
Identification of Genes of Molecular Marker TGS0892 on Chromosome 6 and Its Mechanism of Soluble Solids Metabolism in Tomato


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Determination of Physiological and Quality Indicators
2.3. Identification and Bioinformatics Analysis of Genes within 50 kb Upstream and Downstream of TGS0892
2.4. Extraction of Total RNA and Synthesis of cDNA
2.5. Gene Expression Analysis
2.6. Gene Co-Expression Network Analysis
3. Results
3.1. Growth Status of Different Cultivars of Mature Tomato Fruits
3.2. Physiological and Quality Indicators of Mature Tomato Fruits
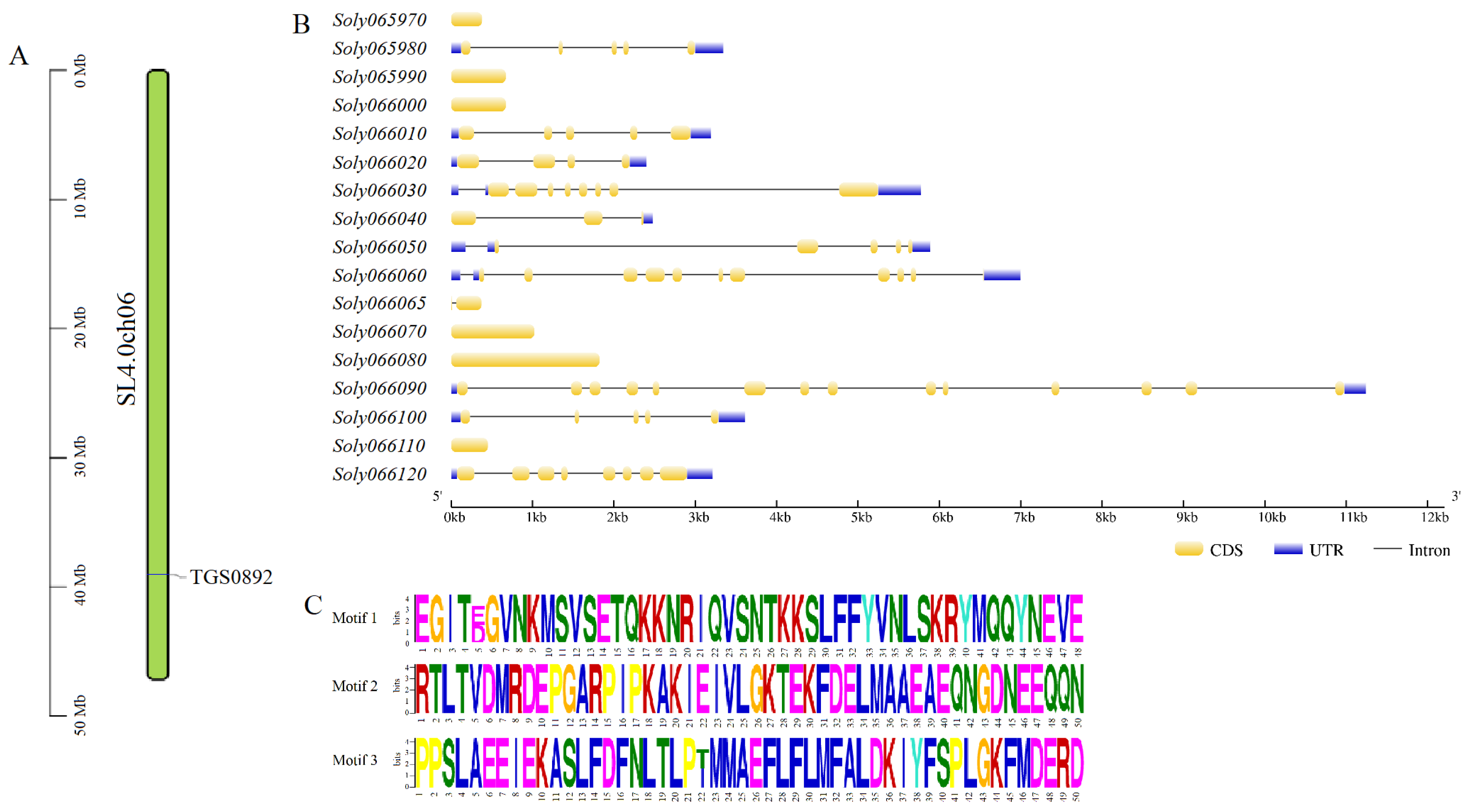
3.3. Identification and Sequence Analysis of Genes within 50 kb Upstream and Downstream of TGS0892
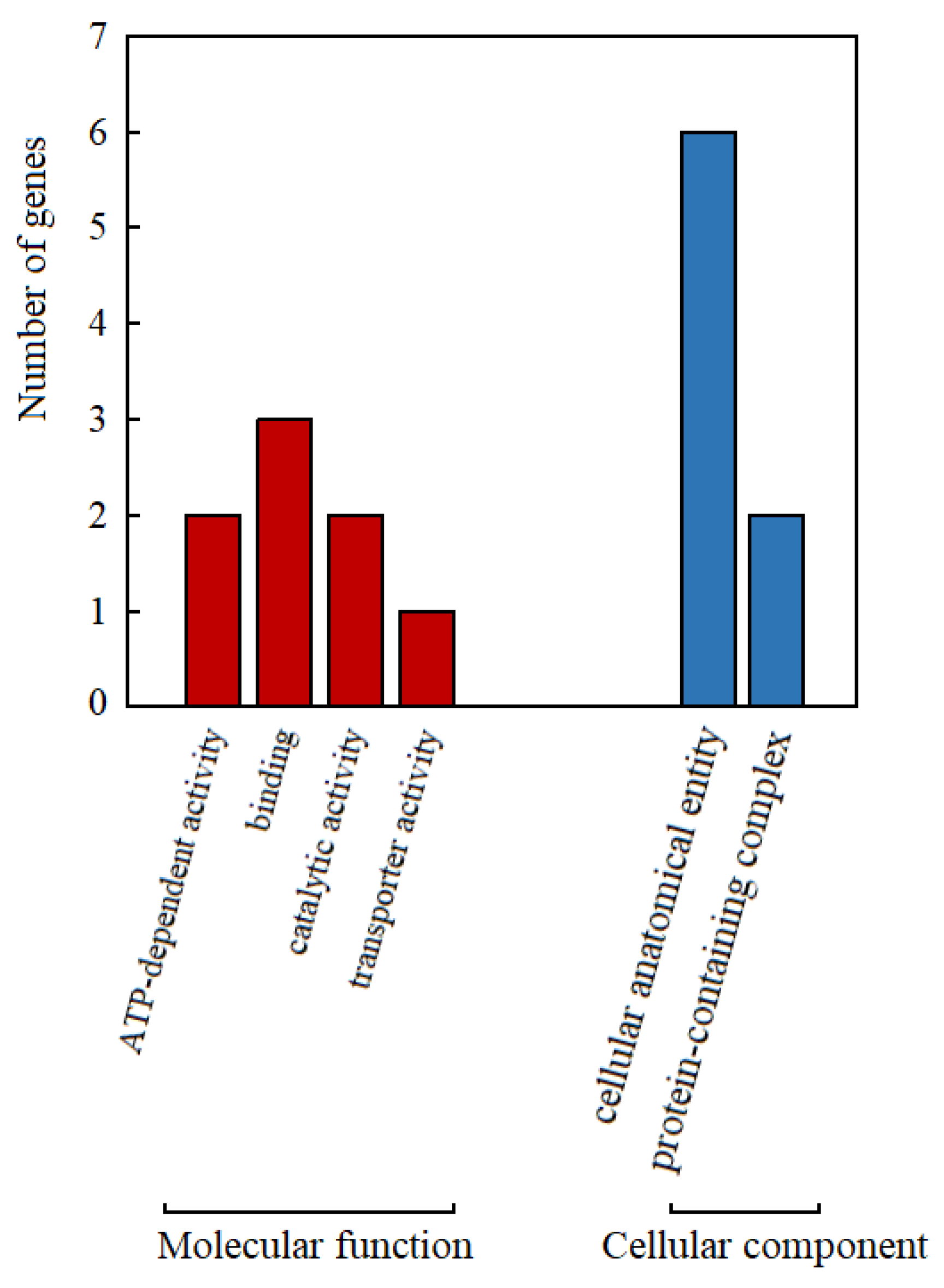
3.4. GO Functional Annotation of Genes within 50 kb Upstream and Downstream of TGS0892
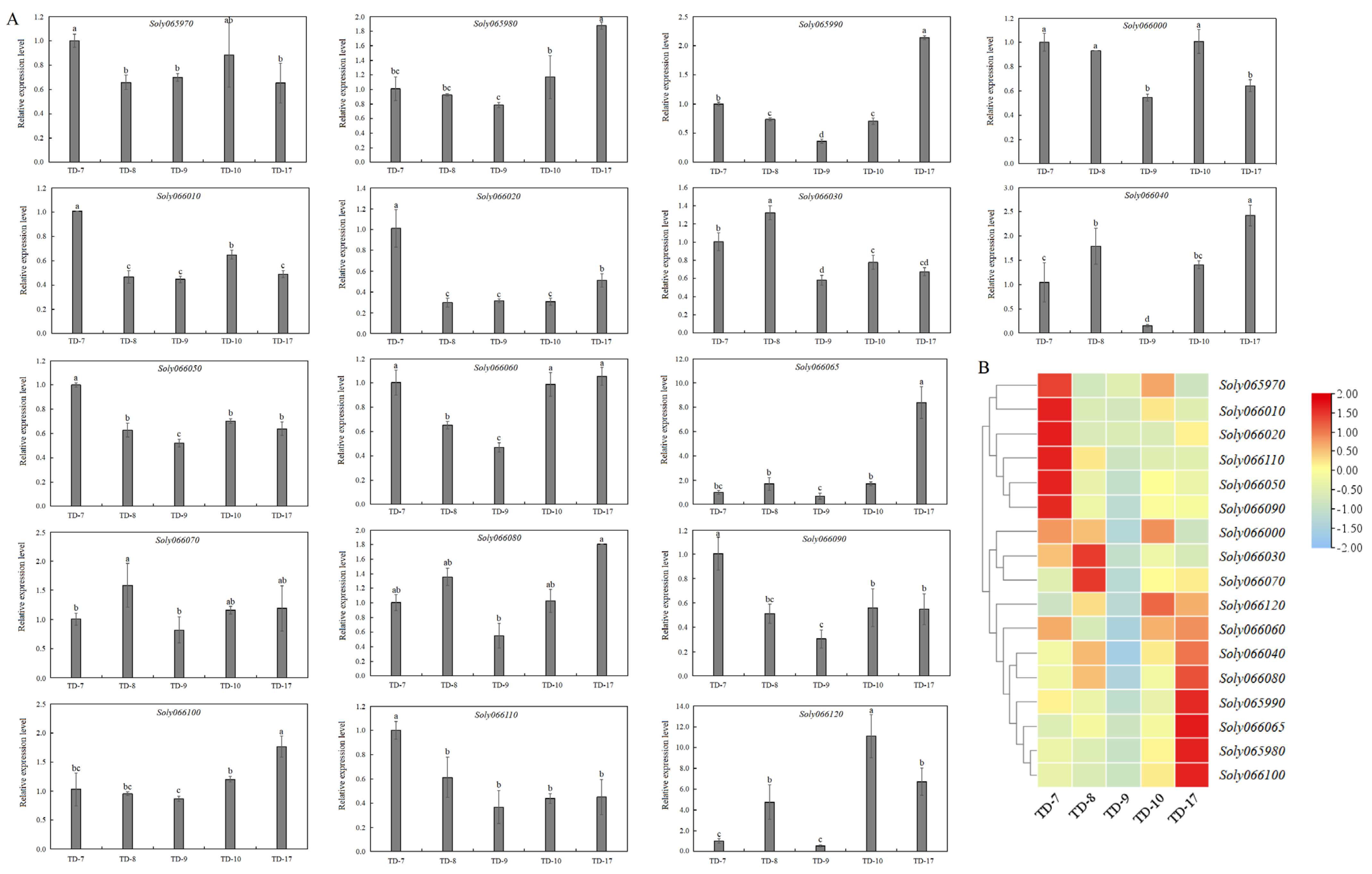
3.5. Expression Profiles of Genes within 50 kb Upstream and Downstream of TGS0892 in Different Tomato Cultivars
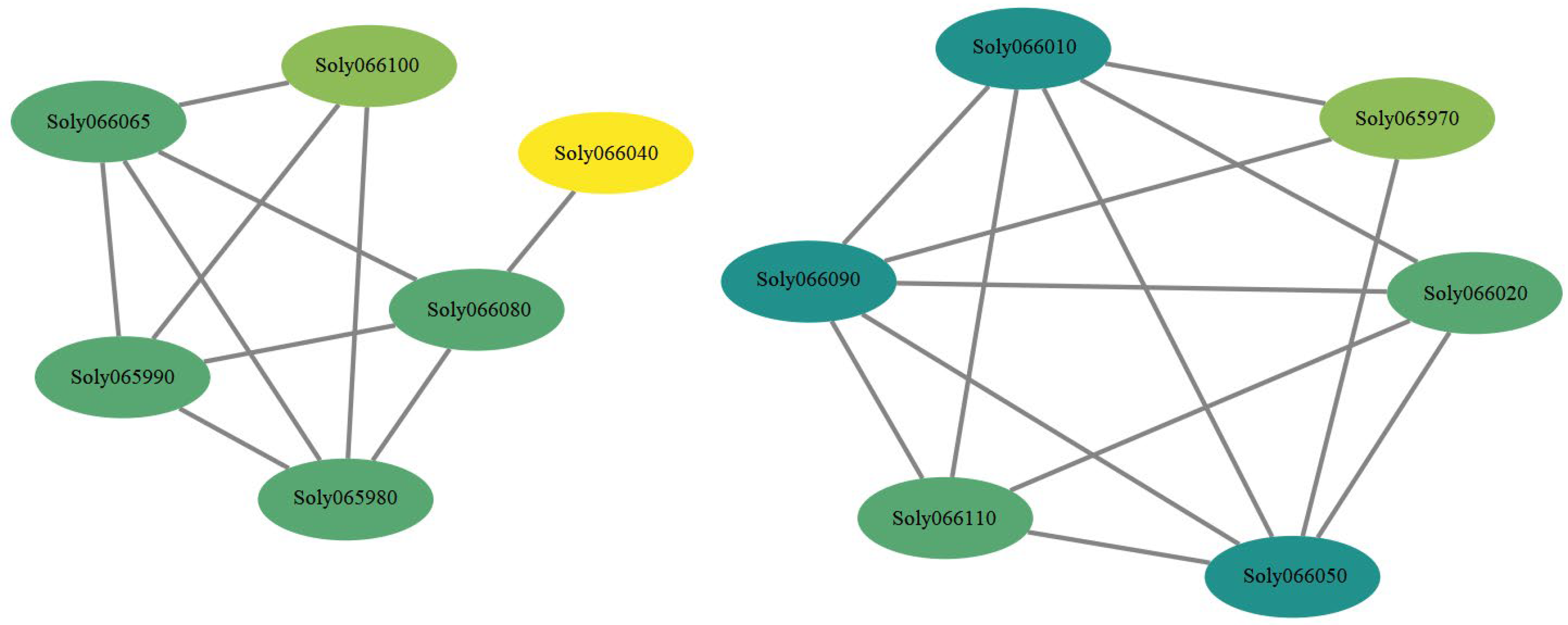
3.6. Interaction Network of Genes within 50 kb Upstream and Downstream of TGS0892
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Choudhary, P.; Rai, G.K.; Bagati, S.; Jamwal, D.; Kumar, P. Evaluation of genetic variability in tomato (Solanum lycopersicum L. Mill) genotypes using microsatellite markers. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2117–2124. [Google Scholar] [CrossRef]
- Ilahy, R.; Riahi, A.; Hdider, C.; Tlili, I.; Dalessandro, G.; Lenucci, M.S. Carotenoids content in intact plastids isolated from ordinary and high-lycopene tomato (Solatium lycopersicum L.) cultivars. Acta Hortic. 2015, 1081, 135–140. [Google Scholar] [CrossRef]
