
Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review

1
College of Science, Gansu Agricultural University, Lanzhou 730070, China
2
College of Food Science and Engineering, Gansu Agricultural University, Lanzhou 730070, China
3
Lanzhou Institutes for Food and Drug Control, Lanzhou 730070, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(8), 843; https://doi.org/10.3390/jof9080843
Submission received: 20 July 2023
/
Revised: 4 August 2023
/
Accepted: 8 August 2023
/
Published: 12 August 2023
(This article belongs to the Special Issue Plant Pathology and Mycotoxins)
Abstract
:Fusarium dry rot is one of the major potato diseases during storage after harvest, which not only results in quality degradation but also causes great economic losses. The disease can be elicited by some species of Fusarium, and the pathogenic fungi of Fusarium causing potato dry rot are considerably diverse in various countries and regions. The disease caused by Fusarium spp. is associated with mycotoxins accumulation, which has phytotoxic and mycotoxic effects on humans and animals. Chemical synthetic fungicide is considered the main control measure for the Fusarium dry rot of potato; nevertheless, it is unfortunate that persistent application inevitably results in the emergency of a resistant strain and environmental contamination. A comprehensive disease control strategy includes potato cultivar selection, appropriate cultural practices (crop rotation, cultivate pattern, fertilization, and irrigation), harvesting processes and postharvest treatments (harvesting, classification, packaging, wound healing), and storage conditions (environmental disinfection, temperature, humidity and gas composition) along with the application of fungicide pre-harvest or post-harvest. Recently, emerging studies have indicated that eco-friendly strategies include physical control, chemical methods (such as the application of generally-recognised-as-safe (GRAS) compounds or chemical (elicitors) and biological control have been introduced to combat the Fusarium dry rot of potato.
1. Introduction
Potato (Solanum tuberosum L.) ranks fourth as the most important food crop all over the world, only behind rice (Oryza sativa L.), wheat (Triticum aestivum L.), and maize (Zeamays L.). It is also a major non-cereal food crop, which plays an irreplaceable role in the global food supply. Potato is rich in plenty of macronutrients (such as carbohydrates and dietary fibre) and micronutrients (such as vitamins and minerals); furthermore, it is also an important source of antioxidants in people’s diet [1]. It is reported that, in 2020, the total world production of potatoes was 359 million tonness (http://www.fao.org/faostat/en/?#data/QC, accessed on 1 January 2021) and China ranked the first with the production of 17.98 million tons [2,3]. More than 85% of potatoes need to be kept for 3–6 months as a vegetable and industrial material, and the losses due to the disease during storage are very large [4]. Some fungi, bacteria, and viruses can result in post-harvest disease. Among them, the infection of potato tubers resulting from Fusarium spp. Can particularly cause severe dry rot during storage, which not only leads to quality deterioration but also reduces the marketable yield. Tuber losses range from 6.25% to 25% due to dry rot during storage annually, and up to 60% when potato tubers are injured [5]. In the Gansu province of China, it is estimated that tuber losses due to dry rot were around 88% of the total post-harvest losses [6]. Importantly, some Fusarium species associated with dry rot produce mycotoxins, which pose an adverse effect on humans and animals due to their mycotoxicoses.
Once the pathogenesis of Fusarium is known, appropriate and effective management measures can be carried out, and the losses will be effectively reduced. At present, some chemical synthetic fungicides (such as carbendazim, mancozeb, and thiabendazole (TBZ)) are employed to prevent and control the dry rot of potato. Nevertheless, chemical synthetic fungicides are not a long-term solution due to the ecological environment and drug resistance. Currently, some eco-friendly control measures such as the use of some organic acids and salts [7], inorganic salts [8,9], chitosan [10], plant essential oil [11,12] and biological antagonists [13,14] for the management of post-harvest disease in potato tubers are being explored and developed. The present review focuses on dry rot occurrence (including the causal pathogens of dry rot, the symptoms of dry rot, and its pathogenesis), mycotoxin production (non-trichothecenes and trichothecenes), and management strategies.
2. Dry Rot of Potato Tubers
2.1. The Causal Agent Causing Dry Rot
Fusarium is a notorious and large fungal genus within the Ascomycota phylum containing hundreds of species, which are primarily isolated from soil and plant survival [1]. It is well-known that Fusarium spp. have the ability to cause potato dry rot, which is a devastating form of post-harvest fungal decay, severely impacting potato tuber quality all over the world [1]. Fusarium dry rot causes a remarkable reduction in potato yield, as well as leading to enormous economic losses. Currently, there are 17 species, and 5 variants of Fusarium recognised globally as causal agents of potato dry rot [15]. Because of the differences in potato cultivars and climatic conditions, diverse Fusarium species were isolated and identified from the Fusarium dry rot of potato in various countries and regions. Among them, F. sambucinum was considered the most predominant pathogenic fungus leading to the Fusarium dry rot of potato in North America and some regions of Europe [16,17,18]; however, some reports suggested that F. solani var. coeruleum was regarded as the most prevalent pathogen causing potato dry rot under low-temperature storage in the United Kingdom, and occasionally, the pathogen of F. coeruleum also caused severe potato harvest disease in the United Kingdom [19,20,21]. F. graminearum was the most frequently Fusarium species caused potato dry rot in North Dakota [22]. F. oxysporum and F. solani were reported as the main pathogens to cause Fusarium dry rot in potato in South Africa [23,24]. F. sulphureum and F. solani were found to have a higher incidence and higher aggressiveness in Iran [25]. In Egypt, F. sambucinum was regarded as the most predominant pathogen, except for F. oxysporum, F. incarnatum and F. verticillioide [26] (Table 1).
There are different climatic conditions in China, and different species of Fusarium were isolated and characterised in different regions. Potato planting regions are divided into four planting regions in China, Northeast, North, Northwest, and South China. In the potato planting regions of Northwest China, F. avenaceum, F. oxysporum, F. sporotrichiodes, F. solani, F. trichothecioides, F. solani var. coeruleum, F. sambucinum, F. semitectum, F. solani, F. sambucinum, F. culmorum, F. gibbosum, F. macroceras, F. solani var. coeruleum, F. acuminatum, F. equiseti and F. redolens were identified in Heilongjiang Province and Inner Mongolia Autonomous Region [27]; in North China, F. sambucinum and F. avenaceum, F. solani var. coeruleum, F. oxysporum, and F. acuminatum were isolated in Shanxi Province [27]. In Northwest China, F. sambucinum, F. avenaceum, F. oxysporum, and F. equiseti were identified in Ningxia Hui Autonomous Region; F. sambucinum, F. solani, F. sulphureum, F. avenaceum, and F. graminearum were identified in Gansu Province [7,8,10,12]; F. tricinctum, F. avenaceum, F. oxysporum, F. solani, F. acuminatum, and F. equiseti were mainly identified in Qinghai Province [6]. In Zhejiang Province of South China, F. solani, F. solani var. coeruleum, F. moniliforme var. intermedium, F. moniliforme var. zhejiangense, and F. redolens were mainly identified [6] (Table 1).
The occurrence of potato dry rot is not only influenced by countries and regions; potato variety and chemical synthetic fungicide application, as well as seed tuber source also play significant roles. For instance, Xue’s research group [28] compared the pathogenicity of F. sulphureum for different potato cultivars, and found that the variety of Longshu No. 3 is susceptible, while the variety of Longshu No.6 is resistant for F. sulphureum.
2.2. Pathogen Infection and the Symptoms of Potato Dry Rot
In general, Fusarium spp. can infect potato tubers through surface wounds or natural openings on tubers during pre-harvest or post-harvest, and the process of Fusarium species infecting potato tubers is shown in Figure 1. Pathogenicity is a crucial factor when understanding the pathogens of the Fusarium infection of potato tubers. Additionally, extracellular enzymes and reactive oxygen species (ROS) play more important roles for the pathogenicity of Fusarium. Pathogens can secrete extracellular enzymes to destroy the cell wall and middle lamellar of the host plant, which makes the pathogen able to spread to the surrounding cell and successfully infect the host plant. Numerous studies indicated that cell wall-degrading enzymes (CWDEs) are important pathogenic factors for Fusarium when in infection and spread [29,30]. Yang et al. [29] suggested that the activities of CWDEs (such as polygalacturonase (PG), carboxymethyl cellulose (Cx), polymethyl-galacturonase (PMG), and so on) were found to increase during the F. sulphureum infection of potato tubers. Cutinase enzymes were also involved in pathogenicity when F. solani infected potato tubers. Moreover, ROS also play a vital role in the pathogenicity of the fungus. As we know, ROS production is an early evens during host-pathogen interactions, and excessive ROS can attack cellular biomolecules, such as lipids, proteins, and DNA, causing cell membrane damage to hosts via lipid peroxidation, and finally leading to infection. For instance, Bao et al. [31] compared the difference in pathogenicity between F. sambucinum and F. sulphureum during pathogens’ infection of potato tubers, and found that F. sulphureum showed higher pathogenicity in inoculated tubers in association with a higher ROS level, which caused a higher malondialdehyde (MDA) content, and a lower level for cell membrane integrity, ultimately leading to a bigger lesion diameter in the inoculated tubers. A contrasting result was observed in tubers inoculated with F. sambucinum due to the lower ROS accumulation. Xue et al. [28] also observed that, compared to F. sambucinum and F. solani, F. sulphureum manifested the strongest infection ability and pathogenicity in inoculated tubers (cv. Longshu No.3) in Gansu province. In addition, the pathogenicity of Fusarium was also related to host-nonspecific phytotoxin trichothecenes; for instance, fusaric acid produced by Fusarium destroyed the cell membrane structure of the host plant, then decreased the respiration rate, which is beneficial for the Fusarium infection of potato tubers.
