Volatiles of Zanthoxylum limoncello as Antifungal Agents against the Postharvest Rot of Manzano Pepper Triggered by Fusarium temperatum

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Obtainment of Zanthoxylum limoncello Leaf Essential Oil
2.3. GC Profiling of Zanthoxylum limoncello Essential Oil (ZlEO)
2.4. Isolation of F. temperatum from Manzano Peppers
2.5. Antifungal Assays
2.6. Determination of pH and Fruit Firmness
2.7. Proximate Analysis and Ascorbic Acid Determination
2.8. Determination of Carotenoids in Manzano Peppers
2.9. Determination of Capsaicinoids in Manzano Peppers
2.10. Statistical Analysis
3. Results
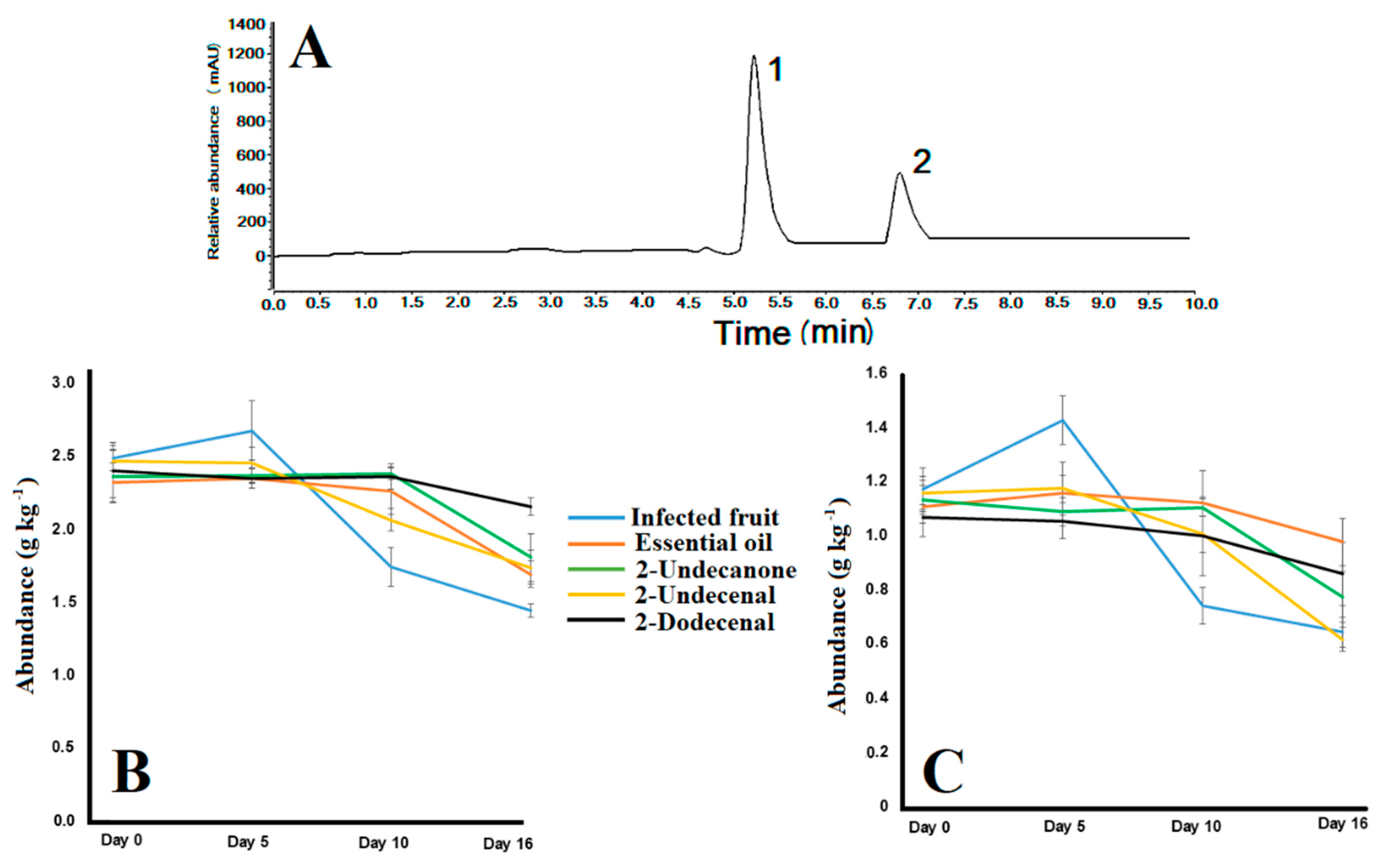
3.1. Essential Oil Composition and In Vitro Antifungal Activity
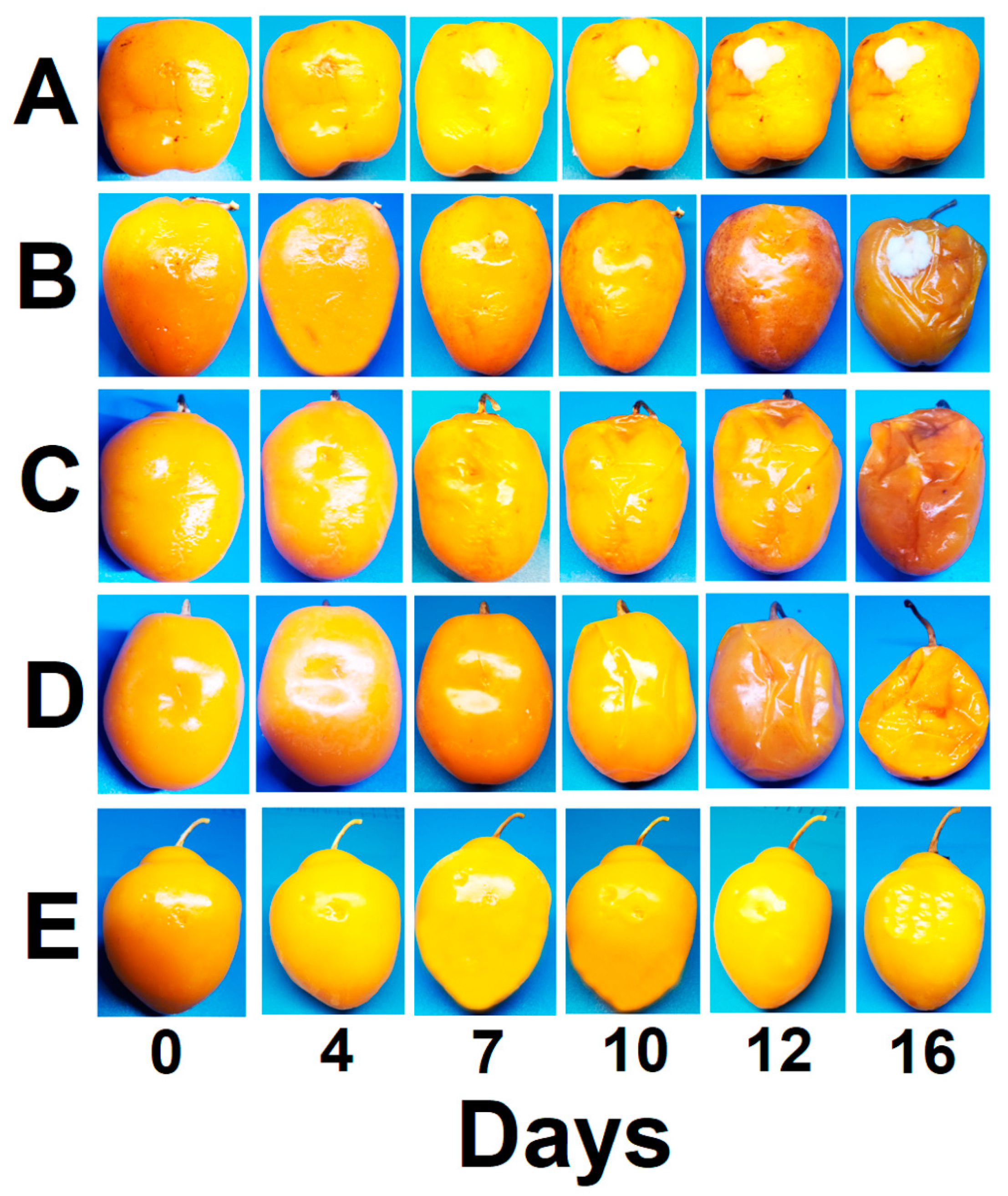
3.2. In Situ Antifungal Activity of ZlEO and Its Major Volatiles
3.3. Protective Effect of ZlEO and Its Volatiles on Fruit Firmness and pH
3.4. Protective Effect of ZlEO and Its Volatiles on the Nutritional Content of Manzano Pepper
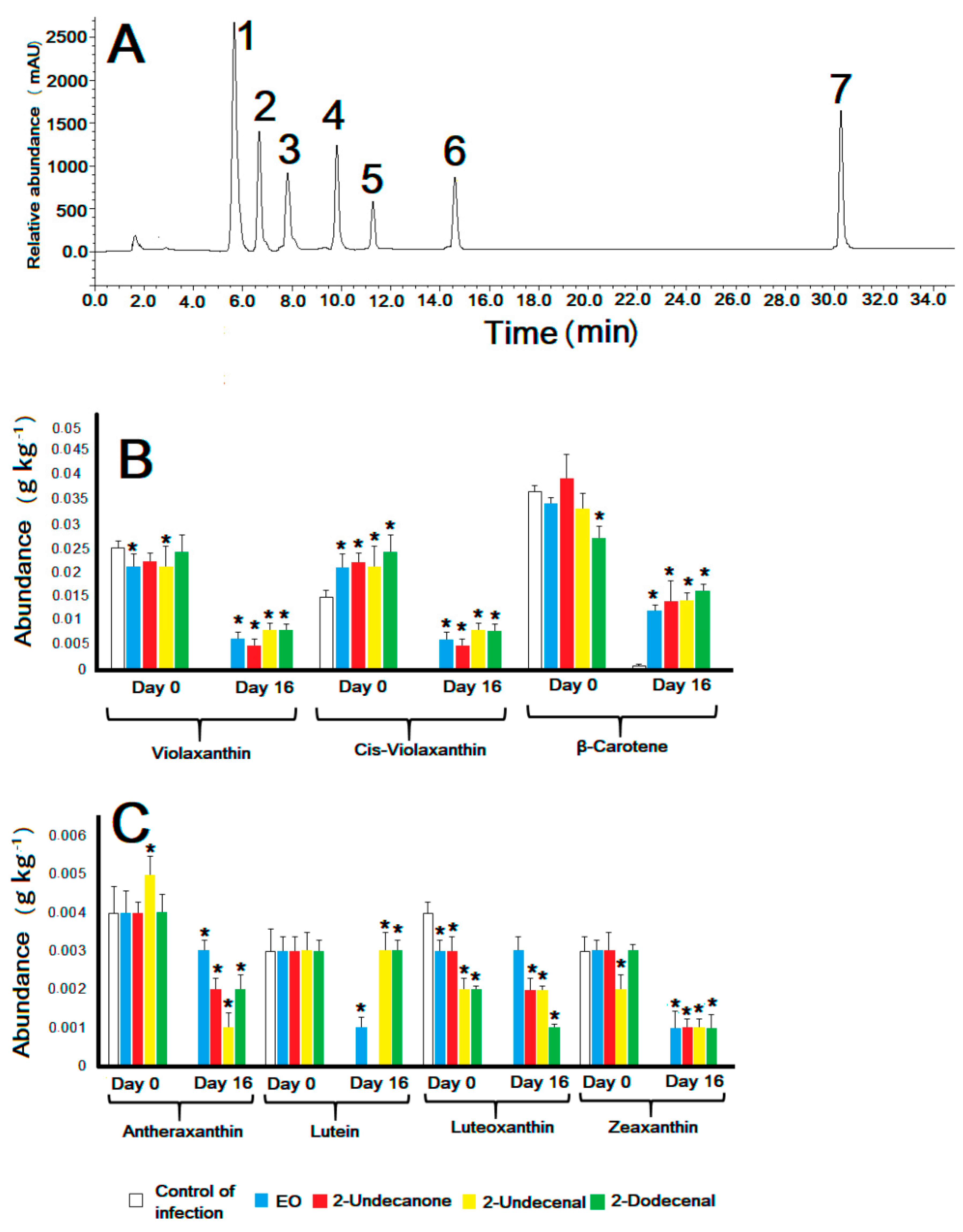
3.5. Protective Effect of ZlEO and Its Volatiles on the Nutraceutical Content of Manzano Pepper
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asnin, L.; Park, S.W. Isolation and analysis of bioactive compounds in Capsicum peppers. Crit. Rev. Food Sci. Nutr. 2015, 55, 254–289. [Google Scholar] [CrossRef] [PubMed]
- Meckelmann, S.; Jansen, C.; Riegel, D.; Zonneveld, M.; Ríos, L.; Peña, K.; Mueller-Seitz, E.; Petz, M. Phytochemicals in native Peruvian Capsicum pubescens (Rocoto). Eur. Food Res. Technol. 2015, 241, 817–825. [Google Scholar] [CrossRef]
