
Proteomic Analysis of the Effect of Accelerated Ageing on Allium mongolicum Seeds
by
Xiaoqing Song
1,2, 
Zhongren Yang
1,2,
Dong Zhang
1,2,
Xiaoyan Zhang
1,2,
Fenglan Zhang
1,2,*,
Jiecai Liu
1,2 and
Chuanzong Yu
3,* 1
College of Horticultural and Plant Protection, Inner Mongolia Agricultural University, Hohhot 010011, China
2
Inner Mongolia Key Laboratory of Wild Peculiar Vegetable Germplasm Resource and Germplasm Enhancement, Inner Mongolia Agricultural University, Hohhot 010011, China
3
Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China
*
Authors to whom correspondence should be addressed.
Horticulturae 2023, 9(10), 1155; https://doi.org/10.3390/horticulturae9101155
Submission received: 25 July 2023
/
Revised: 16 October 2023
/
Accepted: 17 October 2023
/
Published: 21 October 2023
(This article belongs to the Section Propagation and Seeds)
Abstract
:Seed propagation is the most commonly used method for producing Allium mongolicum seeds. However, as storage time increases, the seeds may suffer from ageing to varying degrees. This study aims to investigate the proteome expression of A. mongolicum seeds with different levels of vigor. The findings of this study will provide evidence of protein expression levels that reveal the mechanism behind the ageing of A. mongolicum seeds. An integrated approach of tandem mass tag (TMT) labeling in conjunction with liquid chromatography–mass spectrometry (LC-MS/MS) was used to capture the differential protein expression profiles of A. mongolicum seeds under unaged (Control), lightly aged (LA) and severely aged (SA). In total, 4336 proteins were identified, of which 4318 were quantifiable. After accelerated ageing, 291 differentially expressed proteins (DEPs) were identified. Compared with Control, 36 DEPs (16 up-regulated and 20 down-regulated) were found in LA; and 139 DEPs (64 up-regulated and 75 down-regulated) in SA. Compared with LA, 116 DEPs (51 up-regulated and 65 down-regulated) were found in SA. A number of candidate proteins associated with seed ageing were screened, including ATP-dependent RNA helicase-like protein DB10 (Unigene0060546), β-D-glucan exohydolase (Unigene0008772), Histone H1.2 partial (Unigene0049278), defensin Ec-AMP-D2 (Unigene0042879), NAD(P)H dehydrogenase (quinone) FQR1 (Unigene0047137), glutathione S-transferase (Unigene0000863), oleosin S1-2 (Unigene0011542), probable inactive purple acid phosphatase 2 (Unigene0010352), glutathione S-transferase U8-like (Unigene0006910), and glutathione S-transferase U17-like (Unigene0051938). Additionally, this study was the first to determine the proteome of A. mongolicum seeds. These data provided references for further study of seed ageing mechanism of A. mongolicum.
1. Introduction
A. mongolicum is a perennial herbaceous plant known for its resistance to wind erosion, drought, and barrenness. Unfortunately, wild A. mongolicum is vulnerable to damage caused by climate change and overgrazing. In order to preserve this species, it is crucial to develop and use it through artificial cultivation and domestication. Understanding the seed characteristics of A. mongolicum is essential for successful artificial domestication and cultivation. Seed propagation is the most commonly used method for producing A. mongolicum seeds. However, with prolonged storage time, these seeds undergo varying degrees of senescence, resulting in reduced vigor, germination rate, and uneven seedling emergence. This can make it difficult to conserve germplasm resources for an extended period of time [1]. Previous research found that its seed life is up to 2–4 years [1], and the germination ability of A. mongolicum seeds during storage ageing was significantly and positively correlated with antioxidant enzyme activity [2]. However, proteomic studies on seed ageing in A. mongolicum have not been reported.
Seed ageing is a complex and irreversible process during seed development and storage [3]. The temperature and humidity during seed storage greatly impact seed vigor and the ageing process of seeds is often accelerated under conditions of high temperature and high humidity [4]. At the protein level, there are two main causes of seed senescence: on the one hand, changes in proteins in dry seeds, and on the other hand, the inability of low-activity seeds to express their normal proteome during germination. The key mechanisms that maintain seed viability include seed translocation capacity, seed storage mobilization and antioxidant efficiency [5]. In addition, some late embryogenesis abundant proteins (LEAs) and heat shock proteins (HSPs) play an important role in maintaining seed viability [6]. Similar to the loss of genetic integrity due to natural ageing, accelerated ageing may also have a negative impact on genetic integrity. High temperatures and high seed water content may lead to accelerated ageing, which mimics stress conditions leading to high respiration rates and reserve depletion resulting in seed loss of viability, usually within days or weeks [7]. As seeds age, seed viability decreases, leading to severe economic losses and loss of germplasm resources during seed marketing [8].
In recent years, proteomics has become a new technique to resolve seed characteristics e.g., seed germination [9], seed development [10,11], etc. Chu et al. used proteomics to resolve the variation in the viability of membrane-linked proteins associated with heat tolerance and germination vigor in lotus seeds [12]. Li et al. revealed by proteomics techniques that the differences in seed vigor in maize are due to the influence of growing sites on the seed development process [13]. Yan et al. analyzed the differentially expressed proteins (DEPs) of wheat seed embryos during germination mainly related to carbohydrate metabolism, protein metabolism, and amino acid metabolism using proteomic techniques [14]. Rajjou et al. found that loss of Arabidopsis seed vigor was associated with protein changes in dry seeds by proteomic analysis and the inability of low-viability seeds to produce a normal proteome during germination [5]. Li et al. found that calcium channel blockers affect radicle prominence during rice seed germination by proteomic analysis [15]. In conclusion, proteomics technology provides a powerful tool in seed ageing studies. Therefore, in this study, we utilized an integrated approach of TMT labeling and LC-MS/MS to capture the differential protein expression profiles of A. mongolicum seeds under unaged, lightly aged, and severely aged treatments. Our findings provide evidence of protein expression levels for revealing the ageing mechanism study of A. mongolicum seeds and for propagation renewal of A. mongolicum germplasm resources. In addition, this study will provide a theoretical basis for the mechanism of A. mongolicum seed ageing and lay the groundwork for the mining of related genes.
2. Materials and Methods
2.1. Materials
The test material used was A. mongolicum seeds harvested in 2019 (thousand grain weight of 2.32 g and initial moisture content of 5.18%). The A. mongolicum seeds were artificially aged using the high temperature and high humidity method according to the International Seed Inspection Protocol [16], with a treatment temperature of 50 °C, relative humidity (RH) of 100%, and treatment times of 0 h, 5 h, and 35 h, denoted as Control, LA, and SA, respectively. Finally, they were stored in a sealed bag in a 4 °C refrigerator for backup. Figure 1 shows the germination of the A. mongolicum seeds after different ageing, in which the values of all germination indexes in the A. mongolicum seeds almost tended to 0 at SA treatment, indicating that the A. mongolicum seeds almost reached the state of complete death [1]. For this reason, the materials of unaged (Control), lightly aged (LA), and severely aged (SA) treatment groups were selected for this experiment.
2.2. Methods
2.2.1. Total Protein Extraction from Allium mongolicum Seeds
The A. mongolicum seeds samples were ground into powder at low temperature in a grinder (purchased from Shanghai Jingxin/24-well, Shanghai, China), and quickly transferred to a liquid nitrogen precooled centrifuge tube. The seed protein was extracted using standard methods [17].
2.2.2. TMT Labeling and Peptide Separation
Proteolytic solution was added to each protein sample (100 μL), trypsin was added and 100 mM TEAB buffer, mixed well and then digested at 37 °C for 4 h. More trypsin was added and CaCl2 to digest overnight. Formic acid was then added to adjust the pH to less than 3, mixed well and centrifuged at 12,000× g for 5 min at room temperature. The supernatant was slowly passed through a C18 column for desalting, and the filtrate collected and vacuum freeze dried. To each 100 μL of 0.1 M TEAB buffer was added to dissolve the peptide and 41 µL of TMT labeling reagent, mixed well, and the reaction allowed to proceed at room temperature for 2 h. Afterwards, 8% ammonia was added to terminate the reaction, mixed with an equal volume of labeled sample, desalinated, and freeze-dried under vacuum [18].
