
Recent Advances in Molecular Genetics of Onion


1
Department of Plant and Environmental Sciences, New Mexico State University, Las Cruces, NM 88003, USA
2
Division of Vegetable Science, ICAR—Indian Agricultural Research Institute, New Delhi 110012, India
3
Department of Vegetable Science, Punjab Agricultural University, Ludhiana 141004, India
4
College of Integrated Sciences and Arts—School of Applied Sciences and Arts, Arizona State University, Mesa, AZ 85212, USA
5
Department of Horticultural Sciences, Texas A&M AgriLife Research and Extension Center, Uvalde, TX 78801, USA
*
Author to whom correspondence should be addressed.
Horticulturae 2024, 10(3), 256; https://doi.org/10.3390/horticulturae10030256
Submission received: 16 February 2024
/
Revised: 29 February 2024
/
Accepted: 2 March 2024
/
Published: 7 March 2024
(This article belongs to the Section Genetics, Genomics, Breeding, and Biotechnology (G2B2))
Abstract
:Onion is an important vegetable crop because it adds nutritional value and diversity to food preparation. Understanding recent advancements in onion molecular genetics is essential to improve production, quality, and disease resistance. Cutting-edge genomic technologies like genetic mapping and RNA sequencing reveal important genes and pathways. The review examines the progress in utilizing various molecular markers to study genetic divergence. The exploration extends to understanding the genes and pathways responsible for bulb color and chemical composition and the genetic factors influencing bulbing, flowering, and vernalization. Additionally, the article explores quantitative trait loci associated with resistance to major damaging diseases and delves into the role of different loci in male sterility and hybrid development. The recent publication of the whole genome sequence of onions will lead to further identification of genes and understanding their roles and functions in metabolic pathways.
1. Introduction
Bulb onion (Allium cepa L.) is a widely cultivated and consumed vegetable worldwide. Onions are rich in various chemical compounds, including organosulfur compounds [1,2], phenolics [3,4], polysaccharides [5], and saponins [6,7]. An onion bulb possesses remarkable health benefits, including antioxidant [8], antimicrobial [9], anti-inflammatory [10], anti-obesity [11], anti-diabetic [12], anticancer [13], cardiovascular protective [14], neuroprotective [15], hepatorenal protective [16], respiratory protective [17], digestive system protective [18], reproductive protective [19], and immunomodulatory properties [20].
Conventional breeding involves a selection process in which plants are selected based on their superior phenotypic performance for the trait of interest. The most popular conventional plant breeding strategies include pure-line selections, mass selection, backcross, recurrent selection, and hybridization [21]. Regarding the pace of crop improvement, this breeding method is characterized by a lengthier process and an over-reliance on plant phenotype [22,23]. However, plant phenotype is influenced by various external factors (Genotype × Environment interaction). Therefore, relying only on plant phenotypic performance for selection can sometimes be less accurate. Consequently, plant breeders initiated the integration of different branches of biology in plant breeding to develop modern breeding methods. Plant breeding techniques have moved towards molecular tools, less affected by the environment. Modern crop improvement tools include DNA and RNA sequencing, gene mapping, and marker-assisted selection (MAS). These tools offer significant advantages, including speeding up the breeding process and making it more precise and efficient [24]. Conventional plant breeding approaches have been used to improve onion yield, quality, and resistance against biotic and abiotic stresses. Nevertheless, its biennial life cycle, cross-pollinated nature, and significant inbreeding depression have posed considerable challenges in breeding for improved traits. With the help of modern plant breeding practices, onion breeders can rapidly develop molecular markers and use them to map important onion traits and germplasm selection.
Challenges in onion breeding have made molecular markers highly attractive for breeding programs [25]. The MAS method uses different DNA markers to determine the genomic regions responsible for expressing desired plant traits [26]. Earlier, isozyme markers were utilized in onion breeding, which were later replaced with DNA-based markers such as random amplified polymorphic DNA (RAPD), restriction fragment length polymorphism (RFLP), and amplified fragment length polymorphism (AFLP). Due to their low polymorphism and poor reproducibility [27,28,29], they were replaced with simple sequence repeat (SSR) markers because they are codominant, highly variable, and reproducible. Single-nucleotide polymorphism (SNP) markers, as the name suggests, are based on a single base pair variation, and they are preferred over SRR markers because of their higher density and polymorphism. The SNP markers have recently been validated across a wide range of germplasms and mapped on existing linkage maps [30]. Further, other Allium species have been exploited to develop alien monosomic addition lines (AMALs), which are beneficial for increasing genetic diversity in cultivated species and for comparative genetic studies.
The first onion draft genome assembly was published in 2021. Researchers achieved a milestone by assembling the large genome (16 Gb) of a doubled haploid onion line DHCU066619, resulting in a 14.9 Gb genome, of which 2.4 Gb were organized into eight pseudomolecules [31]. Within this genome, approximately 72.4% comprises repetitive sequences, predominantly (retro)transposons. Gene prediction revealed an unexpectedly large number of putative gene models, totaling 540,925, with 47,066 models supported by RNA sequencing data. Genes were uniformly distributed across the genome without any gene-rich regions. Comparative analysis with garlic (A. sativum) revealed both collinearity and significant rearrangements, providing insights into the evolutionary relationship between these two species. This genome assembly represents a significant advancement in onion research, offering a valuable resource for further exploration and identification of genes influencing different traits. It is important to note that this assembly is primarily based on Illumina short-read sequencing, covering approximately 91% of the expected genome size. However, further refinement and improvement are still necessary.
This review discusses recent progress in onion genomics and molecular genetics, exploring genetic diversity within the germplasm for selection, and the potential for enhancing onion yield, quality, and disease-resistance attributes. The article presents an overview of important onion traits currently being studied and improved using molecular techniques. The traits include bulb color, bulb quality traits, bulbing, flowering, vernalization, disease resistance, and male sterility.
2. Use of Molecular Markers to Study Genetic Divergence
Higher genetic variation within the germplasm makes plants adaptable to environmental fluctuations, ensuring their survival and resilience [32,33]. Earlier in the last decade, most of the genetic diversity studies in the major onion-growing regions of the world were conducted using SSR markers. The study materials were either composed of local populations or, in some cases, of a limited number of commercial cultivars.
In Europe, numerous studies utilized SSR markers to study the genetic diversity of landrace cultivars besides the commercial ones. Mitrova et al. [34] studied the genetic diversity of commercial onion cultivars in the Czech Republic using fifteen polymorphic microsatellite SSR markers. An association study revealed non-specific groupings among the cultivars. However, no groupings were established based on important traits, such as bulb shape or maturity type. Likewise, SSR markers could not establish any clustering between bulb color and shape in a population-level genetic diversity analysis comprising Turkish onion landraces, breeding lines, and commercial cultivars [35]. This study utilized expressed sequence tag-derived SSR (EST-SSR) and genomic SSR (gSSR) markers. González–pérez et al. [36] evaluated the genetic diversity of seventeen onion landraces of Northwest Spain using twelve EST–SSR markers. Two important quality traits were assessed: soluble solids content and pungency. This study’s high heterozygosity and low allele number indicated outcrossing in a smaller geographical area. Additionally, this study highlighted that the geographical separation of landraces was not the determining factor for grouping onions. Another study of important landrace onions in the Galicia region of Northwest Spain utilized gSSR and EST-SSR markers [37]. These landraces were compared with European (Portugal, France, Italy, Germany, Hungary, Bulgaria, Netherlands, and the United Kingdom) and Spanish (northern, central, and southern Spain) accessions. In this study, 121 alleles with an average of 6.05 alleles per marker locus were obtained. The average numbers of alleles per landrace were 52.1, 48, and 51.9 for the Galician, Spanish, and European landraces, respectively. EST-SSR markers showed higher percentages of amplification, while gSSR markers displayed a higher polymorphism rate. Unlike the non-geographical separation of González–pérez et al. [36], Galician landraces formed a distinctive group, geographically separating European and Spanish landrace groups [37]. However, no correlation was found between their origin and the morpho-agronomic characters. The Galician landraces also showed considerable genetic variation regarding alleles per locus despite originating in a smaller region, indicating allele fixation due to local selections and heterogeneity. Karić et al. [38] also found significant genetic variability in local onion cultivars of Bosnia and Herzegovina, which were characterized by seven SSR markers. An Italian-originated ‘Majski Srebrenjak onion’ formed a separate cluster, whereas the ‘Konjic onion’, a landrace collected from the Buturović polje region, shared genetic similarity with a commercial cultivar, Ptujska rdeća. Ricciardi et al. [39] used SSR markers to assess the genetic variation in culturally and economically important “Acquaviva red onion” (ARO) to differentiate it from two other landraces, viz., South Italian landraces, “Tropea red onion” (TRO) and the “Montoro copper onion” (MCO). In the study, 320 individuals of ARO were evaluated for soluble solid content and pungency (pyruvic acid content). Fifty-five alleles were identified, with an average of five alleles per locus. A low Wright’s fixation index (FIS) value indicated high inbreeding depression in the ARO population in Apulia, southern Italy, which was noted as the center of diversity for many horticultural species. A genetic relationship study distinguished the ARO landrace from the other two landraces, TRO and MCO. Interestingly, the mean polymorphic information content (PIC) value, which indicates individual marker efficiency, was 0.48, more than the SNP markers in a similar study, which was ineffective in distinguishing the landraces [40].
SSR markers were also used for the cross-species transferability studies between Allium species. Adeyemo et al. [41] studied the molecular diversity, genetic relationship, and cross-species transferability of seventeen races from three Allium species, namely, A. cepa, A. sativum, and A. fistulosum, grown in different states of Nigeria with RAPD and EST-SSR markers. The bulb onion, A. cepa, was moderately diverse compared to two related Allium species. A lower level of diversity was found among the local landraces with a high degree of inbreeding. Using SSR markers, Jayaswall et al. [42] studied genetic relatedness between Allium germplasms from the gene bank in Pune, India. Eight popular cultivars of bulb onion were characterized along with related species (A. sativum, A. tuberosum, A. fistulosum, A. cepa var. Aggregatum, A. altaicum, and A. ampeloprasum) using thirty polymorphic SSR markers. A low cross-transferability detected by these SSR markers makes them suitable for sustainable management and phylogenetic studies of Allium spp. As expected, the genetic diversity was higher among the wild relatives of Allium than the cultivated species.
Several onion diversity studies used inter-simple sequence repeat (ISSR) markers, such as a genetic diversity study in Uruguay by Monteverde et al. [43], who used ISSR markers to assess twenty-seven landrace onion populations and two commercial cultivars from the Germplasm Bank Facultad de Agronomı’a. As we observed in the previous studies, no significant differences were found regarding their geographical regions or growing cycles. Moreover, genetic variability was observed within populations rather than between populations. Interestingly, this study revealed indirect selections for earlier growing cycles without changing the genetic backgrounds of the commercial cultivars or landraces and unintentional cross-pollination due to gene flow between different maturity groups of onion. Like the SSR markers, ISSR markers also revealed genetic diversity and interspecific relationships between Allium species. Samiei et al. [44] used ISSR markers to divide the Allium germplasm from northeast Iran into four sub-genera: Allium and Melanocrommyum (Webb & Berthel.) Rouy, Cepa (Mill.) Radić, and Reticulatobulbosa (Kamelin) N. Friesen. The highest and lowest variability levels were found in Allium kuhsorkhense and A. stipitatum, respectively. Genetic diversity studies of West and South Indian onion accessions by ISSR markers revealed moderate to high genetic diversity. The authors determined the presence of novel genetic components based on the accessions’ morphological, biochemical, and molecular characterizations and sub-clustering patterns [45,46].
In recent years, SNP markers have gained attention in onion genetic diversity studies. They are preferred over the other markers due to their hyper-variability and multi-allelic nature. Additionally, they can be used in high-throughput automation to genotype many samples simultaneously. Villano et al. [40] used 400 ETS-derived SNP markers to assess the genetic diversity of seventy-three onion (A. cepa) accessions that include landraces, commercial cultivars, and wild species, viz., A. ascalonicum, A. fistulosum, A. altaicum, A. galanthum, and A. schoenoprasum. Even though 72–75% of the markers were informative, moderate loci information was generated. These markers were mapped using a high-throughput kompetitive allele-specific polymerase chain reaction (KASP PCR) technology. The genotypes were divided into five groups, with the wild species (A. galanthum, A. altaicum, A. schoenoprasum, and A. fistulosum) in one group. Even though the study separated the collections geographically, it did not reveal any relationship between the origin of the genotype and markers. The authors also found that the germplasm diversity could be related to the allelic divergence from landraces and the genetic drift due to local environmental influence. SNP genotyping identified seventy-four genes associated with the three phenotypes, bulbing photoperiod (short, intermediate, and long day), bulb color, and bulb shape, fifty-three co-inherited pairs, and three loci with potential roles in bulbing and flowering regulation pathways in onions.
Fang et al. [47] performed a genome-wide assessment study on 122 accessions from twenty-three agro-climatic zones of China. Nine economic traits of onion bulbs and pseudostems were evaluated with 162,321 high-quality SNP markers via the genome-wide specific length amplified fragment (SLAF) sequencing method. Most of the onion species were A. fistulosum L. var. viviparum Makino and A. fistulosum L. var. gigantum Makino, A. mongolicum Regel, and A. cepa L. The SLAF sequencing method was very effective compared to other high-throughput genotyping technologies, e.g., restriction site-associated DNA sequencing (RAD-seq), in finding higher number and quality SNPs using fewer accessions with high genotyping accuracy. The study revealed that, even though A. fistulosum L. var. gigantum Makino, A. mongolicum Regel, and A. cepa L. all diverged from A. fistulosum var. viviparum, some regional accessions of var. gigantum were evolutionarily very close to var. viviparum. This study also identified differences within A. fistulosum var. viviparum due to climate and altitude.
3. Bulb Color
Onion bulb color is the most important trait from a consumer’s point of view and is used to classify onion cultivars. Thousands of low-molecular-weight organic compounds produced by plant cells are categorized into three overlapping groups based on their functions: primary metabolites, secondary metabolites, and hormones [48]. Flavonoids are one of the most abundantly (>9000) found secondary metabolites in plants, consisting of chalcones, flavones, flavonols, anthocyanins, and proanthocyanidins [49]. Among these flavonoids, anthocyanins are associated with red, blue, and purple color pigmentation of flowers, fruit, leaves, and seeds in different vegetables, legumes, and cereals [50,51]. Flavonoids are also associated with health benefits that can reduce the risk of cardiovascular disorders [52], cancer [53,54], obesity, and diabetes [55], and enhance the total antioxidant capacity of plasma [56].
Bulb color is influenced by intricate genetic interactions among many genes involved in the flavonoid biosynthesis pathway. Further, due to variations in the structural and regulatory genes of the pathway, the color of onion bulbs can vary, ranging from red to white, yellow, golden, pink, and chartreuse [57]. Five major loci (I, C, G, R, and L) are involved in the inheritance of onion bulb color. Dark-yellow and red bulb colors are dominant over white [58]. An inhibiter (I) locus, which is dominant, when found homozygous dominant (II), imparts a white color to the bulbs. Heterozygous (Ii) and homozygous recessive (ii) genotypes at the I locus condition a light-yellow or buff color [59,60]. A basic color factor (C) locus requires a homozygous dominant (CC) or heterozygous (Cc) genotype to develop red or yellow bulbs, while homozygous recessive (cc) genotypes are supposed to produce white bulbs. Two complementary loci, R and L, regulate red color. Dominant alleles at the R locus condition red bulb color, while homozygous recessive (rr) genotypes exhibit yellow color [60]. Multiple crosses between North American and Brazilian yellow onions rarely produced red onion bulbs [61], which was recognized and explained by the researchers [62] as an interaction of two complementary loci, the R and the L locus. Another locus (L2) linked to L has been recognized by Khar et al. [63] and Duangjit et al. [64] at which a dominant allele interacts with the R locus to condition red color (R_L_) to bulbs. The homozygous dominant (GG) and heterozygous (Gg) genotypes at the G locus impart a golden-yellow color; however, homozygous recessive (gg) genotypes at the G locus condition chartreuse/light-green bulb color [62]. The epistatic interaction of these loci is associated with the biosynthesis pathway of flavonoids. Understanding genes responsible for flavonoids’ metabolic pathways, their interaction with the environment, and the contribution of physiological processes for activating desired genes is critical for regulating these phytochemicals. The genes coding enzymes for the biosynthesis of anthocyanins have been studied in model plants such as Arabidopsis thaliana L. [65], Zea maize L. [66], and Antirrhinum majus L. [67]. Multiple structural and regulatory genes involved in controlling the expression of the anthocyanin biosynthesis pathway have been identified and isolated using mutant color phenotypes in these model plant species [67]. The majority of reactions of the anthocyanin biosynthesis pathway in Arabidopsis thaliana L., Zea maize L., and Antirrhinum majus L. are the same. However, there are important differences between the different types of anthocyanins produced by these model plants, and the extent of modification varies for each plant species. The sequence information of genes gathered from model plant species and information accumulated from classical genetics underpin the genetic basis of different loci responsible for different bulb colors and anthocyanin biosynthesis pathways in onions [60,62,68] (Figure 1).