- Espinosa-Torres, L.E.; Ramírez-Abarca, O. Rentabilidad de chile manzano (Capsicum pubescens R Y P) producido en invernadero en Texcoco, estado de México. Rev. Mex. Cienc. Agric. 2016, 7, 325–335. [Google Scholar]
- Villa-Ruano, N.; Velásquez-Valle, R.; Zepeda-Vallejo, L.G.; Pérez-Hernández, N.; Velázquez-Ponce, M.; Arcos-Adame, V.M.; Becerra-Martínez, E. 1H NMR-based metabolomic profiling for identification of metabolites in Capsicum annuum cv. mirasol infected by beet mild curly top virus (BMCTV). Food Res. Int. 2018, 106, 870–879. [Google Scholar] [CrossRef]
- Pérez-Grajales, M.; González-Hernández, V.A.; Mendoza-Castillo, M.C.; Peña-Valdivia, C. Physiological characterization of manzano hot pepper (Capsicum pubescens R & P) landraces. J. Am. Soc. Hort. Sci. 2004, 129, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Kumar-Pandey, S.; Kumar-Yadav, S.; Kumar-Singh, V. An overview on Capsicum annum L. J. Pharm. Sci. Technol. 2012, 4, 821–828. [Google Scholar]
- Sánchez, A.A.; Escobar, V.A.; Santamaría, I.F.T.; Angel, D.N. Fusarium sambucinum Fuckel agente causal de la pudrición de frutos de chile manzano (Capsicum pubescens) en México. Rev. Mex. Fitopatol. 2019, 37, 159–169. [Google Scholar] [CrossRef]
- Pérez-Vázquez, M.A.K.; Pacheco Hernández, Y.; Lozoya-Gloria, E.; Mosso-González, C.; Ramírez-García, S.; Romero-Arenas, O.; Villa-Ruano, N. Peppermint essential oil and major volatiles as protective agents against soft rot caused by Fusarium sambucinum in cera pepper (Capsicum pubescens). Chem. Biodivers. 2022, 19, e202100835. [Google Scholar] [CrossRef]