The typical symptoms on the skin of the potato tubers infected by Fusarium spp. mainly include a wrinkled brown appearance, and sunken tissue with a dry and leathery appearance. The initial symptoms are observed to be shallow, small brown spots at tuber wound sites after approximately 30 days of storage. Subsequently, the infected tissue begins to become enlarged in every direction, and the tuber’s periderm gradually undergoes subsidence and collapses. Ultimately, concentric rings are observed on the enlarged lesions, and the dead tissue begins to be desiccated [1,32]. A cottony white, purple, yellow, pink, or brick colour of the spores and mycelia of Fusarium spp. are observed in the cavity under the rotted lesion [33]. As the disease progresses, whole tubers with symptoms of severe decay always have a shrivelled and dehydrated appearance. In severe cases, the affected potatoes may completely decay, resulting in a mushy texture and foul odour. Therefore, it is important to identify and manage potato dry rot to prevent its further spread and minimize economic losses.
2.3. Mycotoxin Accumulation Associated with Fusarium Dry Rot
Potato dry rot resulting from Fusarium is associated with mycotoxin accumulation. Mycotoxins are a type of secondary metabolite produced by toxigenic fungi under suitable temperature and humidity conditions, which can lead to a potential health threat to humans and animals [34,35]. The mycotoxin metabolised by Fusarium is classified into two types of non-trichothecenes and trichothecenes. The main non-trichothecenes metabolised by Fusarium spp. are shown in Table 2. Beauvericin (BEA) and enniatins (ENN) are cyclic hexadepsipeptides with antimicrobial, insecticidal, phytotoxic, and cytotoxic properties, which were detected in potato tubers infected with F. oxysporum [36]. Zearalenone (ZEA) and fusarin C (FUS) were detected in tubers infected with F. sambucinum, F. solani, and F. oxysporum, with oestrogenic syndromes in swine and other experimental animals [37,38,39,40]. Fumonisin (FUM) is linked to leukoencephalomalacia in brain lesions of horses and rabbits with hepatotoxic and carcinogenic influences, and is also associated with esophageal carcinoma in humans with phytotoxic effects, which was detected in potatoes contaminated with F. equiseti, F. sambucinum, and F. oxysporum [41]. El-Hassan and Kim [42,43,44] observed sambutoxin (SAM) in potato dry rot infected by F. sambucinum, F. semitectum, F. solani and F. oxysporum, which resulted in haemorrhage in the stomach and intestines, bodyweight loss, apastia, and death for rats. Sonja et al. [45] detected fusaric acid (FA) production in F. oxysporum-infected potato tubers, and Venter’s group [46] and El-Hassan [41] indicated that FA content was positively correlated with the incidence of dry rot. Pre- and post-harvest strategies were carried out to control FA accumulation during the dry rot of potato tubers [1].
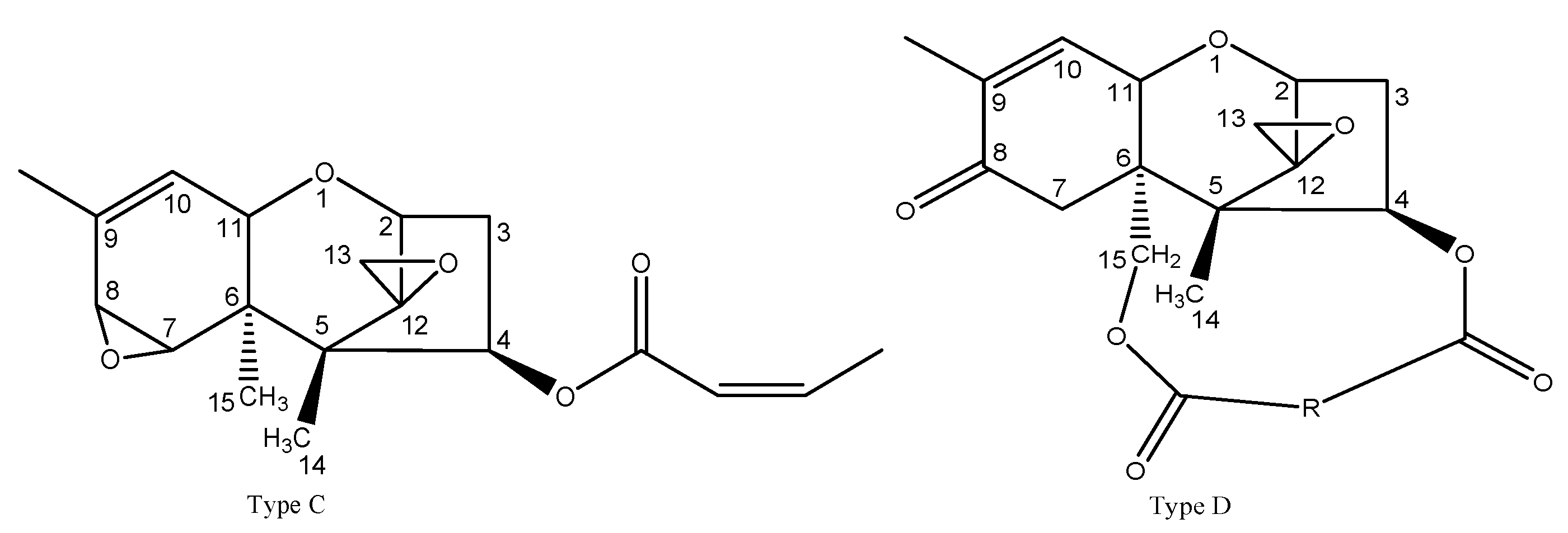
Trichothecenes are categorised as another main type of mycotoxin found in the Fusarium dry rot of potato that is a type of structurally related sesquiterpene compound. Up to the present, there are more than 190 known trichothecenes.
Trichothecenes are classified into four different types, A, B, C, and D, based on chemical structural differences; the chemical structures are listed in Figure 2.
Types A and B of trichothecenes are found in cereal crops and their contaminated products. Additionally, trichothecenes were found in potato dry rot [35], Fusarium dry rot in muskmelon [47], and core rot in apple [48]. As we know, Trichothecenes can pose a serious health threat to humans and animals due to phytotoxicity and mycotoxicoses [28]. For instance, in some severe cases, trichothecenes have potential carcinogenic, teratogenic, and mutagenic effects [49]. Trichothecenes were reported in the Fusarium dry rot of potato tubers (Table 3). Type A and B of trichothecenes are often mainly found in the lesion tissue of rotted potato tubers. Xue et al. [4] found the trichothecenes of 3-ADON, T-2, FUS, and DAS not only in the lesioned part but in the adjacent asymptomatic part of a potato with dry rot contaminated by F. sulphureum, F. solani, and F. sambucinum; it was interesting that the concentration of trichothecenes was negatively correlated with the distance from the infected point. Similarly, Ellner et al. [50] suggested that DAS was detected in rotten tissue, as well as in adjacent asymptomatic tissue in tubers contaminated by F. sambucinum, and a similar changing trend to that of Xue’s report was observed. Delgado et al. [51] suggested that DON, NIV, FX, 3-ADON, and 15-ADON were detected in potatoes inoculated with F. graminearum, and a similar trend to that in Xue’s and Ellner’s reports was found.
2.4. Dry Rot Control
Given the severity of potato dry rot, how to control the disease of dry rot has become an urgent question. Currently, using chemical synthetic fungicides such as thiabendazole, benzimidazole, 2-aminobutane, imazalil, flusilazole, and difenoconazole is the main strategy to control the disease. However, as we all know, a series of problems such as resistance against fungicides, environmental contamination, and pesticide residues have come up, which obliges scientists to develop integrated disease management strategies to combat the problems. An integrated disease control strategy includes potato cultivar selection, and appropriate cultural practices, harvesting processes and post-harvest treatments, and storage conditions along with the application of fungicide pre-harvest or post-harvest.