- Fulton, T.M.; Bucheli, P.; Voirol, E.; Tanksley, S.D. Quantitative trait loci (QTL) affecting sugars, organic acids and other biochemical properties possibly contributing to flavor, identified in four advanced backcross populations of tomato. Euphytica 2002, 127, 163–177. [Google Scholar] [CrossRef]
- Karadeniz, F.; Ekşi, A. A research on chemical composition of tomato pulpe. Turk. J. Agric. For. 1996, 20, 445–448. [Google Scholar] [CrossRef]
- Islam, M.S. Variability in different physical and biochemical characteristics of six tomato genotypes according to stages of ripeness. Bangladesh J. Bot. 1997, 26, 137–147. [Google Scholar]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit-ScienceDirect. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Fridman, E.; Liu, Y.S.; Carmel-Goren, L.; Gur, A.; Shoresh, M.; Pleban, T.; Eshed, Y.; Zamir, D. Two tightly linked QTLs modify tomato sugar content via different physiological pathways. Mol. Genet Genom. 2002, 266, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Chaïb, J.; Lecomte, L.; Buret, M.; Hospital, F. Both additivity and epistasis control. the genetic variation for fruit quality traits in tomato. Theor. Appl. Genet. 2007, 115, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R. Genome mapping and molecular breeding of tomato. Int. J. Plant Genom. 2007, 2007, 64358. [Google Scholar] [CrossRef] [Green Version]
- Causse, M.; Duffe, P.; Gomez, M.C.; Buret, M.; Damidaux, R.; Zamir, D.; Gur, A.; Chevalier, C.; Lemaire-Chamley, M.; Rothan, C. A genetic map of candidate genes and QTLs involved in tomato fruit size and composition. J. Exp. Bot. 2004, 55, 1671–1685. [Google Scholar] [CrossRef]
