M. Mucor dimorphism. Microbiol. Rev. 1991, 55, 234–258. [Google Scholar] [CrossRef]
- McIntyre, M.; Breum, J.; Arnau, J.; Nielsen, J. Growth physiology and dimorphism of Mucor circinelloides (syn. racemosus) during submerged batch cultivation. Appl. Microbiol. Biotechnol. 2002, 58, 495–502. [Google Scholar] [CrossRef]
- Lee, S.C.; Li, A.; Calo, S.; Heitman, J. Calcineurin plays key roles in the dimorphic transition and virulence of the human pathogenic zygomycete Mucor circinelloides. PLoS Pathog. 2013, 9, e1003625. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Pérez, S.P.; Patiño-Medina, J.A.; Valle-Maldonado, M.I.; López-Torres, A.; Jácome-Galarza, I.E.; Anaya-Martínez, V.; Gómez-Ruiz, V.; Campos-García, J.; Nuñez-Anita, R.E.; Ortiz-Alvarado, R.; et al. Alteration of Fermentative Metabolism Enhances Mucor circinelloides Virulence. Infect. Immun. 2020, 88, e00434-19. [Google Scholar] [CrossRef] [PubMed]
- Alejandre-Castañeda, V.; Patiño-Medina, J.A.; Valle-Maldonado, M.I.; Nuñez-Anita, R.E.; Santoyo, G.; Castro-Cerritos, K.V.; Ortiz-Alvarado, R.; Corrales-Escobosa, A.R.; Ramírez-Díaz, M.I.; Gutiérrez-Corona, J.F.; et al. Secretion of the siderophore rhizoferrin is regulated by the cAMP-PKA pathway and is involved in the virulence of Mucor lusitanicus. Sci. Rep. 2022, 12, 10649. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Porras, R.A.; Díaz-Pérez, S.P.; Mendoza-Hernández, J.M.; Romo-Rodríguez, P.; Alejandre-Castañeda, V.; Valle-Maldonado, M.I.; Torres-Guzmán, J.C.; González-Hernández, G.A.; Campos-Garcia, J.; Arnau, J.; et al. Alcohol dehydrogenase 1 participates in the Crabtree effect and connects fermentative and oxidative metabolism in the Zygomycete Mucor circinelloides. J. Microbiol. 2019, 57, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Medina, J.A.; Valle-Maldonado, M.I.; Vargas-Tejeda, D.; Chávez-Jacobo, V.M.; Corrales-Escobosa, A.R.; Ramírez-Emiliano, J.; Ruiz-Herrera, L.F.; Ramírez-Díaz, M.I.; Garre, V.; Meza-Carmen, V. Arf-like proteins (Arl1 and Arl2) are involved in mitochondrial homeostasis in Mucor circinelloides. Fungal Biol. 2020, 124, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Medina, J.A.; Vargas-Tejeda, D.; Valle-Maldonado, M.I.; Alejandre-Castañeda, V.; Jácome-Galarza, I.E.; Villegas-Moreno, J.; Nuñez-Anita, R.E.; Ramírez-Díaz, M.I.; Ortiz-Alvarado, R.; Meza-Carmen, V. Sporulation on blood serum increases the virulence of Mucor circinelloides. Microb. Pathog. 2019, 137, 103737. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.S.; Spellberg, B.; Walsh, T.J.; Kontoyiannis, D.P. Pathogenesis of mucormycosis. Clin. Infect. Dis. 2012, 54, S16–S22. [Google Scholar] [CrossRef]
- Grahl, N.; Shepardson, K.M.; Chung, D.; Cramer, R.A. Hypoxia and fungal pathogenesis: To air or not to air? Eukaryot. Cell 2012, 11, 560–570. [Google Scholar] [CrossRef]
- Roncero, M.I.; Jepsen, L.P.; Strøman, P.; van Heeswijck, R. Characterization of a leuA gene and an ARS element from Mucor circinelloides. Gene 1989, 84, 335–343. [Google Scholar] [CrossRef]
- Navarro-Mendoza, M.I.; Pérez-Arques, C.; Panchal, S.; Nicolás, F.E.; Mondo, S.J.; Ganguly, P.; Pangilinan, J.; Grigoriev, I.V.; Heitman, J.; Sanyal, K.; et al. Early Diverging Fungus Mucor circinelloides Lacks Centromeric Histone CENP-A and Displays a Mosaic of Point and Regional Centromeres. Curr. Biol. 2019, 29, 3791–3802.e6. [Google Scholar] [CrossRef]
- Basu, D.; Kulkarni, R. Overview of blood components and their preparation. Indian J. Anaesth. 2014, 58, 529–537. [Google Scholar] [CrossRef]
