
Cytogenetics Study of Four Edible and Ornamental Zingiber Species (Zingiberaceae) from Thailand
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Chromosome Number and Karyotype Study
2.3. Statistical Analysis
3. Results
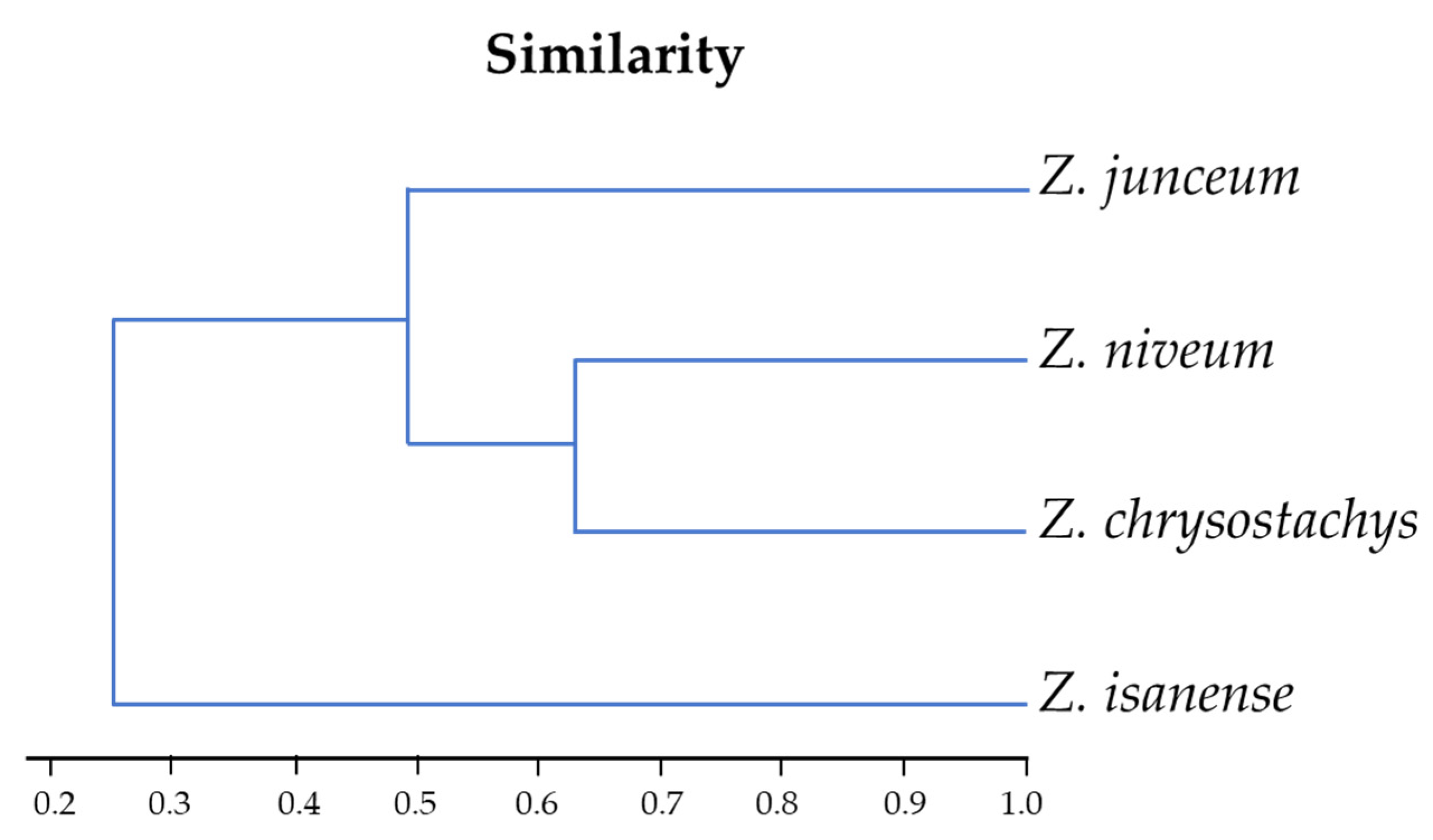
3.1. The Similarity Index of Four Rare Zingiber Species
3.2. The PCA Score Plot of CI
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plant of the World Online, Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 12 February 2023).
