
Inhibitory Mechanisms of trans-2-Hexenal on the Growth of Geotrichum citri-aurantii
by
Qiuli Ouyang
*,
Shiwei Shi
,
Yangmei Liu
,
Yanqin Yang
,
Yonghua Zhang
,
Xingxing Yuan
,
Nengguo Tao
* and
Lu Li
School of Chemical Engineering, Xiangtan University, Xiangtan 411105, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(9), 930; https://doi.org/10.3390/jof9090930
Submission received: 30 July 2023
/
Revised: 7 September 2023
/
Accepted: 8 September 2023
/
Published: 15 September 2023
(This article belongs to the Special Issue Isolation and Control of Fruit and Vegetable Rot Fungi)
Abstract
:Geotrichum citri-aurantii (G. citri-aurantii) is one of the most important postharvest pathogens leading to a postharvest loss of citrus by causing sour rot. In this study, the antifungal activity of trans-2-hexenal, a natural component of essential oil, against G. citri-aurantii was evaluated. Trans-2-hexenal treatment inhibited the mycelia growth of G. citri-aurantii with a minimum inhibitory concentration and minimum fungicidal concentration of trans-2-hexenal at 0.50 and 1.00 μL/mL, respectively. Moreover, trans-2-hexenal efficiently reduced the incidence of sour rot of Satsuma fruit inoculated with G. citri-aurantii. Ultrastructural observations and Fourier transform infrared (FT−IR) results showed that trans-2-hexenal treatment affected the cell wall and cell membrane instructions of G. citri-aurantii. The content of β-1,3-glucan was significantly decreased after trans-2-hexenal treatment, but the cell wall permeability was not changed. The decrease in lipid and ergosterol contents might be responsible for this antifungal activity. Several important genes, FKS1, ERG1, ERG7, and ERG11, showed decreasing expression levels after trans-2-hexenal treatment. Molecule-docking results also indicated that trans-2-hexenal could join with the protein of FKS1, ERG1, ERG7, and ERG11 to impact enzyme activities. These results demonstrated that trans-2-hexenal is a promising fungicide for controlling sour rot of harvested citrus fruit by damaging the membrane integrity of G. citri-aurantii.
1. Introduction
G. citri-aurantii is a necrotrophic fungal pathogen that infects citrus fruit, and it has the characteristics of strong infectivity, fast infectivity, and being difficult to control [1,2]. Synthetic fungicides are exclusively used to control this disease but cause serious hazardous effects on the fruit rind, the environment, and human health [3,4]. Thus, it is necessary to focus on developing alternatives to synthetic fungicides for handling and maintaining the quality of citrus fruit [5,6,7].
Previous studies have shown that plant essential oils and their antimicrobial components have significant inhibitory effects on many postharvest pathogenic fungi of citrus, such as G. citri-aurantii, Penicillium digitatum, and P. italicum, and that they have the advantages of safety, high efficiency, and low residue, which means they have the potential to control postharvest diseases in citrus [8,9]. Trans-2-hexenal is a volatile component of plant essential oil, which naturally exists in citronella oil, camphor oil, apples, and grapes. It is an important signal molecule for plants to respond to and defend against their external environment. Some studies have shown that trans-2-hexenal has good inhibitory effects against Colletotrichum acutatum, Alternaria alternata, P. cyclopium, P. expansum, and Botrytis cinerea [10,11,12,13]. In addition, trans-2-hexenal was also successfully applied in order to control postharvest diseases such as gray mold in tomato fruits, green mold in citrus fruits, and black rot in ‘Zaosu’ pears [9,14,15].
As mentioned above, several studies have shown that trans-2-hexenal is a potential biological alternative to other preservative methods of controlling postharvest diseases, but there are few studies related to the investigation of its exact mechanism of antifungal action. Studies have shown that trans-2-hexenal interferes with the cell wall structure of fungi. For example, Arroyo et al. [10] pointed out that treatment with trans-2-hexenal can cause obvious cracks in the cell wall, disorder cell components to a high degree, and cause the organelle morphology to disappear and the cells to crack. Research by Zhang et al. [13] found that the membrane permeability of P. cyclopium increased with an increase of trans-2-hexenal concentration, resulting in the release of cell components and the leakage of potassium ions. At the same time, the integrity of the cell membrane of P. cyclopium was destroyed due to the decrease in the lipid content. Ma et al. [16] showed that 1.0 μL/mL trans-2-hexenal inhibited spore germination by disrupting the mitochondrial energy metabolism of Aspergillus flavus; the trans-2-hexenal treatment also decreased the acetyl-CoA and ATP contents, and the mitochondrial dehydrogenases activity increased by 65.7 ± 3.7%, 53.9 ± 4.0%, and 23.8 ± 2.2%, respectively. However, the antifungal activity of trans-2-hexenal against G. citri-aurantii has not been determined, and the antifungal mechanism of trans-2-hexenal against G. citri-aurantii has not been studied.
Thus, this study aimed to (1) study the antifungal properties of trans-2-hexenal against G. citri-aurantii in vitro and in vivo, (2) investigate the effect of the cell wall and cell membrane of G. citri-aurantii in the presence of trans-2-hexenal, and (3) further explore the possible mechanism through RT-qPCR and molecular docking.
2. Materials and Methods
2.1. Pathogen
G. citri-aurantii was provided by the Department of Biotechnology and Food Engineering, Xiangtan University, Xiangtan, China. The fungus was purified and preserved at 28 ± 2 °C on potato dextrose agar (PDA). A spore suspension (5 × 106 spores/mL) in potato dextrose broth (PDB) was prepared using a hemocytometer.
2.2. Fruit
Satsuma mandarin fruits (Citrus unshiu Marc. cv. Miyagawa Wase) were harvested on 18 October 2018 from an orchard in Xiangtan, Hunan, China. Healthy fruits of uniform size and without scars were selected for the experiments.
2.3. Chemicals
Trans-2-hexenal (98%) was obtained from Aladdin (Shanghai, China). All the chemicals were analytical grade.
2.4. Antifungal Activity of trans-2-Hexenal against G. citri-aurantii
The inhibition of trans-2-hexenal on the growth of G. citri-aurantii mycelia was tested in vitro through the agar dilution method [17]. Briefly, trans-2-hexenal solutions were prepared by dissolving the requisite amount in Tween-80 (0.5%, Aladdin, Shanghai, China) and adding it to PDA (20 mL) to achieve the desired concentrations (0, 0.25, 0.50, 1.0, 2.0, and 4.0 μL/mL). A 6 mm diameter mycelial disk of inoculate was cut from the actively growing culture of the PDA plates. Then, they were placed at the center of each new Petri plate (90 mm in diameter). The culture plates were then incubated at 28 ± 2 °C for 2 d. Each treatment was performed in triplicate. The percentage of inhibition of mycelial growth (MGI) was calculated according to the following formula:
where dc (cm) is the average diameter of the control and dt (cm) is the average diameter of the treatment. The lowest concentration that completely inhibited the growth of G. citri-aurantii after 2 d of incubation was considered to be the minimum inhibitory concentration (MIC). The minimum fungicidal concentration (MFC) was regarded as the lowest concentration that prevented the growth of the pathogen after 4 d of incubation at 28 ± 2 °C, indicating that more than 99.5% of the original inocula were killed.
MGI (%) = [(dc − dt)/(dc − 6)] × 100%
2.5. In Vivo Experiments of trans-2-Hexenal against G. citri-aurantii
The effect of the trans-2-hexenal on the incidence of sour rot was determined as described previously by Dou et al. [18]. All fresh citrus fruit were surface-sterilized by immersing in 2% sodium hypochlorite solution (v/v) for 2 min, then washed with distilled water, wounded (depth of 3 mm and width of 3 mm) with a sterile needle, inoculated with 20 μL of G. citri-aurantii spore suspension (105 spores mL−1), and left to air-dry. After being inoculated with G. citri-aurantii, the fruit were soaked in wax amended with trans-2-hexenal at 1× MFC and 10× MFC. The fruit with wax and inoculated with the pathogen inoculation was used as a control. The inoculated fruit was kept in sealed incubators at 25 ± 2 °C to ensure a high relative humidity (80–85% relative humidity). Each treatment was performed in triplicate, and each replicate contained 20 Satsuma fruits. The incidence rate of disease (measured by counting the number of green-mold-infected wounds) was calculated as follows:
2.6. Scanning Electron Microscopy (SEM) of trans-2-Hexenal against G. citri-aurantii
The mycelia treated with trans-2-hexenal for 30 min, as described above, were directly examined using a JEOL JSM-6360LV SEM instrument (JEOL, Tokyo, Japan). The hyphae grown on PDA without trans-2-hexenal were used as a control. The procedures for the SEM observation were described in our previous study [19].