The sample was fractionated on a Waters BEH C18 column (4.6 × 250 mm, 5 μm) using an L-3000 HPLC system with the elution gradient shown in Table 1. One tube was collected every minute and divided into 10 fractions, which were lyophilized and dissolved by adding 0.1% formic acid to each.
2.2.3. LC-MS/MS Analysis and Protein Identification
The liquid chromatography elution conditions and mass spectrometry parameters were set according to the method of Zhang [18]. The spray voltage was 2.3 kV, the temperature of the transfer tube was 320 °C, the resolution of the primary mass spectrum was set to 60,000 (200 m/z), the resolution of the secondary mass spectrum was set to 45,000 (200 m/z), the mass spectrum was acquired in data-dependent mode, the threshold intensity was set to 1.2 × 105, the maximum injection time was 86 ms and the dynamic exclusion range was set to 20 s. The raw mass spectrometry data were generated (.raw). The raw data obtained by LC-MS/MS were searched using the search engine Proteome Discoverer 2.4 (PD2.4, Thermo, New York, NY, USA) [19].
The database was queried with the A. mongolicum transcriptome database for comparison [20]. DEPs were defined according to p < 0.05 (Student t-test method) and the absolute value of the fold change (FC) of the protein was greater than 1.3 (up-regulated protein: FC > 1.3; down-regulated protein: FC < 0.77), and the protein was considered to have undergone a significant change if the above conditions were met [21].
2.2.4. Data Analysis
Data were analyzed using Microsoft Excel 2010 statistical packages. Proteomic analyses were performed in three biological replicates. Gene Ontology (GO, http://www.geneontology.org (accessed on 21 March 2022)) functional annotation and classification were performed for the proteins in the experiments using interproscan software [22]. Protein functions were classified by acquiring entries and sub-category terms, including molecular function (MF), biological process (BP) and cellular component (CC), by Fisher’s Exact Test. The GO entry was considered significantly enriched when p < 0.05 by Fisher’s Exact Test [23]. These proteins were further annotated using the Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.genome.jp/kegg/ (accessed on 21 March 2022)) functionally, and the pathway was considered significantly enriched by Fisher’s Exact Test at p < 0.05, proteins were significantly enriched [24]. Unsupervised principal component analysis (PCA) was performed by statistics function prcomp within R v3.5.0 (www.r-project.org (accessed on 21 March 2022)). Volcano map analysis, cluster heat map analysis and pathway enrichment analysis for GO and KEGG were performed for DEP [25].
3. Results
3.1. A General Overview of Protein Identification in Allium mongolicum Seeds under Accelerated Ageing
A total of 401,747 secondary spectra, 29,560 matched spectra, 19,076 identified peptides and 4336 identified proteins were obtained by MS analysis; the total number of quantifiable proteins for all samples was 4318 (Table 2). The results of principal component analysis showed that these nine samples were clearly distinguished into three major categories and are circled with different colors (Figure 2); each circle corresponded to the three replicate samples within the treatment, indicating that protein expression had clear biological reproducibility.
3.2. Quantification and Annotation of Differentially-Expressed Proteins (DEPs) in Allium mongolicum Seeds under Accelerated Ageing
To understand in detail the differences between different ageing treatments and controls, we compared LA and SA with Control (i.e., ‘LA vs. Control’, ‘SA vs. Control’, and ‘SA vs. LA’). Compared with Control, 36 DEPs (16 up-regulated and 20 down-regulated) were found in LA; and 139 DEPs (64 up-regulated and 75 down-regulated) in SA. Compared with LA, 116 DEPs (51 up-regulated and 65 down-regulated) were found in SA (Figure 3A–C). Venn diagram showed that 21 differential proteins were significantly expressed in LA and SA treatments compared with Control, of which, Histone H1.2 partial (Unigene0049278) and defensin Ec-AMP-D2 (Unigene0042879) were significantly up-regulated after LA, but down-regulated after SA. The expression trend of the other 19 differential proteins was consistent between LA and SA (Figure 3D).
3.3. GO Enrichment Analysis of DEPs in Allium mongolicum Seeds under Accelerated Ageing
To resolve the response mechanisms of A. mongolicum seeds to different degrees of accelerated ageing, we used GO analysis of the identified proteins (Figure 4, Table S1). We found that the number of proteins in seeds varied for each GO entry from LA to SA. Compared with Control, the difference of LA proteins are mainly involved in biological processes (BP) and molecular function (MF). The biological processes involved are mainly in response to wounding and vacuolar transport; the molecular functions involved are mainly carbon–sulfur lyase activity, enzyme inhibitor activity, acetyl-CoA carboxylase activity, serine-type endopeptidase inhibitor activity and O-methyltransferase activity (Figure 4A).
Compared with Control, the differential proteins in SA are assigned to various classifications and participate in the embryo development and ceramide metabolic process in terms of biological processes. From the molecular function analysis, it is mainly acid phosphatase activity, hydrolase activity acting on the ester bonds, endonuclease activity, and adenylosuccinate synthase activity. From the analysis of cell components, it mainly includes the monolayer-surrounded lipid storage body and intracellular non-membrane-bounded organelle (Figure 4B).
Compared with LA, the differential proteins in SA are involved in various categories. In biological processes, it is mainly involved in embryo development, cellular amide metabolic process and ceramide metabolic processes, translation, response to oxidative stress, etc.; the main molecular functions involved are carbon–sulfur lyase activity, heme binding, lyase activity and structural constituent of ribosome, adenylosuccinate synthase activity, phosphatidylserine decarboxylase activity, phosphatase activity, mRNA guanylyltransferase activity, and alanine-tRNA ligase activity. The cellular components involved mainly include intracellular non-membrane-bounded organelle and ribosome (Figure 4C).
3.4. KEGG Pathway Enrichment of DEPs in Allium mongolicum Seeds under Accelerated Ageing
To further clarify the metabolic and signal transduction pathways involved in ageing and development-related DEPs of A. mongolicum seeds under accelerated ageing and in Control, and to gain insight into the biological functions of DEPs, the DEPs obtained were subjected to KEGG metabolic pathway enrichment analysis (Figure 5, Table S2). Compared with Control, 18 metabolic pathways were involved in LA, with most proteins involved in biosynthesis of unsaturated fatty acids, glutathione metabolism, spliceosome, fatty acid biosynthesis, and phenylpropanoid biosynthesis (Figure 5A); 28 pathways were involved in SA, with most proteins involved in glutathione metabolism, ribosome, RNA transport, phenylpropanoid biosynthesis, starch and sucrose metabolism, and endocytosis (Figure 5B); compared with LA, 22 pathways were involved in SA, with most proteins involved in ribosome, spliceosome, glutathione metabolism, RNA transport, glycerophospholipid metabolism, glycerolipid metabolism, ribosome biogenesis in eukaryotes, RNA degradation, and oxidative phosphorylation (Figure 5C). From above it could be seen that the number of proteins involved in the same metabolic pathway increased with increasing accelerated ageing.
3.5. Analysis of Candidate Differential Proteins of Allium mongolicum Seeds after Accelerated Ageing
As shown in Figure 6, a total of 21 differential proteins were significantly enriched during the process from LA to SA, which were mainly divided into three categories (Table S3). The first category of proteins was significantly up-regulated expression after ageing treatment, such as ATP-dependent RNA helicase-like protein DB10 (Unigene0060546), β-D-glucan exohydolase (Unigene0008772), prostaglandin reductase-3 (Unigene0003044), aladin isoform X2 (Unigene0048082), trans-resveratrol di-O-methyltransferase-like isoform X2 (Unigene0042829); the second class of proteins is in significantly up-regulated after LA and significantly down-regulated afterSA, such as Histone H1.2 partial (Unigene0049278), defensin Ec-AMP-D2 (Unigene0042879); the third category of proteins is significantly up-regulated after ageing treatment down-regulated expression, such as aldehyde de-hydrogenase family 2 member C4-like (Unigene0049013), NAD(P)H dehydrogenase (quinone) FQR1 (Unigene0047137), tetraketide alpha-pyrone reductase 1-like isoform X3 (Unigene0045869), glutathione S-transferase (Unigene0000863), inhibitor of trypsin and hageman factor-like isoform X2 (Unigene0005112), isoflavone reductase-like protein (Unigene0053605), vacuolar protein sorting-associated protein 32 homolog 2 (Unigene0042292), polyadenylate-binding protein-interacting protein 11 (Unigene0047075), receptor protein kinase TMK1-like (Unigene0053916), oleosin S1-2 (Unigene0011542), mitogen-activated protein kinase kinase 1 isoform X1 (Unigene0045171), probable inactive purple acid phosphatase 2 (Unigene0010352), glutathione S-transferase U8-like (Unigene0006910), glutathione S-transferase U17-like (Unigene0051938).