- Foolad, M.R.; Panthee, D.R. Marker-assisted selection in tomato breeding. Crit. Rev. Plant Sci. 2012, 31, 93–123. [Google Scholar] [CrossRef]
- Gonias, E.D.; Ganopoulos, I.; Mellidou, I.; Bibi, A.C.; Kalivas, A.; Mylona, P.V.; Osanthanunkul, M.; Tsaftaris, A.; Madesis, P.; Doulis, A.G. Exploring genetic diversity of tomato (Solanum lycopersicum L.) germplasm of Genebank collection employing SSR and SCAR markers. Genet. Resour. Crop Evol. 2019, 66, 1295–1309. [Google Scholar] [CrossRef]
- Wang, S.H. QTLs Mapping for Tomato Fruit Weight, Shape and Solube Solid Content in Solanum lycopersicon × Solanum galapagense Recombinant Inbred Line. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, April 2015. [Google Scholar]
- Zhao, J.T. Genome-Wide Association Study for Main Sugars and Organic Acids in Tomato Fruit. Master’s Thesis, Northwest A&F University, Yangling, China, November 2016. [Google Scholar]
- Eshed, Y.; Zamir, D. An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Zamir, D. Less-than-additive epistatic interactions of quantitative trait loci in tomato. Genetics 1996, 143, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H. QTLs Mapping of Soluble Solid Content and Beta-Carotene Content in Solanum cheesmanii. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, June 2012. [Google Scholar]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.R.; Wang, Y.H.; Jia, M.; Zhang, R.R.; Liu, H.; Xu, Z.S.; Xiong, A.S. The phytochrome-interacting factor DcPIF3 of carrot plays a positive role in drought stress by increasing endogenous ABA level in Arabidopsis. Plant Sci. 2022, 322, 111367. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Huang, Y.; Li, M.Y.; Wu, P.; Xu, Z.S.; Que, F.; Wang, F.; Xiong, A.S. Members of WRKY Group III transcription factors are important in TYLCV defense signaling pathway in tomato (Solanum lycopersicum). BMC Genom. 