
The Subunit Nto1 of the NuA3 Complex Is Associated with Conidiation, Oxidative Stress Response, and Pathogenicity in Fusarium oxysporum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Media
2.2. Identification and Phylogenetic Analysis of Nto1
2.3. Construction of Nto1Deleted Mutants
2.4. Complementation of Nto1-Deleted Mutants
2.5. Conidiation
2.6. Oxidant Stressor Treatment
2.7. Plant Material and Inoculation
3. Results
3.1. Identification of Nto1 in F. oxysporum
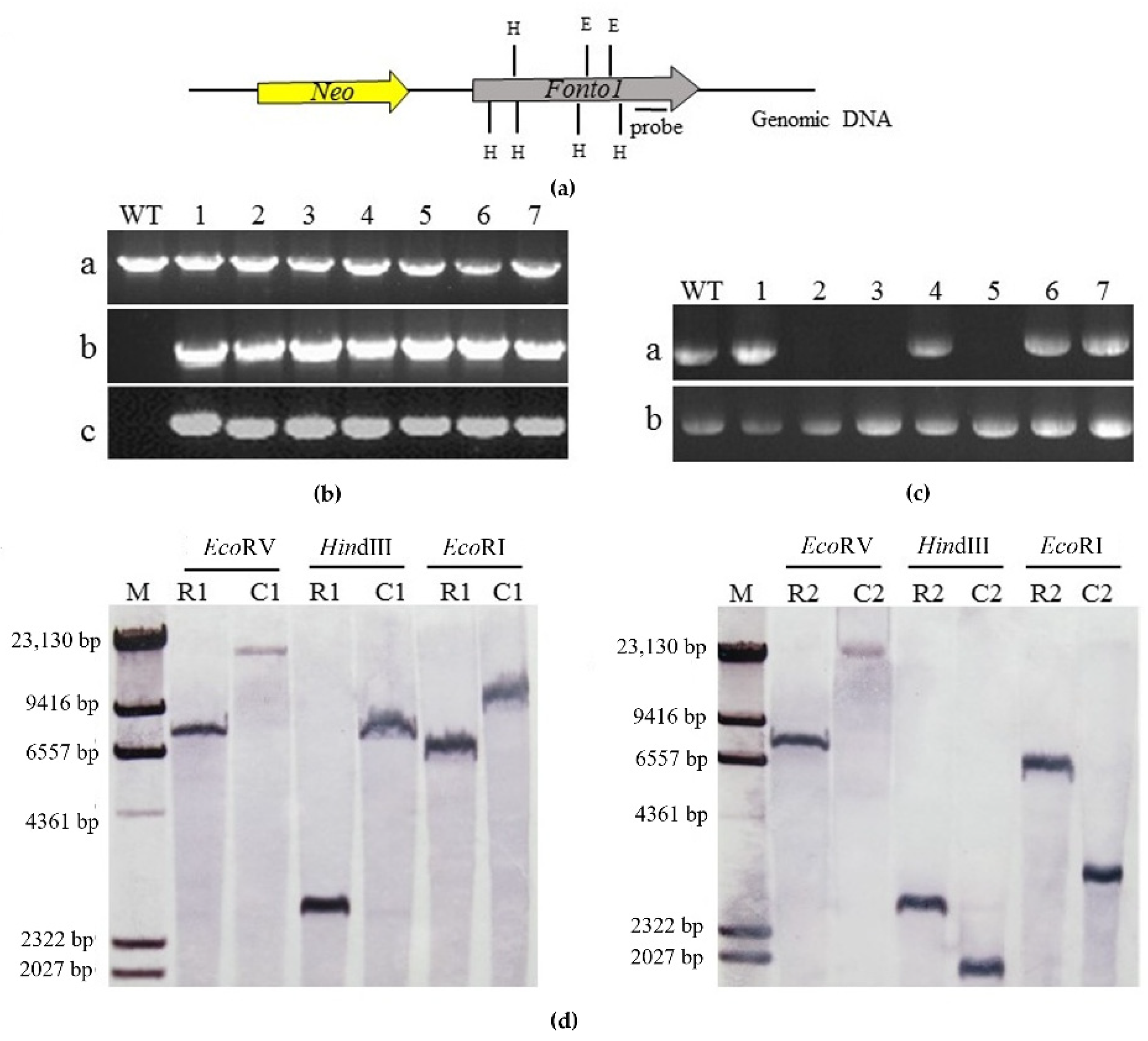
3.2. Deletion of Nto1 in R1 and R2
3.3. Complementation of Nto1-Deleted Mutants
3.4. Nto1 Disruption Resulted in Reduction in Microconidium Formation and Abnormal Conidium Color
3.5. Nto1 Disruption Leads to Hypersensitivity to Oxidative Stress
3.6. Nto1 Is Essential for Full Virulence of F. oxysporum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Correll, J.C. The relationship between formae speciales, races, and vegetative compatibility groups in Fusarium oxysporum. Phytopathology 1991, 81, 1061–1064. [Google Scholar]
- Liu, X.; Ling, J.; Xiao, Z.; Xie, B.; Fang, Z.; Yang, L.; Zhang, Y.; Lv, H.; Yang, Y. Characterization of emerging populations of Fusarium oxysporum f. sp. conglutinans causing cabbage wilt in China. J. Phytopathol. 2017, 165, 813–821. [Google Scholar] [CrossRef]
- Ma, L.J.; van der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Ling, J.; Wang, G.; Xiao, J.; Yang, Y.; Mao, Z.; Wang, X.; Xie, B. Comparative proteomics analyses of two races of Fusarium oxysporum f. sp. conglutinans that differ in pathogenicity. Sci. Rep. 2015, 5, 13663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietro, A.D.; Madrid, M.P.; Caracuel, Z.; Delgado-Jarana, J.; Roncero, M.J.G. Fusarium oxysporum: Exploring the molecular arsenal of a vascular wilt fungus. Mol. Plant Pathol. 2003, 4, 315–325. [Google Scholar] [CrossRef]
- Ohara, T.; Inoue, I.; Namiki, F.; Kunoh, H.; Tsuge, T. REN1 is required for development of microconidia and macroconidia, but not of chlamydospores, in the plant pathogenic fungus Fusarium oxysporum. Genetics 2004, 166, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Ohara, T.; Tsuge, T. FoSTUA, encoding a basic helix-loop-helix protein, differentially regulates development of three kinds of asexual spores, macroconidia, microconidia, and chlamydospores, in the fungal plant pathogen Fusarium oxysporum. Eukaryot. Cell 2004, 3, 1412–1422. [Google Scholar] [CrossRef] [Green Version]
- Wenhui, Z.; Xu, Z.; Qiurong, X.; Qingping, H.; Chengkang, Z.; Huanchen, Z.; Liping, X.; Guodong, L.; Won-Bo, S.; Zonghua, W. A conserved homeobox transcription factor Htf1 is required for phialide development and conidiogenesis in Fusarium species. PLoS ONE 2012, 7, e45432. [Google Scholar]
- Denisov, Y.; Freeman, S.; Yarden, O. Inactivation of Snt2, a BAH/PHD-containing transcription factor, impairs pathogenicity and increases autophagosome abundance in Fusarium oxysporum. Mol. Plant Pathol. 2011, 12, 449–461. [Google Scholar] [CrossRef]