2.4.1. Variety Screening
The screening of resistant varieties plays a crucial role in controlling post-harvest disease. More than 5000 potato varieties were reported to be planted all over the world [61,62]. Most of the varieties are sensitive to Fusarium. Du’s research group investigated 21 potato varieties and 46 breeding lines against F. sambucinum, and found that 67 kinds of potato clones were sensitive to Fusarium in China [27]. Xue’ group [28] investigated the varieties of Longshu No.6 and Longshu No.3 against F. sulphureum and found that the variety of Longshu No.3 showed greater susceptibility to F. sulphureum, had more serious disease and higher levels of FUS, DAS, 3ADON and T-2 toxin in potato tubers contaminated by F. sulphureum compared to the variety of Longshu No. 6. In Tunisia, Trabelsi et al. [63] indicated that the varieties of Mondial, Spunta, and Liseta were less sensitive to F. sambucinum, F. oxysporum, and F. graminearum. In Iran, Esfahani et al. [64] screened 43 potato varieties to F. solani, F. sulphureum, and F. oxysporum, and found that only the variety of Saturna was resistant against the three fungi. In Canada, Yilma et al. [65] indicated that the variety of Owyhee Russet showed significantly higher resistance than that of Russet Burbank did to dry rot. In fact, resistance to every Fusarium spp. is mainly independent and genetically distinct to some extent. The resistance to a species of Fusarium is transmitted to progeny but appears to be associated with recessive alleles [66,67]. Even though numerous potato cultivars and clones were tested for sensitivity, no variety is resistant to all the Fusarium species. At the same time, varieties may be sensitive to one species of Fusarium; nevertheless, the resistance is to another species of Fusarium. Similarly, a certain strain of Fusarium maybe pathogenic to one cultivar, but non-pathogenicity is shown for another cultivar because the susceptibility–resistance outcome varies depending on the strains, the varieties, and the prevailing culture and environmental conditions in different regions of the world. Some studies pointed out the role of storage temperature for a cultivar’s susceptibility against Fusarium species. Mejdoub-Trabelsi et al. [68] found that cultivars at a temperature of 30 °C were less susceptible, while cultivars at a temperature of 15 °C were highly susceptible. Therefore, it is indispensable to study the populations of Fusarium in the field and their pathogenicity to optimize the growth of varieties in each field. At present, breeding resistant cultivars against dry rot is very efficient because of the laborious phenotyping involved [69]. The genome-editing technique using the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/Cas9 system to target genome modifications has become a potential and powerful tool for genetic engineering in potato [70]. The CRISPR/Cas9 system can provide an alternative strategy to that of conventional genetic engineering [71], and is expected to produce disease-resistant cultivars by designing and constructing gene-specific single-guide RNA (sgRNA) vectors.
2.4.2. Cultivation Patterns
Good cultivation patterns play a crucial role and influence the incidence and severity of storage disease after harvest. Generally, cultivation patterns include crop rotation, cultural methods, fertilization, and irrigation. Crop rotation is usually an advised cultural practice in controlling soil-borne disease; however, crop rotation is not very suitable in the management of potato dry rot [72,73]. Because the fungus has a broad host range and can survive in the soil for 5–6 years, it is very difficult to control infection via crop rotation. Potato crop rotation with barley and red clover did not achieve a significant effect for disease incidence and severity in 2–3 years [74].
Cultural methods play a key role in the management of potato dry rot. Dry rot is not only a soil disease but also a tuber-borne disease. Therefore, the seed tuber is usually considered the main source of inoculum [75]. Planting a healthy seed tuber in the field is necessary; planting an infected seed tuber will result in soil infestation around the progeny of the tuber [76]. The contaminated soil adhering to the tuber’s surface will eventually contaminate tubers through wounds or natural openings during storage after harvest (Figure 1). A reasonable sowing time is also an important factor for potatoes, and allows the whole growth period of a potato to have a relatively suitable temperature and humidity, avoiding high temperatures when a potato tuber develops and expands. Finally, reasonable fertilization and irrigation should be considered during different growth periods.
2.4.3. Harvesting, Grading and Packing
Harvesting and processing treatment after harvest significantly influence the control effect disease. Because the fungus of Fusarium spp. attacks potato tubers mainly through wounds, considerable efforts should be focused on avoiding tuber bruising and injuring when harvesting [15]. In addition, a temperature of 10–18 °C for tuber pulp is the best option for harvesting tubers [77]. The maturity level plays an important role when harvesting. Tubers with low maturity have a higher content of sucrose and a poorer skin set; however, a higher level of sucrose provides nutrition for fungus growth, and poorer skin is prone to bruising and the generation wounds. These properties lead to potatoes with low maturity that are more vulnerable to the fungus [18]. In general, it is appropriate for potato tubers to be harvested after 7–14 days of killing the potato vine, which is a sufficient amount of time for wound healing and decreases the chances of fungus attack [15]. It takes 1–2 weeks for A tuber to heal the wound when the environmental humidity is between 95 and 99%, and the tuber pulp temperature ranges from 13 to 16 °C, which is favours the rapid healing of a wound after harvest. Taking some steps to accelerate wound healing not only saves the wound healing time but also decreases labour costs. Our research group’s previous results suggested that sodium silicate or brassinosteroid treatment accelerated the wound healing process of tubers via the activation of phenylpropanoid metabolism [78,79]. Subsequently, Jiang adopted transcriptomics analysis of benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid s-methyl ester (BTH) to induce genes involved in suberin accumulation to accelerate the potato wound healing process [80]. Moreover, a careful examination of tuber grading, and packing should be paid attention to. As mentioned above, the disease of dry rot can easily contaminate potatoes through wounds; when one tuber decays, the rotten tuber will contaminate the tubers around it, which will ultimately result in a disastrous disease during storage. Therefore, tubers with wounds (including pests and disease appearance) must undergo a thorough examination and rejected before storage. Actually, the examination process requires proper grading before storage [81]. Finally, packing can not only reduce disease incidence, but can also increase the beauty and value of potatoes.
2.4.4. Storage Cellar Management
Storage cellar management is a crucial factor in managing the dry rot of potato, and includes storage cellar disinfection, and temperature, humidity, and gas components. During storage, proper disinfection treatment for storage facilities is mandatory, and the common chemical disinfectants include sulphur, potassium permanganate-formaldehyde, peracetic acid, chlorine dioxide, and 2–4% formalin solution. Most of these chemical disinfectants are employed for fumigation processing. Storage temperature is the determining factor of the storage quality of potato tubers. The storage temperature should be decreased to an appropriate level after wound healing at 15–20 °C. In general, the storage temperature is 2–3 °C for seed potatoes and 4–5 °C for commercial potatoes. During storage, the appropriate humidity is 80–93%, and higher humidity will lead to tuber rot and sprouting earlier. In addition, proper cool air circulation is also essential, because stored potato tubers produce excessive carbon dioxide (CO2) and heat, which will facilitate the adherence of Fusarium to spores [82].
2.4.5. Physical, Chemical, and Biological Treatment
Physical Treatment
As we know, with minimal environmental impacts and no residues in the treated product, the development of a physical application for the management of post-harvest plant diseases has been widely carried out. Among all kinds of physical treatments, the extensive application of ultraviolet-C light (UV-C, 190–280 nm) showed a significant effect; on the one hand, UV-C treatment can directly suppress pathogen growth, and on the other hand, UV-C application can induce a defence response in host tissues and increase resistance against disease [83]. The application of UV-C has been shown to control dry rot by inducing the generation of antifungal substances in potato that contribute to disease control [84]. Ranganna et al. [85] also suggested that UV-C irradiation completely prevents the development of dry rot caused by F. solani in potatoes under storage at 8 °C for 3 months. Yu et al. [86] indicated that 35 kJ·m−2 UV-C treatment significantly inhibited the dry rot of potato by increasing the activities of CAT, POD, and PAL. Another report was written by Jakubowski and Krolczyk [87], who suggested that UV-C radiation effectively controlled the disease of dry rot in stored potato tubers by inhibiting the development of fungi.
Chemical Treatment
The most effective strategy to control potato dry rot is the combination of pre- and post-harvest treatment. Firstly, disinfection the seed tuber before planting, and application of chemical treatments after planting plays an important role for management the dry rot. Thiabendazole is currently considered the most widely used chemical fungicide to manage the Fusarium dry rot of potato [88]. Thiophanate-methyl (benzimidazole group) was reported to be extensively applied to manage seed tuber piece disease in Canada. Nevertheless, the employment of thiabendazole has resulted in the occurrence of the drug-resistant strains against the pathogen of F. sambucinum, and it is fortunate that the rest of the Fusarium species viz. F. solani, F. culmorum, F. equiseti, F. acuminatum and F. avenaceum are still sensitive to the fungicide of thiabendazole [72,89]. Some alternative fungicides with high efficiency and low toxicity (such as fludioxonil) also have incomparable effects in managing dry rot. For instance, fludioxonil was used to effectively control tuber seed disease and sprout rot [90]. The application of azoxystrobin and fludioxonil effectively managed dry rot, and the disease incidence decreased to 50% comparing to that of the control after 21 days of storage [15]. However, with the extensive application of synthesised chemical fungicides, the inevitable problems of drug resistance and environmental pollution, as well as food safety are becoming more and more prominent. Therefore, it is an urgent need to develop more safe and efficient fungicides to control the Fusarium dry rot of potato.
Accordingly, generally-recognised-as-safe (GRAS) substances, such as inorganic acids, organic acids, inorganic salts, organic salts, essential oils and phytohormones, all display excellent effects in terms of sustainably controlling the dry rot of potato. Raigond et al. [91] indicated that chitosan application significantly managed dry rot in potato, and he also found that a chitosan coating significantly reduced Fusarium incidence by inhibiting Fusarium growth. Xue et al. [7] suggested that chitosan, sodium silicate and β-aminobutyric acid treatments markedly inhibited the expansion of lesion diameter in tubers infected with F. sulphureum. Interestingly, the trichothecene concentration was also decreased; the involved action mechanism was attributed to the up-regulation of enzyme activities involved in the defence response, and down-regulated genes were related to the trichothecene biosynthesis pathway. Afterwards, Xue’s research group found an interesting result that T-2 toxin, as a kind of trichothecene A, suppressed the spread of dry rot of potato at a low concentration [92]. Later, Han found that the treatments of sodium silicate and brassinosteroid respectively promoted the wound healing of potato and accelerated suberin deposition, ultimately enhancing resistance against the fungi [78,79]. Jiang adopted BTH to treat potato and also observed a similar control effect on potato dry rot, with accelerated wound healing [80]. Ma et al. [93] found that the gene StCDPK23 played an important role in the wound healing of potato and suberin deposition, and constructed StCDPK23-overexpressing plants to conform to StCDPK23 to participate in tuber wound healing and contribute to resistance against the dry rot of potato.
