

Evaluation of Squash (Cucurbita pepo L.) Genotypes for Resistance to Cucurbit Chlorotic Yellows Virus

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Seeds, Whiteflies, and Virus Culture
2.2. CCYV Transmissions
2.3. Disease Phenotyping
2.4. Quantification of CCYV
2.5. Statistical Analyses
3. Results
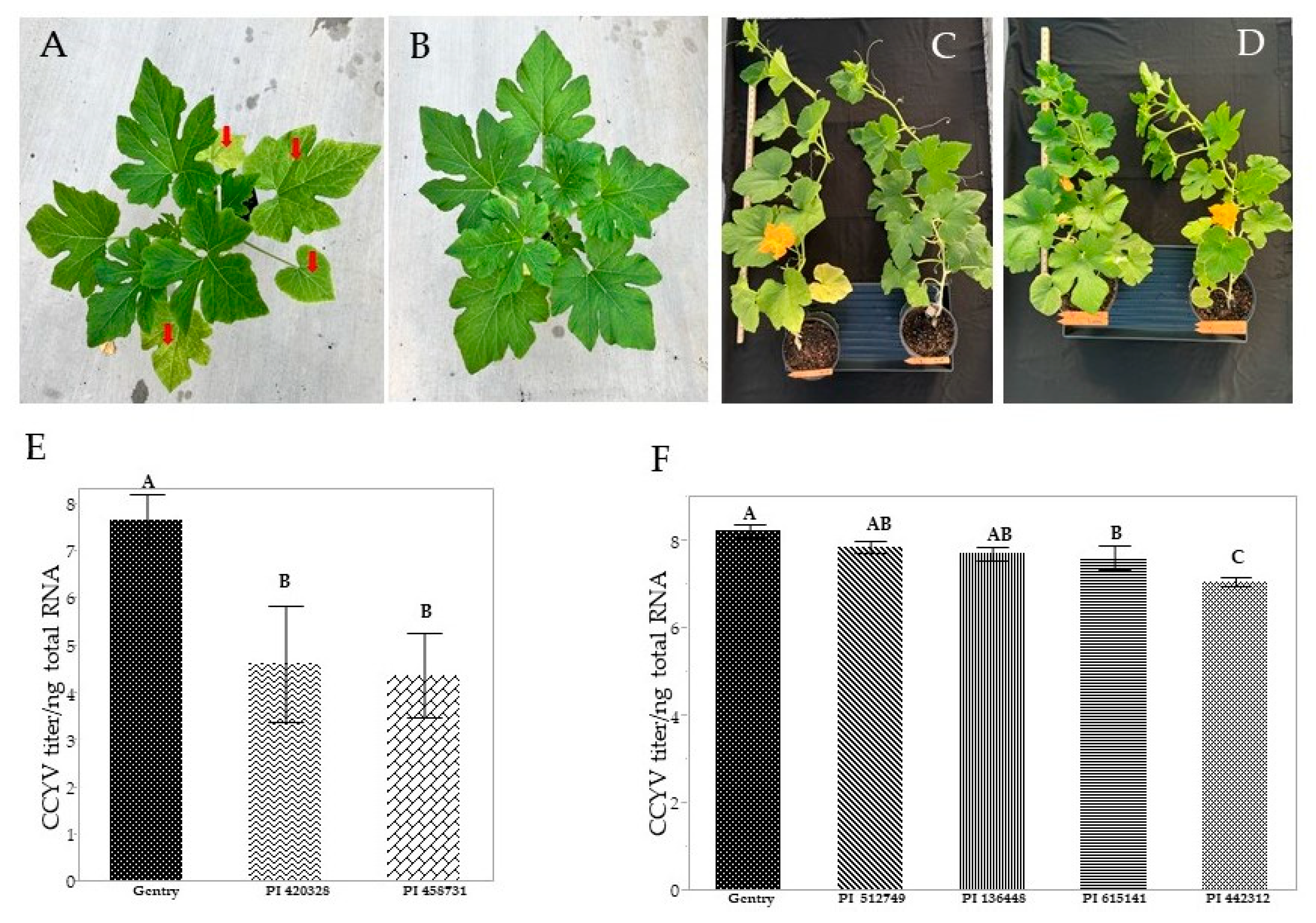
3.1. Symptom Severity
3.2. Virus Titer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wintermantel, W.M. Cucurbit Chlorotic Yellows Virus. 2022. Available online: https://ecucurbitviruses.org/ (accessed on 7 March 2024).