4. Discussion
4.1. Effect of Accelerated Ageing Treatment on the Viability of Allium mongolicum Seeds
Seed is the most basic production material in agricultural production and occupies an important position in the whole process of agricultural production. Seed vigor is the main index of seed quality, which refers to the overall performance of seeds in germination and seedling emergence during activity intensity and characteristics. High vigor seeds germinate early, emerge cleanly and quickly, and have a strong ability to resist adverse conditions, with obvious growth advantages and production potential; low vigor seeds can germinate under suitable conditions, but germination is slow, and seedlings do not emerge cleanly or may not emerge at all under adverse environmental conditions. Seeds are affected by external environmental conditions at every stage from their formation and development to their maturation from the parent plant. Seed ageing occurs when the seed reaches its maximum physiological maturity and dry weight, when the seed has the highest vigor, and then decreases as the seed ages and progresses towards death.
Ageing seeds were found to germinate slowly and with little radicle growth compared to highly viable seeds [26]. In a previous study it was found that both viability and germination rate of A. mongolicum seeds showed a decreasing trend with increasing ageing time, and upon SA treatment the seeds almost completely lost viability [1]. Similar results were found for rice and redwood seeds [3,27]. This may be related to accelerated metabolism due to high temperature and humidity [27].
4.2. Effects of Accelerated Ageing Treatments on Starch and Sucrose Metabolism in Allium mongolicum Seeds
Seeds can lose viability and undergo genetic changes during long-term storage, and genetic integrity can be disrupted [28]. Seed degeneration is an inevitable and irreversible process that begins immediately after seed maturation [29]. During degeneration, the decline in vigor precedes the decline in germination and ends with the death of the seed [30]. In recent years, proteomic approaches for whole-plant expression analysis and protein identification have become very efficient [31]. Proteins are key players in cellular activities, and studying their altered abundance and modifications in different biological processes can greatly help in understanding their functions [32]. Stored proteins in seeds are not only an important source of amino acids during the early stages of germination, but are also important for energy production [33]. In Arabidopsis, proteins are also degraded early and rapidly resume metabolic activity upon seed uptake. After seed uptake, the three major nutrients (sugars, proteins and fats) begin to interconvert to provide energy and substrate for germination. The chemical and physiological changes in the embryo, endosperm and testa and their interactions contribute to the successful germination of seeds [34]. Starch is an important carbohydrate reserve and energy source. Several enzymes are involved in the metabolic activity of starch in germinating seeds [35]. β-Amylase and starch phosphorylase are responsible for the metabolism and degradation of starch [36]. β-Amylase converts starch to UDP-glucose and fructose, which are important for storage function and metabolism [37]. Sucrose synthase, a key class of enzymes in sucrose metabolism, is widely distributed in living organisms and mainly catalyzes the synthesis and degradation of sucrose, but also acts as a signaling molecule to regulate growth and developmental processes in plants [38]. It plays an important role in respiration, carbohydrate biosynthesis and utilization in plants [39]. In this experiment, β-glucosidase was significantly up-regulated in “starch and sucrose metabolism” after ageing treatment, and one protein encoding sucrose synthase (Unigene0063235) was significantly down-regulated after SA treatment, while five proteins encoding sucrose synthase (Unigene0001700, Unigene0063235) were significantly down-regulated after LA treatment. Unigene0001700, Unigene0053027, Unigene0063235, Unigene0050027, Unigene0050026) were up-regulated after LA treatment.
4.3. Effects of Accelerated Ageing Treatment on Proteins Associated with Energy Metabolism in Allium mongolicum Seeds
The ability to produce ATP largely reflects the metabolic activity of the seed, and the amount of ATP during seed germination correlates with seed viability. The ATPase family can act as a proton pump, which is the primary mechanism for lowering intracellular pH, altering membrane potential, and is used as a powerhouse for protein hydrolysis within seed cell tissues [40]. ATP synthesis is mediated by ATP synthases, and studies have shown that the expression of six ATP synthase subunits is down-regulated in maize seeds during senescence, resulting in a decrease in ATP [41]. In contrast, the expression of the ATP synthase subunit beta (Unigene0024535) was up-regulated after senescence treatment in this experiment, and the expression was significantly up-regulated at SA. This may be the result of a “vicious circle” between high metabolic activity and ATP production to maintain a low metabolic level or to defend against the external environment, resulting in a low ATP supply during the germination stage of aged seeds.
It has been shown that there are 29 purple acid phosphatases (PAPs) in the model plant Arabidopsis with different spatio-temporal expression patterns [42]. AtPAP15 may be involved in the mobilization of Pi reserves during seed and pollen germination [43]; AtPAP9 and AtPAP5 may be involved in defense against plant pathogens [44]; while AtPAP2 is targeted to plastids and mitochondria and is involved in plant growth [45]. It has been shown that embryo elongation growth requires cell expansion in specific regions of the radicle and hypocotyl during early seed germination, whereas cell division occurs mainly during seedling growth after emergence. In this experiment, PAP2, PAP22-like and PAP29 were down-regulated after ageing treatment, whereas PAP1 and PAP20 were up-regulated in expression. Among these, PAP2 was down-regulated in expression after ageing treatment, which may be the main reason for the slowing of radicle growth by ageing.
4.4. Effect of Accelerated Ageing Treatment on Glutathione Metabolism-Related Proteins in Allium mongolicum Seeds
Glutathione metabolism plays an important antioxidant role [46], and glutathione and glutathione disulfide are essential for the reduction of various peroxides [47]. Glutathione S-transferases play a direct role in reducing oxidative damage as well as toxic damage caused by foreign substances in response to external environmental stimuli [48,49]. Our results showed that after accelerated ageing, three proteins enriched in the glutathione pathway encoding glutathione S-transferase (glutathione S-transferase U8-like, glutathione S-transferase, glutathione S-transferase U17-like) were significantly down-regulated in expression and upon SA treatment, ascorbate peroxidase (Unigene0050520) was significantly down-regulated, possibly affecting the synthesis of ascorbate to dehydroascorbate and also responding to the reduced antioxidant capacity of low viability seeds. This suggests that the external environment (accelerated ageing) decreases the antioxidant capacity of seeds and reduces seed activity.
4.5. Effects of Accelerated Ageing Treatments on LEA and HSP Proteins in Allium mongolicum Seeds
The role of specific proteins in maintaining seed viability or longevity was verified. Two groups of late embryogenesis-rich (LEA) proteins (dehydrin/RAB group) were shown to contribute to Arabidopsis seed viability by controlled denaturation assays and proteomic analyses [3]. LEA proteins are highly hydrophilic and accumulate in large amounts during the final stages of seed maturation [50]. Mitochondria-specific late embryo development-rich (LEA) proteins found in seeds have been shown to play a key role in protecting these enzymes [51,52], stabilizing cell membranes and preventing seed dehydration [53,54]. In this experiment, five proteins encoding LEA (Unigene0016207, Unigene0016537, Unigene0013380, Unigene0037631, Unigene0048194) and eight proteins (Unigene0030390, Unigene0010414, Unigene0032477, Unigene0011984, Unigene0047667, Unigene0050372, Unigene0007117, Unigene0000326) were down-regulated, but the differences were not significant, whereas six proteins encoding LEA were down-regulated after SA treatment (Unigene0016537, Unigene0013380, Unigene0037631, Unigene0048194). Six proteins (Unigene0007117, Unigene0016537, Unigene0011984, Unigene0047667, Unigene0013380, Unigene0037631) were significantly down-regulated after SA treatment. This suggests that LEA function is reduced in SA and loses its ability to protect against enzymes.