2016, 17, 788. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in realtime RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Szabo, K.; Dulf, F.V.; Diaconeasa, Z.; Dan, C.V. Antimicrobial and antioxidant properties of tomato processing byproducts and their correlation with the biochemical composition. LWT-Food Sci. Technol. 2019, 116, 108558. [Google Scholar] [CrossRef]
- Miller, J.C.; Tanksley, S.D. RFLP analysis of phylogenetic relationships and genetic variation in the genus Lycopersicon. Theor. Appl. Genet. 1990, 80, 437–448. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, J.; Liang, Y.; Zou, Z. Genome-wide association-mapping for fruit quality traits in tomato. Euphytica 2016, 207, 439–451. [Google Scholar] [CrossRef]
- Schouten, H.J.; Tikunov, Y.; Verkerke, W.; Finkers, R.; Bovy, A.; Bai, Y.; Visser, R.G.F. Breeding has increased the diversity of cultivated tomato in the Netherlands. Front. Plant Sci. 2019, 10, 1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, J.J.; Noensie, E.N.; Ruezinsky, D.M.; Lu, X.; Tracy, S.L.; Ganal, M.W.; Martin, G.B.; Pillen, K.; Albert, K.; Tankslev, S.D. Molecular genetic analysis of the ripening-inhibitor and non-ripening loci of tomato: A first step in genetic map-based cloning of fruit ripening genes. Mol. Gen. Genet. 1995, 248, 195–206. [Google Scholar] [CrossRef]
- Chen, F.Q.; Foolad, M.R.; Hyman, J.; Clair, D.A.S.; Beelaman, R.B. Mapping of QTLs for lycopene and other fruit traits in a Lycopersicon esculentum × L. pimpinellifolium cross and comparison of QTLs across tomato species. Mol. Breed. 1999, 5, 283–299. [Google Scholar] [CrossRef]