In addition, essential oils and extracts from plants display an excellent effect in that they suppress the development of Fusarium dry rot via soaking or fumigation treatment [94]. The essential oil of Zanthoxylum bungeanum was found to be efficient in inhibiting the expansion of dry rot disease resulting from F. sulphureum [11]. Cinnamaldehyde, a major component of cinnamon essential oil, displayed a better control effect on potato dry rot resulting from F. sambucinum; the underlying mechanism revealed that cinnamaldehyde suppressed spore germination by impacting the biosynthetic pathway of ergosterol, improving ROS accumulation, and ultimately resulting in a breakdown of cell membrane integrity [12]. Similarly, the essential oils from peppermint and fennel also remarkedly suppressed the growth of F. oxysporum in vitro, and inhibited the development of potato dry rot when treated with a protective emulsifiable concentrate [95]. It is interesting that some essential oils directly influenced mycotoxin metabolism by impacting the biosynthetic pathway of mycotoxins; for instance, the essential oils from palmarose and clove reduced DON and ZEA accumulation by down-regulating the expression of genes involved in mycotoxicity in the biosynthetic pathway of DON and ZEA [96]. Essential oils from plants, as a sustainable alternative to chemical synthetic fungicides, need to be studied in-depth in the future [1,97]. Plant extracts also display excellent effects on plant disease [98]. The extract from black spruce revealed antifungal and suppressive potential to prevent the development of potato dry rot [99]. The extract from cinnamon also significantly inhibited F. sambucinum spore growth in vitro and reduced dry rot development in vivo [100]. Chlorogenic acid, as a kind of polyphenol with antioxidative activity, is mainly sourced from methanol extract, and also displayed better inhibitory activity on the development of Fusarium dry rot of potato. The possible action mechanism is attributed to the alteration of the morphological structure of F. sambucinum after chlorogenic acid application, and the changing of curling, twisting and collapse were observed after exposure to chlorogenic acid [1].
Additionally, chlorine dioxide and ozone, as the two strong oxidants, also perform important and crucial roles in controlling Fusarium dry rot. Chlorine dioxide (ClO2) acts as a water-soluble strong oxidant, whose oxidation ability is 2.5 times higher than that of chlorine. ClO2 can be applied in both a gaseous form and as an aqueous solution to control post-harvest disease in fruits and vegetables. For instance, Li et al. [101] suggested that the application of 0.75 ug/mL of ClO2 solution significantly reduced the incidence of Fusarium dry rot of potato and suppressed the expansion of the lesion diameter by damaging the morphology and ultrastructure of F. sulphureun hyphae. Ozone, as another high-efficiency and non-toxic, strong oxidant, also display an important role in controlling potato dry rot; on the one hand, ozone inhibited the growth of F. sulphureum spores, and destroyed the structure of F. sulphureum [102], while on the other hand, ozone treatment activated the ROS metabolism of potato tubers, and induced resistance against dry rot [103].
In fact, the possible action mechanism of chemical treatment was attributed to two facts: on the one hand, chemical treatments inhibit the growth of pathogens; on the other hand, chemical treatments induce resistance against the dry rot of potato tubers (Figure 3).
Biological Treatment
Biocontrol is regarded as a greener and safer strategy for food safety and human health, comparing to traditional chemical synthetic fungicides. Presently, scientists have been focusing on research on antagonistic microorganisms to manage plant diseases. Antagonistic microorganisms currently regarded as the best potential alternative option with which to manage post-harvest diseases. For instance, antagonistic microorganisms effectively controlled Fusarium dry rot during the potato wound-healing process when tubers were at their most vulnerable. Schisler’s group firstly reported the strains of Pseudomonas Migula spp., Enterobacter Hormaeche and Edwards spp., and Pantoea Gavini. spp. remarkedly decreased the incidence of potato dry rot resulting from F. sambucinum [104]. Later, the group found that mixtures of various antagonist strains were more efficient in controlling potato dry rot than a single strain [105]. Gözdenur and Elif [106] screened 12 bacterial and fungal isolates and found that Pantoea agglomerans had the best efficacy in suppressing the growth of F. oxysporum and in controlling the occurrence of potato dry rot.
Trichoderma harzianum and Bacillus subtilis, as the two most important biological control agents, were registered to manage potato disease. The two agents were the most studied mycoparasitic species for their antagonistic function against a broad spectrum of pathogenic fungi, as well as being recognised as the most promising strategies to inhibit different kinds of pathogenic fungus growth and to control both pre-harvest and post-harvest plant diseases occurrence. Daami-Remadi et al. [107] suggested that T. harzianum and T. viride manifested greater antagonistic activity against Fusarium dry rot in potato in Tunisia. El-Kot [108] compared four strains of fungal, bacterial and bioagent actinomycetes, and suggested that T. harzianum displayed the best efficiency in inhibiting the radial development of F. sambucinum and controlling the occurrence of dry rot in a greenhouse. Paul et al. [109] also observed that the T. harzianum strains CMML20-26 and CMML20-27 significantly decreased post-harvest disease in sweet potato. B. subtilis also plays a vital role in reducing potato disease. Wharton and Kirk [110] used the bioagent of B. subtilis in combination with excellent management practices to significantly reduce seed piece decay by 94.3% in 2007. Hussain et al. [111] compared the biosurfactant extract, culture filtrate and bacterial cell suspension from B. subtilis HussainT-AMU and found that the bacterial cell suspension (49%) and biosurfactant extract (70%) had the most control effects on the net house and field, respectively.
In recent years, Trichothecium roseum was also reported to act as an elicitor to induce resistance against dry rot in potato tubers infected by F. sulphureum. During the defence responses induced by the elicitor, the genes involved in a resistant reaction were up-regulated; accordingly, the enzymes activities and antifungal compound contents were also significantly decreased after T. roseum application [112]. The possible mode of action for the bioagent includes mycoparasitism, competition for nutrients and spaces, and the production of extracellular enzymes and/or secondary metabolites (such as antibiotic compounds and mycotoxins) (Figure 3) [113]. For instance, Xue’s research group [92] indicated that T-2 toxin (secondary metabolites from F. sulphureum) at the concentration of 1µg/mL could act as an elicitor to induce resistance against dry rot by activating ROS metabolism and phenylpropane metabolism in potato.
3. Conclusions
Fusarium dry rot of potato is regarded as a major form of disastrous potato decay that damages tuber quality and causes economic losses and mycotoxin contamination. There are 17 species and 5 variants of Fusarium causing potato dry rot all over the world, and the changes in genetic diversity depend on geographical locations. The frequency of occurrence and aggressiveness of Fusarium dry rot also differ depending on the prevalent variety and ambient conditions in a location-specific manner. Because the susceptibility or resistance of a particular cultivar is related to Fusarium species and storage temperature, a breeding project urgently needs to be designed, for different cultivars to adapt against Fusarium species.
To efficiently control the occurrence of the Fusarium dry rot of potato, an integrated disease-controlling strategy is recommended that includes excellent harvesting conditions to avoid tuber injury and suitable storage conditions (optimum temperature, humidity, and CO2 concentration), as well as planting seed tubers free of visible disease, and registered chemical synthetic fungicide and/or post-harvest GRAS treatment. Fusarium dry rot-controlling strategies eventually integrate the application of alternatives such as GRAS and microbial antagonists. Efficient strategies to manage Fusarium dry rot mainly depend on further research such as a combination of gene editing and molecular breeding, as well as development efforts between scientists and industry to implement an integrated management measure towards the high-efficiency control of the Fusarium dry rot of potato.