Heat shock proteins are involved in a variety of cellular mechanisms, including regulation and prevention of protein folding, transport, activity regulation, degradation and irreversible protein aggregation [55], associated with seed development, protein translocation, reserve synthesis and mobilization [56], and cellular defense [57]. HSP proteins can play a crucial protective role in the process of antioxidant damage to seed proteins and can enhance the resistance of seeds to senescence [58]. hsp70 and hsp90 are important chaperone proteins in eukaryotic cells, heat shock protein 70 is involved in the plasma membrane fluidity of the cell membrane [59], heat shock protein 90-1-like is involved in stress response processes, mainly involved in the regulation of cell activity and apoptosis, and the expression of this protein has a positive effect on the stress response [60]. The results of this experiment showed that in LA, the expression of each heat shock protein was not significantly different from control, whereas in SA treatment, heat shock protein 70-2 (Unigene0053814) and heat shock protein 60, mitochondrial (Unigene0067445) were significantly up-regulated in expression, suggesting their essential role in protein biogenesis in response to accelerated ageing.
5. Conclusions
The present study focused on the problem that A. mongolicum seeds would be aged during storage and could not be preserved for a long time. There were 291 DEPs identifed after accelerated ageing treatments. The ageing-related proteins of A. mongolicum seeds were ATP-dependent RNA helicase-like protein DB10 (Unigene0060546), β-D-glucan exohydolase (Unigene0008772), Histone H1.2 partial (Unigene0049278), defensin Ec-AMP-D2 (Unigene0042879), NAD(P)H dehydrogenase (quinone) FQR1 (Unigene0047137), glutathione S-transferase (Unigene0000863), oleosin S1-2 (Unigene0011542), probable inactive purple acid phosphatase 2 (Unigene0010352), glutathione S-transferase U8-like (Unigene0006910), glutathione S-transferase U17-like (Unigene0051938), and the functional analysis of these proteins needs to be further investigated.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9101155/s1, Table S1: GO enrichment analysis of DEPs under aged A. mongolicum seeds of ‘T1 vs. CK’; Table S2: KEGG enrichment of differential proteins in ‘T1 vs CK’ group; Table S3: List of some important DEPs.
Author Contributions
X.S. helped with data analysis and wrote the manuscript. D.Z. and X.Z. performed the experiment. F.Z. and C.Y. made substantial contributions to conception and critically revised the manuscript. J.L. and Z.Y. designed the research. All authors have read and agreed to the published version of the manuscript.
Funding
This work received funding from the National Natural Science Foundation of China (Grant no. 31760081), Major Science and Technology Projects of China (Grant no. 2019ZD016), Basic Research Funds Project of Colleges and universities directly under Inner Mongolia Autonomous Region Seed industry revitalization leading talent project (Grant no. BR22-11-06), Major Demonstration Project of Science and Technology Innovation in the seed industry of the Autonomous Region (Grant no. 2022JBGS0028), Major Science and Technology Projects of Inner Mongolia Autonomous Region (Grant no. 2021ZD0001), Science and Technology Project of Inner Mongolia Autonomous Region (Grant no. 2021GG0084).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article [and its supplementary information files]. In addition, the datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, D.; Zhang, F.L.; Yang, Z.R.; Hao, L.Z.; Wang, J.Q.; Ren, X.Y. Effects of Ce3+ and La3+ soaking on seed vigor and physiological characteristics of artificially aged Allium mongolicum seeds. Acta Bot. Boreali-Occident. Sin. 2020, 40, 87–94. [Google Scholar]
- Chang, H.W.; Zhang, F.L.; Yang, Z.R.; Kong, D.J.; Zheng, Q.L.; Hao, L.Z. Physiological and biochemical responses of Allium mongolicum seeds to storage ageing. Plant Physiol. J. 2015, 51, 1075–1081. [Google Scholar]
- Liu, Y.; He, J.W.; Yan, Y.T.; Liu, A.M.; Zhang, H.Q. Comparative Transcriptomic Analysis of Two Rice (Oryza sativa L.) Male Sterile Line Seed Embryos Under Accelerated Aging. Plant Mol. Biol. Rep. 2020, 38, 282–293. [Google Scholar] [CrossRef]
- Lakshmi, C.J.; Jijeesh, C.M.; Seethalakshmi, K.K. Impact of accelerated aging process on seed quality and biochemical changes of Dendrocalamus sikkimensis Gamble. Acta Physiol. Plant 2021, 43, 34. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.C.; Maya, B.; Claudette, J.; Dominique, J. Proteome-wide characterization of seed aging in Arabidopsis: A comparison between artificial and natural aging protocols. Plant Physiol. 2008, 148, 620–641. [Google Scholar] [CrossRef]
- Kaur, H.; Petla, B.P.; Kamble, N.U.; Singh, A.; Rao, V.; Salvi, P.; Ghosh, S.; Majee, M. Differentially expressed seed aging responsive heat shock protein OsHSP18.2 implicates in seed vigor, longevity and improves germination and seedling establishment under abiotic stress. Front. Plant Sci. 2015, 6, 713–725. [Google Scholar] [CrossRef]
- Moraes, C.E.; Lopes, J.C.; Farias, C.C.M.; Maciel, K.S. Physiological quality of Tabernaemontana fuchsiaefolia A. DC seeds due to the accelerated aging test. Cienc. Florest. 2016, 26, 213–223. [Google Scholar] [CrossRef]
- Chen, B.X.; Fu, H.; Gao, J.D.; Zhang, Y.X.; Huang, W.J.; Chen, Z.J.; Zhang, Q.; Yan, S.J.; Liu, J. Identification of Metabolomic Biomarkers of Seed Vigor and Aging in Hybrid Rice. Rice 2022, 15, 7. [Google Scholar] [CrossRef]
- Liu, D.M.; Han, C.X.; Deng, X.; Liu, Y.; Liu, N.N.; Yan, Y.M. Integrated physiological and proteomic analysis of embryo and endosperm reveals central salt stress response proteins during seed germination of winter wheat cultivar Zhengmai 366. BMC Plant Biol. 2019, 19, 406. [Google Scholar] [CrossRef]
- Chatelain, E.; Hundertmark, M.; Leprince, O.; Le, G.S.; Satour, P.; Deligny-Penninck, S.; Rogniaux, H.; Buitink, J. Temporal profiling of the heat-stable proteome during late maturation of Medicago truncatula seeds identifies a restricted subset of late embryogenesis abundant proteins associated with longevity. Plant Cell Environ. 2012, 35, 1440–1455. [Google Scholar] [CrossRef]
