Do Extended Cultivation Periods and Reduced Nitrogen Supply Increase Root Yield and Anthocyanin Content of Purple Carrots?

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation
2.2. Yield Determination, Sampling, and Chemical Analyses
2.3. Statistical Analyses
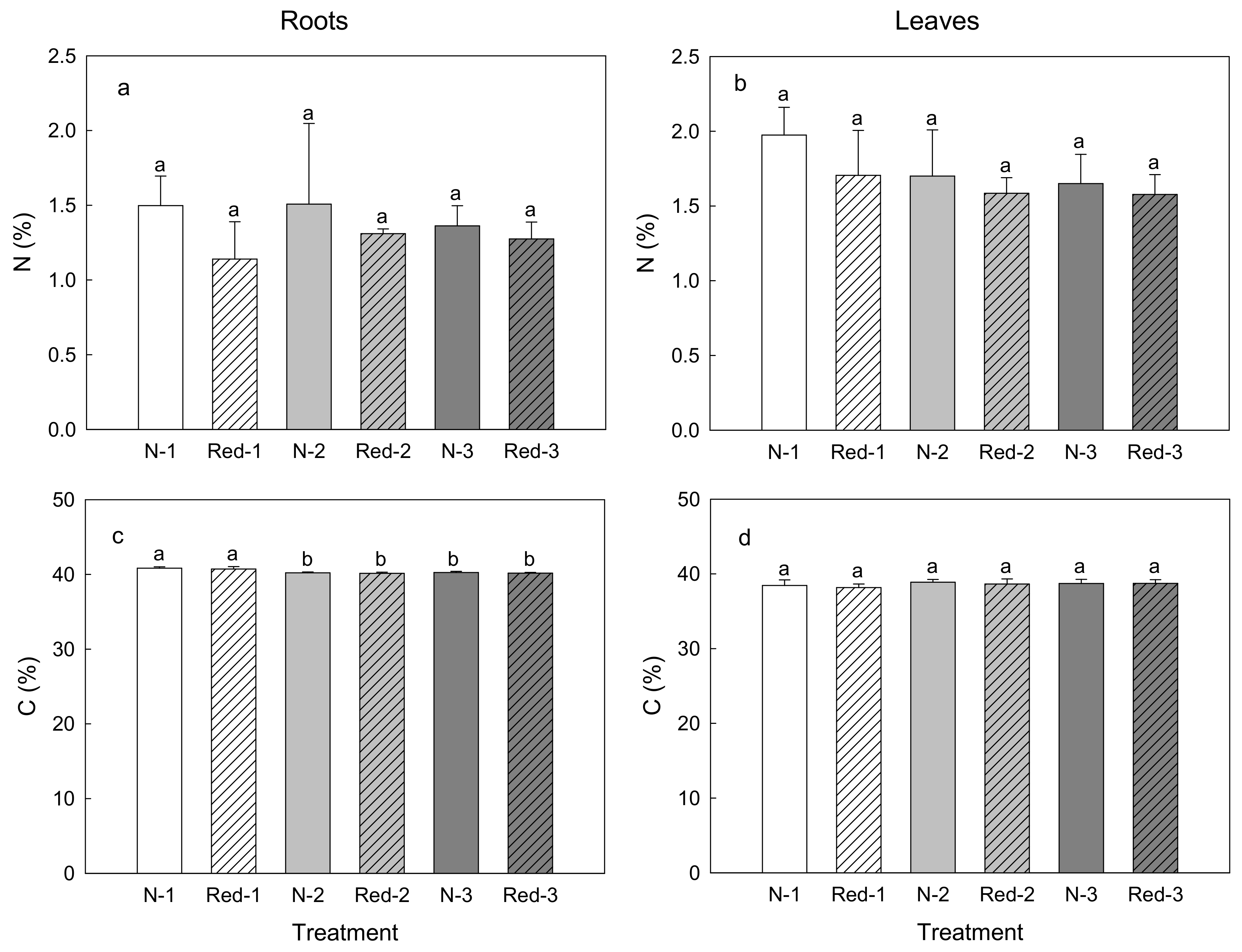
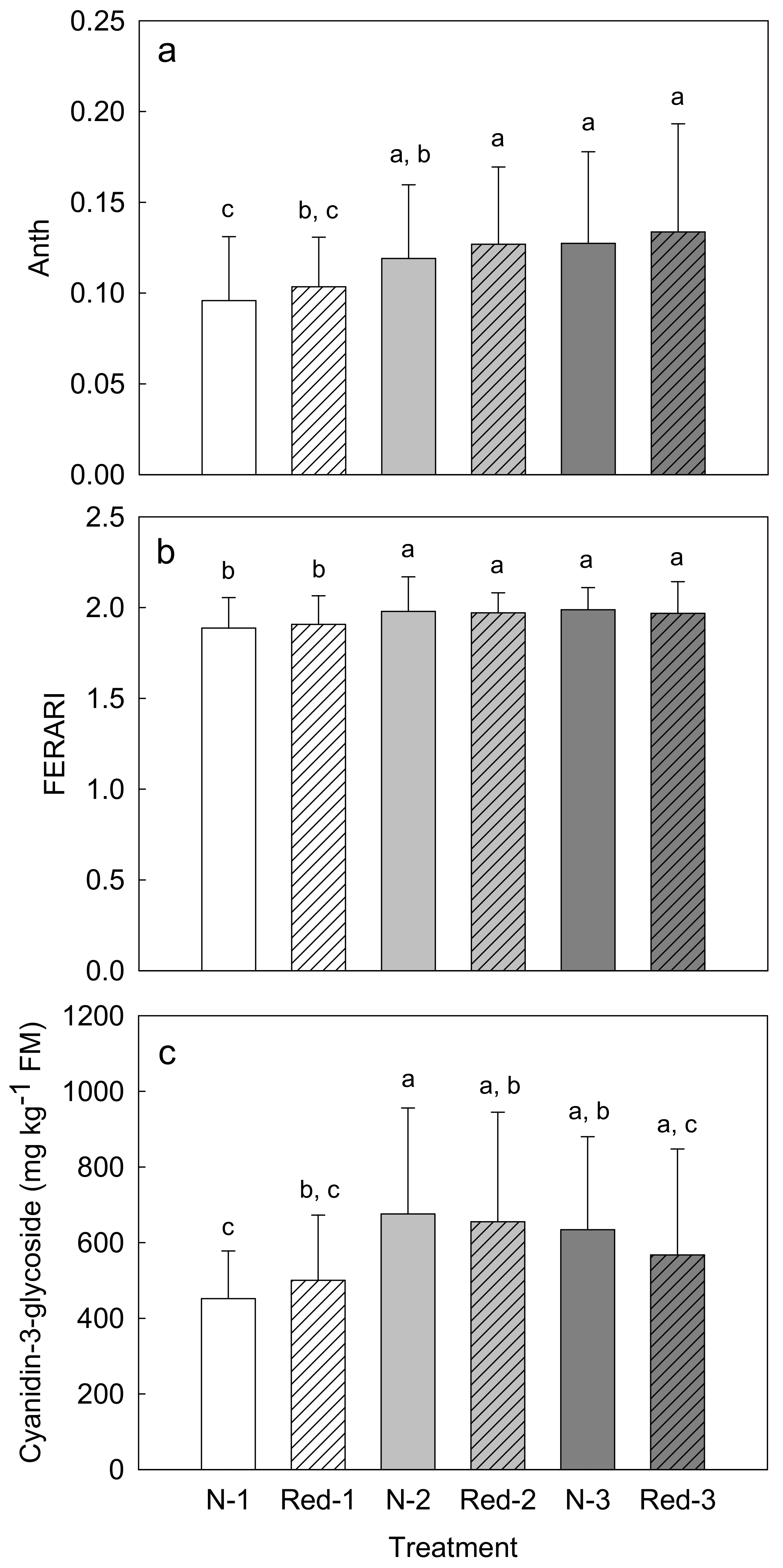
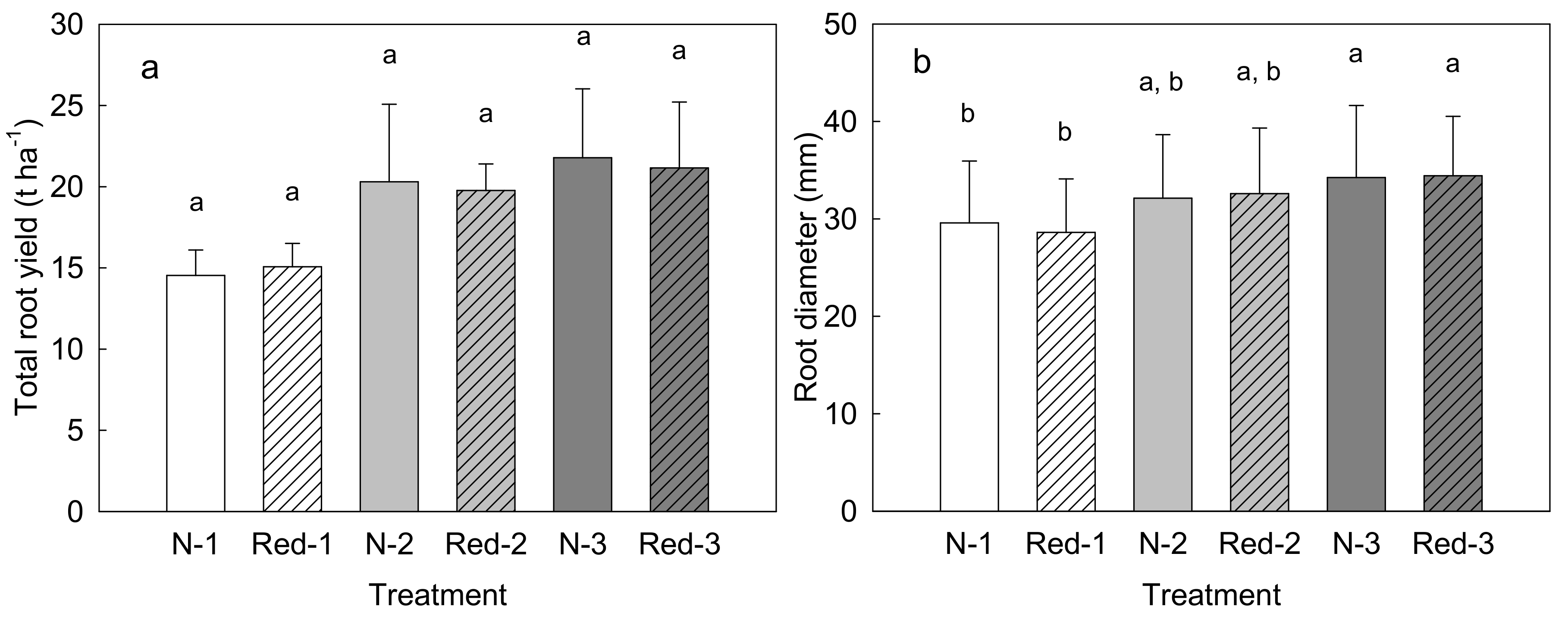
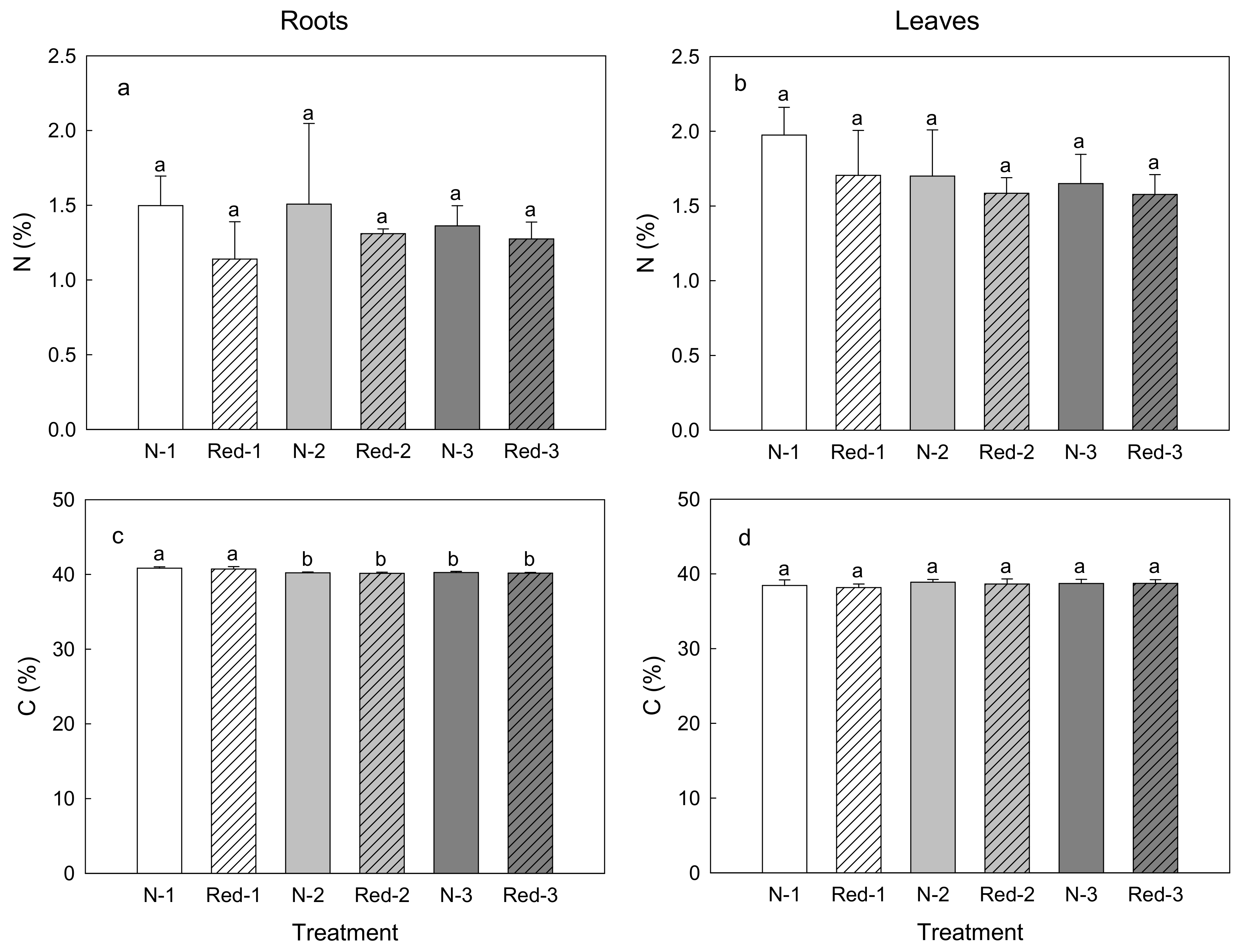
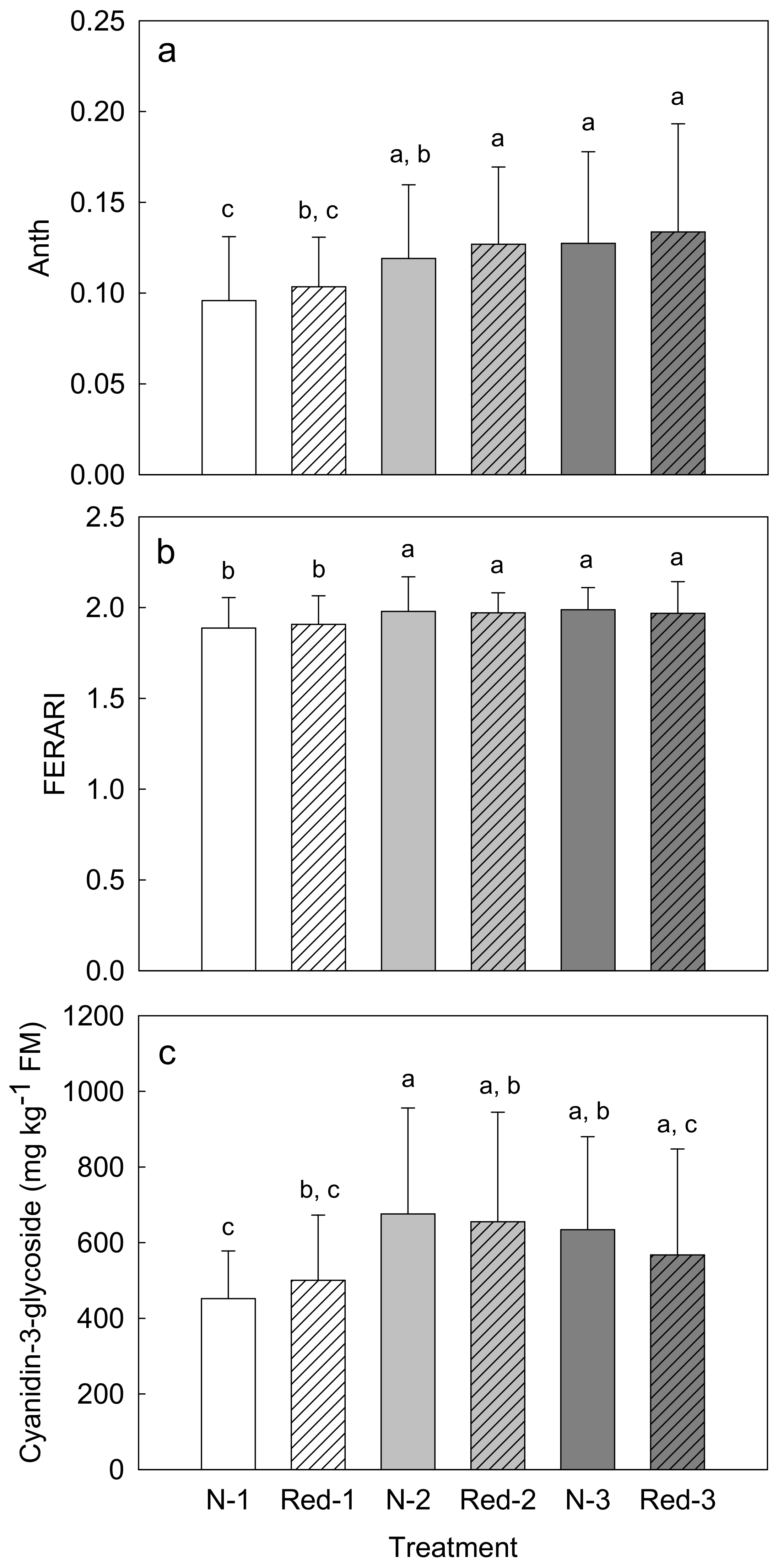
3. Results
4. Discussion
4.1. Reduced N Supply Has Little Effects on Purple Carrot Yield and Composition
4.2. Later Harvests Do Not Increase the Yield but Alter the Root Composition
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photchem. Photbiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Gould, K.S. Nature’s Swiss army knife: The diverse protective roles of anthocyanins in leaves. J. Biomed. Biotechnol. 