Author Contributions
Writing original draft: H.X. Writing—review and editing: H.X., Q.L. and Z.Y. Project administration and supervision: H.X. project management: Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the youth supervisor supporting fund (grant no. (GAU-QDFC-2021–09)), and National Natural Science Foundation of China (grant no. (32060566)).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, J.; Sun, Z.Q.; Zou, Y.P.; Li, W.H.; He, F.Y.; Huang, X.Y.; Lin, C.L.; Cai, Q.N.; Wisniewski, M.; Wu, X.H. Pre- and postharvest measures used to control decay and mycotoxigenic fungi in potato (Solanum tuberosum L.) during storage. Crit. Rev. Food Sci. Nutr. 2022, 62, 415–428. [Google Scholar] [CrossRef]
- Li, L.; Zhu, T.; Song, Y.; Feng, L.; Kear, P.J.; Riseh, R.S.; Sitohy, M.; Datla, R.; Ren, M. Salicylic acid fights against Fusarium wilt by inhibiting target of rapamycin signaling pathway in Fusarium oxysporum. J. Adv. Res. 2022, 39, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Xia, X.; Zhao, Q.; Dong, P. The biocontrol of potato dry rot by microorganisms and bioactive substances: A review. Physiol. Mol. Plant Pathol. 2022, 122, 101919. [Google Scholar] [CrossRef]
- Xue, H.L.; Bi, Y.; Wei, J.M.; Tang, Y.M.; Zhao, Y.; Wang, Y. A new method for the simultaneous analysis of types A and B trichothecenes by ultrahigh-performance liquid chromatography coupled with tandem mass spectrometry in potato tubers inoculated with Fusarium sulphureum. J. Agric. Food Chem. 2013, 61, 9333–9338. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhang, W.; Kang, Y.; Shi, M.; Yang, X.; Yu, H.; Zhang, R.; Liu, Y.; Qin, S. Physiological and dynamic transcriptome analysis of two potato varieties reveal response of lignin and MAPK signal to dry rot caused by Fusarium sulphureum. Sci. Hortic. 2021, 289, 110470. [Google Scholar] [CrossRef]
- Wang, W.Z.; Min, F.X.; Yang, S.; Wei, Q.; Guo, M.; Gao, Y.F.; Hu, L.S.; Sheng, W.M. Research progress on potato dry rot disease in China and its control measures. China Veg. 2020, 4, 22–29. [Google Scholar]
- Xue, H.L.; Bi, Y.; Zong, Y.Y.; Alejandro, C.U.; Wang, H.J.; Pu, L.M.; Wang, Y.; Li, Y.C. Effects of elicitors on trichothecene accumulation and Tri genes expression in potato tubers inoculated with Fusarium sulphureum. Eur. J. Plant Pathol. 2017, 148, 673–685. [Google Scholar]
- Li, Y.C.; Bi, Y.; Ge, Y.H.; Sun, X.J.; Wang, Y. Antifungal activity of sodium silicate on Fusarium sulphureum and its effect on dry rot of potato tubers. J. Food Sci. 2009, 74, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Yaganza, E.S.; Tweddell, R.J.; Arul, J. Postharvest application of organic and inorganic salts to control potato (Solanum tuberosum L.) storage soft rot: Plant tissue-salt physicochemical interactions. J. Agric. Food Chem. 2014, 62, 9223–9231. [Google Scholar] [CrossRef]
- Li, Y.C.; Sun, X.J.; Bi, Y.; Ge, Y.H.; Wang, Y. Antifungal activity of chitosan on Fusarium sulphureum in relation to dry rot of potato tuber. Agric. Sci. China 2009, 8, 597–604. [Google Scholar] [CrossRef]
- Li, X.D.; Xue, H.L. Antifungal activity of the essential oil of Zanthoxylum bungeanum and its major constituent on Fusarium sulphureum and dry rot of potato tubers. Phytoparasitica 2014, 42, 509–517. [Google Scholar]
- Wei, J.; Bi, Y.; Xue, H. Antifungal activity of cinnamaldehyde against Fusarium sambucinum involves inhibition of ergosterol biosynthesis. J. Appl. Microbiol. 2020, 129, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Al-Mughrabi, K.I.; Vikram, A.; Peters, R.D.; Howard, R.J.; Grant, T.L.; Barasubiye, K.; Lynch, R.; Poirier, K.A.; Drake, I.K. Efficacy of Pseudomonas syringae in the management of critical reviews in food science and nutrition potato tuber diseases in storage. Biol. Control 2013, 64, 315–322. [Google Scholar] [CrossRef]
- Aydin, M.H. Evaluation of some Trichoderma species in biological control of potato dry rot caused by Fusarium sambucinum fuckel isolates. Appl. Ecol. Environ. Res. 2019, 17, 533–546. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. Biotechnology 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Ray, H.; Hammerschmidt, R. Responses of potato tuber to infection by Fusarium sambucinum. Physiol. Mol. Plant Pathol. 1998, 53, 82–91. [Google Scholar] [CrossRef]
- Wharton, P.S.; Tumbalam, P.; Kirk, W.W. First report of potato tuber sprout rot caused by Fusarium sambucinum in Michigan. Plant Dis. 2006, 90, 1460. [Google Scholar] [CrossRef] [PubMed]
- Heltoft, P.; Molteberg, E.L.; Nastad, R.; Hermansen, A. Effect of maturity level and potato cultivar on development of Fusarium dry rot in Norway. Potato Res. 2015, 58, 205–219. [Google Scholar] [CrossRef]
- Oosterhaven, K.; Chambel Leitao, A.; Gorris, L.G.M.; Smid, E.J. Comparative study on the action of S-(+)-carvone, in situ, on the potato storage fungi Fusarium solani var. coeruleum and F. sulphureum. J. Appl. Bacteriol. 1996, 80, 535–539. [Google Scholar] [CrossRef]
- Heltoft, P.; Brurberg, M.B.; Skogen, M.; Le, V.H.; Razzaghian, J.; Hermansen, A. Fusarium spp. causing dry rot on potatoes in Norway and development of a real-time PCR method for detection of Fusarium coeruleum. Potato Res. 2016, 59, 67–80. [Google Scholar] [CrossRef]
- Carnegie, S.F.; Cameron, A.M. Occurrence of Polyscytalum pustulans, Phoma foveata and Fusarium solani var. coeruleum in field soils in Scotland. Plant Pathol. 1990, 39, 517–523. [Google Scholar] [CrossRef]
- Ali, S.; Rivera, V.V.; Secor, G.A. First Report of Fusarium graminearum causing dry rot of potato in North Dakota. Plant Dis. 2005, 89, 105. [Google Scholar] [CrossRef] [PubMed]
- Villarino, M.; Cal, A.D.; Melgarejo, P.; Larena, I. Development of a multiplex PCR for the identification of Fusarium solani and F. oxysporum in a single step. J. Plant Dis. Prot. 2021, 128, 1275–1290. [Google Scholar] [CrossRef]
- Theron, D.J.; Holz, G. Effect of temperature on dry rot development of potato tubers inoculated with different Fusarium spp. Potato Res. 1990, 33, 109–117. [Google Scholar] [CrossRef]
- Esfahani, M.N. Present status of Fusarium dry rot of potato tubers in Isfahan. Indian Phytopathol. 2012, 59, 2. [Google Scholar]
- Gherbawy, Y.A.; Hussein, M.A.; El-dawy, E.G.A. Identification of Fusarium spp. associated with potato tubers in upper Egypt by morphological and molecular characters. Asian J. Biochem. Genet. Mol. Biol. 2019, 2, 1–14. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. causing potato dry rot in China and susceptibility evaluation of Chinese potato germplasm to the pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.L.; Bi, Y.; Tang, Y.M.; Zhao, Y.; Wang, Y. Effect of cultivars, Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers. Food Chem. 2014, 151, 236–242. [Google Scholar] [CrossRef]
- Yang, Z.M.; Bi, Y.; Li, Y.C.; Kou, Z.H.; Bao, G.H.; Liu, C.K.; Wang, Y.; Wang, D. Changes of cell wall degrading enzymes in potato tuber tissue slices infected by Fusarium sulphureum. Sci. Agric. Sin. 2012, 45, 127–134. [Google Scholar]
- Chai, Y.; Xu, Y.Q.; Fu, Y.; Li, X.Y.; He, F.M.; Han, Y.Q.; Feng, Z.; Li, F.L. Characteristics of cell wall degradation enzyme produced by main pathogenic Fusarium spp. in potato dry rot. Crops 2018, 4, 154–160. [Google Scholar]
- Bao, G.H.; Bi, Y.; Li, Y.C.; Kou, Z.H.; Hu, L.G.; Ge, Y.H.; Wang, Y.; Wang, D. Overproduction of reactive oxygen species involved in the pathogenicity of Fusarium in potato tubers, Physiol. Mol. Plant Pathol. 2014, 86, 35–42. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wu, C.Y.; Hua, H.H.; Cai, Q.N.; Wu, X.H. Characterization of the first Alternavirus identified in Fusarium avenaceum, the causal agent of potato dry rot. Viruses 2023, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Vatankhah, M.; Saberi Riseh, R.; Moradzadeh Eskandari, M.; Sedaghati, E.; Alaie, H.; Afzali, H. Biological control of Fusarium dry rot of potato using some probiotic bacteria. J. Agric. Sci. Technol. 2019, 21, 1301–1312. [Google Scholar]
- Schultz, B.; Ellner, F.M.; Gossmann, M.; Buettner, C. Investigation into virulence and mycotoxin formation of the dry rot causing pathogen Fusarium sambucinum on potatoes. Mycotoxin Res. 2008, 23, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, H.L.; Bi, Y.; Zhang, R.; Carelle, J.K.; Liu, Q.L.; Nan, M.N.; Pu, L.P.; Dov, P. Ozone treatment inhibits dry rot development and diacetoxyscirpenol accumulation in inoculated potato tuber by influencing growth of Fusarium sulphureum and ergosterol biosynthesis. Postharvest Biol. Technol. 2022, 185, 111796. [Google Scholar] [CrossRef]