- Okuda, M.; Okazaki, S.; Yamasaki, S.; Okuda, S.; Sugiyama, M. Host range and complete genome sequence of Cucurbit chlorotic yellows virus, a new member of the genus Crinivirus. Phytopathology 2010, 100, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Gyoutoku, Y.; Okazaki, S.; Furuta, A.; Etoh, T.; Mizobe, M.; Kuno, K.; Hayashida, S.; Okuda, M. Chlorotic yellows disease of melon caused by cucurbit chlorotic yellows virus, a new crinivirus. Jpn. J. Phytopathol. 2009, 75, 109–111. [Google Scholar] [CrossRef]
- Bananej, K.; Menzel, W.; Kianfar, N.; Vahdat, A.; Winter, S. First report of cucurbit chlorotic yellows virus infecting cucumber, melon, and squash in Iran. Plant Dis. 2013, 97, 1005. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Rout, B.M.; Choudhary, S.; Sureja, A.K.; Baranwal, V.K.; Pant, R.P.; Kaur, B.; Jain, R.K.; Basavaraj, Y.B. First report of cucurbit chlorotic yellows virus infecting pumpkin in India. Plant Dis. 2021, 106, 1767. [Google Scholar] [CrossRef]
- Hamed, K.; Menzel, W.; Dafalla, G.; Gadelseed, A.M.A.; Winter, S. First report of cucurbit chlorotic yellows virus infecting muskmelon and cucumber in Sudan. Plant Dis. 2011, 95, 1321. [Google Scholar] [CrossRef] [PubMed]
- Abrahamian, P.E.; Sobh, H.; Abou-Jawdah, Y. First report of cucurbit chlorotic yellows virus on cucumber in Lebanon. Plant Dis. 2012, 96, 1704. [Google Scholar] [CrossRef] [PubMed]
- Orfanidou, C.; Maliogka, V.; Katis, N. First report of cucurbit chlorotic yellows virus in cucumber, melon, and watermelon in Greece. Plant Dis. 2014, 98, 1446. [Google Scholar] [CrossRef] [PubMed]
- Wintermantel, W.M.; Hladky, L.L.J.; Fashing, P.; Ando, K.; McCreight, J.D. First report of cucurbit chlorotic yellows virus infecting melon in the New World. Plant Dis. 2019, 103, 778. [Google Scholar] [CrossRef]
- Devendran, R.; Kavalappara, S.R.; Simmons, A.M.; Bag, S. Whitefly-transmitted viruses of cucurbits in the Southern United States. Viruses 2023, 15, 2278. [Google Scholar] [CrossRef]
- Peng, J.; Huang, Y. The occurrence of cucurbit chlorotic yellows virus disease in Taiwan and evaluation of the virus infected fruit quality and yield. Phytopathology 2011, 101, S139–S140. [Google Scholar] [CrossRef]
- UGA Farm Gate Value 2022. Available online: https://caed.uga.edu/publications/farm-gate-value.html (accessed on 7 March 2024).