- Bart, L.; Houterman, P.M.; Martijn, R. Effector gene screening allows unambiguous identification of Fusarium oxysporum f. sp. lycopersici races and discrimination from other formae speciales. FEMS Microbiol. Lett. 2009, 300, 201–215. [Google Scholar]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Shahbazian, M.D.; Grunstein, M. Functions of site-specific histone acetylation and deacetylation. Annu. Rev. Biochem. 2007, 76, 75–100. [Google Scholar] [CrossRef]
- Gómez-Diaz, E.; Jordà, M.; Peinado, M.A.; Rivero, A. Epigenetics of host-pathogen interactions: The road ahead and the road behind. PLoS Pathog. 2012, 8, e1003007. [Google Scholar] [CrossRef] [Green Version]
- Xiangjiu, K.; Van, D.A.D.; Van, D.L.T.A.J.; Cees, W.; Jingsheng, X.; Jin, X.; Hao, Z.; Wanquan, C.; Jie, F. The Fusarium graminearum histone acetyltransferases are important for morphogenesis, DON biosynthesis, and pathogenicity. Front. Microbiol. 2018, 9, 654. [Google Scholar]
- Carrozza, M.J.; Utley, R.T.; Workman, J.L.; Cote, J. The diverse functions of histone acetyltransferase complexes. Trends Genet. 2003, 19, 321–329. [Google Scholar] [CrossRef]
- Joseph, T.; Christopher, G.; Michael, G.R. Co-activators and co-repressors in the integration of transcriptional responses. Curr. Opin. Cell Biol. 1998, 10, 373–383. [Google Scholar]
- Vicente-Muoz, S.; Romero, P.; Magraner-Pardo, L.; Martinez-Jimenez, C.P.; Tordera, V.; Pamblanco, M. Comprehensive analysis of interacting proteins and genome-wide location studies of the Sas3-dependent NuA3 histone acetyltransferase complex. FEBS Open Bio 2014, 4, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, T.M.; Mcdaniel, S.L.; Byrum, S.D.; Cades, J.A.; Dancy, B.C.R.; Wade, H.; Tackett, A.J.; Strahl, B.D.; Taverna, S.D. A PWWP domain-containing protein targets the NuA3 acetyltransferase complex via histone H3 lysine 36 trimethylation to coordinate transcriptional elongation at coding regions. Mol. Cell Proteom. 2014, 13, 2883–2895. [Google Scholar] [CrossRef] [Green Version]
- John, S.; Howe, L.A.; Tafrov, S.T.; Grant, P.A.; Sternglanz, R.; Workman, J.L. The something about silencing protein, Sas3, is the catalytic subunit of NuA3, a yTAFII30-containing HAT complex that interacts with the Spt16 subunit of the yeast CP (Cdc68/Pob3)-FACT complex. Genes Dev. 2000, 14, 1196–1208. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Lee, J.; Kwon, S.; Lee, Y.; Jeon, J. A MYST family histone acetyltransferase, MoSAS3, is required for development and pathogenicity in the rice blast fungus. Mol. Plant Pathol. 2019, 20, 1491–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Yoon, C.Y.; Jun, Y.; Lee, B.B.; Lee, J.E.; Ha, S.D.; Woo, H.; Choi, A.; Lee, S.; Jeong, W.; et al. NuA3 HAT antagonizes the Rpd3S and Rpd3L HDACs to optimize mRNA and lncRNA expression dynamics. Nucleic Acids Res. 2020, 48, 10753–10767. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Xia, A.; Ye, M.; Ren, J.; Jiang, C. Opposing functions of Fng1 and the Rpd3 HDAC complex in H4 acetylation in Fusarium graminearum. PLoS Genet. 2020, 16, e1009185. [Google Scholar] [CrossRef]