- Salcedo-Hernandez, R.; Ruiz-Herrera, J. Isolation and characterization of a mycelial cytochrome aa3-deficient mutant and the role of mitochondria in dimorphism of Mucor rouxii. Exp. Mycol. 1993, 17, 142–154. [Google Scholar] [CrossRef]
- Valle-Maldonado, M.I.; Jácome-Galarza, I.E.; Gutiérrez-Corona, F.; Ramírez-Díaz, M.I.; Campos-García, J.; Meza-Carmen, V. Selection of reference genes for quantitative real time RT-PCR during dimorphism in the zygomycete Mucor circinelloides. Mol. Biol. Rep. 2015, 42, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans; WormBook: Pasadena, CA, USA, 2006; pp. 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Lin, Y.; Kemper, T.; Chen, J.; Yuan, Z.; Liu, S.; Zhu, Y.; Broering, R.; Lu, M. AMPK and Akt/mTOR signalling pathways participate in glucose-mediated regulation of hepatitis B virus replication and cellular autophagy. Cell Microbiol. 2020, 22, e13131. [Google Scholar] [CrossRef]
- Qian, W.; Wang, J.; Roginskaya, V.; McDermott, L.A.; Edwards, R.P.; Stolz, D.B.; Llambi, F.; Green, D.R.; Van Houten, B. Novel combination of mitochondrial division inhibitor 1 (mdivi-1) and platinum agents produces synergistic pro-apoptotic effect in drug resistant tumor cells. Oncotarget 2014, 5, 4180–4194. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, N.; Strippoli, V.; Cassone, A. Yeast-mycelial conversion induced by N-acetyl-D-glucosamine in Candida albicans. Nature 1974, 250, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Naseem, S.; Gunasekera, A.; Araya, E.; Konopka, J.B. N-acetylglucosamine (GlcNAc) induction of hyphal morphogenesis and transcriptional responses in Candida albicans are not dependent on its metabolism. J. Biol. Chem. 2011, 286, 28671–28680. [Google Scholar] [CrossRef]
- Aoki, S.; Ito-Kuwa, S.; Nakamura, Y.; Masuhara, T. Comparative pathogenicity of a wild-type strain and respiratory mutants of Candida albicans in mice. Zentralbl Bakteriol. 1990, 273, 332–343. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Li, B.; Chen, T.; Tian, S. Reactive oxygen species: A generalist in regulating development and pathogenicity of phytopathogenic fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3344–3349. [Google Scholar] [CrossRef]
- Dikalov, S. Cross talk between mitochondria and NADPH oxidases. Free Radic. Biol. Med. 2011, 51, 1289–1301. [Google Scholar] [CrossRef]
- Rossi, D.C.P.; Gleason, J.E.; Sanchez, H.; Schatzman, S.S.; Culbertson, E.M.; Johnson, C.J.; McNees, C.A.; Coelho, C.; Nett, J.E.; Andes, D.R.; et al. Candida albicans FRE8 encodes a member of the NADPH oxidase family that produces a burst of ROS during fungal morphogenesis. PLoS Pathog. 2017, 13, e1006763. [Google Scholar] [CrossRef] [PubMed]
- Nasution, O.; Srinivasa, K.; Kim, M.; Kim, Y.J.; Kim, W.; Jeong, W.; Choi, W. Hydrogen peroxide induces hyphal differentiation in Candida albicans. Eukaryot. Cell 2008, 7, 2008–2011. [Google Scholar] [CrossRef] [PubMed]
- Misslinger, M.; Hortschansky, P.; Brakhage, A.A.; Haas, H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118885. [Google Scholar] [CrossRef] [PubMed]
- Expert, D.; Boughammoura, A.; Franza, T. Siderophore-controlled iron assimilation in the enterobacterium Erwinia chrysanthemi: Evidence for the involvement of bacterioferritin and the Suf iron-sulfur cluster assembly machinery. J. Biol. Chem. 2008, 283, 36564–36572. [Google Scholar] [CrossRef] [PubMed]