- Song, H.H.; Lee, H.S.; Jeong, J.H.; Park, H.S.; Lee, C. Diversity in beauvericin and enniatins H, I, and MK1688 by Fusarium oxysporum isolated from potato. Int. J. Food Microbiol. 2006, 122, 296–301. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Diversity of Fusarium spp. associated with dry rot of potato tubers in Poland. Eur. J. Plant Pathol. 2016, 145, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Golinski, P.; Vesonder, R.F.; Latus-Zietkiewicz, D.; Perkowski, J. Formation of fusarenone X, nivalenol, zearalenone, alpha-trans-zearalenol, beta-trans-zearalenol, and fusarin C by Fusarium crookwellense. Appl. Environ. Microbiol. 1998, 54, 2147–2148. [Google Scholar] [CrossRef]
- Latus-Zietkiewicz, D.; Perkowski, J.; Tanaka, T.; Yamamoto, S.; Kawamura, O.; Sugiura, Y.; Ueno, Y. Formation of trichothecenes and zearalenone by Fusarium isolated from potato tubers. Microbiol. Aliment. Nutr. 1990, 8, 143–147. [Google Scholar]
- Sydenham, E.W.; Marasas, W.F.O.; Thiel, P.G.; Shephard, G.S.; Nieuwenhuis, J.J. Production of mycotoxins by selected Fusarium graminearum and F. crookwellense isolates. Food Addit. Contam. 1991, 8, 31–41. [Google Scholar] [CrossRef]
- El-Hassan, K.I.; El-Saman, M.G.; Mosa, A.A.; Mostafa, M.H. Variation among Fusarium spp. the causal of potato tuber dry rot in their pathogenicity and mycotoxins production. Egypt. J. Phytopathol. 2007, 35, 53–68. [Google Scholar]
- Kim, J.C.; Lee, Y.W. Sambutoxin, a new mycotoxin produced by toxic Fusarium isolates obtained from rotted potato tuber. Appl. Environ. Microbiol. 1994, 60, 4380–4386. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Lee, Y.W.; Tamura, H.; Yoshizawa, T. Sambutoxin: A new mycotoxin isolated from Fusarium sambucinum. Tetrahedron Lett. 1995, 36, 1047–1050. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, Y.W.; Yu, S.H. Sambutoxin-producing isolates of Fusarium species and occurrence of sambutoxin in rotten potato tubers. Appl. Environ. Microbiol. 1995, 61, 3750–3751. [Google Scholar] [CrossRef] [PubMed]
- Venter, S.L.; Steyn, P.J.; Hester, S.F. Production of fusaric acid by Fusarium oxysporum. Potato Res. 1996, 39, 79–83. [Google Scholar] [CrossRef]
- Venter, S.L.; Steyn, P.J. Correlation between fusaric acid production and virulence of isolates of Fusarium oxysporum that causes potato dry rot in South Africa. Potato Res. 1998, 41, 289–294. [Google Scholar] [CrossRef]
- Zhang, R.; Xue, H.L.; Li, L.; Bi, Y.; Zong, Y.Y.; Jimdjio, K.C. Study on the adsorption effect of diatomite on neosolaniol (NEO) in muskmelon fruits inoculated with Fusarium sulphureum. Int. J. Food Eng. 2021, 17, 257–264. [Google Scholar] [CrossRef]
- Tang, Y.M.; Xue, H.L.; Bi, Y.; Li, Y.C.; Wang, Y.; Zhao, Y.; Shen, K.P. A method of analysis for T-2 toxin and neosolaniol by UPLC-MS/MS in apple fruit inoculated with Trichothecium roseum. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Neves, M.; Puel, S.; Lippi, Y.; Soler, L.; Miller, J.D.; Oswald, I.P. Intestinal toxicity of the new type A trichothecenes, NX and 3ANX. Chemosphere 2022, 288, 132415. [Google Scholar] [CrossRef] [PubMed]
- Ellner, F.M. Mycotoxins in potato tubers infected by Fusarium sambucinum. Mycotoxin Res. 2002, 18, 57–61. [Google Scholar] [CrossRef]
- Delgado, J.A.; Schwarz, P.B.; Gillespie, J.; Rivera-Varas, V.V.; Secor, G.A. Trichothecene mycotoxins associated with potato dry rot caused by Fusarium graminearum. Phytopathology 2010, 100, 290–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, A.E.; Plattner, R.D. Trichothecene toxin production by strains of Gibberella pulicaris (Fusarium sambucinum) in liquid culture and in potato tubers. J. Agric. Food Chem. 1989, 37, 388–392. [Google Scholar] [CrossRef]
- Jelen, H.H.; Mirocha, C.J.; Wasowicz, E.; Kaminski, E. Production of volatile sesquiterpenes by Fusarium sambucinum strains with different abilities to synthesize trichothecenes. Appl. Environ. Microbiol. 1995, 61, 3815–3820. [Google Scholar] [CrossRef]
- El-Banna, A.A.; Scott, P.M.; Lau, P.Y.; Sakuma, T.; Platt, H.W.; Campbell, V. Formation of trichothecenes by Fusarium solani var. coeruleum and Fusarium sambucinum in potatoes. Appl. Environ. Microbiol. 1984, 47, 1169–1171. [Google Scholar] [CrossRef]
- Lenc, L.; Lukanowski, A.; Sadowski, C. The use of PCR amplification in determining the toxigenic potential of Fusarium sambucinum and F. solani isolated from potato tubers with symptoms of dry rot. Phytopathol. Pol. 2008, 2008, 48. [Google Scholar]
- Vesonder, R.F.; Golinski, P.; Plattner, R.; Zietkiewicz, D.L. Mycotoxin formation by different geographic isolates of Fusarium crookwellense. Mycopathologia 1991, 113, 11–14. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Thrane, U. Fast methods for screening of trichothecenes in fungal cultures using gas chromatography–tandem mass spectrometry. J. Chromatogr. A 2001, 929, 75–87. [Google Scholar] [CrossRef]
- Burlakoti, R.R.; Estrada, R.; Rivera, V.V.; Boddeda, A.; Secor, G.A.; Adhikari, T.B. Real-time PCR quantification and mycotoxin production of Fusarium graminearum in wheat inoculated with isolates collected from potato, sugar beet, and wheat. Phytopathology 2007, 97, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Burlakoti, R.R.; Ali, S.; Secor, G.A.; Neate, S.M.; McMullen, M.P.; Adhikari, T.B. Genetic relationships among populations of Gibberella zeae from barley, wheat, potato, and sugar beet in the Upper Midwest of the United States. Phytopathology 2008, 98, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Latus-Zietkiewicz, D.; Perkowski, J.; Chełkowski, J. Fusarium species as pathogens of potato tubers during storage and their ability to produce mycotoxins. Mycotoxin Res. 1987, 3, 99–104. [Google Scholar] [CrossRef]
- Shams, M.; Mitterbauer, R.; Corradini, R.; Wiesenberger, G.; Dall’Asta, C.; Schuhmacher, R.; Krska, R.; Adam, G.; Berthiller, F. Isolation and characterization of a new less-toxic derivative of the Fusarium mycotoxin diacetoxyscirpenol after thermal treatment. J. Agric. Food Chem. 2011, 59, 9709–9714. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, K.; Akhtar, M.H. Potato production, usage, and Nutrition—A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, B.M.; Abdallah, R.A.B.; Ammar, N.; Kthiri, Z.; Hamada, W. Bio-suppression of Fusarium wilt disease in potato using nonpathogenic potatoassociated fungi. J. Plant Pathol. Microbiol. 2016, 7, 347–354. [Google Scholar]
- Esfahani, M.N. Susceptibility assessment of potato cultivars to Fusarium dry rot species. Potato Res. 2005, 48, 215–226. [Google Scholar] [CrossRef]
- Yilma, S.; Vales, M.I.; Charlton, B.A.; Hane, D.C.; James, S.R.; Shock, C.C.; Mosley, A.R.; Culp, D.; Feibert, E.; Leroux, L.; et al. Owyhee Russet: A variety with high yields of U.S. No. 1 tubers, excellent processing quality, and moderate resistance to Fusarium dry rot (Fusarium solani var. coeruleum). Am. J. Potato Res. 2012, 89, 175–183. [Google Scholar] [CrossRef]
- Valluru, R.; Christ, B.J.; Haynes, K.G.; Vinyard, B.T. Inheritance and stability of resistance to Fusarium tuber rot in tetraploid potatoes. Am. J. Potato Res. 2006, 83, 335–341. [Google Scholar] [CrossRef]
- Burkhart, C.R.; Christ, B.J.; Haynes, K.G. Non-additive genetic variance governs resistance to Fusarium dry rot in a diploid hybrid potato population. Am. J. Potato Res. 2007, 84, 199–204. [Google Scholar] [CrossRef]
- Mejdoub-Trabelsi, B.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Effect of Fusarium species and temperature of storage on the susceptibility ranking of potato cultivars to tuber dry rot biocontrol of soilborne fungal diseases of vegetable crops. Pest Technol. 2012, 6, 41–46. [Google Scholar]
- Chen, D.; Nahar, K.; Bizimungu, B. A simple and efficient inoculation method for Fusarium dry rot evaluations in potatoes. Am. J. Potato Res. 2020, 97, 265–271. [Google Scholar] [CrossRef]
- Nadakuduti, S.S.; Starker, C.G.; Voytas, D.F.; Buell, C.R.; Douches, D.S. Genome editing in potato with CRISPR/Cas9. Methods Mol. Biol. 2019, 1917, 183–201. [Google Scholar]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.P.; Guyon-Debast, A.; Chauvin, J.E.; Nogue, F.; Mazier, M. Transgene-free genome editing in tomato and potato plants using agrobacterium-mediated delivery of a CRISPR/Cas9 cytidine base editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [Green Version]
- Bojanowski, A.; Avis, T.J.; Pelletier, S.; Tweddell, R.J. Management of potato dry rot. Postharvest Biol. Technol. 2013, 84, 99–109. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S. Continuous and emerging challenges of silver scurf disease in potato. Int. J. Pest Manag. 2021, 68, 89–101. [Google Scholar] [CrossRef]
- Qin, J.H.; Bian, C.S.; Duan, S.G.; Wang, W.X.; Li, G.C.; Jin, L.P. Effects of different rotation cropping systems on potato yield, rhizosphere microbial community and soil biochemical properties. Front. Plant Sci. 2022, 13, 999730. [Google Scholar] [CrossRef] [PubMed]