- Stevens, M.A. Inheritance of tomato fruit quality components. Plant Breed. Rev. 1986, 4, 273–311. [Google Scholar]
- Xu, J.; Ranc, N.; Muños, S.; Rolland, S.; Bouchet, J.P.; Desplat, N.; Le Paslier, M.C.; Liang, Y.; Brunel, D.; Causse, M. Phenotypic diversity and association mapping for fruit quality traits in cultivated tomato and related species. Theor. Appl. Genet. 2013, 126, 567–581. [Google Scholar] [CrossRef]
- Fulton, T.M.; Grandillo, S.; Beck-Bunn, T.; Fridman, E.; Frampton, A.; Lopez, J.; Petiard, V.; Uhlig, J.; Zamir, D.; Tanksley, S.D. Advanced backcross QTL analysis of a Lycopersicon esculentum × Lycopersicon parviflorum cross. Theor. Appl. Genet. 2000, 100, 1025–1042. [Google Scholar] [CrossRef]
- Chetelat, R.T.; Klann, E.; DeVerna, J.W.; Yelle, S.; Bennett, A.B. Inheritance and genetic mapping of fruit sucrose accumulation in Lycopersicon chmielewskii. Plant J. 1993, 4, 643–650. [Google Scholar] [CrossRef]
- Levin, I.; Gilboa, N.; Yeselson, E.; Shen, S.; Schaffer, A. Fgr, a major locus that modulates the fructose to glucose ratio in mature tomato fruits. Theor. Appl. Genet. 2000, 100, 256–262. [Google Scholar] [CrossRef]
- Bernacchi, D.; Beck-Bunn, T.; Eshed, Y.; Lopez, J.; Petiard, V.; Uhlig, J.; Zamir, D.; Tanksley, S. Advanced backcross QTL analysis in tomato. I. Identification of QTLs for traits ofagronomic importance from Lycopersicon hirsutum. Theor. Appl. Genet. 1998, 97, 381–397. [Google Scholar] [CrossRef]
- Frary, A.; Fulton, T.M.; Zamir, D.; Tanksley, S.D. Advanced backcross QTL analysis of a Lycopersicon esculentum × L. pennellii cross and identification of possible orthologs in the Solanaceae. Theor. Appl. Genet. 2004, 108, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.D.; Fulton, T.M. Dissecting quantitative trait variation—Examples from the tomato. Euphytica 2007, 154, 365–370. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene | Forward Primer (5’→3’) | Reverse Primer (5’→3’) |