The metabolic pathway for anthocyanin synthesis in onions begins with the amino acid phenylalanine, converted into cinnamic acid by the enzyme phenylalanine ammonia-lyase (PAL). Chalcone synthase (CHS) catalyzes cinnamic acid into chalcone, produced by one molecule of 4-coumaroyl CoA and three molecules of malonyl CoA. Then, chalcone is isomerized to colorless naringenin by chalcone isomerase (CHI). Naringenin is then converted into dihydrokaempferol by flavanone 3-hydroxylase (F3H). Flavonoid 3′,5′-hydroylase (F3′5′H), and flavonoid 3′-hydroxylase (F3′H) then catalyze the hydroxylation of dihydrokaempferol (DHM) to produce dihydromyricetin and dihydroquercetin, respectively. Next, dihydroflavonols are reduced to leucocyanidin by dihydroflavonol 4-reductase (DFR), and during this process, flavonol synthase (FLS) catalyzes the production of quercetin, kaempferol, and myricetin. Anthocyanidin synthase (ANS) further catalyzes the production of cyanidin, and, consequently, UDP-flavonoid glucosyltransferase (UFGT) converts cyanidin into anthocyanins such as cyanidin-3 glucoside [69,70]. Different loci associated with bulb color in onion are structural genes encoding for various key enzymes in the biosynthesis pathway. The gene encoding for CHS [71] is involved in the early steps of the flavonoid biosynthesis pathway in onions. A candidate gene approach was implemented to identify the function of the C locus in the biosynthesis pathway of anthocyanins [72]. Differential gene expression was studied for four different bulb colors in F2, segregating populations developed by crossing two yellow and two red doubled haploid lines. Transcription was significantly reduced for two homologous CHS genes (CHS-A and CHS-B) in dominant and recessive white onions, suggesting that the CHS might be the possible control point for the biosynthesis pathway of anthocyanins. SNP markers tagging different alleles of CHS-A and CHS-B did not co-segregate, suggesting the causal gene for the C locus to be a regulatory gene controlling the expression of the CHS. Moreover, CHI plays a critical role in the anthocyanin biosynthesis pathway because its inactivation leads to the blockage of the accumulation of chalcone derivatives, contributing to the yellow pigment in onion bulbs [73].
Figure 1.
An overview of the anthocyanin biosynthetic pathway in onions. * Hypothesized locus I position in the pathway [68,73,74,75,76,77].
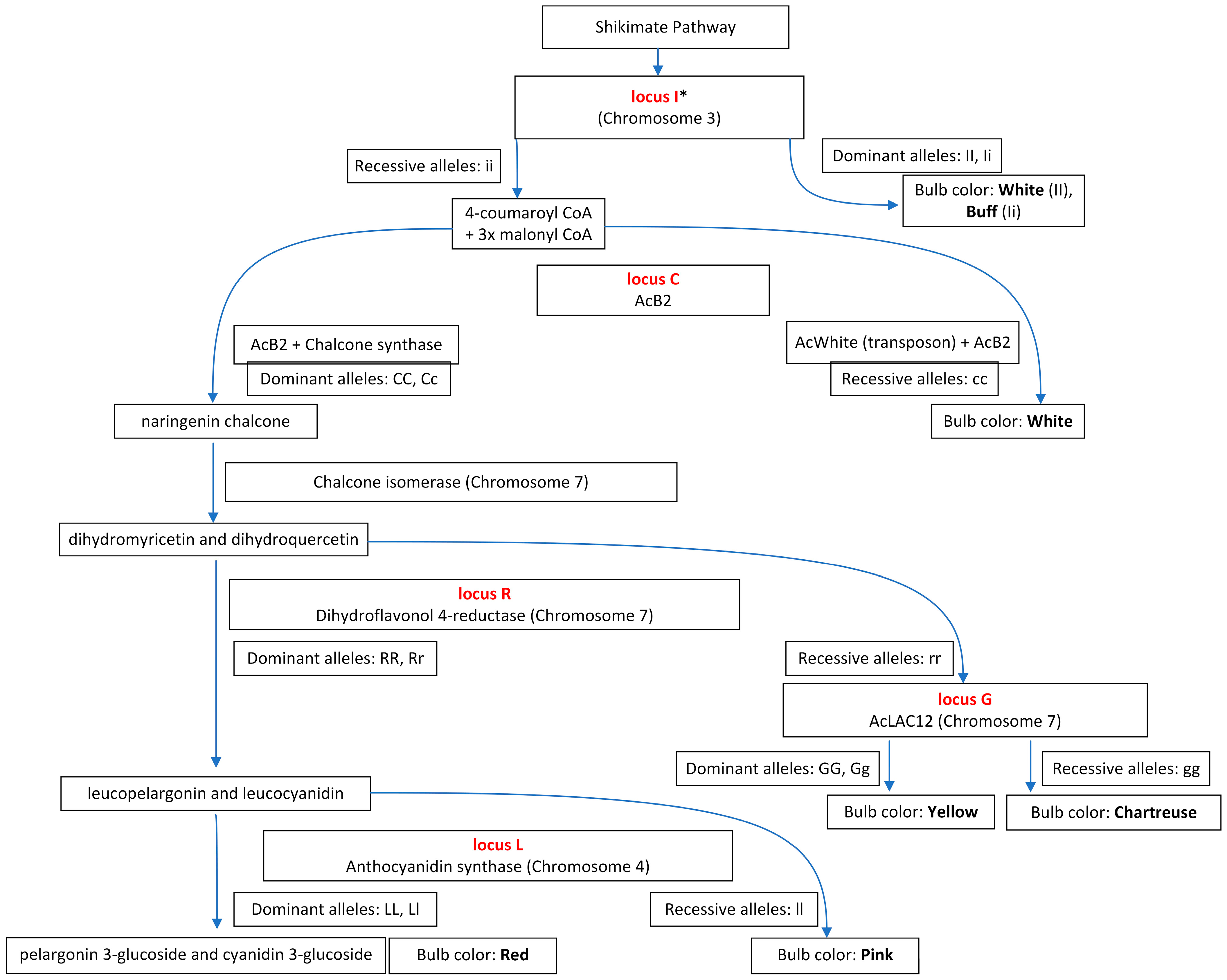
The DFR gene expressed only in red onion is associated with the R locus [74], which complements the L locus encoding ANS involved later in the anthocyanin biosynthesis pathway [68]. The F3 population was derived from a cross between red and yellow parents to study the DFR transcription [74]. The DFR gene was sequenced using genomic DNA from red and yellow onions to develop molecular markers. The co-segregating molecular markers in the F2 population explain that the inactivation of the DFR gene led to the blockage of anthocyanin production, resulting in yellow onion. It was further reported that critical mutations in the DFR gene (DFR-B and DFR-C) are associated with a lack of anthocyanin, imparting yellow pigmentation to onions [69]. Both genes, DFR-B and DFR-C, have 95% sequence identity. However, the promoter region for both genes was completely different except for the 100 bp sequence adjacent to the untranslated region (5′UTR). A single base pair difference was reported in the promoter region of the red and yellow DFR allele, and an approximately 800 bp deletion was reported within the 3′ region of the yellow DFR-A allele. This deletion mutation was further exploited to develop codominant markers, which segregated perfectly in the F2 population, revealing the role of a deletion mutation in deactivating the yellow DFR-A gene, resulting in bulb colors other than red. The role of at least three loci in regulating red color was confirmed [63]. Three families, B2246 × B11159, BYG15-23 × AC43, and B8667B, were developed to study the color variation. Co-segregating families, SSRs, and SNPs in genomic amplicons of DFR and ANS revealed mutations in yellow onion, leading toward the absence of anthocyanins. Besides the DFR-A allele, two novel alleles (DFR-APS and DFR-ADEL) were isolated from yellow onions in Korea and Japan [78]. The DFR-APS allele isolated from the majority of the cultivars, containing a premature stop codon and a 20 bp deletion of a simple sequence repeat in the promoter region, was detected. However, in some cultivars, the complete deletion of the DFR-A gene and the promoter region led to identifying another novel allele designated as DFR-ADEL. Co-segregation of DFR-APS and color phenotypes in the F2 population reported the absence of anthocyanins due to the deactivated DFR-A gene. Also, reverse transcription polymerase chain reaction (RT-PCR) analysis confirmed a significant reduction in the transcription of the DFR-APS allele. However, no PCR products were amplified for DFR-ADEL throughout DFR-A sequences in the F2 population, reiterating the deletion of the entire DFR-A gene. Functional markers were developed based on the premature codon for identifying DFR-APS and DFR-ADEL [70]. These markers were further employed to screen the onion germplasm in Japan and Korea.
New information about dormant alleles connected to the ANS and DFR-A genes has been revealed [79]. Only one previously discovered inactive ANS allele was compared to the eleven dormant DFR-A alleles. A 4 bp insertion that caused a frame-shift mutation and a premature stop codon at the beginning of exon 2 was found to be the mutant ANS allele, ANSPS, in the yellow bulbs of the F2 population. The results of the RT-PCR showed a few ANSPS transcripts, which may imply nonsense-mediated mRNA degradation. In light-red F1 populations exhibiting gene complementation, an additional dormant ANS allele, ANSS188L, was discovered, which involves a crucial transition from serine to leucine. In this study, inactive (ANSPS, ANSS188L, and ANSG229R), leaky (ANSP), and active (ANSL and ANSh1) ANS alleles were identified. In another study, an F1 hybrid was developed to transfer resistance against downy mildew from the yellow-resistant cultivar ‘Santero’ to the yellow breeding line OT803 [80]. The pale pink color of the resulting F1 hybrids indicated possible connections between the ANS and DFR-A genes in the anthocyanin production pathway. OT803 was thought to have both active and inactive DFR-A alleles, even though ‘Santero’ had both active and inactive DFR-A alleles. On the other hand, it was discovered that a homozygous genotype for an active DFR-AR4-like allele was present in some yellow-colored OT803 individuals. Upon deciphering the 4830 bp complete sequence of this allele, a premature stop codon was generated due to a single nucleotide deletion compared to the DFR-AR4 allele. The DFR-A mutant allele, or DFR-APS2, has considerably lower transcript levels, which the nonsense-mediated mRNA decay process might cause. Further, Zhang et al. [81] investigated the role of variations in the cyanidin (Cy) metabolism for bulb color. Six libraries were examined: three dark-red and three white onion bulbs. Variations in the FLS and DFR genes may have an impact on pigment production. However, key genes such as F3′5′H and DFR were shown to be essential for the creation of dark-red bulbs. White bulbs lack Cy, which could account for their intricate coloring synthesis and multi-branching. The study drew attention to how the F3′5′H/F3′H ratio might affect the color diversity of onion bulbs.
Havey [82] generated various groups for the genetic mapping of the G locus by crossing plants with chartreuse bulbs with those with yellow bulbs. The yellow and chartreuse bulb distribution followed the predicted 3:1 ratio (p > 0.05) among the 17 F2 families examined. To create a genetic map of the G locus, DNA from one F2 family was analyzed for the SNPs, and 380 SNPs were found, including 119 that had not been previously mapped. The G locus was located at the terminal of chromosome 7, roughly 6.7 cM away from the closest SNP (isotig28625_2789). With codominant traits and a strong relationship to the G locus, this SNP marker is useful for introducing the recessive chartreuse bulb trait into other onion populations to cultivate this unique colored onion variety commercially. Recently, a candidate gene for the G locus has been identified [75]. A segregating population (JNU21C) was developed by crossing a male-sterile yellow line (JNUA12) with a fertile yellow line (JNUA25). Two more populations, one segregating population involving the self-pollination of JNUA25 to develop a linkage map flanking the G locus and another F2 population by crossing chartreuse SG21 and red UG19 for marker tagging the locus, were generated. This study identified 163 and 143 contigs through DEG and SNP screening, respectively. One of these contigs, designated AcLAC12, was discovered to be shared by the two screens and contained a laccase-like protein with conserved copper-binding domains. AcLAC12 mutations were linked to the color of chartreuse onions, and the chartreuse hue was created by inactivating AcLAC12 and the DFR-A gene. Mapped at the end of chromosome 7, AcLAC12 has a role in influencing the levels of quercetin and anthocyanin production in chartreuse onions. The interaction between the G and R loci was investigated by generating F2 populations from a cross between chartreuse and red parental lines [83]. The genetic analysis of these populations revealed that the chartreuse color of onion bulbs only developed when the R locus was homozygous recessive. On the other hand, the G locus did not influence the appearance of the red color. This suggests an important role of DFR-A inactivation in producing chartreuse bulb color. Additionally, mapping showed a linkage between the G and the R loci on chromosome 7 with a 19.5 cM distance.
Regulatory genes play an important role in the metabolic pathway of anthocyanins in addition to the structural genes. The regulatory role of the R2R3-MYB transcriptional factor in onion anthocyanin biosynthesis has also been revealed [84]. Subgroups (SGs) are frequently linked to either promoting the synthesis of anthocyanins (SG6, AcMYB1) or flavonols (SG7, MYB29), or preventing the synthesis of phenylpropanoid/flavonoid compounds (SG4, MYB4, MYB5). By promoting anthocyanin production in onion tissue following transient overexpression and decreasing pigmentation when briefly inhibited through RNA interference, MYB1’s positive regulatory role in anthocyanin biosynthesis was validated. However, the relationship between MYB1 and the C locus was not established. Six molecular markers were developed, and a linkage map was also constructed using F2 populations using yellow and white onion populations [85]. Two MYB-, one bHLH-, and one WD40-coding regulatory gene exhibited upregulation in the yellow onions. Moreover, Glutathione S-transferase (GST1) was found to be tightly linked to the C locus, but it was not the candidate gene for the C locus. The AcB2 coding for a bHLH transcription factor has been revealed to be the candidate gene encoding for the C locus [76]. Although AcB2 was less active in white bulbs, genetic markers did not always correlate with onion color. They discovered a non-autonomous DNA transposon, AcWHITE, in the promoter region of AcB2. The presence of AcWhite on AcB2 correlated with white onion bulbs. They concluded that AcB2 was the C locus and transcription factor for CHS. Recently, in a study comparing red and white onion bulbs, it was found that the expression of genes AcMYB1 and AcB2 was significantly reduced in white bulbs [86]. They discovered that AcB2 interacts with AcMYB1, and they play a key role in activating other genes (ANS and F3H1) involved in anthocyanin synthesis. Experiments with Arabidopsis showed that, when AcB2 and AcMYB1 were expressed together, the levels of ANS and F3H1 increased compared to when only AcMYB1 was expressed. Furthermore, the increase in the levels of both AcB2 and AcMYB1 in onion epithelial cells led to an induced accumulation of anthocyanins. This knowledge on regulatory genes could potentially help in breeding onions with higher anthocyanin contents.
Bulked segregant analysis and RNA-Seq were combined to locate the I locus on chromosome 3 by identifying 68 contigs with distinct genetic variations between dominant white- and red-bulked RNAs [77]. After examining 1457 F4 individuals, twelve markers were created and utilized to build an extensive linkage map surrounding the I locus. DW51596 and DW35019 markers were closely connected to this locus. Significant decreases in the genes encoding the flavonoid production enzymes were found in the dominant white bulk, and the same patterns were observed in the C locus-controlled recessive white bulbs. Twelve putative regulatory genes have been investigated; however, the precise gene controlling the I locus is still unknown.
4. Bulb Quality
Onions have been valued not only for their culinary purpose but also for their medicinal value. An onion bulb contains water (~90%), carbohydrates (7–9%), and dietary fiber (1–2%). Moreover, onions are a good source of certain vitamins and minerals, such as vitamin C, B6, folate, and potassium. Studies have shown that flavonoids, fructans, fructooligosaccharides (FOS), and organosulfur compounds, including allicin and thiosulfate in onions [87], are associated with antioxidant, anticarcinogenic, antimicrobial, antiplatelet, antithrombotic, and cardioprotective properties benefitting human health. Bulb composition is crucial in determining its suitability for various purposes, focusing on its carbohydrate content (glucose, fructose, sucrose, and fructans) and sulfur derivatives [88]. These compositional attributes are key factors in defining the quality of bulbs for specific applications. Varieties commonly called ‘storage onions’ are characterized by their high dry matter percent (DM%), typically ranging from 15% to 20%. Additionally, they exhibit relatively abundant fructans but lower levels of reducing sugars alongside relatively high concentrations of organosulfur compounds. This unique composition imparts storage onions with an intense and pungent flavor profile, making them highly desirable for culinary purposes, including cooking and various industrial processing applications. The flavor of onion is influenced by the soluble solids content (SSC) and its impact on bulb texture and storage life [89]. Comprehending the QTLs, identifying the associated genes responsible for variation in these nutraceutical compounds, and elucidating their molecular pathways can empower plant breeders to develop cultivars enriched with vital nutraceutical attributes (Table 1).
4.1. Soluble Carbohydrates
The major carbohydrates in an onion bulb are glucose (2–6%), sucrose (1–3%), fructose (1–3%), and fructans (10–20%) [90]. Fructans are considered beneficial in lowering the risk of colorectal cancer [91,92], diarrhea inhibition [93], relieving constipation [94], and reducing the risk of osteoporosis [95]. Researchers have put their efforts into identifying QTLs affecting the types and concentrations of carbohydrates in onion bulbs. It has been reported previously that greater levels of fructans are believed to be correlated with higher SSC [96], more pungency [97], longer bulb dormancy [98], and higher onion-induced antiplatelet activity (OIAA) [99]. Antiplatelet activity refers to reducing the aggregation of blood platelets to benefit cardiovascular health. Platelet aggregation can block arterial blood flow, causing coronary heart disease [100].
Fructans are polyfructose molecules derived from sucrose via enzymes fructosyltransferases [101]. Different types of fructans are biosynthesized and degraded during the life cycle of a plant. Fructan concentrations increase during bulb growth and decrease by the end of its growing season [102], and a further decline has been noticed during storage [103]. The important component for the fructan metabolism is speculated to be sucrose and enzyme sucrose: sucrose 1-fucosyltransferase (1-SST), which initiates the process by transferring a glucosyl residue from sucrose to generate 1-ketose. Further, another enzyme, fructan–fructan 1-fructosyltransferase (1-FFT), breaks the 1-kestose down into higher-DP (degree of polymerization) inulin-type fructans [104]. An enzyme, fructan–fructan 6G-fructosyltransferase (6G-FFT), catalyzes the synthesis of branched inulin neo-series fructans such as 6G-kestoriose, also known as neokestose [105,106,107]. Enzyme fructan exohydrolase (FEH) removes fructose units from fructan chains by breaking linkages at different places to make fructose and glucose molecules in fructan degradation [108,109,110]. Fructan metabolism might be regulated through sucrose-mediated signaling and could be closely related to the crosstalk between sugar signaling and the plant hormone network [111], which needs further exploration.

{kind=link}
Table 1.