2004, 5, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.M.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Brouillard, R. Analysis and biological activities of anthocyanins. Phytochemistry 2003, 64, 923–933. [Google Scholar] [CrossRef]
- He, J.; Giusti, M.M. Anthocyanins. Natural colorants with health-promoting properties. Ann. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Roriz, C.L.; Morales, P.; Barros, L.; Ferreira, I.C.F.R. Food colorants. Challenges, opportunities and current desires of agro-industries to ensure consumer expectations and regulatory practices. Trends Food Sci. Technol. 2016, 52, 1–15. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.D.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins. A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Kämmerer, D.; Carle, R.; Schieber, A. Quantification of anthocyanins in black carrot extracts (Daucus carota ssp. sativus var. atrorubens Alef.) and evaluation of their color properties. Eur. Food Res. Technol. 2004, 219, 479–486. [Google Scholar] [CrossRef]
- Seljasen, R.; Kristensen, H.L.; Lauridsen, C.; Wyss, G.S.; Kretzschmar, U.; Birlouez-Aragone, I.; Kahl, J. Quality of carrots as affected by pre- and postharvest factors and processing. J. Sci. Food Agric. 2013, 93, 2611–2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschold, P.J.; Hey, M.; Kleber, J.; Kürbel, P. Violette Möhrensorten—Was ist anders? Monatsschrift Mag. Gart.-Profi (Sonderh. Möhren) 2008, 12, 14–17. [Google Scholar]