2.7. Transmission Electron Microscopy (TEM) of trans-2-Hexenal against G. citri-aurantii
The mycelia treated with trans-2-hexenal for 30 min, as described above, were directly examined using a transmission electron microscope (JEM-1230; JEOL Ltd., Tokyo, Japan) operated at an accelerating voltage of 80 kV. The hyphae grown on PDA without trans-2-hexenal were used as a control. The procedures for the TEM observation were described in our previous study [19].
2.8. Fourier Transform Infrared (FT−IR) Spectroscopy of trans-2-Hexenal against G. citri-aurantii
The effect of trans-2-hexenal on the mycelia composition of G. citri-aurantii was analyzed using Fourier transform infrared spectroscopy (FT−IR) (Thermo Fisher Scientific, Waltham, MA, USA) [20]. The mycelia treated with 1/2 MIC trans-2-hexenal for 30 min were collected, frozen with liquid nitrogen, and then vacuum freeze-dried. Subsequently, the mycelia were ground (100 mesh) to obtain uniform dried powder. The samples were prepared using the potassium bromide-disk technique for the FT−IR detection. The scanning range was 4000–400 cm−1 with the resolution of 4 cm−1 and 128 separate scans. The infrared spectrum was analyzed using Unscrambler X (Version 10.4).
2.9. Effect of trans-2-Hexenal on the Cell Wall of G. citri-aurantii
The effects of trans-2-hexenal on the cell wall integrity of G. citri-aurantii were analyzed using calcofluor white (Sigma, St. Louis, MO, USA) staining coupled with fluorescence microscopy. The mycelia treated with 1/2 MIC trans-2-hexenal for 0, 30, 60, and 120 min were centrifuged at 4000× g for 10 min. The collected mycelia were stained with 10 μL of calcofluor white stain after the addition of 10 μL KOH (10%) following the manufacturer’s instructions. The samples were observed with a fluorescence microscope (Nikon ECLIPSE TS100, Tokyo, Metropolis, Japan). The fungal culture in PDB without trans-2-hexenal was used as a control.
2.10. Effect of trans-2-Hexenal on the Cell Wall of G. citri-aurantii
2.10.1. Effect of trans-2-Hexenal on the Cell Wall Integrity of G. citri-aurantii
The effects of trans-2-hexenal on the cell wall integrity of G. citri-aurantii were analyzed using calcofluor white (Sigma, St. Louis, MO, USA) staining coupled with fluorescence microscopy. The mycelia treated with 1/2 MIC trans-2-hexenal for 0, 30, 60, and 120 min were centrifuged at 4000× g for 10 min. The collected mycelia were stained with 10 μL of calcofluor white stain after the addition of 10 μL KOH (10%, Aladdin, Shanghai, China) following the manufacturer’s instructions. The samples were observed with a fluorescence microscope (Nikon ECLIPSE TS100, Tokyo Metropolis, Japan). The fungal culture in PDB without trans-2-hexenal was used as a control.
2.10.2. Effect of trans-2-Hexenal on the Chitin Content of G. citri-aurantii
The chitin contents of the G. citri-aurantii treated with the 1/2 MIC trans-2-hexenal treatments in PDB were determined using the method of Francois [21]. A total of 0.5 g of dried mycelia was soaked in 4 mL of concentrated HCl (Soleibao, Beijing, China) at 25 °C for 24 h, then diluted with distilled water until the HCl reached a concentration of 8.5 mol/L, and the solution was further digested in a boiling water bath. After cooling, it was adjusted to neutral with 1 mol/L of NaOH (Aladdin, Shanghai, China) solution, the volume was made constant to 100 mL, then filtered with filter paper, and the supernatant was shaken to obtain the sample to be tested. Then, 200 μL supernatant was mixed with 400 μL acetylacetone reagent and placed in a 90 °C water bath for 1 h. After cooling to room temperature, 4 mL absolute ethyl alcohol and 400 μL 4-dimethylaminobenzaldehyde were added to the supernatant, the volume was fixed to 5 mL with absolute ethyl alcohol, and it was left to stand at room temperature for 1 h. The absorbance of the solution was measured at a wavelength of 530 nm, and glucosamine hydrochloride was used as the standard curve.
2.10.3. Effect of trans-2-Hexenal on the β-1,3-Glucan Content of G. citri-aurantii
The method of Fortwendel et al. [22] was used for the determination of β-1,3-glucan. The mycelia were washed with 0.1 mol/L NaOH solution and then freeze-dried and ground into powder. A certain amount of powder was added to 1 mol/L NaOH solution, ultrasonicated for 30 s, placed in a water bath at 52 °C for 30 min, and centrifuged for 5 min after cooling. A total of 50 μL of the supernatant was taken, 185 μL of aniline blue solution was added, and it was placed in a water bath at 52 °C for 30 min and left to stand for 30 min. The fluorescence value (excitation wavelength: 405 nm; emission wavelength: 460 nm) was measured using a fluorescence spectrophotometer (Lengguang Technology Co., Ltd., Shanghai, China).
2.10.4. Effect of trans-2-Hexenal on the Extracellular Alkaline Phosphatase (AKP) Activities of G. citri-aurantii
The extracellular AKP activities of the G. citri-aurantii mycelia that received different trans-2-hexenal treatments in PDB, as described above, were assayed with a UV-2450 UV/V spectrophotometer (Shimadzu (China) Co., Ltd., Shanghai, China) using a commercially available kit, following the instructions. The fungal culture in PDB without trans-2-hexenal was used as a control. Each experiment was repeated three times. The enzyme activity is expressed as U/g prot.
2.11. Effect of trans-2-Hexenal on the Cell Membrane of G. citri-aurantii
2.11.1. Effect of trans-2-Hexenal on the Cell Membrane Integrity of G. citri-aurantii
The cell membrane integrity of the G. citri-aurantii that received different trans-2-hexenal treatments in PDB, as described above, was analyzed using propidium iodide (PI) staining coupled with an ECLIPSE TS100 microscope (Nikon, Tokyo Metropolis, Japan) and F97 PRO fluorescence spectrophotometer (Lengguang Technology, Shanghai, China) [23].
2.11.2. Effect of trans-2-Hexenal on the Total Lipid Content of G. citri-aurantii
The total lipid content of G. citri-aurantii cells with trans-2-hexenal at various concentrations (0 and 1/2 MIC) for 0, 30, 60, and 120 min was determined using the phosphovanillin method [19]. The fungal culture in PDB without trans-2-hexenal was used as a control.
2.11.3. Effect of trans-2-Hexenal on the Ergosterol Contents of G. citri-aurantii
The ergosterol contents of G. citri-aurantii cells that received different trans-2-hexenal treatments in PDB, as described above, were determined using the HPLC method [23]. The fungal culture in PDA without trans-2-hexenal was used as a control.
2.12. Real-Time Fluorescence Quantitative PCR (RT-qPCR) Analysis
The effects of trans-2-hexenal on the transcriptional profiles of FKS1 (the key gene that synthesizes β-1,3-glucan synthase) and genes related to ergosterol synthesis (ERG1, ERG7, and ERG11) in G. citri-aurantii were evaluated, and the sequences were obtained from a previous RNA-Seq of G. citri-aurantii. RNA was extracted from G. citri-aurantii cells exposed to trans-2-hexenal at concentrations of 0 and 1/2 MIC for 0, 30, 60, and 120 min using Trizol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions. Two micrograms of DNA-free RNA were used for the reverse transcription using M-MLV (Promega, Madison, WI, USA) with oligo dT18. The RT-qPCR was performed on a BIO-RAD CFX Connect Thermal Cycler using FastStart Universal SYBR Green Master (Roche, Basel, Switzerland). All primer pairs for the expression assays are listed in Table 1. The RT-qPCR was programmed as follows: initial denaturation at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 15 s and a combined annealing and extension step at 60 °C for 1 min. The 2−ΔΔCT method was used to quantify the value of every sample using the actin gene as an internal reference [24].