|---|---|---|---|
| Solyc06g065970.1 | Soly065970 | TGGTCCATGTGAGCCAAAGC | GCAATGAGCAACAAGGGGTT |
| Solyc06g065980.3 | Soly065980 | CCCGACCGATACCAAAAGCA | CCTCATTGTCCCCGTTTTGC |
| Solyc06g065990.1 | Soly065990 | TCCCCAGATCACCCAAATCC | GCCATCATGGTGGGAAGTGT |
| Solyc06g066000.3 | Soly066000 | AGACCAGGCTGCTGCAATTA | AGGGCAGCAATCTGAGAATCAA |
| Solyc06g066010.3 | Soly066010 | CGATGGGCGGAAGCTACTTA | CCTCTCGTTCTCCTCTCCGT |
| Solyc06g066020.3 | Soly066020 | GGAGATTGGATGCTTGCTGG | TTGTGCTTCCTCCACTCTTCC |
| Solyc06g066030.4 | Soly066030 | ATCAGATGCCAAACCTCGCA | TTGGCATGGGTGTCGCTTTA |
| Solyc06g066040.2 | Soly066040 | GGCAACCATGACTCAAGGGA | ATGCTTGATGACGGACCAGG |
| Solyc06g066050.3 | Soly066050 | GCACATCTAATCCGGCCCTT | TTGGGTTCTCGAACTGAGGC |
| Solyc06g066060.3 | Soly066060 | GGTTGGTAGGGGCATTGACA | GGCCTTTCGTCCCAAACCTA |
| Solyc06g066065.1 | Soly066065 | TGGTGCTATCTCTTGCTCGT | GTTTCAGGCAACTTGGCGTT |
| Solyc06g066070.1 | Soly066070 | CTCTCCGGCACTGTCATTGT | GGGCACCAATGGGTAAAACG |
| Solyc06g066080.1 | Soly066080 | ACTCACCAGCAAGAACCGAG | TTTGTGAGCACAAGGGGGTT |
| Solyc06g066090.3 | Soly066090 | TGTGGAAGAAGGAGTCGGGA | GCACCTCTCCTACGATCTGC |
| Solyc06g066100.3 | Soly066100 | GGACATGAGGGACGAACCAG | CCTCATTGTCCCCGTTTTGC |
| Solyc06g066110.1 | Soly066110 | CTAAATCGGCAACGGCGAAG | TCACCGCCGATATCAACTCG |
| Solyc06g066120.3 | Soly066120 | TGTCTCCAGGGGGTCTGATT | AGTCCATCTGTTGCTTGGCA |
| Solyc04g077020.2 | Tubulin | TGACGAAGTCAGGACAGGAA | CTGCATCTTCTTTGCCACTG |
| Gene ID | Gene Name | Chromosome Location | gDNA Length (bp) | Amino Acid Length (aa) | Molecular Weight (kDa) | Theoretical pI | Grand Average of Hydropathicity |