- Nogueira, F.C.S.; Palmisano, G.; Schwammle, V.; Soares, E.L.; Soares, A.A.; Roepstorff, P.; Domont, G.B.; Wang, W.Q.; Liu, S.J.; Song, S.Q.; et al. Proteomics of seed development, desiccation tolerance, germination and vigor. Plant Physiol. Biochem. 2015, 86, 1–15. [Google Scholar]
- Chu, P.; Chen, H.H.; Zhou, Y.L.; Li, Y.; Ding, Y.; Jiang, L.W.; Tsang Edward, W.T.; Wu, K.Q.; Huang, S.Z. Proteomic and functional analyses of Nelumbo nucifera annexins involved in seed thermotolerance and germination vigor. Planta 2012, 235, 1271–1288. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qu, H.B.; Zhu, P.Y.; Su, K.M.; Zhang, C.Q. Comparative Proteomics Reveals the Mechanisms Underlying Variations in Seed Vigor Based on Maize (Zea mays L.) Ear Positions. Plant Mol. Biol. Rep. 2018, 36, 738–749. [Google Scholar] [CrossRef]
- Yan, M.K.; Zheng, L.; Li, B.J.; Shen, R.F.; Lan, P. Comparative proteomics reveals new insights into the endosperm responses to drought, salinity and submergence in germinating wheat seeds. Plant Mol. Biol. 2021, 105, 287–302. [Google Scholar] [CrossRef]
- Li, M.; Chen, X.; He, D.L.; Yang, P.F. Proteomic analysis reveals that calcium channel blockers affect radicle protrusion during rice seed germination. Plant Plant Growth Regul. 2020, 90, 393–407. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, D.; Zhang, W.L.; Duan, N.B.; Li, Q.; Yan, T.J.; Dai, S.; Ding, H.F. Effect of artificial aging on soybean seed vigor and ascorbate-glutathione cycle in mitochondria. Plant Physiol. J. 2016, 52, 543–550. [Google Scholar]
- Niu, L.; Zhang, H.; Wu, Z.; Wang, Y.; Liu, H.; Wu, X.; Wang, W.; Thierry, C. Modifed TCA/acetone precipitation of plant proteins for proteomic analysis. PLoS ONE 2018, 13, e0202238. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, T.; Zhang, Z. Integrated Proteogenomic Characterization of Human High-Grade Serous Ovarian Cancer. Cell 2016, 166, 755. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, Z.R.; Song, X.Q.; Zhang, F.L.; Liu, Y. TMT-based proteomic analysis of liquorice root in response to drought stress. BMC Genom. 2022, 23, 524. [Google Scholar] [CrossRef]
- Guo, F. Study on the Response Mechanism of Allium mongolicum Seeds to Storage Ageing. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2017. [Google Scholar]
- Moulder, R.; Lönnberg, T.; Elo, L.L.; Filén, J.J.; Rainio, E.; Corthals, G.; Oresic, M.; Nyman, T.A.; Aittokallio, T.; Lahesmaa, R. Quantitative proteomics analysis of the nuclear fraction of human CD4+ cells in the early phases of IL-4-induced Th2 differentiation. Mol. Cell Proteom. 2010, 9, 1937–1953. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.Z.; McAnulla, C. InterProScan 5: Genome-scale protein function classifcation. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Ringwald, M.; Rubin, G.M.; Sherlock, G. Gene ontology: Tool for the unifcation of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Sveinsdottir, H.; Zhu, Y.Y.; Schubert, S. Seed ageing-induced inhibition of germination and post-germination root growth is related to lower activity of plasma membrane H+-ATPase in maize roots. J. Plant Physiol. 2009, 166, 128–135. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, Y.F.; Liu, X.; Jiang, Y.; Deng, S.M.; Ai, X.R.; Deng, Z.J. Effect of artificially accelerated aging on the vigor of Metasequoia glyptostroboides seeds. J. For. Res. 2020, 31, 769–779. [Google Scholar] [CrossRef]
- Van Treuren, R.; de Groot, E.C.; van Hintum, J.L. Preservation of seed viability during 25 years of storage under standard genebank conditions. Genet. Resour. Crop Evol. 2013, 60, 1407–1421. [Google Scholar] [CrossRef]
- McDonald, M.B. Orthodox seed deterioration and its repair. In Handbook of Seed Physiology: Applications to Agriculture; Benech-Arnold, R.L., Sanchez, R.A., Eds.; The Haworth Press: New York, NY, USA, 2004; pp. 273–304. [Google Scholar]
- Hilhorst, H.W.M.; Toorop, P. Review on dormancy, germinability, and germination in crop and weed seeds. Adv. Agron. 1997, 61, 111–165. [Google Scholar]
- Qin, J.; Zhang, J.N.; Liu, D.; Yin, C.C.; Wang, F.M.; Chen, P.Y.; Chen, H.; Ma, J.B.; Zhang, B.; Xu, J.; et al. iTRAQ-based analysis of developmental dynamics in the soybean leaf proteome reveals pathways associated with leaf photosynthetic rate. Mol. Genet. Genom. 2016, 291, 1595. [Google Scholar] [CrossRef]
- Liu, Y.H.; Lu, S.; Liu, K.F.; Wang, S.; Huang, L.Q.; Guo, L.P. Proteomics: A powerful tool to study plant responses to biotic stress. Plant Methods 2019, 15, 135. [Google Scholar] [CrossRef]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Nomaguchi, M.; Okumoto, N.; Kaneko, Y.; Matsushima, H.; Morohashi, Y. Temporal and spatial pattern of the biochemical activation of the endosperm during and following imbibition of tomato seeds. Physiol. Plant. 1998, 102, 236–242. [Google Scholar] [CrossRef]
- Li, S.F.; Wei, X.J.; Ren, Y.L.; Qiu, J.H.; Jiao, G.A.; Guo, X.P.; Tang, S.Q.; Wan, J.M.; Hu, P.S. OsBT1 encodes an ADP-glucose transporter involved in starch synthesis and compound granule formation in rice endosperm. Sci. Rep. 2017, 7, 40124. [Google Scholar] [CrossRef]
- Smith, A.M.; Zeeman, S.C.; Smith, S.M. Starch degradation. Annu. Rev. Plant Biol. 2005, 56, 73–98. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Zhen, S.M.; Cheng, Z.W.; Cao, H.; Ge, P.; Yan, Y.M. Proteomic analysis reveals key proteins and phosphoproteins upon seed germination of wheat (Triticum aestivum L.). Front. Plant Sci. 2015, 6, 1017–1025. [Google Scholar] [CrossRef]
- Ruan, Y.L. Signaling role of sucrose metabolism in development. Mol. Plant 2012, 5, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Baier, M.C.; Keck, M.; Gdde, V.; Niehaus, K.; Kuster, H.; Hohnjec, N. Knockdown of the symbiotic sucrose synthase Mt SucS1 affects arbuscule maturation and maintenance in mycorrhizal roots of Medicago truncatula. Plant Physiol. 2010, 152, 1000–1014. [Google Scholar] [CrossRef]