- Larsen, K.; Larsen, S.S. Ginger of Thailand; Queen Sirikit Botanic Garden, The Botanical Garden Organization: Chiang Mai, Thailand, 2006. [Google Scholar]
- Kress, W.J.; Prince, L.M.; Williams, J.K. The phylogeny and a new classifcation of the gingers (Zingiberaceae): Evidence from molecular data. Am. J. Bot. 2002, 89, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Mood, J.D.; Ardiyani, M.; Veldkamp, J.F.; Mandáková, T.; Prince, L.M.; de Boer, H.J. Nomenclatural changes in Zingiberaceae: Haplochorema is reduced to Boesenbergia. Gard. Bull. Singap. 2020, 72, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.M.; Tanaka, N.; Miyake, N. Two gingers, Zingiber orbiculatum and Z. flavomaculosum (Zingiberaceae), newly recorded from Myanmar. Bull. Natl. Mus. Nat. Sci. 2015, 41, 107–112. [Google Scholar]
- Saensouk, S.; Saensouk, P.; Pasorn, P.; Chantaranothai, P. Diversity, Traditional Uses and New Record of Zingiberaceae in Nam Nao National Park, Petchabun Province, Thailand. Agric Nat. Resour. 2016, 50, 445–453. [Google Scholar]
- Triboun, T.; Chantaranothai, P.; Larsen, K. Taxonomic changes regarding three species of Zingiber (Zingiberaceae) from Thailand. Acta Phytotax. Sin. 2007, 45, 403–404. [Google Scholar]
- Ikeda, H.; Nam, B.M.; Yamamoto, N.; Funakoshi, H.; Takano, A.; Im, H.T. Chromosome number of myoga ginger (Zingiber mioga: Zingiberaceae) in Korea. Korean J. Plant Taxon. 2021, 51, 100–102. [Google Scholar] [CrossRef]
- Khumkratok, S.; Boontiang, K.; Chutichudet, P.; Pramaul, P. Geographical distributions and ecology of ornamental Curcuma (Zingiberaceae) in Northeastern Thailand. Pak. J. Biol. Sci. 2012, 15, 929–939. [Google Scholar] [CrossRef]
- Saensouk, P.; Saensouk, S.; Phechphakdee, T.; Ragsasilp, A. Cytogenetic study in seven species of Zingiberaceae family from Bueng Kan Province, Thailand. Biodiversitas 2023, 24, 68–77. [Google Scholar] [CrossRef]
- Chen, Z.Y. Evolutionary patterns in cytology and pollen structure of Asian Zingiberaceae. In Tropical Forests; Holm-Nielsen, B., Nielsen, I.C., Balslev, H., Eds.; Academic Press Limited: New York, NY, USA, 1989; pp. 185–191. [Google Scholar]
- Lin, Y.-C.; Chao, C.-T.; Chang, C.-Y.; Tseng, Y.-H. Taxonomic revision of Zingiber (Zingiberaceae) of Taiwan. Eur. J. Taxon. 2022, 839, 74–102. [Google Scholar] [CrossRef]
- Eksomtramage, L.; Boontum, K. Chromosome counts of Zingiberaceae. Songklanakarin J. Sci. Technol. 1995, 17, 291–297. [Google Scholar]
- Eksomtramage, L.; Sirirugsa, P.; Sawangchote, P.; Jornead, S.; Saknimit, T.; Leeratiwong, C. Chromosome numbers of some monocot species from Ton-Nga-Chang Wildlife Sanctuary, Southern Thailand. Thai For. Bull. (Bot.) 2001, 29, 63–71. [Google Scholar]