- Kwambai, T.K.; Griffin, D.; Nyongesa, M.; Byrne, S.; Gorman, M.; Struik, P.C. Dormancy and physiological age of seed tubers from a diverse set of potato cultivars grown at different altitudes and in different seasons in Kenya. Potato Res. 2023, 23, 1–25. [Google Scholar] [CrossRef]
- Zhou, H.L.; Lei, Y.; Wang, P.; Liu, M.Y.; Hu, X.X. Development of SYBR Green real-time PCR and nested RT-PCR for the detection of Potato Mop-top Virus (PMTV) and viral surveys in Progeny tubers derived from PMTV infected Potato tubers. Mol. Cell. Probes 2019, 47, 101438. [Google Scholar] [CrossRef]
- Knowles, N.R.; Plissey, E.S. Maintaining tuber health during harvest, storage, and post-storage handling. In Potato Health Management; Johnson, D.A., Ed.; APS Press: St. Paul, MN, USA, 2008; pp. 79–99. [Google Scholar]
- Han, Y.; Yang, R.R.; Wang, Q.H.; Wang, B.; Prusky, D. Sodium silicate promotes wound healing by inducing the deposition of suberin polyphenolic and lignin in potato tubers, Front. Plant Sci. 2022, 13, 942022. [Google Scholar]
- Han, Y.; Yang, R.R.; Zhang, X.J.; Wang, Q.H.; Wang, B.; Zheng, X.Y.; Li, Y.C.; Prusky, D.; Bi, Y. Brassinosteroid accelerates wound healing of potato tubers by activation of reactive oxygen metabolism and phenylpropanoid metabolism. Foods 2022, 11, 906. [Google Scholar] [CrossRef]
- Jiang, H.; Li, X.; Ma, L.; Ren, Y.Y.; Bi, Y.; Prusky, D. Transcriptome sequencing and differential expression analysis of natural and BTH-treated wound healing in potato tubers (Solanum tuberosum L.). BMC Genom. 2022, 23, 263. [Google Scholar] [CrossRef]
- Su, Q.H.; Kondo, N.; Li, M.Z.; Sun, H.; Al Riza, D.F.; Habaragamuwa, H. Potato quality grading based on machine vision and 3D shape analysis. Comput. Electron. Agric. 2018, 152, 261–268. [Google Scholar] [CrossRef]
- Pinhero, R.G.; Coffin, R.; Yada, R.Y. Post-harvest storage of potatoes. In Advances in Potato Chemistry and Technology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 339–370. [Google Scholar]
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Jakubowski, T. Use of UV-C radiation for reducing storage losses of potato tubers. Bangladesh J. Bot. 2018, 47, 533–537. [Google Scholar] [CrossRef] [Green Version]
- Ranganna, B.; Kushalappa, A.C.; Raghavan, G.S.V. Ultraviolet irradiance to control dry rot and soft rot of potato in storage. Can. J. Plant Pathol. 1997, 19, 30–35. [Google Scholar] [CrossRef]
- Yu, B.; Nan, Y.Y.; Kong, N.N.; Dang, R.M.; Bai, Z.L.; Liu, J. Effect of short-wave Ultraviolet irradiation on dry rot and relevant enzymes activities of post-harvest potato tubers. J. Nucl. Agric. Sci. 2017, 31, 1730–1736. [Google Scholar]
- Jakubowski, T.; Krolczyk, J.B. Method for the reduction of natural losses of potato tubers during their long-term storage. Sustainability 2020, 12, 1048. [Google Scholar] [CrossRef] [Green Version]
- Chudinova, E.M.; Kokaeva, Y.L.; Elansky, S.N.; Kutuzova, I.A.; Pertsev, A.S.; Pobendinskaya, M.A. The occurrence of thiabendazole-resistant isolates of Helminthosporium solani on potato seed tubers in Russia. J. Plant Dis. Prot. 2020, 127, 421–423. [Google Scholar] [CrossRef]
- Gachango, E.; Hanson, L.E.; Rojas, A. Fusarium spp. causing dry rot of seed potato tubers in Michigan and their sensitivity to fungicides. Plant Dis. 2012, 96, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Malyuga1, A.A.; Chulikova, N.S.; Ilyin, M.M.; Khalikov, S.S. Fludioxonil-based preparations for protecting potatoes from diseases and their effectiveness. Russ. Agric. Sci. 2022, 48, S74–S83. [Google Scholar] [CrossRef]
- Raigond, P.; Sagar, V.; Mishra, T. Chitosan: A safe alternative to synthetic fungicides to manage dry rot in stored potatoes. Potato Res. 2019, 62, 393–409. [Google Scholar] [CrossRef]
- Xue, H.; Bi, Y.; Prusky, D. The mechanism of induced resistance against Fusarium dry rot in potato tubers by the T-2 toxin. Postharvest Biol. Technol. 2019, 153, 69–78. [Google Scholar] [CrossRef]
- Ma, L.; Jiang, H.; Ren, Y.Y.; Yang, J.W.; Han, Y.; Si, H.J.; Prusky, D.; Bi, Y.; Wang, Y. Overexpression of StCDPK23 promotes wound healing of potato tubers by regulating StRbohs. Plant Physiol. Biochem. 2022, 185, 279–289. [Google Scholar] [CrossRef]
- Hay, W.T.; Fanta, G.F.; Rich, J.O. Antifungal activity of a fatty ammonium chloride amylose inclusion complex against Fusarium sambucinum; control of dry rot on multiple potato varieties. Am. J. Potato Res. 2019, 96, 79–85. [Google Scholar] [CrossRef]
- Mahmoud, G.A.; El-Tobgy, K.M.K.; Abo-El-Seoud, M.A. Utilisation of biocides for controlling pest attacks on potato tubers. Arch Phytopathol. Plant Prot. 2010, 43, 251–258. [Google Scholar] [CrossRef]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Turon, C.; Marin, S. Impact of essential oils on growth rate, zearalenone and deoxynivalenol production by Fusarium graminearum under different temperature and water activity conditions in maize grain. J. Appl. Microbiol. 2004, 96, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Othmane, M.; Djazouli, Z.; Zebib, B. Aqueous extract of algerian nettle (Urtica dioïca L.) as possible alternative pathway to control some plant diseases. Iran. J. Sci. Technol. Sci. Trans. A 2021, 45, 463–468. [Google Scholar]
- Boivin, M.; Nathalie, B.; Simon, B.; Isabel, D. Black spruce extracts reveal antimicrobial and sprout suppressive potentials to prevent potato (Solanum tuberosum L.) losses during storage. J. Agric. Food Sci. 2021, 5, 100187. [Google Scholar] [CrossRef]
- Mvuemba, H.; Green, S.; Tsopmo, A.; Avis, T. Antimicrobial efficacy of cinnamon, ginger, horseradish and nutmeg extracts against spoilage pathogens. Phytoprotection 2009, 90, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Tian, S.L.; Shen, J.; Wang, X.Z.; Cheng, J.X.; Li, S.Q.; Ge, X.; Tian, J.C. Effects of chlorine dioxide on morphology and ultrastructure of Fusarium sulphureum and its virulence to potato tubers. Int. J. Agric. Biol. Eng. 2017, 10, 242–250. [Google Scholar]
- Liu, Q.L.; Zhang, R.; Xue, H.L.; Bi, Y.; Li, L.; Zhang, Q.Q.; Carelle, J.K.; Nan, M.N.; Prusky, D. Ozone controls potato dry rot development and diacetoxyscirpenol accumulation by targeting the cell membrane and affecting the growth of Fusarium sulphureus. Physiol. Mol. Plant Pathol. 2022, 118, 101785. [Google Scholar] [CrossRef]
- Liu, Z.G.; Yang, X.; Xue, H.L.; Bi, Y.; Zhang, Q.Q.; Liu, Q.L.; Chen, J.Y.; Nan, M.N.; Prusky, D. Reactive oxygen species metabolism and diacetoxyscirpenol biosynthesis modulation in potato tuber inoculated with ozone-treated Fusarium sulphureum. J. Food Process. Preserv. 2023, 2023, 4823679. [Google Scholar] [CrossRef]
- Schisler, D.A.; Slininger, P.J. Selection and performance of bacterial strains for biologically controlling Fusarium dry rot of potatoes incited by Gibberella pulicaris. Plant Dis. 1994, 78, 251–255. [Google Scholar] [CrossRef]
- Schisler, D.A.; Slininger, P.J.; Bothast, R.J. Effects of antagonist cell concentration and two-strain mixtures on biological control of Fusarium dry rot of potatoes. Phytopathology 1997, 87, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çakar, G.; Tozlu, E. The biological control of Fusarium oxysporum, the causal agent of potato rot. Gesunde Pflanz. 2022, 74, 305–315. [Google Scholar] [CrossRef]
- Daami-Remadi, M.; Hibar, K.; Jabnoun-Khiareddine, H.; Ayed, F.; El Mahjoub, M. Effect of two Trichoderma species on severity of potato tuber dry rot caused by Tunisian Fusarium complex. Int. J. Agric. Res. 2006, 1, 432–441. [Google Scholar]
- El-Kot, G.A.N. Biological control of black scurf and dry rot of potato. Egypt J. Phytopathol. 2008, 36, 45–56. [Google Scholar]
- Paul, N.C.; Park, S.; Liu, H.F.; Lee, J.G.; Han, G.H.; Kim, H.; Sang, H. Fungi associated with postharvest diseases of sweet potato storage roots and in vitro antagonistic assay of Trichoderma harzianum against the diseases. J. Fungi 2021, 7, 927. [Google Scholar] [CrossRef]
- Wharton, P.S.; Kirk, W.W. Evaluation of biological seed treatments in combination with management practices for the control of Fusarium dry rot of potato. Biocontrol Control 2014, 73, 23–30. [Google Scholar] [CrossRef]
- Hussain, T.; Khan, A.A.; Mohamed, H.I. Metabolites composition of Bacillus subtilis Hussaint-Amu determined by LC-MS and their effect on Fusarium dry rot of potato seed tuber. Phyton-Int. J. Exp. Bot. 2023, 92, 783–799. [Google Scholar] [CrossRef]
- Yu, X.Y.; Bi, Y.; Yan, L.; Liu, X.; Wang, Y.; Shen, K.P.; Li, Y.C. Activation of phenylpropanoid pathway and PR of potato tuber against Fusarium sulphureum by fungal elicitor from Trichothecium roseum. World J. Microbiol. Biotechnol. 2016, 32, 142. [Google Scholar] [CrossRef]
- Recep, K.; Fikrettin, S.; Erkol, D.; Cafer, E. Biological control of the potato dry rot caused by Fusarium species using PGPR strains. Biol. Control 2009, 50, 194–198. [Google Scholar] [CrossRef]
Figure 1.