- Kurakov, A.V.; Khidirov, K.S.; Sadykova, V.S.; Zviagintsev, D.G. Anaerobic growth ability and alcohol fermentation activity of microscopic fungi. Prikl. Biokhim Mikrobiol. 2011, 47, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Camarasa, C.; Grivet, J.P.; Dequin, S. Investigation by 13C-NMR and tricarboxylic acid (TCA) deletion mutant analysis of pathways for succinate formation in Saccharomyces cerevisiae during anaerobic fermentation. Microbiology 2003, 149, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Ackema, K.B.; Hench, J.; Böckler, S.; Wang, S.C.; Sauder, U.; Mergentaler, H.; Westermann, B.; Bard, F.; Frank, S.; Spang, A. The small GTPase Arf1 modulates mitochondrial morphology and function. EMBO J. 2014, 33, 2659–2675. [Google Scholar] [CrossRef]
- Hassan, M.I.A.; Voigt, K. Pathogenicity patterns of mucormycosis: Epidemiology, interaction with immune cells and virulence factors. Med. Mycol. 2019, 57 (Suppl. S2), S245–S256. [Google Scholar] [CrossRef]
- Rit, K.; Saha, R.; Dey, R.; Barik, G. Rhino-oculo-cerebral Aspergillus and Mucor co-infections in an immunocompromised patient with type 2 diabetes mellitus. Med. J. Dr. DY Patil Univ. 2014, 7, 486–488. [Google Scholar] [CrossRef]
- De Paepe, A.; Dams, K.; Robert, D.; Jacobs, R.; Ten Kate, G.L.; Van Ierssel, S.; Jansens, H.; Lammens, M.; Van Beeck, A.; Jorens, P.G. Two Cases of Post-Traumatic Mucormycosis due to Mucor circinelloides: Salvage Therapy with a Combination of Adjunctive Therapies. Case Rep. Infect. Dis. 2022, 4949426. [Google Scholar] [CrossRef]
- Renvall, S.; Niinikoski, J. Kinetics of oxygen in peritoneal cavity. Effects of chemical peritonitis and intraperitoneally administered colloids in rats. J. Surg. Res. 1980, 28, 132–139. [Google Scholar] [CrossRef]
- Zenewicz, L.A. Oxygen Levels and Immunological Studies. Front. Immunol. 2017, 21, 324. [Google Scholar] [CrossRef]
- Szebenyi, C.; Gu, Y.; Gebremariam, T.; Kocsubé, S.; Kiss-Vetráb, S.; Jáger, O.; Patai, R.; Spisák, K.; Sinka, R.; Binder, U.; et al. cotH Genes Are Necessary for Normal Spore Formation and Virulence in Mucor lusitanicus. mBio 2023, 14, e0338622. [Google Scholar] [CrossRef]
- Giles, S.S.; Batinic-Haberle, I.; Perfect, J.R.; Cox, G.M. Cryptococcus neoformans mitochondrial superoxide dismutase: An essential link between antioxidant function and high-temperature growth. Eukaryot. Cell 2005, 4, 46–54. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patiño-Medina, J.A.; Alejandre-Castañeda, V.; Valle-Maldonado, M.I.; Martínez-Pacheco, M.M.; Ruiz-Herrera, L.F.; Ramírez-Emiliano, J.; Ramírez-Marroquín, O.A.; Castro-Cerritos, K.V.; Campos-García, J.; Ramírez-Díaz, M.I.; et al. Blood Serum Stimulates the Virulence Potential of Mucorales through Enhancement in Mitochondrial Oxidative Metabolism and Rhizoferrin Production. J. Fungi 2023, 9, 1127. https://doi.org/10.3390/jof9121127
Patiño-Medina JA, Alejandre-Castañeda V, Valle-Maldonado MI, Martínez-Pacheco MM, Ruiz-Herrera LF, Ramírez-Emiliano J, Ramírez-Marroquín OA, Castro-Cerritos KV, Campos-García J, Ramírez-Díaz MI, et al. Blood Serum Stimulates the Virulence Potential of Mucorales through Enhancement in Mitochondrial Oxidative Metabolism and Rhizoferrin Production. Journal of Fungi. 2023; 9(12):1127. https://doi.org/10.3390/jof9121127
Chicago/Turabian StylePatiño-Medina, José Alberto, Viridiana Alejandre-Castañeda, Marco Iván Valle-Maldonado, Mauro Manuel Martínez-Pacheco, León Francisco Ruiz-Herrera, Joel Ramírez-Emiliano, Oscar Abelardo Ramírez-Marroquín, Karla Viridiana Castro-Cerritos, Jesús Campos-García, Martha Isela Ramírez-Díaz, and et al. 2023. "Blood Serum Stimulates the Virulence Potential of Mucorales through Enhancement in Mitochondrial Oxidative Metabolism and Rhizoferrin Production" Journal of Fungi 9, no. 12: 1127. https://doi.org/10.3390/jof9121127