- Eksomtramage, L.; Sirirugsa, P.; Jivanit, P.; Maknoi, C. Chromosome counts of some Zingiberaceous species from Thailand. Songklanakarin J. Sci. Technol. 2002, 24, 311–319. [Google Scholar]
- Ragsasilp, A.; Saensouk, P.; Saensouk, S. Ginger family from Bueng Kan Province, Thailand: Diversity, conservation status, and traditional uses. Biodiversitas 2022, 23, 2739–2752. [Google Scholar] [CrossRef]
- Triboun, P.; Larsen, K.; Chantaranothai, P. A key to the genus Zingiber (Zingiberaceae) in Thailand with descriptions of 10 new taxa. Thai J. Bot. 2003, 6, 53–77. [Google Scholar]
- Deng, M.; Yun, X.; Ren, S.; Qing, Z.; Luo, F. Plants of the Genus Zingiber: A review of their ethnomedicine, phytochemistry and pharmacology. Molecules 2022, 27, 2826. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species; Version 2022-2; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2023. [Google Scholar]
- Chaiyasut, K. Cytogenetics and Cytotaxonomy of the Family Zephyranthes; Department of Botany, Faculty of Science, Chulalongkorn University: Bangkok, Thailand, 1989. [Google Scholar]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Addison-Wesley Pub. Co.: San Francisco, CA, USA, 1971. [Google Scholar]
- Beltran, I.C.; Kiew, K.Y. Cytotaxonomic studies in the Zingiberaceae. Notes R. Bot. Gard. Edinb. 1984, 41, 541–559. [Google Scholar]
- Saensouk, S.; Chantaranothai, P. The Family Zingiberaceae in Phu Phan National Park. In Proceedings of the 3rd Symposium on the Family Zingiberaceae, Khon Kaen, Thailand, 7–12 July 2002; pp. 16–25. [Google Scholar]
- Saenprom, K.; Saensouk, S.; Saensouk, P.; Senakun, C. Karyomorphological analysis of four species of Zingiberaceae from Thailand. Nucleus 2018, 61, 111–120. [Google Scholar] [CrossRef]
- Saensouk, S.; Saensouk, P. New report on karyotype and ideogram of two Zingiber species, Z. ligulatum and Z. parishii subsp. phuphanense from Thailand. Nucleus 2021, 64, 115–121. [Google Scholar] [CrossRef]
- Bhadra, S.; Bandyopadhyay, M. New chromosome number counts and karyotype analyses in three important genera of Zingiberaceae. Nucleus 2016, 59, 35–40. [Google Scholar] [CrossRef]
- Levan, A.; Fredya, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosome. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Morinaga, T.; Fukushina, E.; Kanui, T.; Tamasaki, Y. Chromosome numbers of cultivated plants. Bot. Mag. 1929, 43, 589–594. [Google Scholar] [CrossRef]
- Saensouk, S.; Saensouk, P. Chromosome number of some Zingiberaceous in Thailand. KKU Res. J. 2004, 9, 3–9. [Google Scholar]
- Hong, D.Y.; Zhang, S.Z. Observations on chromosomes of some plants from western Sichuan. Cathaya 1990, 2, 191–197. [Google Scholar]
- Das, A.B.; Rai, S.; Das, P. Estimation of 4C DNA and karyotype analysis in ginger (Zingiber officinale Rosc.) II. Cytologia 1998, 63, 133–139. [Google Scholar] [CrossRef]