Genetic mapping of important onion bulb quality traits.
| Bulb Quality Trait | Linked Markers | Mapping Population | Chromosome | Reference |
|---|---|---|---|---|
| Carbohydrates | ||||
| (6G,1)-nystose | API18–AOB77 | ‘Brigham Yellow Globe 15-23’ (BYG15-23) and ‘Ailsa Craig 43’ (AC43) | 4 | [112] |
| Sucrose | AJK84–API89 | 3 | ||
| Dry matter (DM%) | ACM171 | W202A’ × ‘Texas Grano 438’ | 5 | [89] |
| Fructans, fructose, and sucrose | ACM235 and ACM033 | ‘Colossal Grano PVP’ × ‘Early Long-keeper P12’ | 8 | |
| ACM033 and ACABE58 | ‘BYG15-23’ × ’AC43’ | 8 | ||
| DM% | API92 | ‘BYG15-23’ × ’AC43’ | 5 | [113] |
| DM% and fructans | ACM033 | 8 | ||
| Pungency | ||||
| Sulfur assimilation | ACM169 | ‘W202A’ × ‘Texas Grano 438’ | 3 | [114] |
| ACM 171 | 5 | |||
| Lachrymatory factor synthase (LFS) | ACP052 | A. fistulosum–shallot (A. cepa L. Aggregatum group) monosomic addition lines | 5A | [115] |
A study by Galmarini et al. [116] focused on the identification of QTLs for SSC, DM%, pungency, and OIAA using a biparental mapping population derived from a cross between ‘Brigham Yellow Globe 15-23’ (BYG15-23) and ‘Ailsa Craig 43’ (AC43). On linkage group E (chromosome 5), a region was significantly associated with all the traits contributing towards phenotypic variation present in them. The storage duration of onion bulbs significantly affects the detection of QTLs related to various bulb traits, highlighting the importance of considering the storage duration when breeding onion varieties to enhance desirable traits. Using the same cross, Havey et al. [112] utilized interval mapping of family means using DM% as the covariant exhibiting a region on linkage group A (chromosome 4) that was significantly affecting (6G,1)-nystose concentrations [LOD = 2.81] and another region on linkage group D (chromosome 3) for sucrose concentrations (LOD = 3.45). The availability of sucrose in high-solid onions affects fructan synthesis, affecting the accumulation of soluble carbohydrates and their relationship to thiosulfinates, pungency, and OIAA. Genetic variation present in the relative amounts of carbohydrates between storage (high dry matter) and sweet (low dry matter) onions was studied by McCallum et al. [89]. A segregation ratio of 3:1 for high-fructan and low-fructan individuals suggested the involvement of dominant gene action. A genomic region in one of the studied populations revealed a major QTL ‘Frc’ (LOD > 6) on chromosome 8, affecting total fructan content, with 93% of the phenotypic variation explained. In ‘W202A’ × ‘Texas Grano 438’ F2:3 families, two markers, ACM171 and ACM235, were significantly associated with DM%. The recombination estimates with ‘Frc’ for ACM235 and ACM033 markers were 10.5 ± 5 cM and 16.5 ± 5 cM, respectively, as both markers exhibited close linkage in the crosses, ‘Colossal’ × ‘ELK P12’ and A. cepa × A. roylei. Moreover, the QTL location was confirmed using the ‘BYG15-23’ × ’AC43’ mapping population, in which this QTL on chromosome 8 is flanked by with the SSR marker ACM033 and the SNP marker ACABE58, explaining 50.3% of the phenotypic variation. This locus may be responsible for conditioning large differences in carbohydrate concentrations between storage and sweet onions.
The genetic effects and interactions on DM% and soluble carbohydrates were estimated by Raines et al. [113] using segregating families involving a cross between two well-known inbreds, ‘BYG15-23’ (high soluble solids) and ‘AC43’ (low soluble solids). The patterns of segregation on chromosomes 5 and 8 show codominance with markers API66, API92, AOB236, ACM033, and ACABE58, showing a 1:2:1 ratio, and markers API47 and AJB72, showing a 3:1 ratio with a dominant effect. Two markers, API92 and ACM033, were highly significant for all the traits under study. The allele from the ‘high soluble solids’ parent at API92 on chromosome 5 elevated the dry weight and fructan amounts and reduced the levels of reducing sugars. However, the allele from the ‘BYG15-23’ parent at ACM033 elevated the fructan levels and decreased the sugar concentrations compared to the allele from ‘AC43’. These markers, API92 and ACM033, exhibited significant interaction for DM% and neokestose, which suggests the individual contribution of the two chromosomes for higher sucrose and fructan levels. The region on chromosome 5 from API66 to API47 was significantly associated with increased DM%, consistent with the previous study by Havey et al. [112]. Also, chromosome 8 influenced the DM% and fructan content, leading the region from ‘BYG15-23’ to explain 21–28% and 24–69% of the phenotypic variation, respectively.
These genetic studies have revealed some major QTLs for soluble carbohydrates and fructans. Three major loci on chromosomes 3, 5, and 8 influence fructans, glucose, sucrose, and fructose levels. Additionally, two markers (ACM033 and ACABE58) were discovered to be linked to a major locus, ‘Frc’, on chromosome 8, contributing to differences in fructan levels. However, the discovery of tightly linked markers is essential for MAS.
4.2. Pungency
Pungency is a function of bioactive organosulfur compounds [117] associated with the strength of onion flavor, which is either measured by the production of pyruvic acid [118] or the lachrymatory factor (LF) [119]. The major precursors for the onion flavor are 1-propenyl-, propyl-, and methyl cysteine sulfoxides. The flavor comes from the hydrolysis products of these alk(en)l cysteine sulfoxide (ACSO) precursors due to the activity of the enzyme CS-lyase alliinase [120]. The enzyme lachrymatory factor synthase (LFS) acts upon 1-propenyl sulfenic acid to produce lachrymatory factor (thiopropanal-S-oxide) [121]. Hydrolysis of one mole of ACSO by alliinase produces one molecule of enzymatically produced pyruvic acid (EPA), frequently used as an indicator of pungency [122]. Usually, the onion flavor is a mixture of sweetness, overall pungency, and the percentages of different ACSO precursors [120]. Some researchers have performed genetic mapping to identify genomic regions affecting pungency.
To identify genomic regions influencing pungency, McCallum et al. [114] evaluated a ‘W202A’ × ‘Texas Grano 438’ cross using a PCR-based molecular marker map comprising candidate genes for sulfur (S) assimilation. Linkage mapping exhibited that markers for two candidate genes, plastidic ferredoxin-sulfite reductase (SiR) and plastidic ATP sulfurylase (ATPS), are in close vicinity (1–2 cM) on chromosome 3. This close linkage was confirmed using a larger F2 population from another cross of ‘W429A’ × ‘Houston Grano’, which revealed a linkage of 2.2 cM (LOD 33) and showed a wide variation for pungency. A highly significant association between EPA and LF with markers on chromosome 3 was discovered, and significant associations were revealed both with the marker ‘ACM169’ and the region ATPS/SiR at the end of the linkage group. Moreover, the region of ‘ACM169’ was related to SSC, proposing its pleiotropic effect on pungency. The complete dominance of the ‘W202A’ allele over the ‘Texas Grano 438‘ allele confirmed the dominance at the ATPS/SiR locus, which controlled almost 50% of the heritable variation in pungency. Another marker, ‘ACM 171’, also exhibited significant relationships with pungency, and this region also has shown its influence on the bulb DM% [89]. This suggests that the markers ‘ACM169’ on chromosome 3 and ‘ACM171’ on chromosome 5 may help the breeders select less pungent onions without correlated selection for less soluble solids. The observation proposes that phenotypic selection for reduced pungency can lead to correlated selection for reduced solids, possibly negatively impacting storage. Contrarily, selection for storage life might select higher solids with a correlated increase in pungency.
The role of SiR in the regulation of sulfur assimilation in onion was further investigated by McManus et al. [123]. The activity of SiR was studied in Arabidopsis thaliana via a knock-down experiment, which revealed that it reduces the S-assimilation due to the downregulation of the enzyme [124]. In A. thaliana, SiR activity has been controlled by a single gene. Therefore, it is necessary to discover if a single gene copy contributes to S-assimilation in onions. In the study, SiR enzyme activity assays were performed to determine its activity concerning the sulfur supply. One functional gene, SiR, and an unprocessed pseudogene (script phi-SiR) were isolated, and SiR cDNAs were cloned. SiR activity is controlled by a single functional gene present on chromosome 3. They discovered a single copy of a full-length SiR gene and a pseudogene, suggesting that this step is the reductive S-assimilation pathway. Both genes are differentially expressed concerning one another, and the expression of the SiR gene and the enzyme is influenced by the S-supply, with higher expression from plants in S-sufficient conditions. The levels of SiR transcripts were significantly higher in the roots of the S-sufficient than in the S-deficient plants of the pungent cultivar, along with increased enzyme activity in the S-sufficient treatment. The transcription of SiR reductase and the enzyme’s activity can be a regulatory point of S-assimilation.
To understand the evolution of LFS, the genomic organization of LFS genes was studied by Masamura et al. [115]. The LFS gene was mapped to chromosome 5A using alien monosomic addition lines [A. fistulosum-shallot (A. cepa L. Aggregatum group)]. An F2 population from an interspecific cross of A. cepa × A. roylei confirmed it by identifying the co-segregation with chromosome 5 markers via the heteroduplex analysis of LFS amplicons. Several markers developed to onion ESTs, especially the SNP marker ‘ACP052’, were linked to the LFS marker. Further, two artificial bacterial chromosome (BAC) clones with the LFS gene were screened, and eight positive clones were discovered. Southern blot analyses of these eight clones using HindIII and EcoRI digestions suggested the presence of at least two LFS loci. Mapping of clones on onion mitotic metaphase chromosomes via fluorescence in situ hybridization (FISH) karyotype analysis presented signals on the proximal region on the long arm of chromosome 5. Further, the position of clones 2E8/10 and 4F10/155 were found to be 0.31 ± 0.03 and 0.32 ± 0.03, respectively. Additionally, two haplotypes from the double haploid analysis were distinguished: 4F10/155 as ‘haplotype 1’ and 2E8/10 as ‘haplotype 2’, exhibiting a larger variation in the upper flanking region (1.2 kbp) of the LFS between the BACs. This confirmed the involvement of at least two putative loci positioned on chromosome 5 near the dry matter QTL.
5. Bulbing and Flowering
Onion is a biennial crop that produces bulbs in the first and flowers in the second year. The bulb formed in the first year is a modified, underground stem with a flattened basal plate at the base, which acts as a shoot apical meristem (SAM). Each leaf comprises a photosynthetic leaf blade and a non-photosynthetic leaf sheath [125]. During the non-bulbing stage, leaves arise from the SAM, but once bulbing starts, the emergence of new leaves ceases, which leads to the swelling of the leaf sheath to form a “bulb,” which acts as an overwintering stage. Onion plants flower only in the spring, when environmental conditions are favorable to ensure maximum reproductive success [126,127]. Multiple external (environmental cues) and internal factors (plant age and hormones) control bulbing and flowering [126,127,128,129,130]. Among the environmental factors, day length plays a key role in bulbing, as plants only form bulbs once their critical photoperiod requirement is satisfied, and these bulbs flower when their vernalization requirement is fulfilled [125,126,131,132]. Onion is a long-day plant but exhibits tremendous variation for day-length sensitivity. As a result, onions are grown worldwide. Onion cultivars grown at low latitudes require a 10–12 h photoperiod to form bulbs, whereas cultivars grown at mid- and high latitudes need a 12–14 and >16 h photoperiod for bulbing, respectively. Furthermore, germplasm at different latitudes exhibits variation in vernalization sensitivity as cultivars adapted at high latitudes have stronger vernalization than cultivars at low latitudes [125,133,134,135].
The molecular mechanism of the photoperiod and vernalization is well understood in Arabidopsis thaliana [136]. Once plants are vernalized during winter and the days become long during spring, FLOWERING LOCUS T (FT) is produced in the leaves and transported to the shoot apical meristem, where it forms a florigen activation complex (FAC) by interacting with 14-3-3 and FLOWERING LOCUS D (FD). The FAC triggers transcriptional activation of different meristem identity genes, key in transitioning from the vegetative meristem to the reproductive stage [137,138,139,140]. Before winters, FLOWERING LOCUS C (FLC) causes transcriptional suppression of FT to prevent flowering. However, the low temperature of winter lowers the expression of FLC after a few days of cold exposure. Still, it requires extended vernalization for the FLC silencing to be sustained through epigenetic modifications [141]. When the vernalization requirement is fulfilled and the days become long during spring, the circadian clock genes, GIGANTEA (GI) and FLAVIN BINDING KELCH REPEAT F-BOX PROTEIN 1 (FKF1), form a complex to stabilize CONSTANS (CO) to activate FT [142,143,144,145,146,147,148].
In recent years, several remarkable studies have demonstrated that, apart from flowering, FT controls diverse developmental stages in plants, such as bud growth and storage organ formation [149,150,151]. One example is onion, where different FTs (AcFT1, AcFT2, and AcFT4) respond to the photoperiod and vernalization to control bulbing and flowering, respectively [151]. AcFT4 prevents bulbing in young plants or mature plants grown under the non-inductive photoperiod (SD); however, as the plants mature and experience an inductive photoperiod (LD), its expression reduces, resulting in the upregulation of another gene, AcFT1, which acts as a bulbing promoter. In addition to these FTs, several circadian clock genes, such as GI, FKF-1, and CO, have been identified in onion (AcGI, AcFKF-1, and three AcCOLs), but their role in bulbing is not known yet [152,153]. After bulbing, mature bulbs are harvested and replanted in the following year for seed production. During this time, cold exposure causes the transcriptional activation of AcFT2, which promotes flowering [151]. Apart from AcFT2, a genomic region on chromosome one has been identified as responsible for premature bolting, but the candidate gene(s) conditioning this region is unknown [154].
6. Biotic Stress
Onion production faces major limitations due to biotic stresses, such as fungi and bacteria, that cause significant damage to onions [155]. Major fungal diseases severely impacting onions are Fusarium basal rot, pink root, purple blotch, downy mildew, and gray mold. Bacterial diseases that are the major threats to onions include bulb soft rots and Xanthomonas leaf blight. Despite using conventional breeding procedures to develop resistant cultivars, MAS has successfully accelerated crop improvement for most diseases. The information below summarizes the recent molecular studies conducted to comprehend the genetics of resistance for developing resistant onion cultivars for major diseases (Table 2).
6.1. Fusarium Basal Rot
Fusarium basal rot (FBR) is a major disastrous soil-borne fungal disease caused by Fusarium oxysporum Schlechtend.: Fr. f. sp. cepae (H.N. Hans.) W.C. Snyder & H.N. Hans (FOC) [165]. Disease symptoms can be observed at various stages during the life cycle of an onion plant, which includes delayed emergence and the damping-off of seedlings, chlorosis, necrosis of the leaves, brown discoloration of the basal plate tissues of mature bulbs, root death, root abscission, and plant death [166].
In a hybrid of FBR-resistant and FBR-susceptible long-day inbreds, the research identified two partially dominant genes, Foc1 and Foc2, with additive effects on FBR resistance [167]. In a hybrid including five long-day inbreds, Tsutsui [168] discovered a single dominant gene in FBR resistance with cytoplasmic gene possibilities, but Krueger’s studies [169,170] suggested nuclear or cytoplasmic gene participation through reciprocal differences. Multiple gene inheritance was indicated by several investigations [171,172,173]. The genetics behind the inheritance of FBR resistance is still elusive. Recent studies have been conducted to find QTLs conditioning FBR resistance [156,157]. Taylor et al. [157] examined genetic variation across founder onion accessions and between and within half-sib families. Genotyping was carried out using 892 published KASP SNP markers by Duangjit et al. [30]. Of the 892 SNPs, 765 polymorphic markers were selected for further genetic analysis. They found the possible involvement of chromosomes 1, 6B, and 8 in FBR resistance. Linked markers mapped on chromosomes 1, 6B, and 8 were c00676_1004 (149.8 cM), i34519_442 (30.2 cM), and I30594_1021 (1.1 cM), respectively. A patent by Black et al. [158] revealed a region in the linkage group 2 for carrying a codominant FBR resistance with the allele from the resistant parent, which corresponded to chromosome 4. F3 bulbs were obtained from the original mapping population, SYG-75-1706/Serrana-FBR. An onion genomic region defined by loci NQ0345038 and NQ0257326 was associated with FBR resistance. Straley et al. [156] established segregating families to map FBR resistance and estimate the genetic effects. To produce seeds from the hybrid plants and create a segregating F2 family, the FBR-resistant ‘W440’ as the male was crossed with the inbred ‘B5351’ as the female, which was FBR-susceptible. Thirty-three F2 offspring of B5351 × W440 were segregated for 116 SNPs that fit the predicted 1:2:1 segregation ratio (p > 0.001). Three QTLs were identified, one on chromosome 2 and two on chromosome 4. The codominant FBR resistance was most significantly correlated with the marker isotig38484_281 on chromosome 2B. The chromosome 4A marker isotig44683_192 was substantially linked to FBR resistance. On chromosome 4C, the W440 allele at the marker isotig31106_505 was associated with FBR susceptibility. The diversity among FOC isolates may impede the identification of FBR-resistant germplasm and subsequent genetic research. However, the SNPs discovered so far may be useful for future research to comprehend the underlying mechanism of FBR resistance.
Saponins are a type of secondary metabolite synthesized by plants that change levels upon exposure to external stresses [174,175]. Saponins exhibit antifungal properties by forming pores and disrupting the membrane integrity of fungal cells through complex formation with sterols [176,177]. This leads to the leakage of cell contents or programmed cell death in sensitive fungal cells [178]. Allium plants generate antifungal steroidal saponins, and several studies have been conducted to identify and test these saponins against various fungal species. A study investigated the antifungal effects of shallot (a close relative of bulb onion) metabolites on FOC and identified the shallot chromosome responsible for resistance to the disease. The researchers studied a shallot cultivar, A. fistulosum, and a complete set of A. fistulosum–shallot monosomic addition lines (MALs) [179]. After inoculation with the FOC, shallot roots showed a significant increase in the saponin content of root exudates, and a higher concentration of crude saponins induced complete cell death. They identified the line FF + 2A, which contained chromosome 2A of shallot and a specific saponin band derived from shallot, as the most resistant to FBR. This suggests that chromosome 2A of shallot may contain genes related to FBR resistance. Teshima et al. [180] also investigated the antifungal properties of steroidal saponins found in the bulbs. They discovered that crude saponin levels were highest in the roots and basal plate tissue and that total saponins from the basal plate tissue exhibited antifungal activity against several fungal isolates. They isolated two saponins, alliospiroside A (ALA) and alliospiroside B (ALB), which exhibited antifungal activity against Fusarium species, with ALA being more effective than ALB. Further, a study by A. Mostafa et al. [175] aimed to identify the saponin compounds in shallots, including their biosynthetic pathway. The researchers analyzed the phytochemicals in shallot, A. fistulosum (FF), and MALs to identify the candidate genes involved in producing enzymes in the steroidal saponin biosynthesis pathway. They identified a previously identified saponin compound, alliospiroside A, in FF + 2A with potential antifungal activity against FOC strains. By understanding these genes, it may be possible to develop plants with unique saponin content and enhanced resistance to Fusarium diseases.