- Wang, W.Q.; Ye, J.Q.; Rogowska-Wrzesinska, A.; Wojdyla-Katarzyna, I.; Jensen, O.N.; Møller, I.M.; Song, S.Q. Proteomic comparison between maturation drying and prematurely imposed drying of Zea mays seeds reveals a potential role of maturation drying in preparing proteins for seed germination, seedling vigor, and pathogen resistance. J. Proteome Res. 2013, 13, 606–626. [Google Scholar] [CrossRef]
- Xin, X.; Lin, X.H.; Zhou, Y.C.; Chen, X.L.; Liu, X.; Lu, X.X. Proteome analysis of maize seeds: The effect of artificial ageing. Physiol. Plant. 2011, 143, 126–138. [Google Scholar] [CrossRef]
- Zhu, H.F.; Qian, W.Q.; Lu, X.Z.; Li, D.P.; Liu, X.; Liu, K.F.; Wang, D.W. Expression patterns of purple acid phosphatase genes in Arabidopsis organs and functional analysis of AtPAP23 predominantly transcribed in flower. Plant Mol. Biol. 2005, 59, 581–594. [Google Scholar] [CrossRef]
- Kuang, R.B.; Chan, K.H.; Yeung, E.; Lim, B.L. Molecular and biochemical characterization of AtPAP15, a purple acid phosphatase with phytase activity, in Arabidopsis. Plant Physiol. 2009, 151, 199–209. [Google Scholar] [CrossRef]
- Zamani, K.; Lohrasebi, T.; Sabet, M.S.; Malboobi, M.A.; Mousavi, A. Expression pattern and subcellular localization of Arabidopsis purple acid phosphatase AtPAP9. Gene Expr. Patterns 2014, 14, 9–18. [Google Scholar] [CrossRef]
- Sun, F.; Suen, P.K.; Zhang, Y.J.; Liang, C.; Carrie, C.; Whelan, J.; Ward, J.L.; Hawkins, N.D.; Jiang, L.W.; Rajjou, L.; et al. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar]
- Song, W.; Zhou, F.K.; Shan, C.H.; Zhang, Q.; Ning, M.; Liu, X.M.; Zhao, X.X.; Cai, W.C.; Yang, X.Q.; Hao, G.F.; et al. Identification of glutathione S-transferase genes in Hami melon (Cucumis melo var. saccharinus) and their expression analysis under cold stress. Front. Plant Sci. 2021, 12, 672017. [Google Scholar] [CrossRef]
- Milla, M.A.; Maurer, A.; Huete, A.R.; Gustafson, J.P. Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways. Plant J. 2003, 36, 602–615. [Google Scholar] [CrossRef]
- Moons, A. Regulatory and functional interactions of plant growth regulators and plant glutathione S-transferases (GSTs). Vitam. Horm. 2005, 72, 155. [Google Scholar]
- Xu, J.; Tian, Y.S.; Xing, X.J.; Peng, R.H.; Zhu, B.; Gao, J.J.; Yao, H. Over-expression of AtGSTUl9 provides tolerance to salt, drought and methyl viologen stresses in Arabidopsis. Physiol. Plant. 2016, 156, 164–175. [Google Scholar] [CrossRef]
- Espelund, M.; Saeboe-Larssen, S.; Hughes, D.W.; Galan, G.A.; Larsen, F.; Jakobsen, K.S. Late embryogenesis-abundant genes encoding proteins with different numbers of hydrophilic repeats are regulated differentially by abscisic acid and osmotic stress. Plant J. 1992, 2, 241–252. [Google Scholar] [CrossRef]
- Grelet, J.; Benamar, A.; Teyssier, E.; Avelange-Macherel, M.H.; Grunwald, D.; Macherel, D. Identification in pea seed mitochondria of a late-embryogenesis abundant protein able to protect enzymes from drying. Plant Physiol. 2005, 137, 157–167. [Google Scholar] [CrossRef]
- Cao, J.; Li, X. Identification and phylogenetic analysis of late embryogenesis abundant proteins family in tomato (Solanum lycopersicum). Planta 2015, 241, 757–772. [Google Scholar] [CrossRef]
- Pedrosa, A.M.; Martins, C.D.P.S.; Gonçalves, L.P.; Costa, M.G.C. Late embryogenesis abundant (LEA) constitutes a large and diverse family of proteins involved in development and abiotic stress responses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Juszczak, I.; Bartels, D. LEA gene expression, RNA stability and pigment accumulation in three closely related linderniaceae species differing in desiccation tolerance. Plant Sci. 2017, 255, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Li, H.M. Arabidopsis stromal 70-kD heat shock proteins are essential for plant development and important for thermos tolerance of germinating seeds. Plant Physiol. 2008, 146, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Forward, B.S.; Osusky, M.; Misra, S. The Douglas-fir BiP promoter is functional in Arabidopsis and responds to wounding. Planta 2002, 215, 569–576. [Google Scholar] [CrossRef]
- Carpentier, S.C.; Witters, E.; Laukens, K.; Deckers, P.; Swennen, R.; Panis, B. Preparation of protein extracts from recalcitrant plant tissues:an evaluation of different methods for two-dimensional gel electrophoresis analysis. Proteomics 2005, 5, 2497–2507. [Google Scholar] [CrossRef]
- Shen-Miller, J.; Lindner, P.; Xie, Y.; Villa, S.; Wooding, K.; Clarke, S.G.; Loo, R.R.O.; Loo, J.A. Thermal–Stable Proteins of Fruit of Long–Living Sacred Lotus Nelumbo nucifera Gaertn var. China Antique. Trop. Plant Biol. 2013, 6, 69–84. [Google Scholar] [CrossRef]
- Maikova, A.; Zalutskaya, Z.; Lapina, T.; Ermilova, E. The HSP70 chaperone machines of Chlamydomonas are induced by cold stress. J. Plant Physiol. 2016, 204, 85–91. [Google Scholar] [CrossRef]
- Zhang, X.H.; Hong, W.; Tang, S.; Li, Q.N.; Bao, E.D. Apoptosis in response to heat stress is positively associated with heat-shock protein 90 expression in chicken myocardial cells in vitro. J. Vet. Sci. 2017, 18, 129–140. [Google Scholar] [CrossRef]
Figure 1.
Embryonic root growth of Allium mongolicum seeds after different ageing treatments. Notes: Control, LA, and SA represented unaged (Control), lightly aged (LA) and severely aged, respectively.
Figure 1.
Embryonic root growth of Allium mongolicum seeds after different ageing treatments. Notes: Control, LA, and SA represented unaged (Control), lightly aged (LA) and severely aged, respectively.
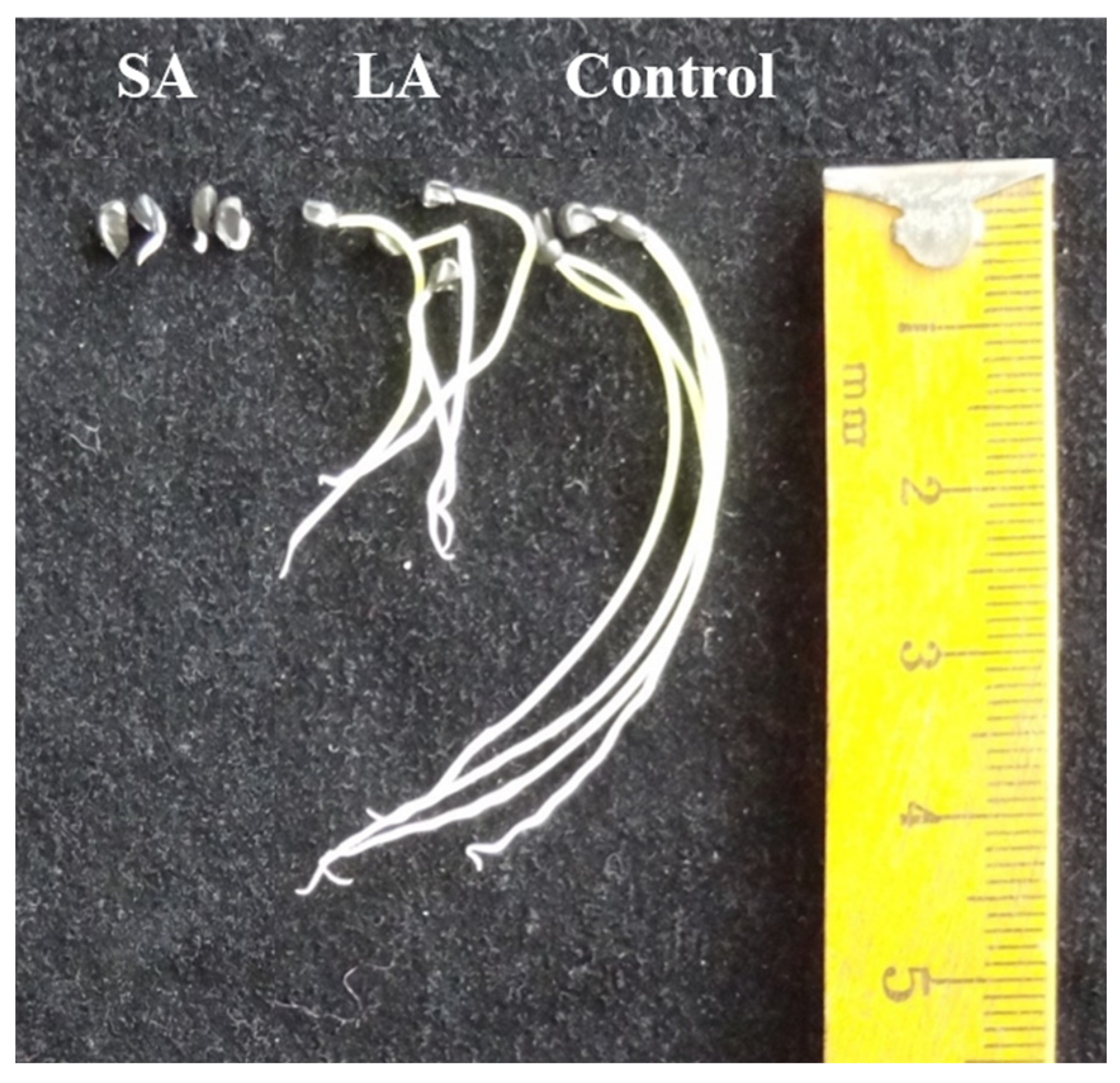
Figure 2.
PCA analysis. Note: The horizontal coordinate PC1 and the vertical coordinate PC2 in the fgure indicate the scores of the first and second ranked principal components, respectively. The scatter color indicates the experimental grouping of the sample.
Figure 2.
PCA analysis. Note: The horizontal coordinate PC1 and the vertical coordinate PC2 in the fgure indicate the scores of the first and second ranked principal components, respectively. The scatter color indicates the experimental grouping of the sample.