- Senavongse, R.; Saensouk, S.; Saensouk, P. Karyological study in three native species of genus Alocasia (Araceae) in the northeast of Thailand. Nucleus 2020, 63, 81–85. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Sengthong, A.; Saensouk, S.; Saensouk, P.; Souladeth, P. Cytogenetic Study of Five Varieties of Callisia repens (Jacq.) L. (Commelinaceae) from Laos. Horticulturae 2023, 9, 1050. [Google Scholar] [CrossRef]
- Bro, R.; Smilde, A.K. Principal component analysis. Anal. Methods 2014, 6, 2812–2831. [Google Scholar] [CrossRef]
- Santhosh, B. Cytological and Palynological Studies on the Family Apocynaceae; Department of Botany, University of Kerala: Thiruvananthapuram, India, 1999. [Google Scholar]
- Dobigny, G.; Ducroz, J.F.; Robinson, T.J.; Volobouev, V. Cytogenetics and Cladistics. Syst. Biol. 2004, 53, 470–484. [Google Scholar] [CrossRef]
- Guerra, M. Chromosome numbers in plant taxonomy: Concepts and implications. Cytogenet. Genome Res. 2008, 120, 339–350. [Google Scholar] [CrossRef]
- Dobigny, G.; Aniskin, V.; Volobouev, V. Explosive chromosome evolution and speciation in the gerbil genus Taterillus (Rodentia, Gerbillinae): A case of two new cryptic species. Cytogenet. Cell Genet. 2002, 96, 117–124. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | n | 2n | NF | Karyotype Formula (Symmetry) | Conservation Status (Based on WCSP/IUCN 2022) | Dominant Characteristics | Traditional Uses | Location (Provinces) | References |
|---|---|---|---|---|---|---|---|---|---|
| Zingiber chrysostachys | - | 22 | 44 * | 16m + 6sm * (Symmetry karyotype) | Rare species/ Endangered | yellow bracts, pale yellow flowers, and a red labellum with white dots |
- Young inflorescences and young leaves are used as vegetables or local food. - Ornamental plants | Thailand (Nakhon Phanom) | Present study |
| 11 | - | - | - | - | Malaysia | [22] | |||
| - | 22 | - | - | - | Thailand (Sakon Nakhon) | [23] | |||
| Z. isanense | - | 22 * | 44 * | 2m + 20sm * (Symmetry karyotype) | Rare species/ Least Concern | red-brown bracts, white flowers, and a white labellum |
- Young leaves are used as vegetables or local food. - Ornamental plants | Thailand (Ubon ratchathani) | Present study * |
| Z. junceum | - | 22 | 44 * | 12m + 10sm * (Symmetry karyotype) | Rare species/ Least Concern | green bracts, yellow flowers, and a yellow labellum |
- Young inflorescences, young leaves, and young rhizomes are used as vegetables or local food. - Ornamental plants | Thailand (Bueng Kan) | Present study |
| - | 22 | - | - | - | Thailand (Sakon Nakhon) | [23] | |||
| Z. niveum | - | 22 * | 44 * | 14m + 4sm + 4st * (Asymmetry karyotype) | Rare species/ Endangered | white bracts, yellow flowers, and a yellow labellum |