2.13. Molecular Docking
Selecting FKS1, ERG1, ERG7, and ERG11 as the receptors and using Alphafold2 (https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2ipynb; accessed on 8 July 2022) for the homology modeling, the structure of the models were optimized using ModRefiner (https://zhanggroup.org/ModRefiner/; accessed on 8 July 2022), and the obtained models were evaluated with SAVES v6.0 (https://saves.mbi.ucla.edu/; accessed on 20 July 2022). In this way, the three-dimensional protein structure models of ERG1, ERG7, ERG11, and FKS1 were obtained. The structure of the ligand small molecule trans-2-hexenal (CID: 5281168; MF: C6H10O) was obtained from the chemical structure database of the PubChem (https://pubchem.ncbi.nlm.nih.gov/; accessed on 20 July 2022) website. Using PyMol (Version 2.5.2) to process the receptor protein, the water molecules and metal ions were deleted. The Openbabel module in PyRx (Version 0.8) was used to minimize the ability of the ligands. The Autodock Vina module in PyRx was used for the molecular docking. The optimal docking model was selected according to the binding energy. PyMol and Ligplot+ (Version 2.2.8) were used to connect the three-dimensional and two-dimensional visual analysis of the model.
2.14. Statistical Analyses
All data are expressed as the mean ± SD (standard deviation), and they were measured using three independent replicates and analyzed using one-way analysis of variance (ANOVA) followed by Duncan’s test. A value of p < 0.05 was considered statistically significant using SPSS statistical software package release 16.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Antifungal Activity of trans-2-Hexenal against G. citri-aurantii
Table 2 shows the effect of trans-2-hexenal on the mycelial growth of G. citri-aurantii in vitro. The results show that the mycelial growth considerably decreased with an increasing trans-2-hexenal concentration and incubation time. Mycelia growth was inhibited to different degrees at 0.25 μL/mL to 0.50 μL/mL of trans-2-hexenal. At a concentration of 0.50 μL/mL, the growth of G. citri-aurantii was completely inhibited after 2 d of incubation. As the duration of the culture was prolonged to 4 d, 72.9 ± 3.9% and 100.0 ± 0.0% of the mycelial growth was inhibited by 0.50 and 1.00 μL/mL of trans-2-hexenal, respectively. Thus, the MIC and MFC of trans-2-hexenal were 0.50 and 1.00 μL/mL, respectively.
3.2. In Vivo Experiments of trans-2-Hexenal against G. citri-aurantii
trans-2-Hexenal (1× and 10× MFC) effectively reduced the decay of citrus fruit inoculated with G. citri-aurantii (Table 3), and the disease progression in the inoculated citrus fruit treated with trans-2-hexenal is presented in Figure 1. The control group began to decay within 2 d with 11 ± 4% decay, while the citrus fruit remained healthy after treatment with trans-2-hexenal. The citrus fruit in the 1× and 10× MFC trans-2-hexenal groups began to rot after 3 d and 5 d of treatment, respectively. After 7 d of storage, the incidence of fruit in the fruit wax control group reached 100%, while that in the 1× and 10× MFC treatment groups were only 85 ± 4% and 33 ± 12%, respectively.
3.3. Scanning Electron Microscopy (SEM) of trans-2-Hexenal against G. citri-aurantii
The effects of trans-2-hexenal on the surface morphology of G. citri-aurantii are shown in Figure 2. The mycelia in the control group were regular in shape, uniform in thickness, smooth on the surface, healthy, and full (Figure 2A,B). However, after the 1/2 MIC trans-2-hexenal treatment, the surface of the mycelia became wrinkled and severely twisted, shrank, and collapsed (Figure 2C,D). The results show that trans-2-hexenal can change the morphology of mycelia of G. citri-aurantii.
3.4. Transmission Electron Microscopy (TEM) of trans-2-Hexenal against G. citri-aurantii
The effects of trans-2-hexenal on the internal morphology of G. citri-aurantii are shown in Figure 3. In the control group, the cells were composed of uniform cell walls, cell membranes, and cytoplasm, with uniform organelles and a complete structure (Figure 3A). After the 1/2 MIC trans-2-hexenal treatment, the internal morphology and ultrastructure of the G. citri-aurantii cells were destroyed, the cell walls became thicker, the cell membranes were irregularly twisted and, in some areas, the organelle structures were disordered, and the mitochondria were enlarged with irregular distribution (Figure 3B).
3.5. FT−IR of trans-2-Hexenal against G. citri-aurantii
In the region of >3000 cm−1, after the trans-2-hexenal treatment, the O–H expansion (3775 cm−1) of alcohols in the carbohydrates shifted, suggesting that the trans-2-hexenal treatment may change the components of the cell walls and cell membranes of G. citri-aurantii (Figure 4). The N-H stretching (3415 cm−1) of the amino groups and amide groups also shifted, which may be related to the Michael addition reaction between the α,β-unsaturated carbonyl groups in trans-2-hexenal and the amino groups in protein, resulting in the degradation of the cell wall proteins. In the 2800–3100 cm−1 region, trans-2-hexenal caused a significant shift of the C-H asymmetric stretching (2928 cm−1) in the methyl and acyl chains, which might be related to the changes in the fatty acids caused by the condensation of the active methylene-containing compounds with aldehydes. The amides in the polypeptides and the protein (1584 cm−1 and 1637 cm−1) in the 1500–1700 cm−1 region all shifted, which might be related to the peroxidation and degradation of protein. In the range of 900–1500 cm−1, the peak positions at 1400 cm−1 and 1077 cm−1, corresponding to fatty acids, proteins, polysaccharides, and nucleic acid, shifted after the treatment with trans-2-hexenal, suggesting that the cell wall’s polysaccharides and nucleic acid were affected by trans-2-hexenal.
3.6. Effect of trans-2-Hexenal on the Cell Wall of G. citri-aurantii
Figure 5A shows the effect of trans-2-hexenal on the chitin content of G. citri-aurantii. There was no significant difference in the chitin content between the trans-2-hexenal-treated group and the control group, suggesting that trans-2-hexenal had no effect on the chitin content of G. citri-aurantii.
The content of β-1,3-glucan in the trans-2-hexenal-treated group was significantly lower than that in the control group (Figure 5B). At 30 min, the trans-2-hexenal-treated group decreased by 17.31% compared with the control group (p < 0.05), which indicates that the content of β-1,3-glucan significantly reduced after trans-2-hexenal treatment.
The activity of extracellular AKP after trans-2-hexenal treatment had no significant difference with the control group (p < 0.05, Figure 5C), which further indicates that trans-2-hexenal did not damage the cell wall integrity of G. citri-aurantii.
3.7. Effect of trans-2-Hexenal on the Cell Membrane of G. citri-aurantii
According to the results of the PI staining (Figure 6A), the fluorescence value of the mycelia treatment group after the 1/2 MIC trans-2-hexenal treatment for 60 min was 1.24 times that of the control group, which was significantly higher than that of the control group (p < 0.05), indicating that the integrity of the cell membrane might begin to be destroyed after 60 min.
The effect of trans-2-hexenal on the total lipid content of G. citri-aurantii is shown in Figure 6B. After the 1/2 MIC trans-2-hexenal treatment for 60 min, the total lipid content was 123.8 ± 1.6 mg/g DW, which was significantly lower than that of the control group (185.4 ± 13.3 mg/g DW) (p < 0.05), suggesting that the integrity of the cell membrane of G. citri-aurantii was destroyed, and the cells were damaged after the 1/2 MIC trans-2-hexenal treatment for 60 min.
The ergosterol content (3.54 ± 0.07 mg/g DW) of G. citri-aurantii was significantly lower than that of the control group (5.89 ± 0.24 mg/g DW) (p < 0.05) after 30 min of treatment with trans-2-hexenal, suggesting that trans-2-hexenal could affect the synthesis of ergosterol (Figure 6C).
3.8. RT-qPCR
In order to determine the effect of trans-2-hexenal on β-1,3-glucan and ergosterol, the key biosynthesis genes related to β-1,3-glucan biosynthesis and ergosterol biosynthesis were selected for RT-qPCR analysis. As revealed in Figure 7, the expression levels of FKS1, ERG1, ERG7, and ERG11 were significantly lower than the control samples in the whole trans-2-hexenal treatment period.