- Xu, H.; Ye, M.; Xia, A.; Jiang, H.; Huang, P.; Liu, H.; Hou, R.; Wang, Q.; Li, D.; Xu, J.R.; et al. The Fng3 ING protein regulates H3 acetylation and H4 deacetylation by interacting with two distinct histone modifying complexes. New Phytol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Kachirskaia, L.; Walter, K.L.; Kuo, J.H.A.; Lake, A.; Davrazou, F.; Chan, S.M.; Martin, D.G.E.; Fingerman, I.M.; Briggs, S.D.; et al. Proteome-wide analysis in Saccharomyces cerevisiae identifies several PHD fingers as novel direct and selective binding modules of histone H3 methylated at either lysine 4 or lysine 36. J. Biol. Chem. 2007, 282, 2450–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Li, J.; Cheng, L.; Ling, J.; Luo, Z.; Bai, M.; Xie, B. A high efficiency gene disruption strategy using a positive-negative split selection marker and electroporation for Fusarium oxysporum. Microbiol. Res. 2014, 169, 835–843. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Z.; Lin, R.; Li, E.; Mao, Z.; Ling, J.; Yang, Y.; Yin, W.B.; Xie, B. Biosynthesis of antibiotic leucinostatins in bio-control fungus Purpureocillium lilacinum and their inhibition on Phytophthora revealed by genome mining. PLoS Pathog. 2016, 12, e1005685. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Zhang, H.; Wang, Y.; Liu, F.; Li, E.; Ma, J.; Yang, B.; Zhang, C.; Li, L.; Liu, Y. Requirement of LaeA, VeA, and VelB on asexual development, ochratoxin A biosynthesis, and fungal virulence in Aspergillus ochraceus. Front. Microbiol. 2019, 10, 2759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, E.; Wang, G.; Xiao, J.; Ling, J.; Yang, Y.; Xie, B. A SIX1 homolog in Fusarium oxysporum f. sp. conglutinans is required for full virulence on cabbage. PLoS ONE 2016, 11, e0152273. [Google Scholar]
- Nicholson, P.; Parry, D.W. Development and use of a PCR assay to detect Rhizoctonia cerealis, the cause of sharp eyespot in wheat. Plant Pathol. 1996, 45, 872–883. [Google Scholar] [CrossRef]
- Gao, T.; Zheng, Z.; Hou, Y.; Zhou, M. Transcription factors spt3 and spt8 are associated with conidiation, mycelium growth, and pathogenicity in Fusarium graminearum. FEMS Microbiol. Lett. 2014, 351, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doke, N.; Miura, Y.; Sanchez, L.; Park, H.; Noritake, T.; Yoshioka, H.; Kawakita, K. The oxidative burst protects plants against pathogen attack: Mechanism and role as an emergency signal for plant bio-defence-a review. Gene 1996, 179, 45–51. [Google Scholar] [CrossRef]
- Gan, Y.; Zhang, L.; Zhang, Z.; Dong, S.; Li, J.; Wang, Y.; Zheng, X. The LCB2 subunit of the sphingolip biosynthesis enzyme serine palmitoyltransferase can function as an attenuator of the hypersensitive response and Bax-induced cell death. New Phytol. 2009, 181, 127–146. [Google Scholar] [CrossRef]
- Guo, M.; Chen, Y.; Du, Y.; Dong, Y.; Guo, W.; Zhai, S.; Zhang, H.; Dong, S.; Zhang, Z.; Wang, Y.; et al. The bZIP transcription factor MoAP1 mediates the oxidative stress response and is critical for pathogenicity of the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1001302. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Schopfer, P. Hydroxyl-radical production in physiological reactions: A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef]
- Averyanov, A. Oxidative burst and plant disease resistance. Front. Biosci. 2009, 1, 142–152. [Google Scholar]