|---|---|---|---|---|---|---|---|
| Solyc06g065970.1 | Solyc06g065970.1, bifunctional inhibitor/lipid-transfer protein, AT2G45180 | chr6: 38992750-38993130 | 381 | 126 | 12.84 | 8.04 | 0.770 |
| Solyc06g065980.3 | Solyc06g065980.3, DNA/RNA-binding Alba-related, AT1G29250 | chr6: 38992750-38993130 | 3346 | 128 | 14.42 | 5.61 | −0.548 |
| Solyc06g065990.1 | Solyc06g065990.1, ATP synthase subunit, AT4G32260 | chr6: 38999236-38999907 | 672 | 223 | 24.41 | 5.79 | −0.079 |
| Solyc06g066000.3 | Solyc06g066000.3, ATP synthase subunit, AT4G32260 | chr6: 39002459-39003130 | 672 | 223 | 24.44 | 5.79 | −0.056 |
| Solyc06g066010.3 | Solyc06g066010.3, ATP synthase subunit, AT5G25340 | chr6: 39003882-39007076 | 3195 | 238 | 27.17 | 9.10 | −0.798 |
| Solyc06g066020.3 | Solyc06g066020.3, auxin-responsive protein IAA, AT2G46990 | chr6: 39015202-39017603 | 2402 | 242 | 28.21 | 5.21 | −0.943 |
| Solyc06g066030.4 | Solyc06g066030.4, targeting protein for Xklp2, AT3G23090 | chr6: 39024791-39030566 | 5776 | 476 | 51.46 | 9.37 | −0.936 |
| Solyc06g066040.2 | Solyc06g066040.2, protein of unknown function, N/A | chr6: 39035654-39038131 | 2478 | 185 | 20.63 | 6.18 | −0.064 |
| Solyc06g066050.3 | Solyc06g066050.3, protein of unknown function, AT5G25360 | chr6: 39039140-39045029 | 5890 | 173 | 19.22 | 8.17 | −0.384 |
| Solyc06g066060.3 | Solyc06g066060.3, ATP-dependent RNA helicase, AT5G11170 | chr6: 39049594-39056592 | 6999 | 410 | 46.50 | 6.41 | −0.159 |
| Solyc06g066065.1 | Solyc06g066065.1, protein of unknown function, N/A | chr6: 39056643-39057016 | 374 | 106 | 12.12 | 6.37 | 0.513 |
| Solyc06g066070.1 | Solyc06g066070.1, solute carrier family, AT2G25520 | chr6: 39058405-39059427 | 1023 | 340 | 38.00 | 9.31 | 0.631 |