3.9. Molecular Docking
Using Alphafold2, FKS1, ERG1, ERG7, and ERG11 were homology modeled; then, trans-2-hexenal was docked against the FKS1, ERG1, ERG7, and ERG11 protein, respectively (Figure 8). The binding energies of trans-2-hexenal to the best conformations of FKS1, ERG1, ERG7, and ERG11 were −4.9 kcal/mol, −4.6 kcal/mol, −4.1 kcal/mol, and −3.9 kcal/mol, respectively. Because of the different binding conformations of the ligand molecules in the conjugates, the amino acid residues around the active sites of their interactions and the strength of their interactions were different. trans-2-Hexenal formed a hydrophobic interaction with Ile1260, Gly1261, Ile1262, Leu1263, Leu1348, Tyr1587, Pro1589, Phe1868, and Leu1867 in FKS1, while the aldehyde group (-COH) of trans-2-hexenal acted as an acceptor to form a hydrogen bond with a bond length of 2.88 Å with the side chain of Gly1264 in FKS1; the binding of trans-2-hexenal to FKS1 with that hydrophobic interaction and hydrogen bonding promote the binding of trans-2-hexenal to FKS1. trans-2-Hexenal formed hydrophobic interactions with amino acid residues Pro399, Leu363, Leu403, Tyr81, Leu367, Val211, Ile83, Leu92, Phe37, and Phe389 in ERG1; His724, Ile725, Glu726, Glu736, Gln127, Tyr722, and Pro405 in ERG7; and Leu371, Val494, Gln490, Tyr493, Asn378, and Gln372 in ERG11. Hydrophobic interactions provide the main driving force for the combination of trans-2-hexenal with ERG1, ERG7, and ERG11, which results in altered interactions among amino acid residues within the protein molecule and plays an important role in maintaining protein conformation.
4. Discussion
G. citri-aurantii is considered one of the most important pathogens in terms of the economic losses that hinder citrus worldwide [2,18,19]. In the present study, trans-2-hexenal showed pronounced antifungal efficacy against G. citri-aurantii in vitro and in vivo. These results are consistent with those of previous studies with regard to the antifungal activity of trans-2-hexenal [9,10,13,15], indicating that the application of trans-2-hexenal is a potent method for controlling G. citri-aurantii.
The fungal cell wall is a dynamic structure that protects cells from osmotic pressure and other environmental stresses. It is of great significance to the growth of fungal cells. The destruction of the cell wall’s structure easily leads to the cell’s dissolution and death. Plant essential oils, such as tea tree essential oil, Hyssopus officinalis essential oil, cinnamaldehyde, and thymol, can destroy the cell wall of fungi [10,25,26]. Cinnamaldehyde mainly inhibits the growth of G. citri-aurantii by inhibiting chitin synthesis, accelerating chitin hydrolysis, and destroying the integrity of the cell wall [19]. In this study, trans-2-hexenal treatment did not change the chitin content and cell wall integrity, but the β-1,3-glucan content of G. citri-aurantii decreased significantly. These results are similar to the results of Li et al. [27] in which o-vanillin mainly reduced the β-1,3-glucan content in the cell wall of A. flavus, but the content of chitin did not change. FKS family genes (FKS1, FKS2, and FKS3) are the key genes that synthesize β-1,3-glucan synthase, and FKS1 is an essential gene, in most, that can regulate β-1,3-glucan synthase activity [28]. García et al. [29] found that the deletion of FKS1 in yeast would lead to a decrease in the glucan content and slow the growth of the strain. The RT-qPCR results show that trans-2-hexenal decreased the expression of FKS1, which is consistent with the decrease in the β-1,3-glucan content, indicating that the β-1,3-glucan synthesis pathway was inhibited. Inhibitors of β-1,3-glucan synthase affect gene expression by targeting key groups of synthases, which, in turn, leads to a decrease in the content of β-1,3-glucan in the cell wall, resulting in cell rupture and death [30]. Douglas et al. [31] found that echinocandin binds to the catalytic subunit of β-1,3-glucan synthase to reduce the integrity of the fungal cell wall. Molecular docking showed that the hydrogen bond and hydrophobic interaction between trans-2-hexenal and FKS1 lead to a change in the FKS1 protein’s structure, rotatable bond dihedral angle, and amino acid residue side chain, which reduces its expression and interferes with the normal function of FKS1. Trans-2-hexenal inhibited the synthesis of the cell wall by combining different amino acid residues with the active site of FKS1 in the cell wall of G. citri-aurantii, which is consistent with the antibacterial mechanism of eugenol and citral against P. roqueforti and A. niger [32]. The decrease in β-1,3-glucan content in the cell wall may lead to an increase in fungal cell wall permeability, as evidenced by the increase in extracellular AKP activity of fungi [26]. However, despite the decrease in β-1,3-glucan content, the cell wall became thicker and the activity of extracellular AKP activity remained unchanged, suggesting that trans-2-hexenal did not destroy the cell wall permeability of G. citri-aurantii but induced its cell wall components to restructure, resulting in a decrease in the β-1,3-glucan content.
The integrity and fluidity of the cell membrane are crucial for the survival and growth of fungi, and they are key points in many kinds of drug treatment [33]. The fungal cell membrane is the protective barrier of fungal cells, and it is selective for substances to enter and leave the cells. Therefore, damage to the integrity of the cell membrane will cause the leakage of components, leading to cell apoptosis [34,35]. Lipids are an important component of the cell membrane, and they play important roles in cell membrane integrity and material transportation [36,37]. Ergosterol, as a unique component of fungal cell membranes, plays important roles in cell membrane fluidity and membrane protein function [38,39,40]. It was found that the synthesis of ergosterol in A. flavus cell membranes was significantly inhibited after being treated with Artemisia annua essential oil; thus, the fungal cell membranes were the main site [41]. After being treated with trans-2-hexenal, the lipid content of G. citri-aurantii decreased at 60 min. The ergosterol content in the treatment group decreased at 30 min, indicating that ergosterol was the primary target of trans-2-hexenal in inhibiting G. citri-aurantii. ERG1, ERG7, and ERG11 are key genes in the sterol biosynthesis pathway, encoding squalene epoxidase, lanosterol synthase, and lanosterol-14-α-demethylase in the sterol biosynthesis pathway, respectively [19,42]. ERG1, ERG7, and ERG11 in G. citri-aurantii were significantly downregulated after trans-2-hexenal treatment, which is consistent with the results of the ergosterol content, further indicating that the ergosterol synthesis pathway was attacked. Meanwhile, it is worth noting that the molecular docking analysis of the formed complexes shows that the formation of hydrophobic interactions may lead to a longer residence time of trans-2-hexenal in ERG1, ERG7, and ERG11, and the hydrophobic interactions may enhance the effect of trans-2-hexenal on the ergosterol synthesis pathway. This result is consistent with that of Song et al. [43], who found that trans-2-hexenal fumigation reduced the ergosterol content of Botrytis cinerea and influenced ergosterol biosynthetic gene expression levels.
5. Conclusions
In summary, trans-2-hexenal reduced the β-1,3-glucan content and ergosterol content of G. citri-aurantii by inhibiting the expressions of related genes. With the prolongation of the treatment time, the total lipid content decreased, and the cell membrane integrity was further destroyed.