- Kavalappara, S.R.; Milner, H.; Konakalla, N.C.; Morgan, K.; Sparks, A.N.; McGregor, C.; Culbreath, A.K.; Wintermantel, W.M.; Bag, S. High throughput sequencing-aided survey reveals widespread mixed infections of whitefly-transmitted viruses in cucurbits in Georgia, USA. Viruses 2021, 13, 988. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Friedman, M. Breeding for resistance to whitefly-transmitted geminiviruses. Ann. Appl. Biol. 2002, 140, 109–127. [Google Scholar] [CrossRef]
- Morales, F.J. Conventional breeding for resistance to Bemisia tabaci-transmitted geminiviruses. Crop Prot. 2001, 20, 825–834. [Google Scholar] [CrossRef]
- Candian, J.S.; Coolong, T.; Dutta, B.; Srinivasan, R.; Sparks, A.; Barman, A.; Ribeiro da Silva, A.L.B. Yellow squash and zucchini cultivar selection for resistance to cucurbit leaf crumple virus in the Southeastern United States. HortTechnology 2021, 31, 504–513. [Google Scholar] [CrossRef]
- Okuda, S.; Okuda, M.; Sugiyama, M.; Sakata, Y.; Takeshita, M.; Iwai, H. Resistance in melon to cucurbit chlorotic yellows virus, a whitefly-transmitted crinivirus. Eur. J. Plant Pathol. 2013, 135, 313–321. [Google Scholar] [CrossRef]
- Kawazu, Y.; Shimomura, K.; Maeda, S.; Yamato, Y.; Ueda, S.; Okuda, S.; Okuda, M.; Sugiyama, M. QTL mapping for resistance to cucurbit chlorotic yellows virus in melon (Cucumis melo L.). Euphytica 2018, 214, 239. [Google Scholar] [CrossRef]
- Luckew, A.; Meru, G.; Wang, Y.-Y.; Mwatuwa, R.; Paret, M.; Carvalho, R.; Kalischuk, M.; da Silva, A.L.B.R.; Candian, J.; Dutta, B. Field evaluation of cucurbita germplasm for resistance to whiteflies and whitefly-transmitted viruses. HortScience 2022, 57, 337–344. [Google Scholar] [CrossRef]
- Cohen, S.; Kern, J.; Harpaz, I.; Ben-Joseph, R. Epidemiological studies of the tomato yellow leaf curl virus (TYLCV) in the Jordan Valley, Israel. Phytoparasitica 1988, 16, 259–270. [Google Scholar] [CrossRef]
- Lapidot, M.; Ben-Joseph, R.; Cohen, L.; Machbash, Z.; Levy, D. Development of a scale for evaluation of tomato yellow leaf curl virus resistance level in tomato plants. Phytopathology 2006, 96, 1404–1408. [Google Scholar] [CrossRef]
- Costa, H.; Ullman, D.; Johnson, M.; Tabashnik, B. Squash silverleaf symptoms induced by immature, but not adult, Bemisia tabaci. Phytopathology 1993, 83, 763. Available online: https://www.apsnet.org/publications/phytopathology/backissues/Documents/1993Abstracts/Phyto_83_763.htm (accessed on 7 March 2024). [CrossRef]
- Tzanetakis, I.E.; Martin, R.R.; Wintermantel, W.M. Epidemiology of criniviruses: An emerging problem in world agriculture. Front. Microbiol. 2013, 4, 119. [Google Scholar] [CrossRef]
- Young, K.; Kabelka, E.A. Characterization of resistance to squash silverleaf disorder in summer squash. HortScience 2009, 44, 1213–1214. [Google Scholar] [CrossRef]
- Tamang, P.; Ando, K.; Wintermantel, W.M.; McCreight, J.D. QTL mapping of cucurbit yellow stunting disorder virus resistance in melon Accession PI 313970. HortScience 2021, 56, 424–430. [Google Scholar] [CrossRef]
- McCreight, J.D.; Wintermantel, W.M. Genetic resistance in melon PI 313970 to cucurbit yellow stunting disorder virus. HortScience 2011, 46, 1582–1587. [Google Scholar] [CrossRef]
- Orfanidou, C.; Katsiani, A.; Papayiannis, L.; Katis, N.I.; Maliogka, V.I. Interplay of cucurbit yellow stunting disorder virus with cucurbit chlorotic yellows virus and transmission dynamics by Bemisia tabaci MED. Plant Dis. 2021, 105, 416–424. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Riley, D.G.; Cremonez, P.S.G.; Perier, J.D.; Bag, S. Wild Radish (Raphanus raphanistrum L.) is a potential reservoir host of cucurbit chlorotic yellows virus. Viruses 2022, 14, 593. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-virus interactions in a plant host and in a hemipteran vector: Implications for vector fitness and virus epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Krishna Kumar, N.K.; Ullman, D.E.; Montero-Astúa, M.; Willis, D.K.; German, T.L.; Whitfield, A.E. Variation in tomato spotted wilt virus titer in Frankliniella occidentalis and its association with frequency of transmission. Phytopathology 2009, 99, 404–410. [Google Scholar] [CrossRef] [PubMed]
- McCreight, J.D.; Davis, A.A.; Reitsma, K. Melon (Cucumis melo) Crop Vulnerability Statement. 2020. Available online: https://www.ars-grin.gov/documents/cgc/cvs/2020-Melon%20Crop%20Vulnerability%20Statement.pdf (accessed on 7 March 2024).