| Solyc06g066080.1 | Solyc06g066080.1, DYW family of nucleic acid deaminases, AT1G47580 | chr6: 39060931-39062754 | 1824 | 607 | 68.49 | 7.79 | −0.037 |
| Solyc06g066090.3 | Solyc06g066090.3, ATPase family protein, AT2G25530 | chr6: 39064180-39075423 | 11,244 | 594 | 68.43 | 7.35 | −0.487 |
| Solyc06g066100.3 | Solyc06g066100.3, DNA/RNA-binding Alba-related, AT1G29250 | chr6: 39075900-39079511 | 3612 | 128 | 14.43 | 5.63 | −0.548 |
| Solyc06g066110.1 | Solyc06g066110.1, protein of unknown function, AT1G72510 | chr6: 39082573-39083025 | 453 | 150 | 16.66 | 8.67 | −0.426 |
| Solyc06g066120.3 | Solyc06g066120.3, endoglucanase, AT3G43860 | chr6: 39087035-39090249 | 3215 | 486 | 54.12 | 8.59 | −0.401 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.-R.; Tao, J.-P.; Song, L.-X.; Zhang, J.-Q.; Liu, H.; Zhu, W.-M.; Zhao, T.-M.; Xiong, A.-S. Identification of Genes of Molecular Marker TGS0892 on Chromosome 6 and Its Mechanism of Soluble Solids Metabolism in Tomato. Horticulturae 2023, 9, 52. https://doi.org/10.3390/horticulturae9010052
Zhang R-R, Tao J-P, Song L-X, Zhang J-Q, Liu H, Zhu W-M, Zhao T-M, Xiong A-S. Identification of Genes of Molecular Marker TGS0892 on Chromosome 6 and Its Mechanism of Soluble Solids Metabolism in Tomato. Horticulturae. 2023; 9(1):52. https://doi.org/10.3390/horticulturae9010052
Chicago/Turabian StyleZhang, Rong-Rong, Jian-Ping Tao, Liu-Xia Song, Jia-Qi Zhang, Hui Liu, Wei-Min Zhu, Tong-Min Zhao, and Ai-Sheng Xiong. 2023. "Identification of Genes of Molecular Marker TGS0892 on Chromosome 6 and Its Mechanism of Soluble Solids Metabolism in Tomato" Horticulturae 9, no. 1: 52. https://doi.org/10.3390/horticulturae9010052