6.2. Pink Root
Pink root (PR) is another soil-borne disease that affects onion plants and is caused by the fungus Pyrenochaeta terrestris. The disease is characterized by developing pinkish to dark-purple lesions on the plant roots, which disintegrate later [181].
Genetic resistance to PR was conditioned by a single recessive locus [182,183]. A recent study by Marzu et al. [159] showed that the same chromosomal region conditions confer codominance for PR resistance originating from various sources. Two segregating F2 families (PR1 and PR2) were generated by crossing the male-sterile inbred line ‘DehyA’ and female inbreds ‘B5351C’ and ‘Ski-MsMs’ to develop F2M families. Later, a susceptible female inbred ‘B5351C’ was crossed to the inbred ‘W446B’. F1MS families (PR3 and PR4) were also developed by selfing randomly chosen plants from F1M families. Using 92 F2M progenies from the PR2 family and 93 F2M progenies from the PR1 family, a total of 111 SNPs were mapped. The PR resistance QTL was consistent and detected on chromosome 4 with LODs of 8.3 and 10.1 and phenotypic variations of 27.6% and 35.1%, respectively, on the PR1 and PR2 families. PR resistance from the ‘DehyA’ parent lowered disease severity and was codominantly inherited in both families, where additive effects were greater than dominance effects. For the PR3 and PR4 families, 54 and 39 F1MS families were screened, respectively, and 151 SNPs were employed to map PR survival. The most significant QTL, localized to the same area on chromosome 4 as the PR1 and PR2 families, explained 54% of the phenotypic variation for PR survival. The SNP marker isotig26045_1046, the closest to the PR resistance QTL, showed a positive additive effect, exhibiting an increase in PR survival by 24% for each additional W446B allele. In a patent by Black et al. [158], PR resistance was mapped in the short-day population, SYG-75-1706, and the resistance was discovered to have a strong additive effect that boosted seedling survival by 32%. Two loci, NQ0257277 and NQ0258453, surrounding an onion genomic region linked to PR resistance, were mapped on chromosome 4. According to these findings, three sources of PR resistance exhibit codominance in seedling survival assessments and map to the same area on chromosome 4.
6.3. Purple Blotch
Purple blotch (PB) is a devastating foliage disease of onion that is caused by the fungus Alternaria porri (Ellis) Ciferri [184]. This disease influences leaves and bulbs and reduces the bulb yield by up to 97% [185]. Symptoms of the disease include sunken purple lesions, which enlarge and produce brownish-purple necrotic lesions, causing delayed bulb development [186,187].
Two SSR markers sets, matK, and rbcL, were evaluated for marker-assisted selection for PB resistance s1/s2 locus on chromosome 8 [160]. They were dominant markers for resistance. The PCR products for resistant lines were 214 bp and 67 bp for matk and rbcL, respectively. In a recent study, three populations (F1, F2, and BC1) generated by crossing ‘Arka Kalyan’ (resistant) and ‘Agrifound Rose’ (susceptible) were evaluated to study the inheritance and develop molecular markers linked to PB resistance [161]. Using a virulent isolate of A. porri, it was discovered that the F1 was resistant, while 498 F2 plants segregated in a 3:1 resistant (R) to susceptible (S) phenotypic ratio and 128 BC1 lines segregated in a 1R:1S ratio, indicating that PB resistance is regulated by a single dominant gene known as Alternaria porri resistance 1 (ApR1). In a bulk segregant analysis, 59 of the 288 [ISSR (146) and SSR (142)] primer sets that discriminated between two parental lines were used to connect them to the alleged ApR1 gene. SSR marker, AcSSR7, and STS marker, ApR-450, were found to be strongly coupled to the ApR1 locus at distances of 1.3 and 1.1 cM, respectively, which can be successfully used for MAS. Six KASP markers and the ApR1 gene were strongly associated at a genetic distance of 1.3 cM according to a recent study including 153 recombinant inbred lines (RILs) [162]. Co-segregation of the ApR1 locus and ApRsnip14 was observed. MAS for resistance to purple blotch is advised for ApRsnip14 and ApRsnip23, which are closely linked to ApR1.
6.4. Downy Mildew
Downy mildew of onion caused by an oomycete fungus Peronospora destructor (Pd) is another foliage disease [188]. It is one of the major fungal diseases threatening onion production worldwide. Infected plants produce soft, shriveled, and sprouted onions prematurely in storage, causing significant yield losses [189]. A wild relative of Allium roylei Stearn was found to be completely resistant to downy mildew, and this resistance was transferred into a bulb onion through interspecific hybridization and backcross breeding [190,191]. One dominant gene called Pd was found to be responsible for the introgressed downy mildew resistance [190].
In a patent by Harrewijn et al. [192], an investigation was carried out to identify the locus providing resistance to downy mildew. They identified a homozygous plant of A. cepa with a Pd resistance locus in the long arm of chromosome 3, similar to the genomic segment of A. roylei responsible for resistance. Later, the locus data from the high-density linkage maps [30] and transcriptome sequences generated by RNA-Seq [193] were used in a study by Kim et al. [163] to build a straightforward and reliable PCR marker that was connected to the downy mildew resistance gene. Additionally, molecular markers for recombinant selection in marker-assisted backcrossing (MAB) were created after measuring the range of an introgressed A. roylei fragment in chromosome 3 of the downy mildew-resistant onion cultivar. So, from a transcriptome database, cDNA sequences of the loci found at the end of chromosome 3 on two linkage maps were acquired. Eight loci’s exon sequences were used to build primer pairs; among these, the i25255 locus showed length variation between A. roylei and bulb onion, and both large- and small-sized PCR products were observed in the resistant cultivar. A 67 bp indel in the intron sequences, a PCR marker known as DMR1, was discovered. The resistant cultivar ‘Santero’ was the only accession with the A. roylei-specific marker. These outcomes showed that the A. roylei fragment carrying the downy mildew resistance gene was successfully tagged by the DMR1 marker.
By employing a straightforward and reliable molecular marker in the interspecific breeding of bulb onions resistant to downy mildew, the benefits of genomic in situ hybridization (GISH) were utilized to track the introgression process at the chromosome level [194]. Using GISH and the previously identified DMR1 marker linked to the resistance gene Pd, homozygous introgression lines with downy mildew resistance were successfully produced in a relatively short breeding period. It took seven years from the production of the F1 hybrid to the development of the S2BC2 homozygous lines that are resistant to downy mildew, which is not a long time given that the bulb onion is a biennial crop. A homozygous fragment of A. roylei was found in three S2BC2 offspring plants’ long arms of chromosome 3 in A. cepa genetic background.
6.5. Gray Mold
Gray mold is caused by the necrotrophic fungi B. cinerea and B. squamosa at the stage of bulb development and by B. cinerea during storage [195,196]. In a study by Kim et al. [164], they developed the sequence-characterized amplified region (SCAR) marker by studying polymorphism between resistant and susceptible onion lines using four resistant and three susceptible lines. This study aimed to create molecular markers for breeding onions resistant to gray mold. From the sequencing of the amplified product, the SCAR-OPAN1 marker was designed by including the 5′-ACT CCA CGT C-3′ sequence of the OPAN-1 random primer, and this marker was used to screen resistant and susceptible germplasms. The marker amplified the specific band only from the resistant lines, demonstrating its potential as a molecular marker for selecting resistant lines. Among the 1636 transcripts, 320 transcripts showed higher expression levels in the resistant group, and 1316 showed higher expression in the susceptible group. Further, resistance-related transcripts were confirmed to be related to disease resistance to breed gray mold-resistant onions, which were later used to develop SNP markers to screen gray mold-resistant onions. One SNP showed the ‘T’ allele in the resistant lines and the ‘C’ allele in the susceptible lines, while the other SNP showed the ‘C’ allele in the resistant lines and the ‘G’ allele in the susceptible ones. It was confirmed that the SNP-3 high-resolution melting (HRM) marker is versatile and can be used to select resistant onions. Since the SNP-3 HRM marker was gene-based, a qPCR analysis was conducted to confirm the expression level of the aldo-keto reductase (AKR) gene. The expression level of AKR was higher in the resistant group than in the susceptible group. Therefore, the SCAR-OPAN1 and SNP-3 HRM markers were developed to select resistant onion lines in breeding programs to reduce the damage caused by gray mold disease.
Lee et al. [197] also explored the expression of genes involved in resistance to gray mold, where the screening resistance and the genetic relationships of 54 onion lines based on RAPD and in vitro-cultured onion lines infected with gray mold were studied. One onion line with a resistant phenotype (Asia-23) and one with a susceptible phenotype (Asia-24) were selected. Leaf samples were obtained after infection for time-series gene expression analyses. Onion leaves were sampled at 0 (control), 4, 8, 16, and 32 h post-inoculation (hpi). The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway ‘-linoleic acid metabolism’ was chosen because it had a strong correlation with gray mold-resistant phenotypes when the time series expression patterns of the genes for jasmonate-resistant 1 (JAR1), coronatine-insensitive protein 1 (COI 1), and transcription factor MYC2 were compared between resistant and susceptible lines. As confirmed by HRM analysis, the JAR1, COI1, and MYC2 gene sequences had SNPs between the Asia-23 and Asia-24 lines. Future research on the genes involved in onion disease resistance and breeding onions with traits resistant to gray mold are both anticipated to benefit from these findings.
7. Male Sterility and Hybrid Development
Cytoplasmic male sterility (CMS) refers to a maternally inherited inability to produce viable pollen grains and is widespread among higher plants. It is a valuable tool in facilitating the production of hybrid seeds in self-pollinating crops like rice, cotton, and maize, and several vegetable crops such as onion, pepper, carrot, cabbage, broccoli, and radish. CMS is characterized by the de novo expression of mitochondrial chimeric mutants resulting from novel ORFs generated by spontaneous rearrangements of mitochondrial genomic sequences. They may vary in sequence, structure, and function and involve sub-stoichiometric shifting [198]. Expression of such mitochondrial mutation confers pollen disruption, with the plant failing to produce functional pollen. However, full pollen fertility can be restored in the male-sterile cytoplasm by nuclear-encoded restorer-of-fertility (Rf) genes through post-transcriptional or post-translational modification involving altering or suppressing the expression of the CMS-associated ORFs [199].
Hybrid onions started with the discovery of the sterile (S) cytoplasm in the Italian Red cultivar [200]. In this cytoplasm, fertility was restored by the dominant allele at the Ms locus [201]. Another type of male sterility, CMS-T [202], was discovered in the cultivar ‘Jaune paille des Vertus’. This cytoplasm can be restored by the interaction of the T cytoplasm with three independent loci [203] comprising an independent gene (A) and two complementary genes (B and C). In the CMS-S system, male-sterile lines (Smsms) are maintained by crossing with the isogenic maintainer line (Nmsms), whereas hybrids can be developed by using either a maintainer (Nmsms) or restorer (NMsMs/NMsms) line. To identify male-sterile and maintainer lines through conventional breeding, progeny testing was used [204]. However, genotyping by progeny tests to extract the maintainer individuals from onion populations was difficult and time-consuming because of the biennial generation time, high frequency of the dominant allele at the Ms locus [205], and prevalence of the S cytoplasm [206]. Four to six years was required to isolate male-sterile and maintainer lines. Molecular markers capable of distinguishing cytoplasm types and Ms locus genotypes are an easy way to speed up the breeding process and save time and effort for the breeders. MAS is a powerful tool that can facilitate the rapid transfer of economically important genes into cultivars [207]. Further, MAS will be helpful in the accelerated development of male-sterile and maintainer lines from open-pollinated populations [208].
Research to distinguish cytotypes started with de Courcel et al. [209], differentiating the onion cytoplasm into two groups, (i) the M group containing the N and T cytoplasms and (ii) the S group containing the S cytoplasm based on RFLP profiling of mitochondrial and chloroplast DNA. They concluded that the N and T cytoplasms were autoplasmic in origin and may have been introgressed from Allium vavilovii [210]. The S cytoplasm was thought to have originated from an interspecific cross [211]. Havey [212] proposed ‘Pran’, a triploid viviparous onion, as the putative donor of the S cytoplasm, and the T cytoplasm was thought to have been extracted from Dutch or Japanese populations [213]. Satoh et al. [214] reported mtDNA markers for differentiating the N and S cytoplasms. PCR markers identifying the N and S cytoplasms led the way toward identifying maintainer lines from open-pollinated populations [207,215]. Of the two cytotypes, S is widely used for F1 hybrid development because of its simple restoration mechanism and stability under diverse environments [213]. A PCR-based marker system, based on the combination of cob marker with orfA501, distinguishing the S, N, and T cytoplasms, was reported by Engelke et al. [216]. Further, it was observed that the T cytoplasm is also present in Turkish and German varieties besides the French varieties. Kim et al. [217] designed a new PCR marker based on the relative copy numbers of orf725 and cox1 to distinguish three cytoplasms with one PCR marker. Sequencing and annotating the chloroplast led to developing PCR markers, targeting small indels and deletions in the accD gene for distinguishing N and S cytotypes [218]. With the advancement of sequencing technologies, two variants, each of N and CMS-T cytotypes (Cytotype X and Y), have been identified [219,220]. Deeper insights into the cytoplasms of various geographic regions have led to the postulation of new variants of the T cytoplasm (T, T-like, Y) [220]. The main hypothesis that three independent loci can restore male sterility caused by CMS-T was challenged by Kim [221]. It was hypothesized that fertility of ‘T’-like cytoplasm [202] could be restored by the dominant Ms allele, which contrasted with the studies of Havey [213], who reported that the dominant alleles at Ms locus could not restore T cytoplasmic male-sterile lines. De Vries and Wietsma [222] also reported male fertile plants by crossing male-sterile lines of ‘Rijnsburger’ onion with Allium roylei. These studies signify that additional variation in CMS systems may be prevalent in onion. Havey and Kim [223] suggested that four cytotypes, viz., N, R, S, and T, exist. The S and R cytoplasms are commonly used, whereas the T cytoplasm is rarely used commercially.
Diversifying the male sterility-inducing cytoplasm in onions is an important task to avoid epidemics like southern corn leaf blight on maize caused by only one cytoplasmic source [224]. Hybridization of onion with Allium galanthum resulted in the galanthum cytoplasmic population, with the complete absence of anthers, that can be used as an alternative source of sterility [225]. The exploitation of Allium roylei to develop new male-sterile cytoplasmic lines was successful, and A. roylei can be used as a new source of male sterility [226].
Identification of the cytoplasm is the preliminary step towards isolating male-sterile lines. The next step is to identify the restorer-of-fertility (Ms) locus in the open-pollinated populations to isolate the maintainer (Nmsms) and restorer (NMsMs/NMsms) lines. Since the commercial part of the onion is a bulb, the isolation of the restorer line is unimportant. Molecular markers that can effectively distinguish the Ms locus are indispensable, and several molecular markers for the allelic selection of the nuclear Ms locus have been reported. AOB272 (0.9 cM) is the closest RFLP marker to the Ms locus in the onion genome [227] and AGF136 is positioned on the opposite side of the AOB272 marker [228]. The OPT and PsaO markers derived separately from AOB272 and AGF136 are linked in opposite directions to the Ms locus at distances of 1.5 and 6.4 cM, respectively [229]. ACms.1100 is a CAPS (cleaved amplified polymorphic sequence) marker of the Ms locus, and the genetic distance between the Ms and the ACms.1100 marker is presumed to be much closer than the distance between the Ms and the OPT marker [230]. Two CAPS markers, jnurf05 and jnurf17, are linked very close to the Ms locus, and jnurf05 was estimated to be 0.05 cM from the Ms locus [231]. These markers are useful for selecting specific alleles at the Ms locus in segregating families with linkage disequilibrium. The male-fertility restorer (Ms) locus has been mapped to chromosome 2 of the onion and linked genetic markers have been identified. Havey [232] identified three tightly linked SNPs on chromosome 2 and remained in linkage disequilibrium with genotypes at Ms locus. Based on AFLP and hi TAIL-PCR, Yang et al. [233] developed two SCAR markers to identify Ms locus. Out of 14 PCR markers, 2 PCR markers, jnurf12 and jnurf13, were developed, and jnurf13 was reported to be in linkage disequilibrium with Ms locus in diverse breeding lines [221]. Two new PCR-based markers, AcSKP1 [234] and AcPMS1 [219], were developed to identify and tag the Ms locus in onion. AcSKP1, a multiplex PCR-based codominant marker, is composed of four compatible primers (FU898, FD898, SU628, and SD628) to distinguish homozygous, heterozygous, and recessive alleles at the Ms locus. The utility of this primer has been established in 25 breeding lines, seven hybrid cultivars, one F2 population, two BC1 populations, and four open-pollinated (OP) populations. It was observed that the AcSKP1 marker perfectly predicted the genotypes at the Ms locus in the four OP populations according to the phenotypes of testcross progenies. A combination of BSA and RNA-seq was performed to identify candidates for the restorer-of-fertility gene in onion [219], and AcPMS1, a gene involved in DNA mismatch repair, was identified as the best candidate. Khar and Saini [235] pointed out the limitations of these PCR markers, especially those linked to the Ms locus, for identifying maintainer lines. The GISH technique revealed that the assignment of PCR markers linked to the Ms locus was placed at the proximal locations close to the centromere on the long arm of chromosome 2, a region of lower recombination [236]. This explained that the reported tightly linked markers may be distant from the Ms locus, leading to the limitations of these PCR markers. But Havey and Kohn [237] reported that two PCR markers, jnurf13 and AcPMS1, predicted the genotypes at the Ms locus accurately in the North American onion populations, and AcPMS1 was better among the two. In Indian onion populations, AcPMS1 was also better than AcSKP1 in determining the Ms locus [238]. A novel AcPMS1 allele in one onion breeding line (ORI854), which is rarely distributed in the onion germplasm, was reported by Kim and Kim [239]. Recently, a new locus for restoring male fertility (Ms2) was reported [240]. Comparative mapping revealed that Ms2 is positioned at the end of chromosome 2 with a distance of 70 cM from the Ms locus. The transcription level suggests that the male fertility mechanism of Ms2 is different from the Ms locus.