Author Contributions
Conceptualization, Q.O.; data curation, Q.O.; formal analysis, Q.O.; funding acquisition, Q.O.; investigation, Q.O., S.S., Y.L. and Y.Y.; methodology, S.S., Y.L., Y.Z., X.Y. and N.T.; supervision, Q.O.; writing—original draft, Q.O.; writing—review & editing, Q.O., N.T. and L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China, grant number 32001755, Scientific Research Fund of Hunan Provincial Education Department, grant number 22B0165, and Hunan Provincial Natural Science Foundation of China, grant number 2021JJ40533.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ferraz, L.P.; Cunha, T.D.; da Silva, A.C.; Kupper, K.C. Biocontrol ability and putative mode of action of yeasts against Geotrichum citri-aurantii in citrus fruit. Microbiol. Res. 2016, 188–189, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.X.; Feng, L.Y.; Sun, J.; Mao, L.T.; Li, X.J.; Jiang, Y.M.; Duan, X.W.; Li, T.T. Antifungal activities of a natural trisaccharide ester against sour rot in mandarin fruit. Postharvest Biol. Technol. 2022, 191, 111981. [Google Scholar] [CrossRef]
- McKay, A.H.; Förster, H.; Adaskaveg, J.E. Efficacy and application strategies for propiconazole as a new postharvest fungicide for managing sour rot and green mold of citrus fruit. Plant Dis. 2012, 96, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, D.Y.; Wang, Z.; Tian, Z.H.; Fan, Y.; Lu, X.J.; Long, C.A. Genome sequencing and transcriptome analysis of Geotrichum citri-aurantii on citrus reveal the potential pathogenic- and guazatine-resistance related genes. Genomics 2020, 112, 4063–4071. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yang, Y.; Zhu, X.; Yuan, P.; Shan, Y. Inhibitory mechanisms of cinnamic acid on the growth of Geotrichum citri-aurantii. Food Control 2022, 131, 108459. [Google Scholar] [CrossRef]
- Li, Y.H.; Shao, X.F.; Xu, J.Y.; Wei, Y.Y.; Xu, F.; Wang, H.F. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Wuryatmo, E.; Able, A.J.; Ford, C.M.; Scott, E.S. Effect of volatile citral on the development of blue mould, green mould and sour rot on navel orange. Australas. Plant Path. 2014, 43, 403–411. [Google Scholar] [CrossRef]
- Bazioli, J.M.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; Fill, T.P. Biological control of citrus postharvest phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Ouyang, Q.L.; Duan, B.; Reymick, O.O.; Chen, Y.; Tan, Y.Z.; Zhu, X.R.; Su, D.L.; Li, G.Y.; Tao, N.G. Trans-2-hexenal/beta-cyclodextrin effectively reduces green mold in citrus fruit. Postharvest Biol. Technol. 2022, 187, 111871. [Google Scholar] [CrossRef]
- Arroyo, F.T.; Moreno, J.; Daza, P.; Boianova, L.; Romero, F. Antifungal activity of strawberry fruit volatile compounds against Colletotrichum acutatum. J. Agric. Food Chem. 2007, 55, 5701–5707. [Google Scholar] [CrossRef]
- Myung, K.; Hamilton-Kemp, T.R.; Archbold, D.D. Interaction with and Effects on the profile of proteins of Botrytis cinerea by C6 aldehydes. J. Agric. Food Chem. 2007, 55, 2182–2188. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Fu, M.R.; Qu, X.Q.; Liu, J.J.; Bu, J.W.; Feng, S.R.; Zhao, H.D.; Jiao, W.X.; Sun, F. (E)-2-Hexenal-based coating induced acquired resistance in apple and its antifungal effects against Penicillium expansum. LWT-Food Sci. Technol. 2022, 163, 113536. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, H.; Sun, H.; Wang, X. Antifungal activity of trans-2-hexenal against Penicillium cyclopium by a membrane damage mechanism. J. Food Biochem. 2017, 41, e12289. [Google Scholar] [CrossRef]
- Dong, Y.P.; Li, C.Y.; Long, H.T.; Liu, Z.T.; Huang, Y.; Zhang, M.; Wang, T.L.; Liu, Y.X.; Bi, Y.; Prusky, D.B. Preparation and use of trans-2-hexenal microcapsules to preserve ‘Zaosu’ pears. Sci. Hortic. 2021, 283, 110091. [Google Scholar] [CrossRef]
- Guo, M.; Feng, J.; Zhang, P.; Jia, L.; Chen, K. Postharvest treatment with trans-2-hexenal induced resistance against Botrytis cinerea in tomato fruit. Australas. Plant Path. 2015, 44, 121–128. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, L.; Zhao, W.; Xie, Y.L. (E)-2-Hexenal, as a potential natural antifungal compound, inhibits Aspergillus flavus spore germination by disrupting mitochondrial energy metabolism. J. Agric. Food Chem. 2019, 67, 1138–1145. [Google Scholar] [CrossRef]
- Wu, Y.L.; Ouyang, Q.L.; Tao, N.G. Plasma membrane damage contributes to antifungal activity of citronellal against Penicillium digitatum. J. Food Sci. Technol. 2016, 53, 3853–3858. [Google Scholar] [CrossRef]
- Dou, S.W.; Ouyang, Q.L.; You, K.Y.; Qian, J.J.; Tao, N.G. An inclusion complex of thymol into β-cyclodextrin and its antifungal activity against Geotrichum citri-aurantii. Postharvest Biol. Technol. 2018, 138, 31–36. [Google Scholar] [CrossRef]
- Ouyang, Q.L.; Duan, X.F.; Li, L.; Tao, N.G. Cinnamaldehyde exerts its antifungal activity by disrupting the cell wall integrity of Geotrichum citri-aurantii. Front. Microbiol. 2019, 10, 55. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, K.; Yang, H.; Yuan, Y.H.; Yue, T.L. Antifungal mechanism of cinnamaldehyde and citral combination against Penicillium expansum based on FT-IR fingerprint, plasma membrane, oxidative stress and volatile profile. RSC Adv. 2018, 8, 5806–5815. [Google Scholar] [CrossRef]
- Francois, J.M. A simple method for quantitative determination of polysaccharides in fungal cell walls. Nat. Protoc. 2016, 1, 2995–3000. [Google Scholar] [CrossRef]
- Fortwendel, J.R.; Juvvadi, P.R.; Pinchai, N.; Perfect, B.Z.; Alspaugh, J.A.; Perfect, J.R.; Steinbach, W.J. Differential effects of inhibiting chitin and 1,3-β-D-glucan synthesis in ras and calcineurin mutants of Aspergillus fumigatus. Antimicrob. Agents Chemother. 2009, 53, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.L.; Okwong, R.O.; Chen, Y.P.; Tao, N.G. Citronellal exerts its antifungal activity by targeting ergosterol biosynthesis in Penicillium digitatum. J. Fungi 2021, 7, 432. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2021, 25, 402–408. [Google Scholar] [CrossRef]
- Ghfir, B.; Fonvieille, J.L.; Dargent, R. Influence of essential oil of Hyssopus officinalis on the chemical composition of the walls of Aspergillus fumigatus (Fresenius). Mycopathologia 1997, 138, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Cheng, S.; Wang, H.; Yu, D.; Mungai, C. The possible mechanism of antifungal action of tea tree oil on Botrytis cinerea. J. Appl. Microbiol. 2013, 11114, 1642–1649. [Google Scholar] [CrossRef]
- Li, Q.; Zhu, X.M.; Xie, Y.L.; Zhong, Y. o-Vanillin, a promising antifungal agent, inhibits Aspergillus flavus by disrupting the integrity of cell walls and cell membranes. Appl. Microbiol. Biotechnol. 2021, 105, 5147–5158. [Google Scholar] [CrossRef]
- Healey, K.R.; Paderu, P.; Hou, X.; Ortigosa, C.J.; Bagley, N.; Patel, B.; Zhao, Y.N.; Perlin, D.S. Differential regulation of echinocandin targets Fks1 and Fks2 in Candida glabrataby the Post-Transcriptional Regulator Ssd1. J. Fungi 2020, 4, 143. [Google Scholar] [CrossRef]
- García-Rodriguez, L.J.; Trilla, J.A.; Castro, C.; Valdivieso, M.H.; Durán, A.; Roncero, C. Characterization of the chitin biosynthesis process as a compensatory mechanism in the fks1 mutant of Saccharomyces cerevisiae. FEBS Lett. 2000, 478, 84–88. [Google Scholar] [CrossRef]
- Ma, L.; Salas, O.; Bowler, K.; Bar-Peled, M.; Sharon, A. UDP-4-keto-6-deoxyglucose, a transient antifungal metabolite, weakens the fungal cell wall partly by inhibition of UDP-galactopyranose mutase. MBio 2017, 8, e01559-17. [Google Scholar] [CrossRef]
- Douglas, C.M.; Foor, F.; Marrinan, J.A.; Morin, N.; Nielsen, J.B.; Dahl, A.M.; Mazur, P.; Baginsky, W.; Li, W.; el-Sherbeini, M. The Saccharomyces cerevisiae FKS1 (ETG1) gene encodes an integral membrane protein which is a subunit of 1,3-β-D-glucan synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 12907–12911. [Google Scholar] [CrossRef] [PubMed]
- Ju, J. Study on the Synergistic Inhibitory Mechanism of Eugenol and Citral against Penicillium Roqueforti and Aspergillus niger; Jiangnan University: Wuxi, China, 2021; pp. 27–35. [Google Scholar]
- Li, L.; Tang, X.; Ouyang, Q.L.; Tao, N.G. Combination of sodium dehydroacetate and sodium silicate reduces sour rot of citrus fruit. Postharvest Biol. Technol. 2019, 151, 19–25. [Google Scholar] [CrossRef]
- Liu, K.; Zhou, X.; Fu, M. Inhibiting effect of epsilon-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Sant, D.G.; Tupe, S.G.; Ramana, C.V.; Deshpande, M.V. Fungal cell membrane-promising drug target for antifungal therapy. J. Appl. Microbiol. 2016, 121, 1498–1510. [Google Scholar] [CrossRef]
- Segawa, K.; Nagata, S. An apoptotic ‘eat me’signal: Phosphatidylserine exposure. Trends Cell Biol. 2015, 25, 639–650. [Google Scholar] [CrossRef]
- Vartabedian, V.F.; Savage, P.B.; Teyton, L. The processing and presentation of lipids and glycolipids to the immune system. Immunol. Rev. 2016, 272, 109–119. [Google Scholar] [CrossRef]
- Dupont, S.; Lemetais, G.; Ferreira, T.; Cayot, P.; Gervais, P.; Beney, L. Ergosterol biosynthesis: A fungal pathway for life on land? Evolution 2012, 66, 2961–2968. [Google Scholar] [CrossRef]
- Krumpe, K.; Frumkin, I.; Herzig, Y.; Rimon, N.; Oezbalci, C.; Bruegger, B.; Rapaport, D.; Schuldiner, M. Ergosterol content specifies targeting of tail-anchored proteins to mitochondrial outer membranes. Mol. Biol. Cell 2012, 23, 3927–3935. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Gamarra, S.; Garcia-Effron, G.; Park, S.; Perlin, D.S.; Rao, R. Requirement for ergosterol in V-ATPase function underlies antifungal activity of azole drugs. PLoS Pathog. 2010, 6, e1000939. [Google Scholar] [CrossRef]
- Kumar, M.; Dwivedy, A.K.; Sarma, P.; Dkhar, M.S.; Kayang, H.; Raghuwanshi, R.; Dubey, N.K. Chemically characterised Artemisia nilagirica (Clarke) Pamp. essential oil as a safe plant-based preservative and shelf-life enhancer of millets against fungal and aflatoxin contamination and lipid peroxidation. Plant Biosyst. 2020, 154, 269–276. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces cerevisiae. mBio 2018, 9, e01291-18. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Du, S.L.; Sun, H.L.; Liang, Q.W.; Wang, H.H.; Yan, L.M.; Zhang, J.H. Antifungal mechanism of (E)-2-hexenal against Botrytis cinerea growth revealed by transcriptome analysis. Front. Microbiol. 2022, 13, 951751. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Disease progression in inoculated citrus fruit treated with trans-2-hexenal (0×, 1× and 10× MFC) during storage at 25 ± 2 °C and 80–85% RH. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 20).