An illustration of the process of Fusarium species infecting potato tubers.

Figure 2.
Basic chemical structure of trichothecenes.


Figure 3.
The strategy of controlling potato dry rot via physical, chemical and biological methods and the possible action mechanism.
Figure 3.
The strategy of controlling potato dry rot via physical, chemical and biological methods and the possible action mechanism.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The reported Fusarium spp. to lead to dry rot of potato in diverse countries and regions.
| Fusarium Species | Region | Reference |
|---|---|---|
| F. sambucinum | North American and some regions of Europe | [16,17,18] |
| F. coeruleum | United Kingdom and Great Britain | [19,20,21] |
| F. graminearum | North Dakota | [22] |
| F. solani and | South Africa | [23,24] |
| F. oxysporum | Michigan | |
| F. sulphureum and F. solani | Iran | [25] |
| F. sambucinum | Egypt | [26] |
| F. oxysporum | ||
| F. verticillioides | ||
| F. incarnatum | ||
| F. avenaceum, | Heilongjiang Province and Inner Mongolia Autonomous Region | [27] |
| F. oxysporum, | ||
| F. sporotrichiodes | ||
| F. solani, | ||
| F. trichothecioides, | ||
| F. solani var. coeruleum | ||
| F. sambucinum | ||
| F. semitectum, | ||
| F. solani | ||
| F. sambucinum | ||
| F. culmorum, | ||
| F. gibbosum, | ||
| F. macroceras, | ||
| F. solani var. coeruleum, F. acuminatum, | ||
| F. equiseti and F. redolens | ||
| F. sambucinem | North of China | [27] |
| F. avenaceum | Shanxi Province | |
| F. solani var. coeruleum, | ||
| F. oxysporum, | ||
| F. acuminatum | ||
| F. sambucinem | Northwest of China, | [7,8,10,11,12,27] |
| F. avenaceum | Gansu Province, Ningxia Hui Autonomous Region | |
| F. graminearum | ||
| F. solani | ||
| F. sulphureum | ||
| F. tricinctum, | Northwest of China, Qinghai Province | [6] |
| F. avenaceum, | ||
| F. oxysporum, | ||
| F. solani, | ||
| F. acuminatum, | ||
| F. equiseti | ||
| F. solani, | South of China Zhejiang Province | [6] |
| F. moniliform, | ||
| F. redolens |
Table 2.
Non-trichothecenes generated by Fusarium spp. in potato dry rot.
| Fusarium Species | Non-Trichothecenes | Reference |
|---|---|---|
| F. oxysporum | BEA, ENNs | [36] |
| F. sambucinum | ZEA, FUS | [37] |
| F. solani | ZEA, FUS | [37] |
| F. oxysporum | ZEA, FUS | [37] |
| F. crookwellense | ZEA, FUS | [38] |
| F. crookwellense | ZEA, FUS | [39] |
| F. crookwellense | ZEA, FUS | [40] |
| F. graminearum | ZEA | [40] |
| F. equiseti | FUM, ZEA | [41] |
| F. oxysporum | FA, FUM, ZEA | [41] |
| F. sambucinum | SAM | [42] |
| F. sambucinum | SAM | [43] |
| F. sambucinum | SAM | [44] |
| F. oxysporum | FA | [45] |
| F. oxysporum | FA | [46] |
Note: BEA: beauvericin, ENNs: enniatins, ZEA: zearalenone, FA: fusaric acid, FUM: fumonisin, FUS: fusarin C, SAM: sambutoxin.
Table 3.
Trichothecenes generated by Fusarium spp. in potato dry rot.
| Fusarium Species | Trichothecenes | Reference |
|---|---|---|
| F. sambucinum | trichothecene | [37] |
| F. solani | trichothecene | [37] |
| F. oxysporum | trichothecene | [37] |
| F. crookwellense | NIV, FX | [38] |
| F. sambucinum | DAS | [39] |
| F. crookwellense | NIV, FX | [39] |
| F. sulphureum | 3-ADON, T-2, FUS, DAS | [4] |
| F. solani | 3-ADON, T-2, FUS, DAS | [4] |
| F. sambucinum | 3-ADON, T-2, FUS, DAS | [4] |
| F. equiseti | T-2 | [41] |
| F. oxysporum | T-2 | [41] |
| F. sambucinum | DAS | [50] |
| F. sambucinum | DAS, MAS, NEO, T-2, HT-2 | [52] |
| F. sambucinum | 4,15-DAS, 15-MAS, 4-MASc | [53] |
| F. solani | DON, HT-2, 3-ADON | [54] |
| F. sambucinum | DON, NIV, HT-2 | [54] |
| F. sambucinum | MAS, DAS | [55] |
| F. crookwellense | NIV, DAS | [56] |
| F. culmorum | NIV, FX, 3-ADON, DON | [57] |
| F. crookwellense | FX | [57] |
| F. equiseti | NIV, FX, 4-MAS, 15-MAS, DAS, SCR | [57] |
| F. graminearum | NIV, FX, DON, 3-ADON, 15-ADON | [57] |
| F. sambucinum | SCR, 4-MAS, 15-MAS, DAS, NEO, HT-2,T-2 | [57] |
| F. graminearum | DON, NIV, FX, 3-ADON, 15- ADON | [51] |
| F. graminearum | NIV, T-2, 3-ADON, 15-ADON,15-SCR | [58] |
| F. graminearum | DON, 3ADON, 15-ADON | [59] |
| F. culmorum | DON, 3-ADON | [60] |
Note: DAS: diacetoxyscirpenol; 4,15-DAS: 4,15-diacetoxyscirpenol; DON: deoxynivalenol; 3-ADON: 3-acetyldeoxynivalenol; 15-ADON: 15-acetyldeoxynivalenol; HT-2: HT-2 toxin; MAS: mono-acetoxyscirpenol; 4-MAS: 4-acetyl-monoacetoxyscirpenol; NIV: nivalenol; FX: fusarenone X; NEO: neosolaniol; SCR: scirpentriol; 15-MAS: 15-acetyl-monoacetoxyscirpenol; T-2: T-2 toxin; 15-SCRP: 15-acetylscripenol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xue, H.; Liu, Q.; Yang, Z. Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. J. Fungi 2023, 9, 843. https://doi.org/10.3390/jof9080843
AMA Style
Xue H, Liu Q, Yang Z. Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. Journal of Fungi. 2023; 9(8):843. https://doi.org/10.3390/jof9080843
Chicago/Turabian StyleXue, Huali, Qili Liu, and Zhimin Yang. 2023. "Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review" Journal of Fungi 9, no. 8: 843. https://doi.org/10.3390/jof9080843
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.