These PCR markers have been used to predict the frequency of the cytoplasm and Ms locus in onion populations from various countries. The frequency of the T cytoplasm (48%) was more than the S cytoplasm (15%) in Korean onion [217], the 2% male-sterile line (both the S and T cytoplasms) [241], the S (56%) and T (25.8%) cytoplasms in Brazilian onion [242], and the S cytoplasm (2.15%) in Indian onion VL Piaz-3 [243], whereas the S cytoplasm in the range of 88% (Sel. 121-2), 33% (Pusa Red), and 17% (Pusa Madhavi) was reported by Khar and Saini [235]. Until 2015, the primers reported by Sato [215] and Engelke et al. [216] were mostly used for cytoplasm determination [242,243,244]. Recently, new primers for the cytoplasm [217,218] and Ms locus [219,234] have been utilized for the identification of the cytoplasm and Ms locus in onions from Brazil [245], India [238,246], and South Africa [247]. Transcriptomic studies of male-sterile and maintainer lines predicted that cytoplasmic genes (mitochondrial energy metabolism-related genes cox1 and atp9) and nuclear genes (tapetal development-related genes AG, AMS, and SERK1) are likely involved in the CMS in onions [248].
Early research in male sterility might have emerged in the pursuit of exploiting the value of hybrid vigor in the agricultural enterprise, but further research at the molecular level has provided an understanding of the sterile nature as a reproductive strategy in plants. To conclude, the isolation of male-sterile and maintainer lines can be accelerated by using these PCR markers at a preliminary stage. These markers are important, mostly in countries where onion hybrids have not been commercialized at a larger scale. With the draft genome of onion published [31], in-depth research in male sterility systems of onion from various geographic regions may be useful to develop markers in linkage disequilibrium with the Ms locus.
8. Conclusions
In the genetics of onions, several traits have been found and located on genetic maps; nevertheless, to facilitate marker-assisted selection, it is necessary to find tightly linked markers with the gene of interest. For this purpose, large-scale, diverse mapping populations must be developed and utilized. This is an essential step that will greatly increase the accuracy of the onion linkage map and make marker identification easier. Association mapping methods, especially when applied to large germplasm collections, can potentially uncover previously unknown alleles affecting many phenotypes. Additionally, the continued progress in sequencing technology and collaborative efforts to develop draft genome sequences for bulb onions are expected to increase the amount of genetic information available for onions significantly.
Author Contributions
Conceptualization, S.S. and S.M. (Subas Malla); writing—original draft preparation, S.S., A.K., J.S.K., S.M. (Subhankar Mandal) and S.M. (Subas Malla); writing—review and editing, S.M. (Subas Malla). All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by United States Department of Agriculture—National Institute of Food and Agriculture 2023-51181-41172, Texas Department of Agriculture Specialty Crop Block Grant GSC2022045, and Texas A&M AgriLife Vegetable Seed Grant.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Moreno-Rojas, J.M.; Moreno-Ortega, A.; Ordóñez, J.L.; Moreno-Rojas, R.; Pérez-Aparicio, J.; Pereira-Caro, G. Development and Validation of UHPLC-HRMS Methodology for the Determination of Flavonoids, Amino Acids and Organosulfur Compounds in Black Onion, a Novel Derived Product from Fresh Shallot Onions (Allium cepa var. aggregatum). LWT 2018, 97, 376–383. [Google Scholar] [CrossRef]
- Zamri, N.; Hamid, H.A. Comparative Study of Onion (Allium cepa) and Leek (Allium ampeloprasum): Identification of Organosulphur Compounds by UPLC-QTOF/MS and Anticancer Effect on MCF-7 Cells. Plant Foods Hum. Nutr. 2019, 74, 525–530. [Google Scholar] [CrossRef]
- Ren, F.; Reilly, K.; Kerry, J.P.; Gaffney, M.; Hossain, M.; Rai, D.K. Higher Antioxidant Activity, Total Flavonols, and Specific Quercetin Glucosides in Two Different Onion (Allium cepa L.) Varieties Grown under Organic Production: Results from a 6-Year Field Study. J. Agric. Food Chem. 2017, 65, 5122–5132. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Parks, J.S.; Kang, H.W. Quercetin, a Functional Compound of Onion Peel, Remodels White Adipocytes to Brown-like Adipocytes. J. Nutr. Biochem. 2017, 42, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Zhu, D.Y.; Thakur, K.; Wang, C.H.; Wang, H.; Ren, Y.F.; Zhang, J.G.; Wei, Z.J. Antioxidant and Antibacterial Evaluation of Polysaccharides Sequentially Extracted from Onion (Allium cepa L.). Int. J. Biol. Macromol. 2018, 111, 92–101. [Google Scholar] [CrossRef]
- Lanzotti, V.; Romano, A.; Lanzuise, S.; Bonanomi, G.; Scala, F. Antifungal Saponins from Bulbs of White Onion, Allium cepa L. Phytochemistry 2012, 74, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Dahlawi, S.M.; Nazir, W.; Iqbal, R.; Asghar, W.; Khalid, N. Formulation and Characterization of Oil-in-Water Nanoemulsions Stabilized by Crude Saponins Isolated from Onion Skin Waste. RSC Adv. 2020, 10, 39700–39707. [Google Scholar] [CrossRef]
- Ouyang, H.; Hou, K.; Peng, W.; Liu, Z.; Deng, H. Antioxidant and Xanthine Oxidase Inhibitory Activities of Total Polyphenols from Onion. Saudi J. Biol. Sci. 2018, 25, 1509–1513. [Google Scholar] [CrossRef]
- Loredana, L.; Adiletta, G.; Nazzaro, F.; Florinda, F.; Marisa, D.M.; Donatella, A. Biochemical, Antioxidant Properties and Antimicrobial Activity of Different Onion Varieties in the Mediterranean Area. J. Food Meas. Charact. 2019, 13, 1232–1241. [Google Scholar] [CrossRef]
- Jakaria, M.; Azam, S.; Cho, D.Y.; Haque, M.E.; Kim, I.S.; Choi, D.K. The Methanol Extract of Allium cepa L. Protects Inflammatory Markers in LPS-Induced BV-2 Microglial Cells and Upregulates the Antiapoptotic Gene and Antioxidant Enzymes in N27-A Cells. Antioxidants 2019, 8, 348. [Google Scholar] [CrossRef]
- Lee, J.S.; Cha, Y.J.; Lee, K.H.; Yim, J.E. Onion Peel Extract Reduces the Percentage of Body Fat in Overweight and Obese Subjects: A 12-Week, Randomized, Double-Blind, Placebo-Controlled Study. Nutr. Res. Pract. 2016, 10, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Jini, D.; Sharmila, S. Green Synthesis of Silver Nanoparticles from Allium cepa and Its in Vitro Antidiabetic Activity. Mater. Today Proc. 2020, 22, 432–438. [Google Scholar] [CrossRef]
- Tsuboki, J.; Fujiwara, Y.; Horlad, H.; Shiraishi, D.; Nohara, T.; Tayama, S.; Motohara, T.; Saito, Y.; Ikeda, T.; Takaishi, K.; et al. Onionin A Inhibits Ovarian Cancer Progression by Suppressing Cancer Cell Proliferation and the Protumour Function of Macrophages. Sci. Rep. 2016, 6, 29588. [Google Scholar] [CrossRef] [PubMed]
- Colina-Coca, C.; Gonzalez-Pena, D.; de Ancos, B.; Sánchez-Moreno, C. Dietary Onion Ameliorates Antioxidant Defence, Inflammatory Response, and Cardiovascular Risk Biomarkers in Hypercholesterolemic Wistar Rats. J. Funct. Foods 2017, 36, 300–309. [Google Scholar] [CrossRef]
- Salami, M.; Tamtaji, O.R.; Mohammadifar, M.; Talaei, S.A.; Tameh, A.A.; Abed, A.R.; Shirkhoda, S.; Dadgostar, E.; Taghizadeh, M. Neuroprotective Effects of Onion (Allium cepa) Ethanolic Extract on Animal Model of Parkinson’s Disease Induced by 6-Hydroxydopamine: A Behavioral, Biochemical, and Histological Study. Gazi Med. J. 2020, 31, 25–29. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Al-Yousef, H.M.; Al-Qahtani, J.H.; Al-Said, M.S. A Hepatonephro-Protective Phenolic-Rich Extract from Red Onion (Allium cepa L.) Peels. Pak. J. Pharm. Sci. 2017, 30, 1971–1979. [Google Scholar] [PubMed]
- El-Hashim, A.Z.; Khajah, M.A.; Orabi, K.Y.; Balakrishnan, S.; Sary, H.G.; Abdelali, A.A. Onion Bulb Extract Downregulates EGFR/ERK1/2/AKT Signaling Pathway and Synergizes With Steroids to Inhibit Allergic Inflammation. Front. Pharmacol. 2020, 11, 551683. [Google Scholar] [CrossRef]
- Khajah, M.A.; EL-Hashim, A.Z.; Orabi, K.Y.; Hawai, S.; Sary, H.G. Onion Bulb Extract Can Both Reverse and Prevent Colitis in Mice via Inhibition of Pro-Inflammatory Signaling Molecules and Neutrophil Activity. PLoS ONE 2020, 15, e0233938. [Google Scholar] [CrossRef]
- Shokoohi, M.; Madarek, E.O.S.; Khaki, A.; Shoorei, H.; Khaki, A.A.; Soltani, M.; Ainehchi, N. Investigating the Effects of Onion Juice on Male Fertility Factors and Pregnancy Rate after Testicular Torsion/Detorsion by Intrauterine Insemination Method. Int. J. Women’s Health Reprod. Sci. 2018, 6, 499–505. [Google Scholar] [CrossRef]
- Alkhedaide, A.; Soliman, M.M.; Ismail, T.A. Protective effect of onion extract against experimental immunesuppression in wistar rats: Biological and molecular study. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 96–103. [Google Scholar] [CrossRef]
- Lamichhane, S.; Thapa, S. Advances from Conventional to Modern Plant Breeding Methodologies. Plant Breed. Biotechnol. 2022, 10, 1–14. [Google Scholar] [CrossRef]
- Bharti, G.; Chimata, M.K. Review on New Plant Breeding Techniques. Int. J. Sci. Res. 2019, 8, 723–730. [Google Scholar]
- Lema, M. Marker Assisted Selection in Comparison to Conventional Plant Breeding: Review Article. Agric. Res. Technol. 2018, 14, 555914. [Google Scholar] [CrossRef]
- Liang Jiang, G. Plant Marker-Assisted Breeding and Conventional Breeding: Challenges and Perspectives. Adv. Crop Sci. Technol. 2013, 1, e106. [Google Scholar] [CrossRef]
- McCallum, J. Genome Mapping and Molecular Breeding in Plants; Chittaranjan, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 5. [Google Scholar]
- Das, G.; Patra, J.K.; Baek, K.H. Insight into MAS: A Molecular Tool for Development of Stress Resistant and Quality of Rice through Gene Stacking. Front. Plant Sci. 2017, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Bark, O.H.; Havey, M.J. Similarities and Relationships among Populations of the Bulb Onion as Estimated by Nuclear RFLPs. Theor. Appl. Genet. 1995, 90, 407–414. [Google Scholar] [CrossRef]
- Bradeen, J.M.; Havey, M.J. Randomly Amplified Polymorphic DNA in Bulb Onion and Its Use to Assess Inbred Integrity. J. Am. Soc. Hortic. Sci. 1995, 120, 752–758. [Google Scholar] [CrossRef]
- Cramer, C.S.; Havey, M.J. Morphological, Biochemical, and Molecular Markers in Onion. HortScience 1999, 34, 590. [Google Scholar] [CrossRef]
- Duangjit, J.; Bohanec, B.; Chan, A.P.; Town, C.D.; Havey, M.J. Transcriptome Sequencing to Produce SNP-Based Genetic Maps of Onion. Theor. Appl. Genet. 2013, 126, 2093–2101. [Google Scholar] [CrossRef]
- Finkers, R.; van Kaauwen, M.; Ament, K.; Burger-Meijer, K.; Egging, R.; Huits, H.; Kodde, L.; Kroon, L.; Shigyo, M.; Sato, S.; et al. Insights from the First Genome Assembly of Onion (Allium cepa). G3 Genes Genomes Genet. 2021, 11, jkab243. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, K.; Zhang, G.; Zeng, F. Metabolite Profiling of Barley Grains Subjected to Water Stress: To Explain the Genotypic Difference in Drought-Induced Impacts on Malting Quality. Front. Plant Sci. 2017, 8, 1547. [Google Scholar] [CrossRef] [PubMed]
- Mitrová, K.; Svoboda, P.; Ovesná, J. The Selection and Validation of a Marker Set for the Differentiation of Onion Cultivars from the Czech Republic. Czech J. Genet. Plant Breed. 2015, 51, 62–67. [Google Scholar] [CrossRef]
- Hanci, F.; Gökçe, A.F. Molecular Characterization of Turkish Onion Germplasm Using SSR Markers. Czech J. Genet. Plant Breed. 2016, 52, 71–76. [Google Scholar] [CrossRef]
- González-Pérez, S.; Mallor, C.; Garcés-Claver, A.; Merino, F.; Taboada, A.; Rivera, A.; Pomar, F.; Perovic, D.; Silvar, C. Exploring Genetic Diversity and Quality Traits in a Collection of Onion (Allium cepa L.) Landraces from North-West Spain. Genetika 2015, 47, 885–900. [Google Scholar] [CrossRef]
- Rivera, A.; Mallor, C.; Garcés-Claver, A.; García-Ulloa, A.; Pomar, F.; Silvar, C. Assessing the Genetic Diversity in Onion (Allium cepa L.) Landraces from Northwest Spain and Comparison with the European Variability. N. Z. J. Crop Hortic. Sci. 2016, 44, 103–120. [Google Scholar] [CrossRef]
- Karić, L.; Golzardi, M.; Glamočlija, P.; Šutković, J. Genetic Diversity Assessment of Allium cepa L. Cultivars from Bosnia and Herzegovina Using SSR Makers. Genet. Mol. Res. 2018, 17, gmr16039870. [Google Scholar] [CrossRef]
- Ricciardi, L.; Mazzeo, R.; Marcotrigiano, A.R.; Rainaldi, G.; Iovieno, P.; Zonno, V.; Pavan, S.; Lotti, C. Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace. Plants 2020, 9, 260. [Google Scholar] [CrossRef]
- Villano, C.; Esposito, S.; Carucci, F.; Iorizzo, M.; Frusciante, L.; Carputo, D.; Aversano, R. High-Throughput Genotyping in Onion Reveals Structure of Genetic Diversity and Informative SNPs Useful for Molecular Breeding. Mol. Breed. 2019, 39, 5. [Google Scholar] [CrossRef]
- Adeyemo, O.A.; Oko, I.; Onadipe, A.S. Molecular Diversity of 3 Allium spp. Local Races Based on Rapd Data and Assessment of SSR Markers in Cross-Species Transferability; University of Lagos Research Repository: Lagos, Nigeria, 2018; Volume 3. [Google Scholar]
- Jayaswall, K.; Bhandawat, A.; Sharma, H.; Yadav, V.K.; Mahajan, V.; Singh, M. Characterization of Allium Germplasms for Conservation and Sustainable Management Using SSR Markers. Indian J. Tradit. Know. 2019, 18, 193–199. [Google Scholar]
- Monteverde, E.; Galván, G.A.; Speranza, P. Genetic Diversification of Local Onion Populations under Different Production Systems in Uruguay. Plant Genet. Resour. Characterisation Util. 2015, 13, 238–246. [Google Scholar] [CrossRef]
- Samiei, L.; Kiani, M.; Zarghami, H.; Memariani, F.; Joharchi, M.R. Genetic Diversity and Interspecific Relationships of Some Allium Species Using Inter Simple Sequence Repeat Markers. Bangladesh J. Plant Taxon. 2015, 22, 67. [Google Scholar] [CrossRef]
- Sudha, G.S.; Ramesh, P.; Sekhar, A.C.; Krishna, T.S.; Bramhachari, P.V.; Riazunnisa, K. Genetic Diversity Analysis of Selected Onion (Allium cepa L.) Germplasm Using Specific RAPD and ISSR Polymorphism Markers. Biocatal. Agric. Biotechnol. 2019, 17, 110–118. [Google Scholar] [CrossRef]
- Nikhil, B.S.K.; Jadhav, A.S. Characterization of Onion (Allium cepa L) Genotypes Using Molecular Markers. Int. J. Trop. Agric. 2017, 35, 25–31. [Google Scholar]
- Fang, H.; Liu, H.; Ma, R.; Liu, Y.; Li, J.; Yu, X.; Zhang, H.; Yang, Y.; Zhang, G. Genome-Wide Assessment of Population Structure and Genetic Diversity of Chinese Lou Onion Using Specific Length Amplified Fragment (SLAF) Sequencing. PLoS ONE 2020, 15, e0231753. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Williams, C.A.; Grayer, R.J. Anthocyanins and Other Flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef]
- Song, S.; Kim, C.-W.; Moon, J.S.; Kim, S. At Least Nine Independent Natural Mutations of the DFR-A Gene Are Responsible for Appearance of Yellow Onions (Allium cepa L.) from Red Progenitors. Mol. Breed. 2014, 33, 173–186. [Google Scholar] [CrossRef]