- Vallejo-Gutiérrez, A.J.; Mejía-Carranza, J.; García-Velasco, R.; Ramírez-Gerardo, M.G. Response of Capsicum pubescens genotypes to damage caused by the fungal wilt complex. Rev. Mex. Fitopatol. 2019, 37, 50–70. [Google Scholar] [CrossRef]
- Pérez-Vázquez, M.A.K.; Morales-Mora, L.A.; Romero-Arenas, O.; Rivera, A.; Landeta-Cortés, G.; Villa-Ruano, N. First report of Fusarium temperatum causing fruit blotch of Capsicum pubescens in Puebla, México. Plant Dis. 2022, 106, 1758. [Google Scholar] [CrossRef]
- Scauflaire, J.; Gourgue, M.; Munaut, F. Fusarium temperatum sp. nov. from maize, an emergent species closely related to Fusarium Subglutinans. Mycologia 2011, 103, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Fumero, M.V.; Reynoso, M.M.; Chulze, S. Fusarium temperatum and Fusarium subglutinans isolated from maize in Argentina. Int. J. Food Microbiol. 2015, 199, 86–92. [Google Scholar] [CrossRef]
- Sarkhosh, A.; Schaffer, B.; Vargas, A.I.; Palmateer, A.J.; López, P.; Soleymani, A. In vitro evaluation of eight plant essential oils for controlling Colletotrichum, Botryosphaeria, Fusarium and Phytophthora fruit rots of avocado, mango and papaya. Plant Protect. Sci. 2018, 54, 153–162. [Google Scholar] [CrossRef]
- Said-Omar, M.; Kordali, S. Review of essential oils as anti-fungal agents for plant fungal diseases. Zir. Fak. Derg. 2019, 14, 294–301. [Google Scholar]
- Pacheco-Hernández, Y.; Santamaría-Juárez, J.D.; Hernández-Silva, N.; Cruz-Durán, R.; Mosso-González, M.; Villa-Ruano, N. Essential oil of Lepidium virginicum: Protective activity on anthracnose disease and preservation effect on the nutraceutical content of tamarillo fruit (Solanum betaceum). Chem. Biodivers. 2021, 18, e2000941. [Google Scholar] [CrossRef]