- Li, H.Y.; Deng, Z.Y.; Zhu, H.H.; Hu, C.L.; Liu, R.H.; Young, J.C.; Tsao, R. Highly pigmented vegetables. Anthocyanin compositions and their role in antioxidant activities. Food Res. Int. 2012, 46, 250–259. [Google Scholar] [CrossRef]
- Montilla, E.C.; Arzaba, M.R.; Hillebrand, S.; Winterhalter, P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) cultivars Antonina, Beta Sweet, Deep Purple, and Purple Haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Close, D.C.; Beadle, C.L. The ecophysiology of foliar anthocyanin. Bot. Rev. 2003, 69, 149–161. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Liu, Z.J.; Liu, J.P.; Lin, S.; Wang, J.F.; Lin, W.X.; Xu, W.F. GA-DELLA pathway is involved in regulation of nitrogen deficiency-induced anthocyanin accumulation. Plant Cell Rep. 2017, 36, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Caretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon fluxes between primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [PubMed]
- Schaller, R.G.; Schnitzler, W.H. Nitrogen nutrition and flavour compounds of carrots (Daucus carota L.) cultivated in Mitscherlich pots. J. Sci. Food Agric. 2000, 80, 49–56. [Google Scholar] [CrossRef]
- Feller, C.; Fink, M.; Laber, H.; Maync, A.; Paschold, P.; Scharpf, H.C.; Schlaghecken, J.; Strohmeyer, K.; Weier, U.; Ziegler, J. Düngung im Freilandgemüsebau, 3rd ed.; Schriftenreihe des Leibniz-Instituts für Gemüse- und Zierpflanzenbau (IGZ): Großbeeren, Germany, 2011; Available online: http://www.igzev.de/publikationen/IGZ_Duengung_im_Freilandgemuesebau.pdf (accessed on 11 August 2017).