- Gu, K.-D.; Wang, C.-K.; Hu, D.-G.; Hao, Y.-J. How Do Anthocyanins Paint Our Horticultural Products? Sci. Hortic. 2019, 249, 257–262. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The Flavonoid Quercetin in Disease Prevention and Therapy: Facts and Fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef]
- Clere, N.; Faure, S.; Carmen Martinez, M.; Andriantsitohaina, R. Anticancer Properties of Flavonoids: Roles in Various Stages of Carcinogenesis. Cardiovasc. Hematol. Agents Med. Chem. (Former. Curr. Med. Chem.-Cardiovasc. Hematol. Agents) 2011, 9, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Nishiumi, S.; Miyamoto, S.; Kawabata, K.; Ohnishi, K.; Mukai, R.; Murakami, A.; Ashida, H.; Terao, J. Dietary Flavonoids as Cancer-Preventive and Therapeutic Biofactors. Front. Biosci.-Sch. 2011, 3, 1332–1362. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, O.; Shiojima, Y.; Igarashi, K. Anti-Obesity Effects of Onion Extract in Zucker Diabetic Fatty Rats. Nutrients 2012, 4, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Lotito, S.B.; Frei, B. Consumption of Flavonoid-Rich Foods and Increased Plasma Antioxidant Capacity in Humans: Cause, Consequence, or Epiphenomenon? Free Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef] [PubMed]
- Fossen, T.; Andersen, Ø.M.; Øvstedal, D.O.; Pedersen, A.T.; Raknes, Å. Characteristic Anthocyanin Pattern from Onions and Other Allium spp. J. Food Sci. 1996, 61, 703–706. [Google Scholar] [CrossRef]
- Meunissier, A. Expkriences Gknktiques Faitesii Verri2re. Bull. Soc. Acclim. Fr. 1918, 65, 81–90. [Google Scholar]
- Rieman, G.H. Genetic Factors for Pigmentation in the Onion and Their Relation to Disease Resistance; CABI Digital Library: Wallingford, CT, USA, 1931. [Google Scholar]
- Clarke, A.E.; Jones, H.A.; Little, T. Inheritance of Bulb Color in the Onion. Genetics 1944, 29, 569. [Google Scholar] [CrossRef]
- Jones, H.A. Complementary Factors for Light-Red Bulb Color in Onions. Proc. Am. Soc. Hortic. Sci. 1952, 59, 457. [Google Scholar]
- El-Shafie, M.; Davis, G.N. Inheritance of Bulb Color in the Onion (Allium cepa L.). Hilgardia 1967, 38, 607–622. [Google Scholar] [CrossRef]
- Khar, A.; Jakse, J.; Havey, M.J. Segregations for Onion Bulb Colors Reveal That Red Is Controlled by at Least Three Loci. J. Am. Soc. Hortic. Sci. 2008, 133, 42–47. [Google Scholar] [CrossRef]
- Duangjit, J.; Welsh, K.; Wise, M.L.; Bohanec, B.; Havey, M.J. Genetic Analyses of Anthocyanin Concentrations and Intensity of Red Bulb Color among Segregating Haploid Progenies of Onion. Mol. Breed. 2014, 34, 75–85. [Google Scholar] [CrossRef]
- Wilmouth, R.C.; Turnbull, J.J.; Welford, R.W.D.; Clifton, I.J.; Prescott, A.G.; Schofield, C.J. Structure and Mechanism of Anthocyanidin Synthase from Arabidopsis thaliana. Structure 2002, 10, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.; Carpenter, R.; Coen, E.S. A Common Gene Regulates Pigmentation Pattern in Diverse Plant Species. Cell 1992, 68, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Quattrocchio, F.; Wing, J.F.; Leppen, H.T.C.; Mol, J.N.M.; Koes, R.E. Regulatory Genes Controlling Anthocyanin Pigmentation Are Functionally Conserved among Plant Species and Have Distinct Sets of Target Genes. Plant Cell 1993, 5, 1497–1512. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jones, R.; Yoo, K.S.; Pike, L.M. The L Locus, One of Complementary Genes Required for Anthocyanin Production in Onions (Allium cepa), Encodes Anthocyanidin Synthase. Theor. Appl. Genet. 2005, 111, 120–127. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.S.; Pike, L.M. Development of a PCR-Based Marker Utilizing a Deletion Mutation in the Dihydroflavonol 4-Reductase (DFR) Gene Responsible for the Lack of Anthocyanin Production in Yellow Onions (Allium cepa). Theor. Appl. Genet. 2005, 110, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Cho, D.Y.; Moon, J.S.; Yoon, M.-K.; Kim, S. Development of Functional Markers for Detection of Inactive DFR-A Alleles Responsible for Failure of Anthocyanin Production in Onions (Allium cepa L.). Hortic. Sci. Technol. 2013, 31, 72–79. [Google Scholar] [CrossRef]
- Masuzaki, S.-I.; Shigyo, M.; Yamauchi, N. Complete Assignment of Structural Genes Involved in Flavonoid Biosynthesis Influencing Bulb Color to Individual Chromosomes of the Shallot (Allium cepa L.). Genes Genet. Syst. 2006, 81, 255–263. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.-S.; Pike, L.M. The Basic Color Factor, the C Locus, Encodes a Regulatory Gene Controlling Transcription of Chalcone Synthase Genes in Onions (Allium cepa). Euphytica 2005, 142, 273–282. [Google Scholar] [CrossRef]
- Kim, S.; Jones, R.; Yoo, K.S.; Pike, L.M. Gold Color in Onions (Allium cepa): A Natural Mutation of the Chalcone Isomerase Gene Resulting in a Premature Stop Codon. Mol. Genet. Genom. 2004, 272, 411–419. [Google Scholar] [CrossRef]
- Kim, S.; Binzel, M.L.; Park, S.; Yoo, K.S.; Pike, L.M. Inactivation of DFR (Dihydroflavonol 4-Reductase) Gene Transcription Results in Blockage of Anthocyanin Production in Yellow Onions (Allium cepa). Mol. Breed. 2004, 14, 253–263. [Google Scholar] [CrossRef]
- Jeon, S.; Han, J.; Moon, J.-H.; Kim, S. Identification of a Candidate Gene Responsible for the G Locus Determining Chartreuse Bulb Color in Onion (Allium cepa L.) Using Bulked Segregant RNA-Seq. Theor. Appl. Genet. 2022, 135, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.; Kim, S. Transposition of a Non-Autonomous DNA Transposon in the Gene Coding for a BHLH Transcription Factor Results in a White Bulb Color of Onions (Allium cepa L.). Theor. Appl. Genet. 2020, 133, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Seo, I.; Kim, J.G.; Moon, J.H.; Kim, S. Construction of a Linkage Map Flanking the I Locus Controlling Dominant White Bulb Color and Analysis of Differentially Expressed Genes between Dominant White and Red Bulbs in Onion (Allium cepa L.). Euphytica 2020, 216, 97. [Google Scholar] [CrossRef]
- Kim, S.; Baek, D.; Cho, D.Y.; Lee, E.-T.; Yoon, M.-K. Identification of Two Novel Inactive DFR-A Alleles Responsible for Failure to Produce Anthocyanin and Development of a Simple PCR-Based Molecular Marker for Bulb Color Selection in Onion (Allium cepa L.). Theor. Appl. Genet. 2009, 118, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Kim, C.W.; Kim, S. Identification of Two Novel Mutant ANS Alleles Responsible for Inactivation of Anthocyanidin Synthase and Failure of Anthocyanin Production in Onion (Allium cepa L.). Euphytica 2016, 212, 427–437. [Google Scholar] [CrossRef]
- Kim, B.; Cho, Y.; Kim, S. Identification of a Novel DFR-A Mutant Allele Determining the Bulb Color Difference between Red and Yellow Onions (Allium cepa L.). Plant Breed. Biotechnol. 2017, 5, 45–53. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Zhan, Z.; Cao, L.; Zeng, A.; Chang, G.; Liang, Y. Transcriptome Sequencing and Metabolism Analysis Reveals the Role of Cyanidin Metabolism in Dark-Red Onion (Allium cepa L.) Bulbs. Sci. Rep. 2018, 8, 14109. [Google Scholar] [CrossRef]
- Havey, M.J. Genetic Mapping of chartreuse Bulb Color in Onion. J. Am. Soc. Hortic. Sci. 2020, 145, 110–119. [Google Scholar] [CrossRef]
- Han, J.W.; Kim, C.W.; Kim, S. Genetic Analysis of the Interaction between G and R Loci Involved in the Determination of Bulb Colors in Onion (Allium cepa L.). Hortic. Environ. Biotechnol. 2023, 64, 801–810. [Google Scholar] [CrossRef]
- Schwinn, K.E.; Ngo, H.; Kenel, F.; Brummell, D.A.; Albert, N.W.; McCallum, J.A.; Pither-Joyce, M.; Crowhurst, R.N.; Eady, C.; Davies, K.M. The Onion (Allium cepa L.) R2R3-MYB Gene MYB1 Regulates Anthocyanin Biosynthesis. Front. Plant Sci. 2016, 7, 1865. [Google Scholar] [CrossRef] [PubMed]
- Baek, G.; Kim, C.-W.; Kim, S. Development of a Molecular Marker Tightly Linked to the C Locus Conferring a White Bulb Color in Onion (Allium cepa L.) Using Bulked Segregant Analysis and RNA-Seq. Mol. Breed. 2017, 37, 1–13. [Google Scholar] [CrossRef]
- Li, X.; Cao, L.; Jiao, B.; Yang, H.; Ma, C.; Liang, Y. The BHLH Transcription Factor AcB2 Regulates Anthocyanin Biosynthesis in Onion (Allium cepa L.). Hortic. Res. 2022, 9, uhac128. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Butt, M.S.; Anjum, F.M.; Saeed, F.; Khalid, N. Onion: Nature Protection against Physiological Threats. Crit. Rev. Food Sci. Nutr. 2015, 55, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Gubb, I.R.; Mactavish, H.S. Onion pre-and postharvest considerations. In Allium Crop Science: Recent Advances; CABI Digital Library: Wallingford, UK, 2002; pp. 233–265. [Google Scholar] [CrossRef]
- McCallum, J.; Clarke, A.; Pither-Joyce, M.; Shaw, M.; Butler, R.; Brash, D.; Scheffer, J.; Sims, I.; Van Heusden, S.; Shigyo, M.; et al. Genetic Mapping of a Major Gene Affecting Onion Bulb Fructan Content. Theor. Appl. Genet. 2006, 112, 958–967. [Google Scholar] [CrossRef]
- Darbyshire, B.; Henry, R.J. Differences in Fructan Content and Synthesis in Some Allium Species. N. Phytol. 1981, 87, 249–256. [Google Scholar] [CrossRef]
- Taper, H.S.; Delzenne, N.M.; Roberfroid, M.B. Growth Inhibition of Transplantable Mouse Tumors by Non-Digestible Carbohydrates. Int. J. Cancer 1997, 71, 1109–1112. [Google Scholar] [CrossRef]
- Rowland, I.R.; Rumney, C.J.; Coutts, J.T.; Lievense, L.C. Effect of Bifidobacterium Longum and Inulin on Gut Bacterial Metabolism and Carcinogen-Induced Aberrant Crypt Foci in Rats. Carcinogenesis 1998, 19, 281–285. [Google Scholar] [CrossRef]
- Gibson, G.R.; Wang, X. Regulatory Effects of Bifidobacteria on the Growth of Other Colonic Bacteria. J. Appl. Microbiol. 1994, 77, 412–420. [Google Scholar] [CrossRef]
- Kleessen, B.; Svkura, B.; Zunft, H.-J.; Blaut, M. Effects of Inulin and Lactose on Fecal Microflora, Microbial Activity, and Bowel Habit in Elderly Constipated Persons. Am. J. Clin. Nutr. 1997, 65, 1397–1402. [Google Scholar] [CrossRef]
- Niness, K.R. Inulin and Oligofructose: What Are They? J. Nutr. 1999, 129, 1402S–1406S. [Google Scholar] [CrossRef]
- Sinclair, P.J.; Blakeney, A.B.; Barlow, E.W.R. Relationships between Bulb Dry Matter Content, Soluble Solids Concentration and Non-Structural Carbohydrate Composition in the Onion (Allium cepa). J. Sci. Food Agric. 1995, 69, 203–209. [Google Scholar] [CrossRef]
- Lin, M.-W.; Watson, J.F.; Baggett, J.R. Inheritance of Soluble Solids and Pyruvic Acid Content of Bulb Onions. J. Am. Soc. Hortic. Sci. 1995, 120, 119–122. [Google Scholar] [CrossRef]
- Suzuki, M.; Cutcliffe, J.A. Fructans in Onion Bulbs in Relation to Storage Life. Can. J. Plant Sci. 1989, 69, 1327–1333. [Google Scholar] [CrossRef]
- Debaene, J.E.P.; Goldman, I.L.; Yandell, B.S. Postharvest Flux and Genotype × Environment Effects for Onion-Induced Antiplatelet Activity, Pungency, and Soluble Solids in Long-Day Onion during Postharvest Cold Storage. J. Am. Soc. Hortic. Sci. 1999, 124, 366–372. [Google Scholar] [CrossRef]
- Mcgill, H.C.; Mcmahan, C.A.; Zieske, A.W.; Tracy, R.E.; Malcom, G.T.; Herderick, E.E.; Strong, J.P. Association of Coronary Heart Disease Risk Factors With Microscopic Qualities of Coronary Atherosclerosis in Youth. Circulation 2000, 102, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Jaime, L.; Martín-Cabrejas, M.A.; Mollá, E.; López-Andréu, F.J.; Esteban, R.M. Effect of Storage on Fructan and Fructooligosaccharide of Onion (Allium cepa L.). J. Agric. Food Chem. 2001, 49, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, N.; Onodera, S.; Sakai, H. Fructo-Oligosaccharide Content and Fructosyltransferase Activity during Growth of Onion Bulbs. N. Phytol. 1997, 136, 105–113. [Google Scholar] [CrossRef]
- Pak, C.; van der Plas, L.H.W.; de Boer, A.D. Importance of Dormancy and Sink Strength in Sprouting of Onions (Allium cepa) during Storage. Physiol. Plant 1995, 94, 277–283. [Google Scholar] [CrossRef]
- Van Den Ende, W.; Clerens, S.; Vergauwen, R.; Boogaerts, D.; Le Roy, K.; Arckens, L.; Van Laere, A. Cloning and Functional Analysis of a High DP Fructan:Fructan 1-Fructosyl Transferase from Echinops Ritro (Asteraceae): Comparison of the Native and Recombinant Enzymes. J. Exp. Bot. 2006, 57, 775–789. [Google Scholar] [CrossRef]
- Vijn, I.; Van Dijken, A.; Sprenger, N.; Van Dun, K.; Weisbeek, P.; Wiemken, A.; Smeekens, S. Fructan of the Inulin Neoseries Is Synthesized in Transgenic Chicory Plants (Cichorium intybus L.) Harbouring Onion (Allium cepa L.) Fructan:Fructan 6G-Fructosyltransferase. Plant J. 1997, 11, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Onodera, S.; Kawakami, A.; Yoshida, M.; Shiomi, N. Molecular Characterization and Expression of a CDNA Encoding Fructan:Fructan 6G-Fructosyltransferase from Asparagus (Asparagus Officinalis). N. Phytol. 2005, 165, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Romero, C.; Martínez-Hernández, A.; Mellado-Mojica, E.; López, M.G.; Simpson, J. Molecular and Functional Characterization of Novel Fructosyltransferases and Invertases from Agave tequilana. PLoS ONE 2012, 7, e35878. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Yoshida, M.; Van Den Ende, W. Molecular Cloning and Functional Analysis of a Novel 6&1-FEH from Wheat (Triticum aestivum L.) Preferentially Degrading Small Graminans like Bifurcose. Gene 2005, 358, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.I.; Sanada, Y.; Tase, K.; Komatsu, T.; Yoshida, M. Pp6-FEH1 Encodes an Enzyme for Degradation of Highly Polymerized Levan and Is Transcriptionally Induced by Defoliation in Timothy (Phleum pratense L.). J. Exp. Bot. 2011, 62, 3421–3431. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liang, M.; Xu, L.; Li, H.; Zhang, X.; Kang, J.; Zhao, Q.; Zhao, H. Cloning and Functional Characterization of Two Abiotic Stress-Responsive Jerusalem Artichoke (Helianthus tuberosus) Fructan 1-Exohydrolases (1-FEHs). Plant Mol. Biol. 2015, 87, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Valluru, R. Fructan and Hormone Connections. Front. Plant Sci. 2015, 6, 180. [Google Scholar] [CrossRef]
- Havey, M.J.; Galmarini, C.R.; Gökçe, A.F.; Henson, C. QTL Affecting Soluble Carbohydrate Concentration in Stored Onion Bulbs and Their Association with Flavor and Health-Enhancing Attributes. Genome 2004, 47, 463–468. [Google Scholar] [CrossRef]
- Raines, S.; Henson, C.; Havey, M.J. Genetic Analyses of Soluble Carbohydrate Concentrations in Onion Bulbs. J. Am. Soc. Hortic. Sci. 2009, 134, 618–623. [Google Scholar] [CrossRef]
- Imai, S.; Tsuge, N.; Tomotake, M.; Nagatome, Y.; Sawada, H.; Nagata, T.; Kumagai, H. An Onion Enzyme That Makes the Eyes Water. Nature 2002, 419, 685. [Google Scholar] [CrossRef]
- Khan, M.S.; Haas, F.H.; Allboje Samami, A.; Moghaddas Gholami, A.; Bauer, A.; Fellenberg, K.; Reichelt, M.; Hänsch, R.; Mendel, R.R.; Meyer, A.J. Sulfite Reductase Defines a Newly Discovered Bottleneck for Assimilatory Sulfate Reduction and Is Essential for Growth and Development in Arabidopsis thaliana. Plant Cell 2010, 22, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Galmarini, C.R.; Goldman, I.L.; Havey, M.J. Genetic Analyses of Correlated Solids, Flavor, and Health-Enhancing Traits in Onion (Allium cepa L.). Mol. Genet. Genom. 2001, 265, 543–551. [Google Scholar] [CrossRef] [PubMed]