- Villa-Ruano, N.; Pacheco-Hernández, Y.; Zárate-Reyes, J.A.; Cruz-Durán, R.; Lozoya-Gloria, E. Volatile composition and biological activities of the leaf essential oil from Zanthoxylum limoncello grown in Oaxaca, México. Chem. Biodivers. 2019, 16, e1800498. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 3rd ed.; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Chadha, S.; Kale, S.P. Simple fluorescence-based high throughput cell viability assay for filamentous fungi. Lett. Appl. Microbiol. 2015, 61, 238–244. [Google Scholar] [CrossRef]
- Villa-Ruano, N.; Pacheco-Hernández, Y.; Cruz-Duran, R.; Lozoya-Gloria, E.; Betancourt-Jiménez, M.B. Seasonal variation in phytochemicals and nutraceutical potential of Begonia nelumbiifolia consumed in Puebla, México. J. Food Sci. Technol. 2017, 54, 1484–1490. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rabanales, Q.N.; Cóyotl-Pérez, W.A.; Rubio-Rosas, E.; Cortes-Ramírez, G.S.; Sánchez-Ramírez, J.F.; Villa-Ruano, N. Antifungal properties of hybrid films containing the essential oil of Schinus molle: Protective effect against postharvest rot of tomato. Food Control 2022, 134, 108766. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; González-Mas, M.C.; Nuez, F. Carotenoid composition and vitamin A value in ají (Capsicum baccatum L.) and rocoto (C. pubescens R. & P.), 2 pepper species from the Andean region. Int. J. Food Sci. Technol. 2010, 75, S446–S453. [Google Scholar] [CrossRef]
- Teerarakm, M.; Chamroon, L. Essential oil from ginger as a novel agent in delaying senescence of cut fronds of the fern (Davallia solida (G. Forst.) Sw.). Postharvest Biol. Technol. 2019, 156, 110927. [Google Scholar] [CrossRef]