- Schmidt, L.; Zinkernagel, J. Yield and anthocyanin content in purple carrots with reduced nitrogen supply. DGG Proc. 2014, 4. [Google Scholar] [CrossRef]
- Hamilton, J.G.; Zangerl, A.R.; DeLucia, E.H.; Berenbaum, M.R. The carbon-nutrient balance hypothesis. Its rise and fall. Ecol. Lett. 2001, 4, 86–95. [Google Scholar] [CrossRef]
- Bumgarner, N.R.; Scheerens, J.C.; Mullen, R.W.; Bennett, M.A.; Ling, P.P.; Kleinhenz, M.D. Root-zone temperature and nitrogen affect the yield and secondary metabolite concentration of fall- and spring-grown, high-density leaf lettuce. J. Sci. Food Agric. 2012, 92, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Warnasooriya, S.N.; Montgomery, B.L. Mesophyll-localized phytochromes gate stress- and light-inducible anthocyanin accumulation in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e28013. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Yang, J.; Ma, C.L.; Zhang, Y.; Ge, T.; Qi, Z.; Kang, Y. Arabidopsis ROOT HAIR DEFECTIVE3 is involved in nitrogen starvation-induced anthocyanin accumulation. J. Integr. Plant Biol. 2015, 57, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Rahman, Z.A. Involvement of nitrogen on flavonoids, glutathione, anthocyanin, ascorbic acid and antioxidant activities of Malaysian medicinal plant Labisia pumila Blume (Kacip Fatimah). Int. J. Mol. Sci. 2012, 13, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, L.; Ravishankar, G.A.; Venkataraman, L.V.; Prathiba, K.R. Anthocyanin production in callus cultures of Daucus carota as influenced by nutrient stress and osmoticum. Biotechnol. Lett. 1992, 14, 707–712. [Google Scholar] [CrossRef]
- Seljasen, R.; Lea, P.; Torp, T.; Riley, H.; Berentsen, E.; Thomsen, M.; Bengtsson, G.B. Effects of genotype, soil type, year and fertilisation on sensory and morphological attributes of carrots (Daucus carota L.). J. Sci. Food Agric. 2012, 92, 1786–1799. [Google Scholar] [CrossRef] [PubMed]
- Salo, T.; Raisio, R.; Tiilikkala, K. Effectiveness of fertilizer recommendations in Finnish carrot and pea production. Acta Hortic. 1999, 506, 37–40. [Google Scholar] [CrossRef]
- Smoleń, S.; Sady, W. The effect of various nitrogen fertilization and foliar nutrition regimes on the concentrations of sugars, carotenoids and phenolic compounds in carrot (Daucus carota L.). Sci. Hortic. 2009, 120, 315–324. [Google Scholar] [CrossRef]
- Bleasdale, J.K.A. Control of size and yield in relation to harvest date of carrot roots. Acta Hortic. 1973, 27, 134–143. [Google Scholar] [CrossRef]
- Venter, F. Nitrate contents in carrots (Daucus carota L.) as influenced by fertilization. Acta Hortic. 1979, 93, 163–172. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2014. Available online: http://www.r-project.org/ (accessed on 23 October 2017).
- Ensminger, I.; Xyländer, M.; Hagen, C.; Braune, W. Strategies providing success in a variable habitat. III. Dynamic control of photosynthesis in Cladophora glomerata. Plant Cell Environ. 2001, 24, 769–779. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Durst, R.W.; Lee, J. Tracking color and pigment changes in anthocyanin products. Trends Food Sci. Technol. 2005, 16, 423–428. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 1 December 2017).