Figure 1.
Disease progression in inoculated citrus fruit treated with trans-2-hexenal (0×, 1× and 10× MFC) during storage at 25 ± 2 °C and 80–85% RH. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 20).

Figure 2.
SEM images of (A,B) untreated control culture of G. citri-aurantii; (C,D) culture after incubation with the trans-2-hexenal.
Figure 2.
SEM images of (A,B) untreated control culture of G. citri-aurantii; (C,D) culture after incubation with the trans-2-hexenal.
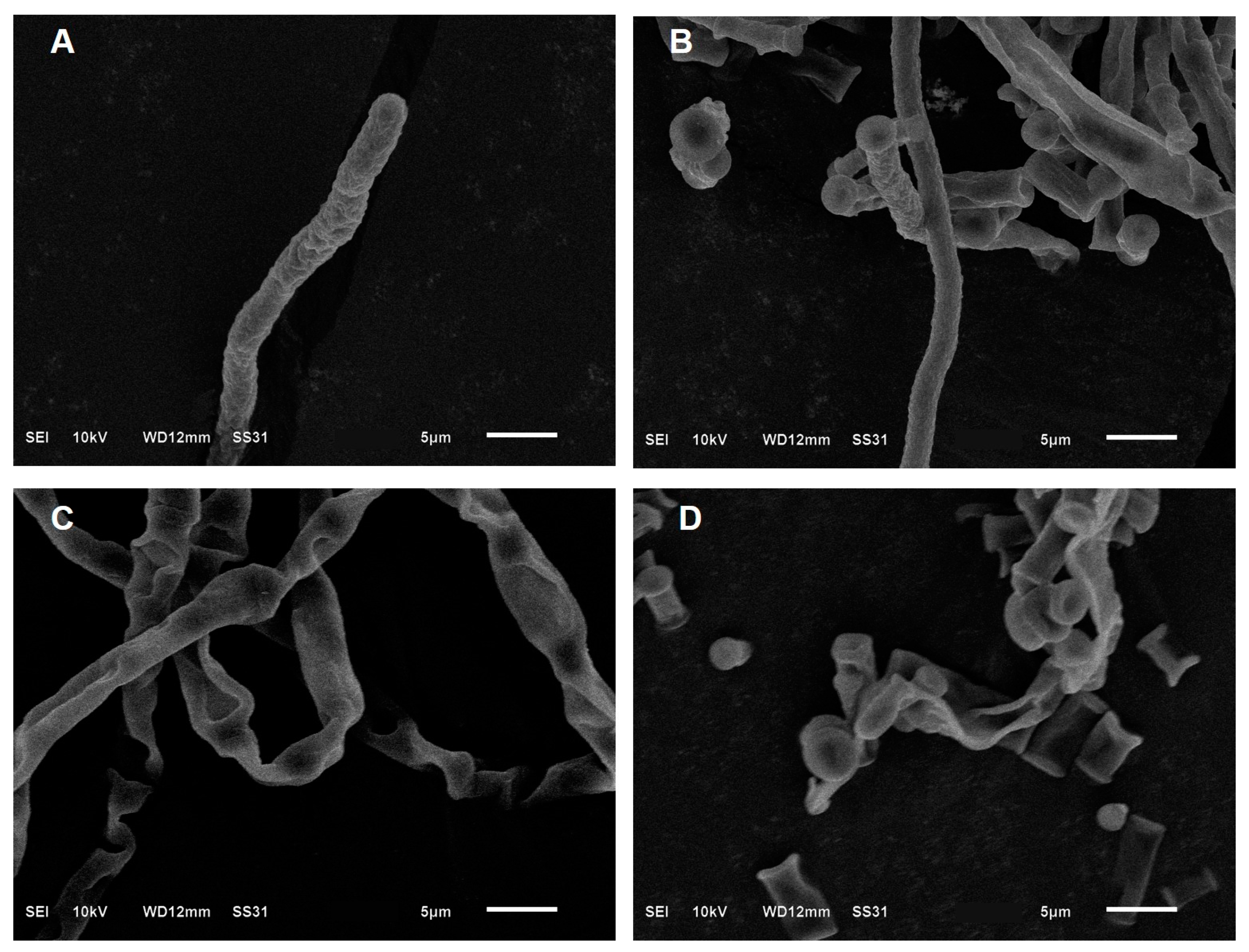
Figure 3.
TEM images of (A) untreated control culture of G. citri-aurantii; (B) culture after incubation with the trans-2-hexenal. CW: cell wall; CM: cell membrane; MIT: mitochondrion.
Figure 3.
TEM images of (A) untreated control culture of G. citri-aurantii; (B) culture after incubation with the trans-2-hexenal. CW: cell wall; CM: cell membrane; MIT: mitochondrion.