- Pacheco-Hernández, Y.; Villa-Ruano, N.; Lozoya-Gloria, E.; Barrales-Cortés, C.A.; Jiménez-Montejo, F.E.; Cruz-López, M.C. Influence of environmental factors on the genetic and chemical diversity of Brickellia veronicifolia populations growing in fragmented shrublands from Mexico. Plants 2021, 10, 325. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Shi, X.; Wang, B.; Li, M.; Wang, Q.; Zhang, S. Antifungal effect of volatile organic compounds from Bacillus velezensis CT32 against Verticillium dahliae and Fusarium oxysporum. Processes 2020, 8, 1674. [Google Scholar] [CrossRef]
- Jha, S.N.; Jaiswal, P.; Narsaiah, K.; Gupta, M.; Bhardwaj, R.; Singh, A.K. Non-destructive prediction of sweetness of intact mango using near infrared spectroscopy. Sci. Hort. 2012, 138, 171–175. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Veloso, J.; Prego, C.; Varela, M.M.; Carballeira, R.; Bernal, A.; Merino, F.; Díaz, J. Properties of capsaicinoids for the control of fungi and oomycetes pathogenic to pepper. Plan Biol. 2014, 16, 177–185. [Google Scholar] [CrossRef]
- Otaka, J.; Seo, S.; Nishimura, M. Lutein, a natural carotenoid, induces α-1,3-Glucan accumulation on the cell wall surface of fungal plant pathogens. Molecules 2016, 21, 980. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Number | Compound | RI | 1TRI | Abundance (%) | +Concentration |
|---|---|---|---|---|---|
| 1 | 1-Hexen-3-ol | 768 | 769 | 1.4 | ND |
| 2 | Camphene | 946 | 946 | 1.8 | ND |
| 3 | 1-Octanal | 996 | 998 | 1.1 | ND |
| 4 | β-Ocimene, (E)- | 1042 | 1044 | 2.0 | ND |