- McCallum, J.; Pither-Joyce, M.; Shaw, M.; Kenel, F.; Davis, S.; Butler, R.; Scheffer, J.; Jakse, J.; Havey, M.J. Genetic Mapping of Sulfur Assimilation Genes Reveals a QTL for Onion Bulb Pungency. Theor. Appl. Genet. 2007, 114, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Masamura, N.; McCallum, J.; Khrustaleva, L.; Kenel, F.; Pither-Joyce, M.; Shono, J.; Suzuki, G.; Mukai, Y.; Yamauchi, N.; Shigyo, M. Chromosomal Organization and Sequence Diversity of Genes Encoding Lachrymatory Factor Synthase in Allium cepa L. G3 Genes Genomes Genet. 2012, 2, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A Global Benefit to Health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Schwimmer, S.; Weston, W.J. Onion Flavor and Odor, Enzymatic Development of Pyruvic Acid in Onion as a Measure of Pungency. J. Agric. Food Chem. 1961, 9, 301–304. [Google Scholar] [CrossRef]
- Randle, W.M. Onion Flavor Chemistry and Factors Influencing Flavor Intensity; ACS Publications: Washington, DC, USA, 1997; ISBN 1947-5918. [Google Scholar]
- Randle, W.M.; Lancaster, J.E. Sulphur Compounds in Alliums in Relation to Flavour Quality. In Allium Crop Science: Recent Advances; CABI Digital Library: Wallingford, UK, 2002; pp. 329–356. [Google Scholar] [CrossRef]
- Schwimmer, S.; Guadagni, D.G. Relation between Olfactory Threshold Concentration and Pyruvic Acid Content of Onion Juice. J. Food Sci. 1962, 27, 94–97. [Google Scholar] [CrossRef]
- McManus, M.T.; Joshi, S.; Searle, B.; Pither-Joyce, M.; Shaw, M.; Leung, S.; Albert, N.; Shigyo, M.; Jakse, J.; Havey, M.J.; et al. Genotypic Variation in Sulfur Assimilation and Metabolism of Onion (Allium cepa L.) III. Characterization of Sulfite Reductase. Phytochemistry 2012, 83, 34–42. [Google Scholar] [CrossRef]
- Rabinowitch, H.D. Onions and Allied Crops; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Heath, O.V.S. Studies in the Physiology of the Onion Plant: I. An Investigation of Factors Concerned in the Flowering (‘Bolting’) of Onions Grown from Sets and Its Prevention. Ann. Appl. Biol. 1943, 30, 208–220. [Google Scholar] [CrossRef]
- Heath, O.V.S.; Mathur, P.B. Studies in the Physiology of the Onion Plant: II. Inflorescence Initiation and Development, and Other Changes in the Internal Morphology of Onion Sets, as Influenced by Temperature and Day Length. Ann. Appl. Biol. 1944, 31, 173–186. [Google Scholar] [CrossRef]
- Austin, R.B. Bulb Formation in Onions as Affected by Photoperiod and Spectral Quality of Light. J. Hortic. Sci. 1972, 47, 493–504. [Google Scholar] [CrossRef]
- Lercari, B. Role of Phytochrome in Photoperiodic Regulation of Bulbing and Growth in the Long Day Plant Allium cepa. Physiol. Plant 1984, 60, 433–436. [Google Scholar] [CrossRef]
- Sobeih, W.Y.; Wright, C.J. The Photoperiodic Regulation of Bulbing in Onions (Allium cepa L.). II. Effects of Plant Age and Size. J. Hortic. Sci. 1986, 61, 337–341. [Google Scholar] [CrossRef]
- Brewster, J.L. Environmental Physiology of the Onion: Towards Quantitative Models for the Effects of Photoperiod, Temperature and Irradiance on Bulbing, Flowering and Growth. In Proceedings of the I International Symposium on Edible Alliaceae 433, Mendoza, Argentina, 14–18 March 1994; pp. 347–374. [Google Scholar]
- Brewster, J.L. Onions and Other Vegetable Alliums; CABI: Wallingford, UK, 2008; Volume 15, ISBN 1845936221. [Google Scholar]
- Shishido, Y.; Saito, T. Studies on the Flower Bud Formation in Onion Plants I. Effects of Temperature, Photoperiod and Light Intensity on the. Low Temperature Induction of Flower Buds. J. Jpn. Soc. Hortic. Sci. 1975, 44, 122–130. [Google Scholar] [CrossRef]
- Wiebe, H.-J. Vernalization of Vegetable Crops-a Review. In Proceedings of the VI Symposium on the Timing of Field Production of Vegetables 267, Wageningen, The Netherlands, 21–25 August 1989; pp. 323–328. [Google Scholar]
- Khokhar, K.M.; Hadley, P.; Pearson, S. Effect of Cold Temperature Durations of Onion Sets in Store on the Incidence of Bolting, Bulbing and Seed Yield. Sci. Hortic. 2007, 112, 16–22. [Google Scholar] [CrossRef]
- Sung, S.; Schmitz, R.J.; Amasino, R.M. A PHD Finger Protein Involved in Both the Vernalization and Photoperiod Pathways in Arabidopsis. Genes. Dev. 2006, 20, 3244–3248. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a BZIP Protein Mediating Signals from the Floral Pathway Integrator FT at the Shoot Apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef]
- Yoo, S.K.; Chung, K.S.; Kim, J.; Lee, J.H.; Hong, S.M.; Yoo, S.J.; Yoo, S.Y.; Lee, J.S.; Ahn, J.H. Constans Activates Suppressor of Overexpression of Constans 1 through Flowering Locus T to Promote Flowering in Arabidopsis. Plant Physiol. 2005, 139, 770–778. [Google Scholar] [CrossRef]
- Greb, T.; Mylne, J.S.; Crevillen, P.; Geraldo, N.; An, H.; Gendall, A.R.; Dean, C. The PHD Finger Protein VRN5 Functions in the Epigenetic Silencing of Arabidopsis FLC. Curr. Biol. 2007, 17, 73–78. [Google Scholar] [CrossRef]
- Nakamichi, N. Molecular Mechanisms Underlying the Arabidopsis Circadian Clock. Plant Cell Physiol. 2011, 52, 1709–1718. [Google Scholar] [CrossRef]
- Whittaker, C.; Dean, C. The FLC Locus: A Platform for Discoveries in Epigenetics and Adaptation. Annu. Rev. Cell Dev. Biol. 2017, 33, 555–575. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Robson, F.; Lee, K.; Simon, R.; Coupland, G. The CONSTANS Gene of Arabidopsis Promotes Flowering and Encodes a Protein Showing Similarities to Zinc Finger Transcription Factors. Cell 1995, 80, 847–857. [Google Scholar] [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA Complex Formation Is Required for Day-Length Measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, T.; Schultz, T.F.; Harmon, F.G.; Ho, L.A.; Kay, S.A. FKF1 F-Box Protein Mediates Cyclic Degradation of a Repressor of CONSTANS in Arabidopsis. Science 2005, 309, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Marchal, V.; Panigrahi, K.C.S.; Wenkel, S.; Soppe, W.; Deng, X.; Valverde, F.; Coupland, G. Arabidopsis COP1 Shapes the Temporal Pattern of CO Accumulation Conferring a Photoperiodic Flowering Response. EMBO J. 2008, 27, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Fornara, F.; de Montaigu, A.; Coupland, G. SnapShot: Control of Flowering in Arabidopsis. Cell 2010, 141, 550. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering Time Regulation: Photoperiod-and Temperature-Sensing in Leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef]
- Song, J.; Angel, A.; Howard, M.; Dean, C. Vernalization–a Cold-Induced Epigenetic Switch. J. Cell Sci. 2012, 125, 3723–3731. [Google Scholar] [CrossRef]
- Navarro, C.; Abelenda, J.A.; Cruz-Oró, E.; Cuéllar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of Flowering and Storage Organ Formation in Potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef]
- Hsu, C.-Y.; Adams, J.P.; Kim, H.; No, K.; Ma, C.; Strauss, S.H.; Drnevich, J.; Vandervelde, L.; Ellis, J.D.; Rice, B.M. FLOWERING LOCUS T Duplication Coordinates Reproductive and Vegetative Growth in Perennial Poplar. Proc. Natl. Acad. Sci. USA 2011, 108, 10756–10761. [Google Scholar] [CrossRef]
- Lee, R.; Baldwin, S.; Kenel, F.; McCallum, J.; Macknight, R. FLOWERING LOCUS T Genes Control Onion Bulb Formation and Flowering. Nat. Commun. 2013, 4, 2884. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Massiah, A.J.; Thomas, B. Conservation of Arabidopsis Thaliana Photoperiodic Flowering Time Genes in Onion (Allium cepa L.). Plant Cell Physiol. 2010, 51, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.A.; Thomas, B. Diurnal Expression of Arabidopsis Gene Homologs during Daylength-Regulated Bulb Formation in Onion (Allium cepa L.). Sci. Hortic. 2020, 261, 108946. [Google Scholar] [CrossRef]
- Baldwin, S.; Revanna, R.; Pither-Joyce, M.; Shaw, M.; Wright, K.; Thomson, S.; Moya, L.; Lee, R.; Macknight, R.; McCallum, J. Genetic Analyses of Bolting in Bulb Onion (Allium cepa L.). Theor. Appl. Genet. 2014, 127, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Khar, A.; Galván, G.A.; Singh, H. Allium Breeding against Biotic Stresses. In Genomic Designing for Biotic Stress Resistant Vegetable Crops; Chittaranjan, K., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 233–259. [Google Scholar]
- Straley, E.; Marzu, J.C.; Havey, M.J. Genetic Analyses of Resistance to Fusarium Basal Rot in Onion. Horticulturae 2021, 7, 538. [Google Scholar] [CrossRef]
- Taylor, A.; Teakle, G.R.; Walley, P.G.; Finch-Savage, W.E.; Jackson, A.C.; Jones, J.E.; Hand, P.; Thomas, B.; Havey, M.J.; Pink, D.A.C.; et al. Assembly and Characterisation of a Unique Onion Diversity Set Identifies Resistance to Fusarium Basal Rot and Improved Seedling Vigour. Theor. Appl. Genet. 2019, 132, 3245–3264. [Google Scholar] [CrossRef] [PubMed]
- Black, L.; Chan, E.K.; Colcol, J.F.; Jones, R.; Kramer, C.; Xiang, W. QTLs Conferring Resistance to Fusarium Basal Rot, Pink Root and Complementary Pinks in Onions. U.S. Patent 11,457,582, 4 October 2022. [Google Scholar]
- Marzu, J.C.; Straley, E.; Havey, M.J. Genetic Analyses and Mapping of Pink-Root Resistance in Onion. J. Am. Soc. Hortic. Sci. 2018, 143, 503–507. [Google Scholar] [CrossRef]
- Chakraborty, M.; Quddus, T.; Rahman, S.; Azad, M.; Hossain, M.; Hoque, A.; Saha, M.; Haque, M. Molecular Characterization of Selected Mutant Lines of Onion (Allium cepa L.) against Purple Leaf Blotch Disease Using SSR Markers. Am. J. Exp. Agric. 2015, 8, 261–267. [Google Scholar] [CrossRef]
- Chand, S.K.; Nanda, S.; Joshi, R.K. Genetics and Molecular Mapping of a Novel Purple Blotch-Resistant Gene ApR1 in Onion (Allium cepa L.) Using STS and SSR Markers. Mol. Breed. 2018, 38, 109. [Google Scholar] [CrossRef]
- Sahoo, J.; Mahanty, B.; Mishra, R.; Joshi, R.K. Development of SNP Markers Linked to Purple Blotch Resistance for Marker-Assisted Selection in Onion (Allium cepa L.) Breeding. 3 Biotech 2023, 13, 137. [Google Scholar] [CrossRef]
- Kim, S.; Kim, C.W.; Choi, M.S.; Kim, S. Development of a Simple PCR Marker Tagging the Allium Roylei Fragment Harboring Resistance to Downy Mildew (Peronospora Destructor) in Onion (Allium cepa L.). Euphytica 2016, 208, 561–569. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, J.S.; Park, T.; Lee, H.M.; Choi, J.R.; Park, Y.D. Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through Rapd-Pcr and Transcriptome Analysis. Horticulturae 2021, 7, 436. [Google Scholar] [CrossRef]
- Brayford, D. Fusarium Oxysporum f. Sp. Cepae. Mycopathologia 1996, 133, 39–40. [Google Scholar]
- Cramer, C.S. Breeding and Genetics of Fusarium Basal Rot Resistance in Onion. Euphytica 2000, 115, 159–166. [Google Scholar] [CrossRef]
- Bacher, J.W. Inheritance of Resistance to Fusarium Oxysporum f. Sp. Cepae in Cultivated Onions; Michigan State University: East Lansing, MI, USA, 1989; ISBN 9798641845807. [Google Scholar]
- Tsutsui, K. Inheritance of Resistance to Fusarium Oxysporum in Onion; University of Wisconsin: Madison, WI, USA, 1991. [Google Scholar]
- Krueger, S.K. Resistance to Fusarium Basal Rot in Onions (Allium cepa L.); University of Wisconsin: Madison, WI, USA, 1986. [Google Scholar]
- Krueger, S.K.; Weinman, A.A.; Gabelman, W.H. Combining Ability among Inbred Onions for Resistance to Fusarium Basal Rot. HortScience 1989, 24, 1021–1023. [Google Scholar] [CrossRef]
- Kehr, A.E.; O’Brien, M.J.; Davis, E.W. Pathogenicity of Fusarium oxysporum f. Sp. Cepae and Its Interaction with Pyrenochaeta terrestris on Onion. Euphytica 1962, 11, 197–208. [Google Scholar] [CrossRef]
- Lorbeer, J.W.; Stone, K.W. Reaction of Onion to Fusarium Basal Rot. Plant Dis. Rep. 1965, 49, 522–526. [Google Scholar]
- Holz, G.; Knox-Davies, P.S. Resistance of Onion Selections to Fusarium oxysporum f. sp. Cepae* (Met Opsomming in Afrikaans). Phytophylactica 1974, 6, 153–156. [Google Scholar]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two Classes of Plant Antibiotics: Phytoalexins versus “Phytoanticipins”. Plant Cell 1994, 6, 1191. [Google Scholar] [CrossRef]
- Abdelrahman, M.; El-Sayed, M.; Sato, S.; Hirakawa, H.; Ito, S.I.; Tanaka, K.; Mine, Y.; Sugiyama, N.; Suzuki, M.; Yamauchi, N.; et al. RNA-Sequencing-Based Transcriptome and Biochemical Analyses of Steroidal Saponin Pathway in a Complete Set of Allium fistulosum—A. cepa Monosomic Addition Lines. PLoS ONE 2017, 12, e0181784. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular Activities, Biosynthesis and Evolution of Triterpenoid Saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.P.; Osbourn, A.E. Fungal Resistance to Plant Antibiotics as a Mechanism of Pathogenesis. Microbiol. Mol. Biol. Rev. 1999, 63, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.I.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H.; Dissanayake, C.; Abdel-Motaal, F.F.; El-Sayed, M.A. α-Tomatine, the Major Saponin in Tomato, Induces Programmed Cell Death Mediated by Reactive Oxygen Species in the Fungal Pathogen Fusarium oxysporum. FEBS Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.Q.; El-Sayed, M.A.; Ito, S.I.; Yamauchi, N.; Shigyo, M. Discovery of a New Source of Resistance to Fusarium oxysporum, Cause of Fusarium Wilt in Allium fistulosum, Located on Chromosome 2 of Allium cepa Aggregatum Group. Genome 2012, 55, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Teshima, Y.; Ikeda, T.; Imada, K.; Sasaki, K.; El-Sayed, M.A.; Shigyo, M.; Tanaka, S.; Ito, S.I. Identification and Biological Activity of Antifungal Saponins from Shallot (Allium cepa L. Aggregatum Group). J. Agric. Food Chem. 2013, 61, 7440–7445. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Lee, S.B.; Shim, H.S.; Lee, C.J.; Kim, H.D. Pink Root of Onion Caused by Pyrenochaeta terrestris (Syn. Phoma Terrestris). Plant Pathol. J. 2003, 19, 195–199. [Google Scholar] [CrossRef]
- Jones, H.A.; Perry, B.A. Inheritance of Resistance to Pink Root in the Onion. J. Hered. 1956, 47, 33–34. [Google Scholar] [CrossRef]
- Nichols, C.G.; Gabelman, W.H.; Larson, R.H.; Walker, J.C. The Expression and Inheritance of Resistance in Onion to Pink Root in Seedling Assays. Phytopathology 1965, 55, 752–756. [Google Scholar]
- Dar, A.A.; Sharma, S.; Mahajan, R.; Mushtaq, M.; Salathia, A.; Ahamad, S.; Sharma, J.P. Overview of Purple Blotch Disease and Understanding Its Management through Chemical, Biological and Genetic Approaches. J. Integr. Agric. 2020, 19, 3013–3024. [Google Scholar] [CrossRef]
- Kareem, M.A.; Murthy, K.V.; Hasansab, A.N.; Waseem, M.A. Effect of Temperature, Relative Humidity and Light on Lesion Length Due to Alternaria Porri in Onion. BIOINFOLET-A Q. J. Life Sci. 2012, 9, 264–266. [Google Scholar]
- Miller, M.E.; Lacy, M.L. Purple Blotch. In Compendium of Onion and Garlic Disease; Schwartz, H.F., Mohan, S.K., Eds.; APS Press: St. Paul, MN, USA, 1995. [Google Scholar]
- Suheri, H.; Price, T.V. The Epidemiology of Purple Leaf Blotch on Leeks in Victoria, Australia. Eur. J. Plant Pathol. 2001, 107, 503–510. [Google Scholar] [CrossRef]
- Yarwood, C.E. Onion Downy Mildew. Hilgardia 1943, 14, 595–691. [Google Scholar] [CrossRef]
- Schwartz, H.F.; Mohan, S.K. Compendium of Onion and Garlic Diseases and Pests, 2nd ed.; APS Press: St. Paul, MN, USA, 2008. [Google Scholar]