- Xia, L.L.; Lam, S.K.; Chen, D.L.; Wang, J.Y.; Tang, Q.; Yan, X.Y. Can knowledge-based N management produce more staple grain with lower greenhouse gas emission and reactive nitrogen pollution? A meta-analysis. Glob. Chang. Biol. 2017, 23, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Veitch, R.S.; Lada, R.R.; Adams, A.; MacDonald, M.T. Carrot yield and quality as influenced by nitrogen application in cut-and-peel carrots. Commun. Soil Sci. Plant Anal. 2014, 45, 887–895. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Seepaul, R.; George, S.; Wright, D.L. Comparative response of Brassica carinata and B. napus vegetative growth, development and photosynthesis to nitrogen nutrition. Ind. Crops Prod. 2016, 94, 872–883. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Heens, B. Fertilisation azotée des légumes industriels. Biotechnol. Agron. Soc. Environ. 2013, 17, 207–214. Available online: http://popups.ulg.ac.be/1780-4507/index.php?id=9695 (accessed on 1 December 2017).
- Westerveld, S.M.; McKeown, A.W.; Scott-Dupree, C.D.; McDonald, M.R. How well do critical nitrogen concentrations work for cabbage, carrot, and onion crops? Hortscience 2003, 38, 1122–1128. [Google Scholar]
- Westerveld, S.M.; McKeown, A.W.; McDonald, M.R. Seasonal nitrogen partitioning and nitrogen uptake of carrots as affected by nitrogen application in a mineral and an organic soil. Hortscience 2006, 41, 1332–1338. [Google Scholar]
- Westerveld, S.M.; McKeown, A.W.; McDonald, M.R. Carrot yield, quality, and storability in relation to preplant and residual nitrogen on mineral and organic soils. HortTechnology 2006, 16, 286–293. [Google Scholar]
- Sorensen, J.N. Nitrogen effects on vegetable crop production and chemical composition. Acta Hortic. 1999, 506, 41–49. [Google Scholar] [CrossRef]
- Beys, P.F. Einfluss von Wasserverfügbarkeit und Erntezeitpunkt auf den Ertrag und Wertgebende Inhaltsstoffe der violetten Möhre. Bachelor’s Thesis, FH Bingen, Bingen, Germany, 25 June 2012. [Google Scholar]
- Westerveld, S.M.; McDonald, M.R.; McKeown, A.W. Nitrogen utilization timeline of carrot over the growing season. Can. J. Plant Sci. 2007, 87, 587–592. [Google Scholar] [CrossRef]
- Fritz, D.; Weichmann, J. Influence of the harvesting date of carrots on quality and quality preservation. Acta Hortic. 1979, 93, 91–100. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N Supply (% of Recommendation) | Harvesting Date |
|---|---|---|
| N-1 | 100 | 21 September 2015 |
| Red-1 | 33 | 21 September 2015 |
| N-2 | 100 | 12 October 2015 |
| Red-2 | 33 | 12 October 2015 |
| N-3 | 100 | 2 November 2015 |
| Red-3 | 33 | 2 November 2015 |
| Treatment | FMleaf (g per plant) |
|---|---|
| N-1 | 594 a |
| Red-1 | 525 a,b |
| N-2 | 419 b,c |
| Red-2 | 407 b,d |
| N-3 | 359 c,d |
| Red-3 | 329 c,d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, L.; Sorg, S.; Tittmann, S.; Max, J.F.J.; Zinkernagel, J. Do Extended Cultivation Periods and Reduced Nitrogen Supply Increase Root Yield and Anthocyanin Content of Purple Carrots? Horticulturae 2018, 4, 7. https://doi.org/10.3390/horticulturae4020007
Schmidt L, Sorg S, Tittmann S, Max JFJ, Zinkernagel J. Do Extended Cultivation Periods and Reduced Nitrogen Supply Increase Root Yield and Anthocyanin Content of Purple Carrots? Horticulturae. 2018; 4(2):7. https://doi.org/10.3390/horticulturae4020007
Chicago/Turabian StyleSchmidt, Lilian, Sophia Sorg, Susanne Tittmann, Johannes F. J. Max, and Jana Zinkernagel. 2018. "Do Extended Cultivation Periods and Reduced Nitrogen Supply Increase Root Yield and Anthocyanin Content of Purple Carrots?" Horticulturae 4, no. 2: 7. https://doi.org/10.3390/horticulturae4020007