| 5 | 2-Nonanone | 1088 | 1087 | 2.3 | ND |
| 6 | * Linalool | 1095 | 1095 | 1.3 | 23.1 |
| 7 | Nonanal | 1102 | 1100 | 1.9 | ND |
| 8 | 2-decanone | 1190 | 1190 | 1.5 | ND |
| 9 | * 1-Decanal | 1203 | 1201 | 2.8 | 63.6 |
| 10 | (2E)-Decenal | 1258 | 1260 | 1.1 | ND |
| 11 | * 2-Undecanone | 1292 | 1293 | 34.0 | 313.9 |
| 12 | Undecanal | 1306 | 1035 | 1.3 | ND |
| 13 | * 2-Undecenal | 1362 | 1357 | 32.0 | 276.5 |
| 14 | * 2-Dodecenal | 1454 | 1454 | 8.4 | 128.2 |
| 15 | 2-Tridecanone | 1493 | 1495 | 2.9 | ND |
| Total | 95.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Arenas, O.; Pérez-Vázquez, M.A.K.; Rivera, A.; Pacheco-Hernández, Y.; Ramirez-Garcia, S.A.; Landeta-Cortés, G.; Villa-Ruano, N. Volatiles of Zanthoxylum limoncello as Antifungal Agents against the Postharvest Rot of Manzano Pepper Triggered by Fusarium temperatum. Horticulturae 2022, 8, 700. https://doi.org/10.3390/horticulturae8080700
Romero-Arenas O, Pérez-Vázquez MAK, Rivera A, Pacheco-Hernández Y, Ramirez-Garcia SA, Landeta-Cortés G, Villa-Ruano N. Volatiles of Zanthoxylum limoncello as Antifungal Agents against the Postharvest Rot of Manzano Pepper Triggered by Fusarium temperatum. Horticulturae. 2022; 8(8):700. https://doi.org/10.3390/horticulturae8080700
Chicago/Turabian StyleRomero-Arenas, Omar, Marco A. Kevin Pérez-Vázquez, Antonio Rivera, Yesenia Pacheco-Hernández, Sergio Alberto Ramirez-Garcia, Gerardo Landeta-Cortés, and Nemesio Villa-Ruano. 2022. "Volatiles of Zanthoxylum limoncello as Antifungal Agents against the Postharvest Rot of Manzano Pepper Triggered by Fusarium temperatum" Horticulturae 8, no. 8: 700. https://doi.org/10.3390/horticulturae8080700