- Kofoet, A.; Kik, C.; Wietsma, W.A.; De Vries, J.N. Inheritance of Resistance to Downy Mildew (Peronospora destructor [Berk.] Casp.) from Allium Roylei Stearn in the Backcross Allium cepa L.×(A. Roylei× A. cepa). Plant Breed. 1990, 105, 144–149. [Google Scholar] [CrossRef]
- Van der Meer, Q.P.; De Vries, J.N. An Interspecific Cross between Allium roylei Stearn and Allium cepa L., and Its Backcross to A. cepa. Euphytica 1990, 47, 29–31. [Google Scholar] [CrossRef]
- Harrewijn, J.L.; Hoogenboom, J.P.H. Resistance to Downy Mildew of Onion Caused by the Fungus Peronospora Destructor 2014. U.S. Patent 8,119,856, 21 February 2012. [Google Scholar]
- Kim, S.; Kim, M.S.; Kim, Y.M.; Yeom, S.I.; Cheong, K.; Kim, K.T.; Jeon, J.; Kim, S.; Kim, D.S.; Sohn, S.H.; et al. Integrative Structural Annotation of de Novo RNA-Seq Provides an Accurate Reference Gene Set of the Enormous Genome of the Onion (Allium cepa L.). DNA Res. 2015, 22, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Khrustaleva, L.; Mardini, M.; Kudryavtseva, N.; Alizhanova, R.; Romanov, D.; Sokolov, P.; Monakhos, G. The Power of Genomic in Situ Hybridization (GISH) in Interspecific Breeding of Bulb Onion (Allium cepa L.) Resistant to Downy Mildew (Peronospora destructor [Berk.] Casp.). Plants 2019, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.M.; Simon, A.; Viaud, M. Botrytis cinerea Virulence Factors: New Insights into a Necrotrophic and Polyphageous Pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef]
- Carisse, O.; Tremblay, D.M.; McDonald, M.R.; Brodeur, L.; McRoberts, N. Managemant of Botrytis Leaf Blight of Onion: The Québec Experience of 20 Tears of Continual Improvement. Plant Dis. 2011, 95, 504–514. [Google Scholar] [CrossRef]
- Lee, H.M.; Park, J.S.; Kim, S.J.; Kim, S.G.; Park, Y.D. Using Transcriptome Analysis to Explore Gray Mold Resistance-Related Genes in Onion (Allium cepa L.). Genes 2022, 13, 542. [Google Scholar] [CrossRef]
- Hanson, M.R.; Bentolila, S. Interactions of Mitochondrial and Nuclear Genes That Affect Male Gametophyte Development. Plant Cell 2004, 16, S154–S169. [Google Scholar] [CrossRef]
- Tang, H.; Xie, Y.; Liu, Y.G.; Chen, L. Advances in Understanding the Molecular Mechanisms of Cytoplasmic Male Sterility and Restoration in Rice. Plant Reprod. 2017, 30, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.A.; Emsweller, S.L. A Male-Sterile Onion. In Proceedings of the American Society for Horticultural Science, Atlantic City, NJ, USA, 29–31 December 1936; pp. 582–585. [Google Scholar]
- Jones HA, C.A. Inheritance of Male-Sterility in the Onion and the Production of Hybrid Seed. Proc. Am. Soc. Hortic. Sci. 1943, 43, 189–194. [Google Scholar]
- Berninger, E. Contribution to the Study of Male Sterility in the Onion (Allium cepa L.). Ann. L’amélioration Plantes 1965, 15, 183–199. [Google Scholar]
- Schweisguth, B. A New Type of Male-Sterility in Onion, Allium cepa L. Ann. Amélior Plant 1973, 23, 221–233. [Google Scholar]
- Yen, D.E. Pollen Sterility in Pukekohe Longkeeper Onions. N. Z. J. Agric. Res. 1959, 2, 605–612. [Google Scholar] [CrossRef]
- Havey, M.J.; Randle, W.M. Combining Abilities for Yield and Bulb Quality among Long-and Intermediate-Day Open-Pollinated Onion Populations. J. Am. Soc. Hortic. Sci. 1996, 121, 604–608. [Google Scholar] [CrossRef]
- Havey, M.J. A Putative Donor of S-Cytoplasm and Its Distribution among Open-Pollinated Populations of Onion. Theor. Appl. Genet. 1993, 86, 128–134. [Google Scholar] [CrossRef]
- Havey, M.J. Identification of Cytoplasms Using the Polymerase Chain Reaction to Aid in the Extraction of Maintainer Lines from Open-Pollinated Populations of Onion. Theor. Appl. Genet. 1995, 90, 263–268. [Google Scholar] [CrossRef]
- Khar, A.; Singh, H. Rapid Methods for Onion Breeding. In Accelerated Plant Breeding; Singh, G.S., Hussain, W.S., Eds.; Springer: Cham, Switzerland, 2020; Volume 2, pp. 77–99. [Google Scholar]
- De Courcel, A.G.; Vedel, F.; Boussac, J.M. DNA Polymorphism in Allium cepa Cytoplasms and Its Implications Concerning the Origin of Onions. Theor. Appl. Genet. 1989, 77, 793–798. [Google Scholar] [CrossRef]
- Havey, M.J. On the Origin and Distribution of Normal Cytoplasm of Onion. Genet. Resour. Crop Evol. 1997, 44, 307–313. [Google Scholar] [CrossRef]
- Holford, P.; Croft, J.H.; Newbury, H.J. Differences between, and Possible Origins of, the Cytoplasms Found in Fertile and Male-Sterile Onions (Allium cepa L.). Theor. Appl. Genet. 1991, 82, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Havey, M.J. Phylogenetic Relationships among Cultivated Allium Species from Restriction Enzyme Analysis of the Chloroplast Genome. Theor. Appl. Genet. 1991, 81, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Havey, M.J. Diversity among Male-Sterility-Inducing and Male-Fertile Cytoplasms of Onion. Theor. Appl. Genet. 2000, 101, 778–782. [Google Scholar] [CrossRef]
- Satoh, Y.; Nagai, M.; Mikami, T.; Kinoshita, T. The Use of Mitochondrial DNA Polymorphism in the Classification of Individual Onion Plants by Cytoplasmic Genotypes. Theor. Appl. Genet. 1993, 86, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y. PCR Amplification of CMS-Specific Mitochondrial Nucleotide Sequences to Identify Cytoplasmic Genotypes of Onion (Allium cepa L.); Springer: Berlin/Heidelberg, Germany, 1998; Volume 96. [Google Scholar]
- Engelke, T.; Terefe, D.; Tatlioglu, T. A PCR-Based Marker System Monitoring CMS-(S), CMS-(T) and (N)-Cytoplasm in the Onion (Allium cepa L.). Theor. Appl. Genet. 2003, 107, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, E.T.; Cho, D.Y.; Han, T.; Bang, H.; Patil, B.S.; Ahn, Y.K.; Yoon, M.K. Identification of a Novel Chimeric Gene, Orf725, and Its Use in Development of a Molecular Marker for Distinguishing among Three Cytoplasm Types in Onion (Allium cepa L.). Theor. Appl. Genet. 2009, 118, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Von Kohn, C.; Kiełkowska, A.; Havey, M.J.; Bonen, L. Sequencing and Annotation of the Chloroplast DNAs and Identification of Polymorphisms Distinguishing Normal Male-Fertile and Male-Sterile Cytoplasms of Onion. Genome 2013, 56, 737–742. [Google Scholar] [CrossRef]
- Kim, S.; Kim, C.W.; Park, M.; Choi, D. Identification of Candidate Genes Associated with Fertility Restoration of Cytoplasmic Male-Sterility in Onion (Allium cepa L.) Using a Combination of Bulked Segregant Analysis and RNA-Seq. Theor. Appl. Genet. 2015, 128, 2289–2299. [Google Scholar] [CrossRef]
- Kim, B.; Kim, C.W.; Kim, S. Inheritance of Fertility Restoration of Male-Sterility Conferred by Cytotype Y and Identification of Instability of Male Fertility Phenotypes in Onion (Allium cepa L.). J. Hortic. Sci. Biotechnol. 2019, 94, 341–348. [Google Scholar] [CrossRef]
- Kim, S. A Codominant Molecular Marker in Linkage Disequilibrium with a Restorer-of-Fertility Gene (Ms) and Its Application in Reevaluation of Inheritance of Fertility Restoration in Onions. Mol. Breed. 2014, 34, 769–778. [Google Scholar] [CrossRef]
- De Vries, J.N.; Wietsma, W.A. Allium Roylei Stearn Restores Cytoplasmic Male Sterility of Rijnsburger Onion (A. cepa L.). J. Genet. Breed. 1992, 46, 379–382. [Google Scholar]
- Havey, M.J.; Kim, S. Molecular Marker Characterization of Commercially Used Cytoplasmic Male Sterilities in Onion. J. Am. Soc. Hortic. Sci. 2021, 146, 351–355. [Google Scholar] [CrossRef]
- Miller, R.J.; Koeppe, D.E. Southern Corn Leaf Blight: Susceptible and Resistant Mitochondria. Science 1971, 173, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Havey, M.J. Seed Yield, Floral Morphology, and Lack of Male-Fertility Restoration of Male-Sterile Onion (Allium cepa) Populations Possessing the Cytoplasm of Allium galanthum. J. Am. Soc. Hortic. Sci. 1999, 124, 626–629. [Google Scholar] [CrossRef]
- Vu, H.Q.; Iwata, M.; Yamauchi, N.; Shigyo, M. Production of Novel Alloplasmic Male Sterile Lines in Allium cepa Harbouring the Cytoplasm from Allium roylei. Plant Breed. 2011, 130, 469–475. [Google Scholar] [CrossRef]
- Gökçe, A.F.; Havey, M.J. Linkage Equilibrium among Tightly Linked RFLPs and the Ms Locus in Open-Pollinated Onion Populations. J. Am. Soc. Hortic. Sci. 2002, 127, 944–946. [Google Scholar] [CrossRef]
- Martin, W.J.; McCallum, J.; Shigyo, M.; Jakse, J.; Kuhl, J.C.; Yamane, N.; Pither-Joyce, M.; Gokce, A.F.; Sink, K.C.; Town, C.D.; et al. Genetic Mapping of Expressed Sequences in Onion and in Silico Comparisons with Rice Show Scant Colinearity. Mol. Genet. Genom. 2005, 274, 197–204. [Google Scholar] [CrossRef]
- Bang, H.; Cho, D.Y.; Yoo, K.S.; Yoon, M.K.; Patil, B.S.; Kim, S. Development of Simple PCR-Based Markers Linked to the Ms Locus, a Restorer-of-Fertility Gene in Onion (Allium cepa L.). Euphytica 2011, 179, 439–449. [Google Scholar] [CrossRef]
- Bang, H.; Kim, S.; Park, S.O.; Yoo, K.S.; Patil, B.S. Development of a Codominant CAPS Marker Linked to the Ms Locus Controlling Fertility Restoration in Onion (Allium cepa L.). Sci. Hortic. 2013, 153, 42–49. [Google Scholar] [CrossRef]
- Park, J.; Bang, H.; Cho, D.Y.; Yoon, M.K.; Patil, B.S.; Kim, S. Construction of High-Resolution Linkage Map of the Ms Locus, a Restorer-of-Fertility Gene in Onion (Allium cepa L.). Euphytica 2013, 192, 267–278. [Google Scholar] [CrossRef]
- Havey, M.J. Single Nucleotide Polymorphisms in Linkage Disequilibrium with the Male-Fertility Restoration (Ms) Locus in Open-Pollinated and Inbred Populations of Onion. J. Am. Soc. Hortic. Sci. 2013, 138, 306–309. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Huo, Y.M.; Miao, J.; Liu, B.J.; Kong, S.P.; Gao, L.M.; Liu, C.; Wang, Z.B.; Tahara, Y.; Kitano, H.; et al. Identification of Two SCAR Markers Co-Segregated with the Dominant Ms and Recessive Ms Alleles in Onion (Allium cepa L.). Euphytica 2013, 190, 267–277. [Google Scholar] [CrossRef]
- Huo, Y.M.; Liu, B.J.; Yang, Y.Y.; Miao, J.; Gao, L.M.; Kong, S.P.; Wang, Z.B.; Kitano, H.; Wu, X. AcSKP1, a Multiplex PCR-Based Co-Dominant Marker in Complete Linkage Disequilibrium with the Male-Fertility Restoration (Ms) Locus, and Its Application in Open-Pollinated Populations of Onion. Euphytica 2015, 204, 711–722. [Google Scholar] [CrossRef]
- Khar, A.; Saini, N. Limitations of PCR-Based Molecular Markers to Identify Male-Sterile and Maintainer Plants from Indian Onion (Allium cepa L.) Populations. Plant Breed. 2016, 135, 519–524. [Google Scholar] [CrossRef]
- Khrustaleva, L.; Jiang, J.; Havey, M.J. High-Resolution Tyramide-FISH Mapping of Markers Tightly Linked to the Male-Fertility Restoration (Ms) Locus of Onion. Theor. Appl. Genet. 2016, 129, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Havey, M.J.; von Kohn, C. Efficacy of Molecular Markers Jnurf13 and AcPms1 for Prediction of Genotypes at the Nuclear Ms Locus in Three Open-Pollinated Populations of Onion from North America. HortScience 2017, 52, 1052–1053. [Google Scholar] [CrossRef]
- Khar, A.; Zimik, M.; Verma, P.; Singh, H.; Mangal, M.; Singh, M.C.; Gupta, A.J. Molecular Marker-Based Characterization of Cytoplasm and Restorer of Male Sterility (Ms) Locus in Commercially Grown Onions in India. Mol. Biol. Rep. 2022, 49, 5535–5545. [Google Scholar] [CrossRef]
- Kim, T.; Kim, S. Identification of a Novel Haplotype of the Ms Locus Controlling Restoration of Male-Fertility and Its Implication in Origination of Cytoplasmic Male-Sterility in Onion (Allium cepa L.). J. Hortic. Sci. Biotechnol. 2021, 96, 750–758. [Google Scholar] [CrossRef]
- Yu, N.; Kim, S. Identification of Ms2, a Novel Locus Controlling Male-Fertility Restoration of Cytoplasmic Male-Sterility in Onion (Allium cepa L.), and Development of Tightly Linked Molecular Markers. Euphytica 2021, 217, 191. [Google Scholar] [CrossRef]
- Santos, C.A.; Leite, D.L.; Oliveira, V.R.; Rodrigues, M.A. Marker-Assisted Selection of Maintainer Lines within an Onion Tropical Population. Sci. Agric. 2010, 67, 223–227. [Google Scholar] [CrossRef]
- Ragassi, C.F.; Santos, M.D.M.; Fonseca, M.E.N.; Oliveira, V.R.; Buzar, A.G.R.; Costa, C.P.; Boiteux, L.S. Genotyping of Polymorphisms Associated with Male-Sterility Systems in Onion Accessions Adapted for Cultivation in Brazil. Hortic. Bras. 2012, 30, 409–414. [Google Scholar] [CrossRef]
- Saini, N.; Hedau, N.K.; Khar, A.; Yadav, S.; Bhatt, J.C.; Agrawal, P.K. Successful Deployment of Marker Assisted Selection (MAS) for Inbred and Hybrid Development in Long-Day Onion (Allium cepa L.). Indian J. Genet. Plant Breed. 2015, 75, 93–98. [Google Scholar] [CrossRef]
- Ferreira, R.R.; Santos, C.A.; Oliveira, V.R. Fertility Restoration Locus and Cytoplasm Types in Onion. Genet. Mol. Res. 2017, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.R.; Santos, C.A.F. Partial Success of Marker-Assisted Selection of ‘A’ and ‘B’ Onion Lines in Brazilian Germplasm. Sci. Hortic. 2018, 242, 110–115. [Google Scholar] [CrossRef]
- Manjunathagowda, D.C.; Anjanappa, M. Identification and Development of Male Sterile and Their Maintainer Lines in Short-Day Onion (Allium cepa L.) Genotypes. Genet. Resour. Crop Evol. 2020, 67, 357–365. [Google Scholar] [CrossRef]
- Gazendam, I.; Greyling, M.M.; Laurie, S.M. The Application of Molecular Markers to Accelerate the Recovery of Cytoplasmic and Nuclear Male Sterility in South African Onion (Allium cepa L.) Hybrid Parental Lines. J. Agric. Sci. 2018, 10, 95. [Google Scholar] [CrossRef]
- Yuan, Q.; Song, C.; Gao, L.; Zhang, H.; Yang, C.; Sheng, J.; Ren, J.; Chen, D.; Wang, Y. Transcriptome de Novo Assembly and Analysis of Differentially Expressed Genes Related to Cytoplasmic Male Sterility in Onion. Plant Physiol. Biochem. 2018, 125, 35–44. [Google Scholar] [CrossRef]
Table 2.
Genetic mapping of major diseases in onion.
| Disease | Linked Markers | Chromosome | Reference |
|---|---|---|---|
| Fusarium basal rot | isotig38484_281 | 2B | [156] |
| isotig44683_192 | 4A | ||
| isotig31106_505 | 4C | ||
| I30594_1021 | 8 | [157] | |
| i34519_442 | 6B | ||
| c00676_1004 | 1 | ||
| i33746_1093 | Unmapped | ||
| i33439_640 | Unmapped | ||
| NQ0345038 and NQ0257326 | 4 | [158] | |
| Pink root | isotig26045_1046 | 4 | [159] |
| NQ0257277 and NQ0258453 | 4 | [158] | |
| Purple blotch | matK and rbcL | 8 | [160] |
| AcSSR7 and ApR-450 | Unmapped | [161] | |
| ApRsnip14 and ApRsnip23 | Unmapped | [162] | |
| Downy mildew | DMR1 | 3 | [163] |
| Gray mold | SCAR-OPAN1 | Unmapped | [164] |
| SNP-3 HRM | Unmapped |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharma, S.; Khar, A.; Khosa, J.S.; Mandal, S.; Malla, S. Recent Advances in Molecular Genetics of Onion. Horticulturae 2024, 10, 256. https://doi.org/10.3390/horticulturae10030256
AMA Style
Sharma S, Khar A, Khosa JS, Mandal S, Malla S. Recent Advances in Molecular Genetics of Onion. Horticulturae. 2024; 10(3):256. https://doi.org/10.3390/horticulturae10030256
Chicago/Turabian StyleSharma, Suman, Anil Khar, Jiffinvir S. Khosa, Subhankar Mandal, and Subas Malla. 2024. "Recent Advances in Molecular Genetics of Onion" Horticulturae 10, no. 3: 256. https://doi.org/10.3390/horticulturae10030256
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.