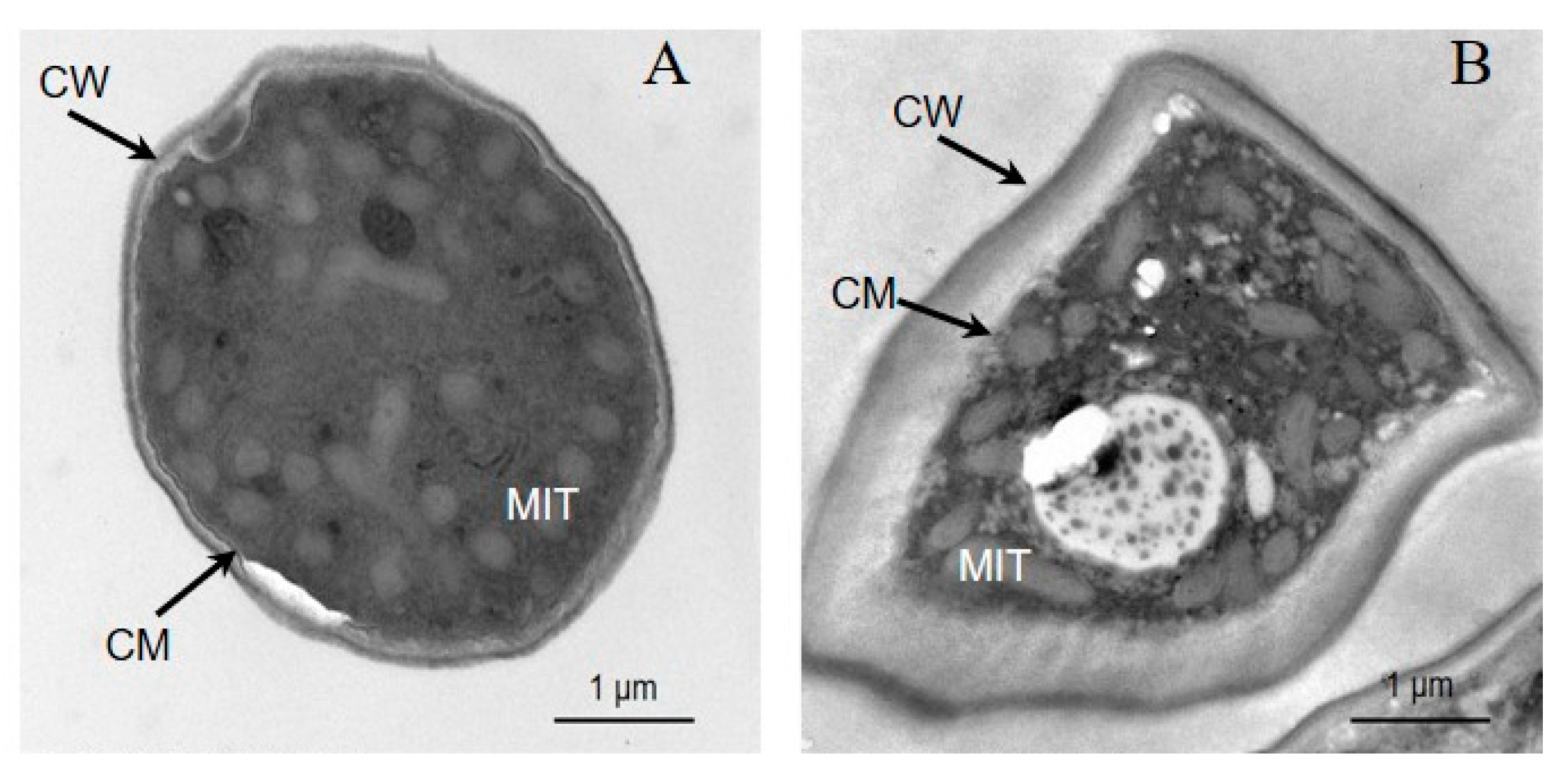
Figure 4.
FT−IR spectra analysis of G. citri-aurantii under trans-2-hexenal treatment. (A) FT−IR spectra of the second derivative (400–4000 cm−1); (B) the second derivative of FT−IR spectra of the nucleic acid and polysaccharide region (900−1200 cm−1); (C) the second derivative of FT−IR spectra of the mixed region of fatty acid, protein, and polysaccharide (1200−1500 cm−1); (D) the second derivative of FT−IR spectra of the mixed region (1500−1700 cm−1); (E) the second derivative of FT−IR spectra of the lipid region (2800−3100 cm−1).
Figure 4.
FT−IR spectra analysis of G. citri-aurantii under trans-2-hexenal treatment. (A) FT−IR spectra of the second derivative (400–4000 cm−1); (B) the second derivative of FT−IR spectra of the nucleic acid and polysaccharide region (900−1200 cm−1); (C) the second derivative of FT−IR spectra of the mixed region of fatty acid, protein, and polysaccharide (1200−1500 cm−1); (D) the second derivative of FT−IR spectra of the mixed region (1500−1700 cm−1); (E) the second derivative of FT−IR spectra of the lipid region (2800−3100 cm−1).

Figure 5.
The effects of trans-2-hexenal on the cell walls of G. citri-aurantii. (A) The chitin content of G. citri-aurantii; (B) the β-1,3-glucan content of G. citri-aurantii; (C) the extracellular AKP activity of G. citri-aurantii. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a” indicated that there was no difference between different treatment groups (p < 0.05).
Figure 5.
The effects of trans-2-hexenal on the cell walls of G. citri-aurantii. (A) The chitin content of G. citri-aurantii; (B) the β-1,3-glucan content of G. citri-aurantii; (C) the extracellular AKP activity of G. citri-aurantii. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a” indicated that there was no difference between different treatment groups (p < 0.05).

Figure 6.
The effects of trans-2-hexenal on the cell membranes of G. citri-aurantii. (A) The plasma membrane integrity of G. citri-aurantii mycelia and mycelia fluorescence times; (B) the total lipids contents and (C) ergosterol contents of G. citri-aurantii mycelia. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a,b” indicates the difference among different treatment groups (p < 0.05).
Figure 6.
The effects of trans-2-hexenal on the cell membranes of G. citri-aurantii. (A) The plasma membrane integrity of G. citri-aurantii mycelia and mycelia fluorescence times; (B) the total lipids contents and (C) ergosterol contents of G. citri-aurantii mycelia. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a,b” indicates the difference among different treatment groups (p < 0.05).
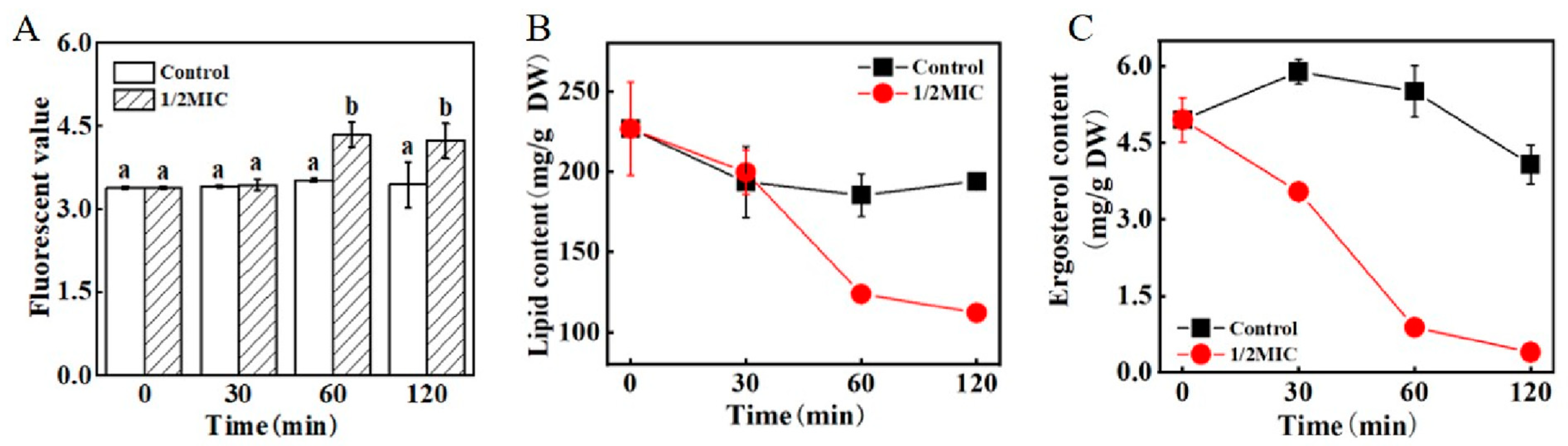
Figure 7.
Changes in the expression of the β-1,3-glucan and ergosterol biosynthesis genes of G. citri-aurantii by control and 1/2 MIC of trans-2-hexenal for 0, 30, 60, and 120 min ((A) FKS1; (B) ERG1; (C) ERG7; (D) ERG11). (E) Ergosterol biosynthesis pathway. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a,b” indicates the difference among different treatment groups (p < 0.05).
Figure 7.
Changes in the expression of the β-1,3-glucan and ergosterol biosynthesis genes of G. citri-aurantii by control and 1/2 MIC of trans-2-hexenal for 0, 30, 60, and 120 min ((A) FKS1; (B) ERG1; (C) ERG7; (D) ERG11). (E) Ergosterol biosynthesis pathway. The data presented are the means of pooled data. Error bars indicate the SDs of the means (n = 3). “a,b” indicates the difference among different treatment groups (p < 0.05).