- Young inflorescences, young leaves, and young rhizomes are used as vegetables or local food. - Ornamental plants | Thailand (Ubon ratchathani) | Present study * |
| Zingiber chrysostachys (2n = 22) | ||||||
| Chro. Pair | Ls ± SD (µm) | Ll ± SD (µm) | LT ± SD (µm) | RL (%) | CI | Chromosome Type |
| 1. | 1.40 ± 0.09 | 2.17 ± 0.65 | 3.57 ± 0.73 | 12.72 ± 0.04 | 0.61 ± 0.08 | Submetacentric |
| 2. | 1.09 ± 0.07 | 1.92 ± 0.13 | 3.02 ± 0.21 | 10.75 ± 0.06 | 0.64 ± 0.10 | Submetacentric |
| 3. | 1.20 ± 0.08 | 1.81 ± 0.12 | 3.01 ± 0.20 | 10.74 ± 0.08 | 0.60 ± 0.06 | Submetacentric |
| 4. | 1.18 ± 0.07 | 1.61 ± 0.12 | 2.78 ± 0.18 | 9.92 ± 0.06 | 0.58 ± 0.04 | Metacentric |
| 5. | 1.10 ± 0.06 | 1.49 ± 0.10 | 2.59 ± 0.16 | 9.24 ± 0.04 | 0.57 ± 0.08 | Metacentric |
| 6. | 1.15 ± 0.07 | 1.36 ± 0.09 | 2.51 ± 0.16 | 8.94 ± 0.08 | 0.54 ± 0.06 | Metacentric |
| 7. | 1.03 ± 0.06 | 1.40 ± 0.10 | 2.43 ± 0.16 | 8.68 ± 0.08 | 0.58 ± 0.04 | Metacentric |
| 8. | 1.00 ± 0.06 | 1.36 ± 0.08 | 2.36 ± 0.14 | 8.41 ± 0.04 | 0.58 ± 0.04 | Metacentric |
| 9. | 0.91 ± 0.06 | 1.18 ± 0.08 | 2.08 ± 0.14 | 7.43 ± 0.04 | 0.56 ± 0.06 | Metacentric |
| 10. | 0.91 ± 0.06 | 1.13 ± 0.08 | 2.04 ± 0.13 | 7.27 ± 0.08 | 0.56 ± 0.06 | Metacentric |
| 11. | 0.68 ± 0.04 | 0.98 ± 0.06 | 1.66 ± 0.11 | 5.91 ± 0.04 | 0.59 ± 0.08 | Metacentric |
| Zingiber isanensis (2n = 22) | ||||||
| Chro. Pair | Ls ± SD (µm) | Ll ± SD (µm) | LT ± SD (µm) | RL (%) | CI | Chromosome Type |
| 1. | 1.37 ± 0.05 | 2.45 ± 0.01 | 3.83 ± 0.06 | 13.00 ± 0.02 | 0.64 ± 0.01 | Submetacentric |
| 2. | 1.29 ± 0.02 | 2.22 ± 0.01 | 3.51 ± 0.02 | 12.00 ± 0.01 | 0.63 ± 0.01 | Submetacentric |
| 3. | 1.12 ± 0.02 | 2.03 ± 0.01 | 3.15 ± 0.03 | 11.00 ± 0.01 | 0.64 ± 0.01 | Submetacentric |
| 4. | 1.07 ± 0.03 | 1.96 ± 0.01 | 3.03 ± 0.04 | 10.00 ± 0.01 | 0.65 ± 0.01 | Submetacentric |
| 5. | 1.04 ± 0.03 | 1.58 ± 0.01 | 2.62 ± 0.03 | 9.00 ± 0.01 | 0.59 ± 0.01 | Metacentric |
| 6. | 0.98 ± 0.01 | 1.54 ± 0.01 | 2.52 ± 0.01 | 9.00 ± 0.01 | 0.61 ± 0.01 | Submetacentric |
| 7. | 0.94 ± 0.02 | 1.50 ± 0.01 | 2.44 ± 0.02 | 8.00 ± 0.01 | 0.61 ± 0.01 | Submetacentric |
| 8. | 0.83 ± 0.01 | 1.44 ± 0.01 | 2.26 ± 0.01 | 8.00 ± 0.01 | 0.64 ± 0.01 | Metacentric |
| 9. | 0.77 ± 0.01 | 1.39 ± 0.01 | 2.16 ± 0.01 | 7.00 ± 0.01 | 0.64 ± 0.01 | Submetacentric |
| 10. | 0.75 ± 0.01 | 1.28 ± 0.01 | 2.04 ± 0.01 | 7.00 ± 0.01 | 0.63 ± 0.01 | Submetacentric |
| 11. | 0.68 ± 0.01 | 1.08 ± 0.01 | 1.76 ± 0.01 | 6.00 ± 0.01 | 0.61 ± 0.01 | Submetacentric |
| Zingiber junceum (2n = 22) | ||||||
| Chro. Pair | Ls ± SD (µm) | Ll ± SD (µm) | LT ± SD (µm) | RL (%) | CI | Chromosome Type |
| 1. | 1.24 ± 0.08 | 2.22 ± 0.65 | 3.46 ± 0.73 | 12.00 ± 0.16 | 0.64 ± 0.13 | Submetacentric |
| 2. | 1.27 ± 0.07 | 1.83 ± 0.13 | 3.10 ± 0.20 | 10.76 ± 0.15 | 0.59 ± 0.12 | Metacentric |