Figure 3.
Volcano plot and Venn diagram of differential protein expression in Allium mongolicum seeds after different levels. Note: (A–C) The horizontal co-ordinates indicate the difference multiplicity of differential proteins (log2 value), the vertical axis indicates the p value (−log10 value), black represents proteins that are not differentially expressed compared with the control; red represents up-regulated proteins; and green represents down-regulated proteins; (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA). (D) Each circle in the graph represents a comparison group, and the numbers in the overlapping part of the circles represent the number of differentially expressed proteins shared between the comparison groups; the numbers in the non-overlapping part represent the number of differentially expressed proteins that are specific to that group.
Figure 3.
Volcano plot and Venn diagram of differential protein expression in Allium mongolicum seeds after different levels. Note: (A–C) The horizontal co-ordinates indicate the difference multiplicity of differential proteins (log2 value), the vertical axis indicates the p value (−log10 value), black represents proteins that are not differentially expressed compared with the control; red represents up-regulated proteins; and green represents down-regulated proteins; (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA). (D) Each circle in the graph represents a comparison group, and the numbers in the overlapping part of the circles represent the number of differentially expressed proteins shared between the comparison groups; the numbers in the non-overlapping part represent the number of differentially expressed proteins that are specific to that group.
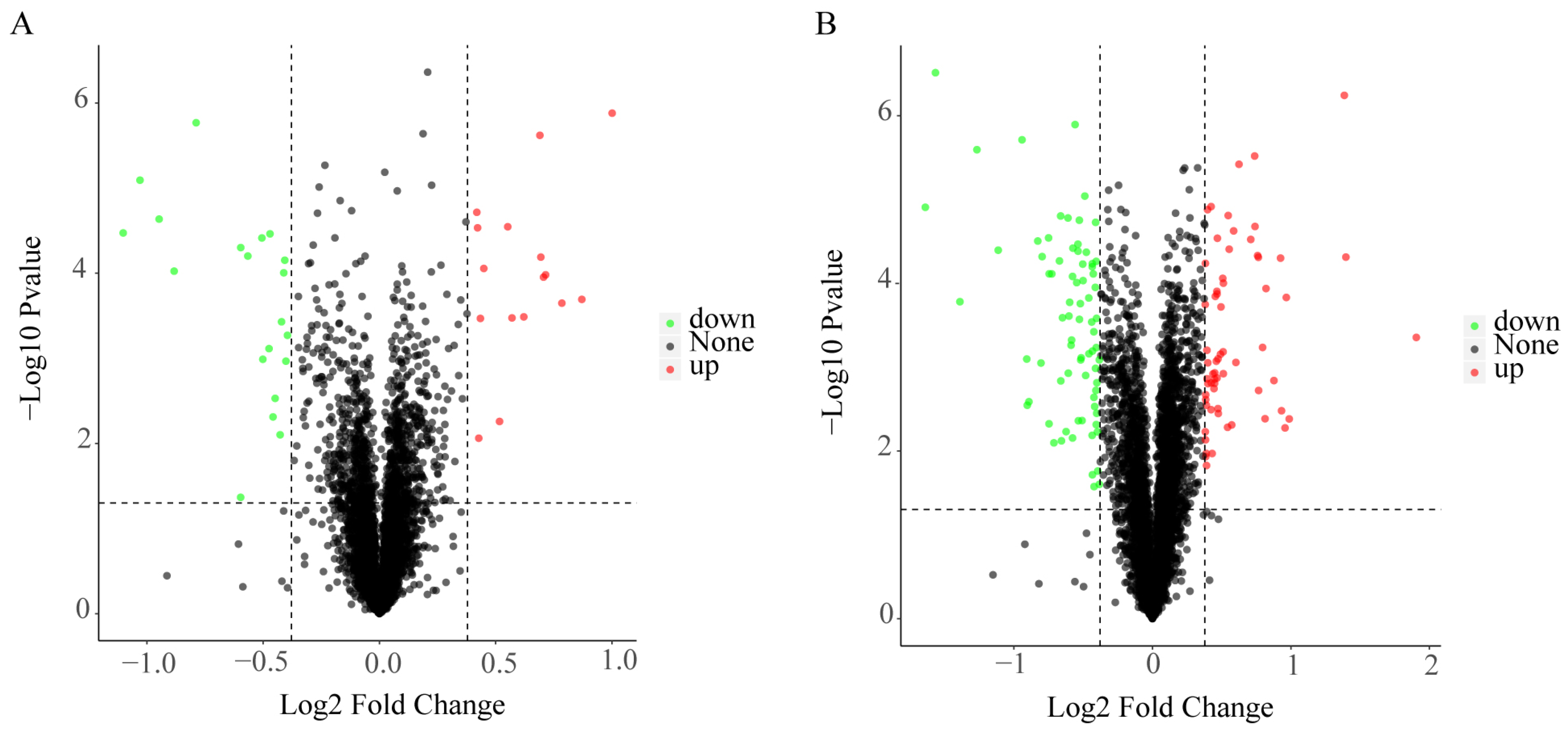

Figure 4.
GO enrichment analysis of DEPs under aged Allium mongolicum seeds of different comparison groups. Note: (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA).
Figure 4.
GO enrichment analysis of DEPs under aged Allium mongolicum seeds of different comparison groups. Note: (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA).
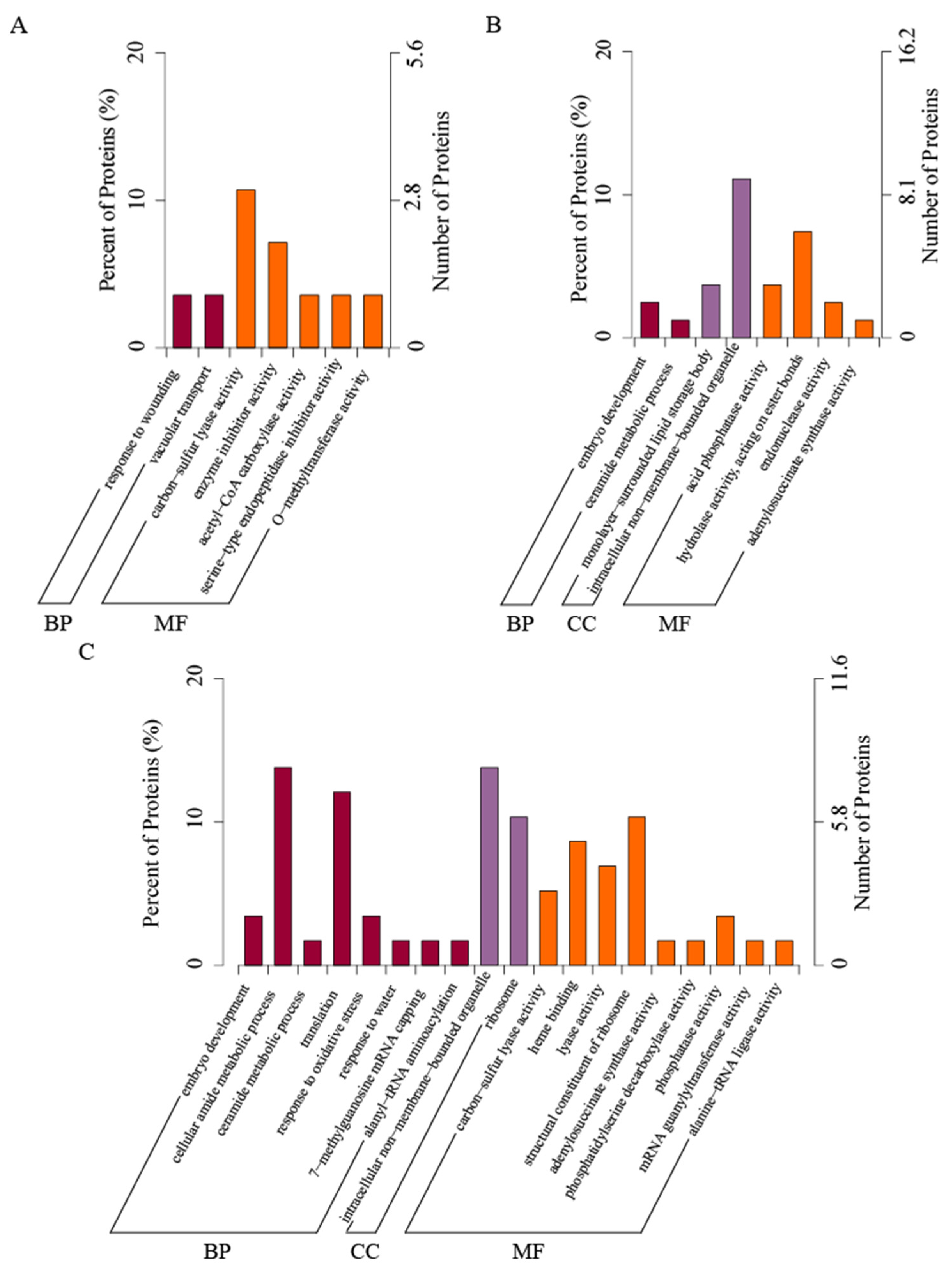
Figure 5.