- Mayer, A.M.; Staples, R.C.; Gil-ad, N.L. Mechanisms of survival of necrotrophic fungal plant pathogens in hosts expressing the hypersensitive response. Phytochemistry 2001, 58, 33–41. [Google Scholar] [CrossRef]
- Moye-Rowley, W.S. Regulation of the transcriptional response to oxidative stress in fungi: Similarities and differences. Eukaryot. Cell 2003, 2, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisov, Y.; Yarden, O.; Freeman, S. The transcription factor SNT2 is involved in fungal respiration and reactive oxidative stress in Fusarium oxysporum and Neurospora crassa. Physiol. Mol. Plant Pathol. 2011, 76, 137–143. [Google Scholar] [CrossRef]
- Baker, L.A.; Ueberheide, B.M.; Dewell, S.; Chait, B.T.; Zheng, D.; Allis, C.D. The yeast Snt2 protein coordinates the transcriptional response to hydrogen peroxide-mediated oxidative stress. Mol. Cell. Biol. 2013, 33, 3735–3748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Gonzalez, M.; Kabbaj, M.H.M.; Gunjan, A. Novel E3 ubiquitin ligases that regulate histone protein levels in the budding yeast Saccharomyces cerevisiae. PLoS ONE 2012, 7, e36295. [Google Scholar] [CrossRef] [Green Version]
- Papamichos-Chronakis, M.; Petrakis, T.; Ktistaki, E.; Topalidou, I.; Tzamarias, D. Cti6, a PHD domain protein, bridges the Cyc8-Tup1 corepressor and the SAGA coactivator to overcome repression at GAL1. Mol. Cell 2002, 9, 1297–1305. [Google Scholar] [CrossRef]
- Puig, S.; Lau, M.; Thiele, D.J. Cti6 is an Rpd3-Sin3 histone deacetylase-associated protein required for growth under iron-limiting conditions in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 30298–30306. [Google Scholar] [CrossRef] [Green Version]
- Bian, C.; Xu, C.; Ruan, J.; Lee, K.K.; Burke, T.L.; Tempel, W.; Barsyte, D.; Li, J.; Wu, M.; Zhou, B.; et al. Sgf29 binds histone H3K4me2/3 and is required for SAGA complex recruitment and histone H3 acetylation. EMBO J. 2011, 30, 2829–2842. [Google Scholar] [CrossRef] [Green Version]
- Pray-Grant, M.G.; Daniel, J.A.; Schieltz, D.; Yates, J.R.; Grant, P.A. Chd1 chromodomain links histone H3 methylation with SAGA-and SLIK-dependent acetylation. Nature 2005, 433, 434–438. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Microbiology, biosynthesis of naphthoquinone pigments by fungi of the genus Fusarium. Appl. Biochem. Microbiol. 2005, 41, 503–507. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, E.; Xiao, J.; Yang, Y.; Xie, B.; Mao, Z. The Subunit Nto1 of the NuA3 Complex Is Associated with Conidiation, Oxidative Stress Response, and Pathogenicity in Fusarium oxysporum. Horticulturae 2022, 8, 540. https://doi.org/10.3390/horticulturae8060540
Li E, Xiao J, Yang Y, Xie B, Mao Z. The Subunit Nto1 of the NuA3 Complex Is Associated with Conidiation, Oxidative Stress Response, and Pathogenicity in Fusarium oxysporum. Horticulturae. 2022; 8(6):540. https://doi.org/10.3390/horticulturae8060540
Chicago/Turabian StyleLi, Erfeng, Jiling Xiao, Yuhong Yang, Bingyan Xie, and Zhenchuan Mao. 2022. "The Subunit Nto1 of the NuA3 Complex Is Associated with Conidiation, Oxidative Stress Response, and Pathogenicity in Fusarium oxysporum" Horticulturae 8, no. 6: 540. https://doi.org/10.3390/horticulturae8060540