- Pico, B.; Díez, M.; Nuez, F. Evaluation of whitefly-mediated inoculation techniques to screen Lycopersicon esculentum and wild relatives for resistance to tomato yellow leaf curl virus. Euphytica 1998, 101, 259–271. [Google Scholar] [CrossRef]
- Orfanidou, C.; Baltzi, A.; Dimou, N.; Katis, N.; Maliogka, V. Cucurbit chlorotic yellows virus: Insights into its natural host range, genetic variability, and transmission parameters. Plant Dis. 2017, 101, 2053–2058. [Google Scholar] [CrossRef]
- Akhter, M.S.; Bhor, S.A.; Hlalele, N.; Nao, M.; Sekine, K.-T.; Yaeno, T.; Yamaoka, N.; Nishiguchi, M.; Gubba, A.; Kobayashi, K. Review of beet pseudoyellows virus genome structure built the consensus genome organization of cucumber strains and highlighted the unique feature of strawberry strain. Virus Genes 2016, 52, 828–834. [Google Scholar] [CrossRef]
- Orílio, A.F.; Navas-Castillo, J. The complete nucleotide sequence of the RNA2 of the crinivirus tomato infectious chlorosis virus: Isolates from North America and Europe are essentially identical. Arch. Virol. 2009, 154, 683–687. [Google Scholar] [CrossRef]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef]
- Cooper, J.; Jones, A. Responses of plants to viruses: Proposals for the use of terms. Phytopathology 1983, 73, 127–128. Available online: https://www.apsnet.org/publications/phytopathology/backissues/Documents/1983Articles/Phyto73n02_127.PDF (accessed on 7 March 2024). [CrossRef]
- Fraser, R. The genetics of resistance to plant viruses. Annu. Rev. Phytopathol. 1990, 28, 179–200. [Google Scholar] [CrossRef]
- Kang, B.C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Annu. Rev Phytopathol 2005, 43, 581–621. [Google Scholar] [CrossRef] [PubMed]
- Pagán, I.; García-Arenal, F. Tolerance to plant pathogens: Theory and experimental evidence. Int. J. Mol. Sci. 2018, 19, 810. [Google Scholar] [CrossRef] [PubMed]
- Kavalappara, S.R.; Bag, S.; Luckew, A.; McGregor, C.E. Small RNA profiling of cucurbit yellow stunting disorder virus from susceptible and tolerant squash (Cucurbita pepo) lines. Viruses 2023, 15, 788. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Accession/Cultivar | Origin | Name | Symptom Severity Scores | |||
|---|---|---|---|---|---|---|
| Mass Exposure | Clip Cage Inoculations | |||||
| 15 DPI | 30 DPI | 15 DPIDPI | 30 DPI | |||
| Gentry | Seedway, USA | NA | 2 | 5 | 2 | 5 |
| PI 512749 | Castilla y León, Spain | AS-CU-1 | 0 | 3 | 0 | 3 |
| PI 615141 | Alma-Ata, Kazakhstan | Ames 19040 | 0 | 3 | 0 | 2 |
| PI 136448 | Manchuria, China | NA | 0 | 1 | 0 | 1 |
| PI 442312 | Guanajuato, Mexico | Calabaza de India | 0 | 1 | 0 | 1 |
| PI 458731 | Buenos Aires, Argentina | VAV 3738 | 0 | 1 | 0 | 1 |
| PI 420328 | Sakarya, Turkey | NA | 0 | 1 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavalappara, S.R.; Bag, S.; Luckew, A.; McGregor, C.E.; Culbreath, A.K.; Simmons, A.M. Evaluation of Squash (Cucurbita pepo L.) Genotypes for Resistance to Cucurbit Chlorotic Yellows Virus. Horticulturae 2024, 10, 264. https://doi.org/10.3390/horticulturae10030264
Kavalappara SR, Bag S, Luckew A, McGregor CE, Culbreath AK, Simmons AM. Evaluation of Squash (Cucurbita pepo L.) Genotypes for Resistance to Cucurbit Chlorotic Yellows Virus. Horticulturae. 2024; 10(3):264. https://doi.org/10.3390/horticulturae10030264
Chicago/Turabian StyleKavalappara, Saritha Raman, Sudeep Bag, Alexander Luckew, Cecilia E. McGregor, Albert K. Culbreath, and Alvin M. Simmons. 2024. "Evaluation of Squash (Cucurbita pepo L.) Genotypes for Resistance to Cucurbit Chlorotic Yellows Virus" Horticulturae 10, no. 3: 264. https://doi.org/10.3390/horticulturae10030264