| 3. | 1.06 ± 0.07 | 1.95 ± 0.13 | 3.01 ± 0.19 | 10.45 ± 0.13 | 0.65 ± 0.12 | Submetacentric |
| 4. | 1.00 ± 0.06 | 1.76 ± 0.12 | 2.76 ± 0.18 | 9.58 ± 0.14 | 0.64 ± 0.13 | Submetacentric |
| 5. | 1.12 ± 0.06 | 1.54 ± 0.10 | 2.66 ± 0.16 | 9.21 ± 0.13 | 0.58 ± 0.12 | Metacentric |
| 6. | 1.14 ± 0.07 | 1.44 ± 0.09 | 2.58 ± 0.16 | 8.95 ± 0.12 | 0.56 ± 0.12 | Metacentric |
| 7. | 1.13 ± 0.06 | 1.40 ± 0.10 | 2.54 ± 0.16 | 8.80 ± 0.12 | 0.55 ± 0.10 | Metacentric |
| 8. | 1.00 ± 0.06 | 1.45 ± 0.09 | 2.46 ± 0.14 | 8.51 ± 0.10 | 0.59 ± 0.10 | Metacentric |
| 9. | 0.93 ± 0.05 | 1.45 ± 0.09 | 2.38 ± 0.14 | 8.26 ± 0.10 | 0.61 ± 0.09 | Submetacentric |
| 10. | 0.89 ± 0.05 | 1.39 ± 0.08 | 2.28 ± 0.13 | 7.92 ± 0.09 | 0.61 ± 0.09 | Submetacentric |
| 11. | 0.66 ± 0.04 | 0.95 ± 0.06 | 1.61 ± 0.10 | 5.57 ± 0.08 | 0.59 ± 0.10 | Metacentric |
| Zingiber niveum (2n = 22) | ||||||
| Chro. Pair | Ls ± SD (µm) | Ll ± SD (µm) | LT ± SD (µm) | RL (%) | CI | Chromosome Type |
| 1. | 1.65 ± 0.10 | 2.33 ± 0.68 | 3.98 ± 0.78 | 12.50 ± 0.20 | 0.58 ± 0.10 | Metacentric |
| 2. | 1.61 ± 0.09 | 2.24 ± 0.15 | 3.85 ± 0.24 | 12.08 ± 0.12 | 0.58 ± 0.10 | Metacentric |
| 3. | 1.47 ± 0.09 | 2.25 ± 0.14 | 3.72 ± 0.23 | 11.66 ± 0.09 | 0.61 ± 0.12 | Subtalocentric |
| 4. | 1.19 ± 0.08 | 1.88 ± 0.13 | 3.07 ± 0.21 | 9.63 ± 0.12 | 0.61 ± 0.12 | Subtalocentric |
| 5. | 1.09 ± 0.07 | 1.67 ± 0.11 | 2.76 ± 0.18 | 8.68 ± 0.09 | 0.60 ± 0.10 | Submetacentric |
| 6. | 1.29 ± 0.08 | 1.47 ± 0.10 | 2.76 ± 0.18 | 8.66 ± 0.10 | 0.53 ± 0.09 | Metacentric |
| 7. | 1.05 ± 0.07 | 1.63 ± 0.11 | 2.68 ± 0.18 | 8.42 ± 0.09 | 0.61 ± 0.12 | Submetacentric |
| 8. | 1.06 ± 0.06 | 1.51 ± 0.09 | 2.56 ± 0.16 | 8.05 ± 0.08 | 0.59 ± 0.09 | Metacentric |
| 9. | 1.03 ± 0.07 | 1.22 ± 0.09 | 2.25 ± 0.15 | 7.06 ± 0.06 | 0.54 ± 0.08 | Metacentric |
| 10. | 1.02 ± 0.06 | 1.26 ± 0.08 | 2.28 ± 0.15 | 7.16 ± 0.09 | 0.55 ± 0.09 | Metacentric |
| 11. | 0.86 ± 0.06 | 1.09 ± 0.07 | 1.94 ± 0.13 | 6.10 ± 0.12 | 0.56 ± 0.010 | Metacentric |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saensouk, P.; Saensouk, S.; Senavongse, R.; Maensiri, D.; Souladeth, P. Cytogenetics Study of Four Edible and Ornamental Zingiber Species (Zingiberaceae) from Thailand. Horticulturae 2024, 10, 409. https://doi.org/10.3390/horticulturae10040409
Saensouk P, Saensouk S, Senavongse R, Maensiri D, Souladeth P. Cytogenetics Study of Four Edible and Ornamental Zingiber Species (Zingiberaceae) from Thailand. Horticulturae. 2024; 10(4):409. https://doi.org/10.3390/horticulturae10040409
Chicago/Turabian StyleSaensouk, Piyaporn, Surapon Saensouk, Rattanavalee Senavongse, Duangkamol Maensiri, and Phetlasy Souladeth. 2024. "Cytogenetics Study of Four Edible and Ornamental Zingiber Species (Zingiberaceae) from Thailand" Horticulturae 10, no. 4: 409. https://doi.org/10.3390/horticulturae10040409