Bubbles representing different comparison groups of Allium mongolicum seeds following differential protein KEGG enrichment. Note: (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA). The horizontal coordinate in the graph is the ratio of the number of differential proteins in the corresponding pathway to the number of total proteins identified in that pathway; the larger the value, the higher the differential protein enrichment in that pathway. The color of the dot represents the p value of the hypergeometric test; the colors range from blue to red, the redder the color, the smaller the value and the more reliable and statistically significant the test is. The size of the dots represents the number of differential proteins in the corresponding pathway, the larger the dot, the more differential proteins are present in the pathway.
Figure 5.
Bubbles representing different comparison groups of Allium mongolicum seeds following differential protein KEGG enrichment. Note: (A) compared LA with Control (LA vs. Control); (B) compared SA with Control (SA vs. Control); (C) compared SA with LA (SA vs. LA). The horizontal coordinate in the graph is the ratio of the number of differential proteins in the corresponding pathway to the number of total proteins identified in that pathway; the larger the value, the higher the differential protein enrichment in that pathway. The color of the dot represents the p value of the hypergeometric test; the colors range from blue to red, the redder the color, the smaller the value and the more reliable and statistically significant the test is. The size of the dots represents the number of differential proteins in the corresponding pathway, the larger the dot, the more differential proteins are present in the pathway.

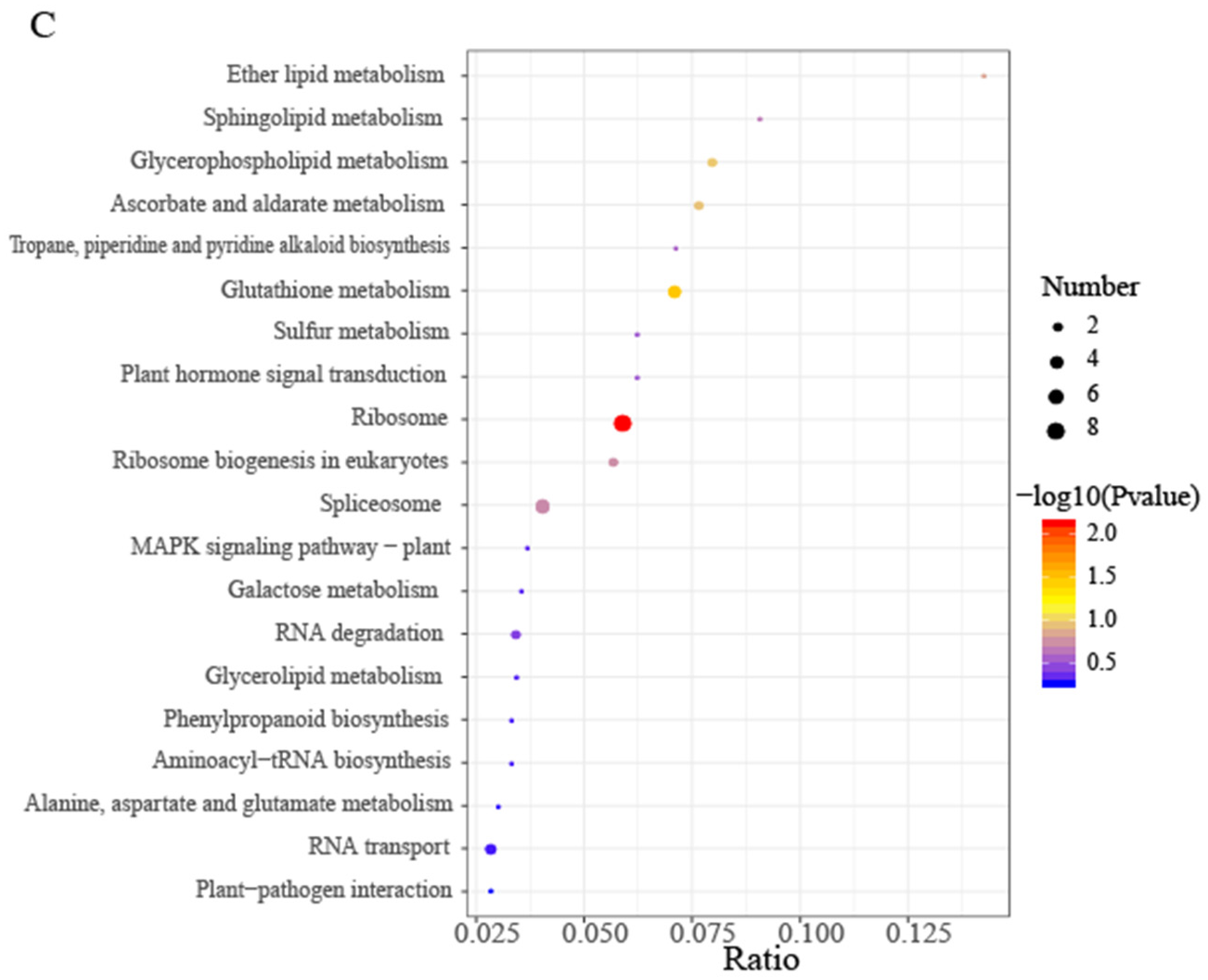
Figure 6.
Heat map of clustering of co-significantly estimated differential proteins.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Peptide fraction separation liquid chromatography elution gradient table.
| Time (min) | Flow Rate (nL/min) | Mobile Phase A (%) | Mobile Phase B (%) |
|---|---|---|---|
| 0 | 1 | 97 | 3 |
| 10 | 1 | 95 | 5 |
| 30 | 1 | 80 | 20 |
| 48 | 1 | 60 | 40 |
| 50 | 1 | 50 | 50 |
| 53 | 1 | 30 | 70 |
| 54 | 1 | 0 | 100 |
Note: Liquid A (2% acetonitrile, 98% water, ammonia adjusted to pH = 10) and Liquid B (98% acetonitrile, 2% water).
Table 2.
Protein Identification Information Statistical Summary.
| Total Spectra | Matched Spectrum | Peptide | Identified Protein | ALL |
|---|---|---|---|---|
| 401,747 | 29,560 | 19,076 | 4336 | 4318 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, X.; Yang, Z.; Zhang, D.; Zhang, X.; Zhang, F.; Liu, J.; Yu, C. Proteomic Analysis of the Effect of Accelerated Ageing on Allium mongolicum Seeds. Horticulturae 2023, 9, 1155. https://doi.org/10.3390/horticulturae9101155
AMA Style
Song X, Yang Z, Zhang D, Zhang X, Zhang F, Liu J, Yu C. Proteomic Analysis of the Effect of Accelerated Ageing on Allium mongolicum Seeds. Horticulturae. 2023; 9(10):1155. https://doi.org/10.3390/horticulturae9101155
Chicago/Turabian StyleSong, Xiaoqing, Zhongren Yang, Dong Zhang, Xiaoyan Zhang, Fenglan Zhang, Jiecai Liu, and Chuanzong Yu. 2023. "Proteomic Analysis of the Effect of Accelerated Ageing on Allium mongolicum Seeds" Horticulturae 9, no. 10: 1155. https://doi.org/10.3390/horticulturae9101155
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.