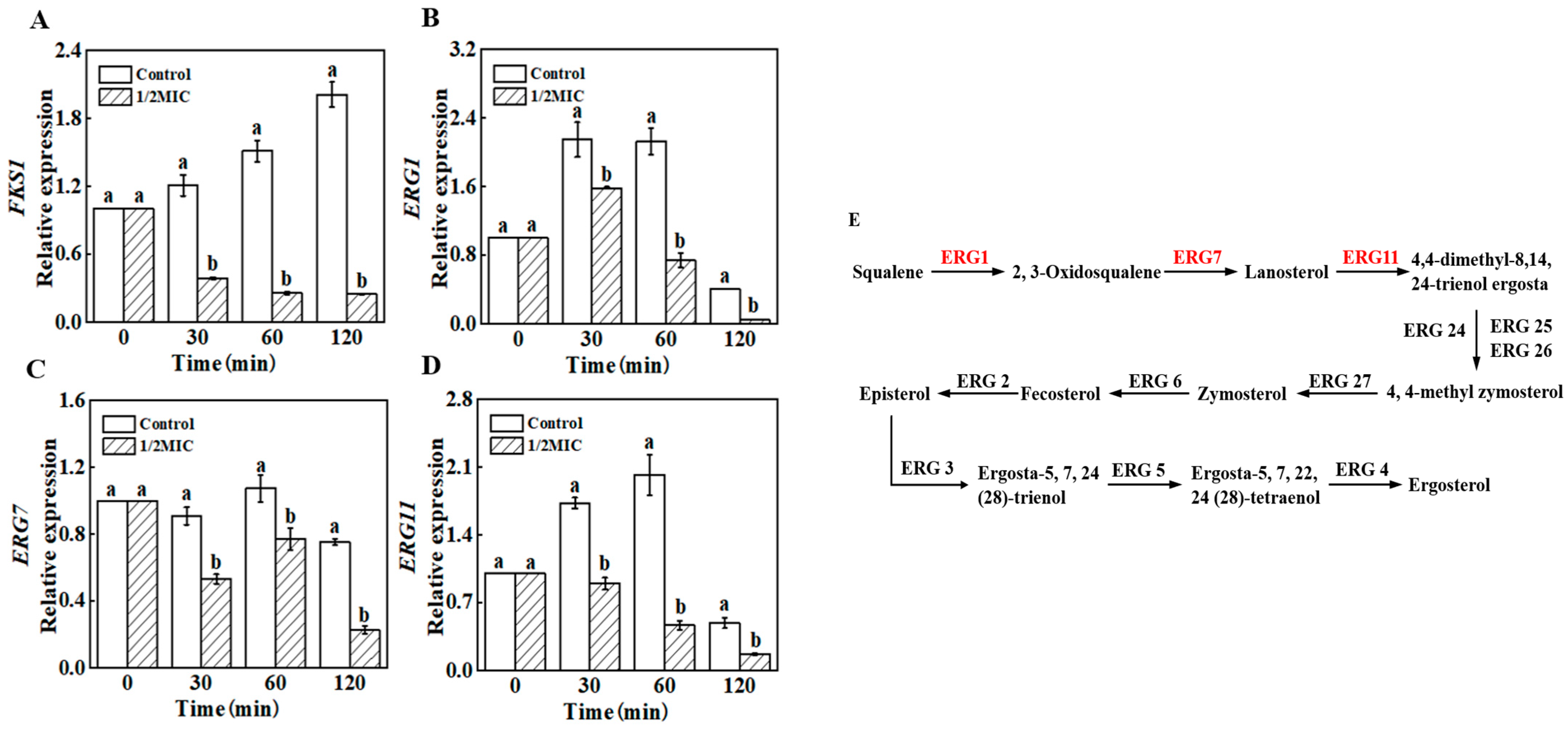
Figure 8.
The 3D images of FKS1 (A), ERG1 (B), ERG7 (C), and ERG11 (D) proteins and the 2D interaction diagram of trans-2-hexenal with FKS1 (A), ERG1 (B), ERG7 (C), and ERG11 (D) proteins. The dark green rod-like structure in the diagram is trans-2-hexenal, and the other color rod-like structures are amino acid residues.
Figure 8.
The 3D images of FKS1 (A), ERG1 (B), ERG7 (C), and ERG11 (D) proteins and the 2D interaction diagram of trans-2-hexenal with FKS1 (A), ERG1 (B), ERG7 (C), and ERG11 (D) proteins. The dark green rod-like structure in the diagram is trans-2-hexenal, and the other color rod-like structures are amino acid residues.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer pair sequences designed for validation of differentially expressed genes in control and 1/2 MIC trans-2-hexenal treatment of G. citri-aurantii using RT-qPCR.
Table 1.
Primer pair sequences designed for validation of differentially expressed genes in control and 1/2 MIC trans-2-hexenal treatment of G. citri-aurantii using RT-qPCR.
| Gene ID | Genes | Primer Sequence (5′-3′) |
|---|---|---|
| CL1729.Contig1_All | FKS1-F | AGGTTGAAGGCAAGCGTACTCT |
| FKS1-R | CAGGAAGTGGCTCAGGAATAGGT | |
| Unigene4828_All | ERG1-F | AAGTCCTACACCTCCAAGGCTAC |
| ERG1-R | GAATATCGGCGTCAGTGAGAACC | |
| Unigene3819_All | ERG7-F | TAACGCATATCCAGGACGACCAA |
| ERG7-R | CGCACAATCTCAATTCGCTCTTC | |
| Unigene3920_All | ERG11-F | CGCCGTAAGGAAGGAAACATTGA |
| ERG11-R | AAGACGAAGTAGCAGCCGAAGT | |
| CL313.Contig2 | Actin1-F | TTACGCCGGTTTCTCCCTCC |
| Actin1-R | GACGATTTCACGCTCGGCAG |
Table 2.
Antifungal activity of trans-2-hexenal against G. citri-aurantii.
| Concentration (µL/mL) | Inhibitory Rate (%) | ||||
|---|---|---|---|---|---|
| 1 d | 2 d | 3 d | 4 d | 5 d | |
| 0.25 | 100.0 ± 0.0 a | 53.7 ± 5.2 b | 24.8 ± 1.7 c | 27.1 ± 3.0 c | 23.2 ± 1.4 c |
| 0.50 | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 90.5 ± 1.7 b | 72.9 ± 3.9 b | 64.0 ± 8.7 b |
| 1.00 | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a |
| 2.00 | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a |
| 4.00 | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 100.0 ± 0.0 a |
Note: Mean values ± SD (standard deviation) followed by different letters (a–c) represent significantly different scores in the same phase (p < 0.05).
Table 3.
Effect of trans-2-hexenal on the incidence of Satsuma fruit inoculated with G. citri-aurantii.
Table 3.
Effect of trans-2-hexenal on the incidence of Satsuma fruit inoculated with G. citri-aurantii.
| Treatments | Incidence Rate (%) | ||||||
|---|---|---|---|---|---|---|---|
| 1 d | 2 d | 3 d | 4 d | 5 d | 6 d | 7 d | |
| Control | 0 ± 0 a | 11 ± 4 a | 22 ± 4 a | 49 ± 4 a | 73 ± 0 a | 89 ± 4 a | 100 ± 0 a |
| 1× MFC trans-2-hexenal | 0 ± 0 a | 0 ± 0 b | 16 ± 4 b | 22 ± 8 b | 60 ± 7 b | 80 ± 0 b | 85 ± 4 b |
| 10× MFC trans-2-hexenal | 0 ± 0 a | 0 ± 0 b | 0 ± 0 c | 9 ± 0 c | 22 ± 8 c | 29 ± 8 b | 33 ± 12 c |
Note: “a–c” indicates the difference among different treatment groups with the same storage time (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ouyang, Q.; Shi, S.; Liu, Y.; Yang, Y.; Zhang, Y.; Yuan, X.; Tao, N.; Li, L. Inhibitory Mechanisms of trans-2-Hexenal on the Growth of Geotrichum citri-aurantii. J. Fungi 2023, 9, 930. https://doi.org/10.3390/jof9090930
AMA Style
Ouyang Q, Shi S, Liu Y, Yang Y, Zhang Y, Yuan X, Tao N, Li L. Inhibitory Mechanisms of trans-2-Hexenal on the Growth of Geotrichum citri-aurantii. Journal of Fungi. 2023; 9(9):930. https://doi.org/10.3390/jof9090930
Chicago/Turabian StyleOuyang, Qiuli, Shiwei Shi, Yangmei Liu, Yanqin Yang, Yonghua Zhang, Xingxing Yuan, Nengguo Tao, and Lu Li. 2023. "Inhibitory Mechanisms of trans-2-Hexenal on the Growth of Geotrichum citri-aurantii" Journal of Fungi 9, no. 9: 930. https://doi.org/10.3390/jof9090930
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.