
Effects of Temperature on Growth and Isoprene Metabolism Pathway in Eucommia ulmoides Oliv

1
Key Laboratory of Plant Resource Conservation and Germplasm Innovation in Mountainous Region (Ministry of Education), College of Life Sciences, Institute of Agro-Bioengineering, Guizhou University, Guiyang 550025, China
2
Plant Conservation Technology Center, Guizhou Provincial Key Laboratory of Agricultural Biotechnology, Guizhou Academy of Agricultural Sciences, Guiyang 550006, China
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(12), 1298; https://doi.org/10.3390/horticulturae9121298
Submission received: 24 October 2023
/
Revised: 27 November 2023
/
Accepted: 28 November 2023
/
Published: 1 December 2023
(This article belongs to the Special Issue Advancements in Enhancing Environmental Stress Tolerance of Specialty Crops in Horticulture)
Abstract
:This study aimed to explore the influence of temperature on the growth and isoprene metabolism pathways of Eucommia ulmoides seedlings. Growth indices and levels of rubber and total triterpenoids in leaves were assessed under three temperature treatments: low (15 °C/10 °C), control (25 °C/20 °C), and high (35 °C/30 °C). High-throughput sequencing identified 2309 differentially expressed genes, of which 1608 were upregulated and 701 were downregulated. After 40 days, leaf length, leaf width, and plant height were significantly lower in low- and high-temperature treatments compared with the control. Specific leaf weight was higher in LT-treated leaves. Chlorophyll a and b contents were 1.372 mg.g−1 and 0.594 mg.g−1, respectively, in control leaves, followed by low- and high-temperature treatments. Carotenoid content was the highest in LT treatment. The rubber content of LT and HT groups significantly decreased by 16.5% and 38%, respectively, compared with that of the control group. Total triterpene content was the lowest in control leaves at 1.02%, which was 30% and 20% less than that in low- and high-temperature treatments, respectively. This study provides insights into the efficient cultivation of E. ulmoides and the regulatory network of secondary metabolic pathways.
1. Introduction
The isoprene metabolic pathway is important for the synthesis of secondary metabolites in plants. The intermediate isoprenyl pyrophosphate is generated by two pathways: the mevalonate (MVA) pathway, which occurs in the cytoplasm, and the pyruvate/phosphoglyceraldehyde (MEP) pathway, which occurs in the plastids [1]. According to a previous study, the isopentenyl pyrophosphate (IPP) that is used as a precursor for sesquiterpene and triterpene synthesis is produced by the MVA pathway [2], whereas IPP that is used as a precursor for monoterpene, diterpene, and carotenoid synthesis is produced by the MEP pathway [3,4]. Based on the number of isoprene residues, terpenes can be classified into monoterpenes, sesquiterpenes, diterpenes, and triterpenes [5]. Terpenoids play a crucial role in regulating plant growth, development, and environmental adaptation [6]. When environmental temperatures increase, plants adapt by increasing the production and emission of isoprene [7,8]. Geranyl pyrophosphate (GPP), farnesyl pyrophosphate (FPP), and geranylgeranyl pyrophosphate (GGPP) are direct precursors used in the synthesis of monoterpenes, sesquiterpenes, and diterpenes, respectively, using terpene synthase (TPS) [9]. The TPS family is categorized into seven subfamilies: TPS-a, TPS-b, TPS-c, TPS-d, TPS-e/f, TPS-g, and TPS-h [10]. This family is classified into monoterpene synthases, sesquiterpene synthases, diterpene synthases, and triterpene synthases based on terpene products. Isoprene metabolites in plants encompass artemisinin, gibberellins [11], abscisic acid, chlorophyll, terpenoids, sterols, perfumes, flavors, and rubbers [12].
Eucommia ulmoides (E. ulmoides), a tertiary relict tree species unique to China, is not only a precious traditional medicinal herb but also a high-quality source of rubber [13,14,15]. The pericarp, bark, leaves, and roots of E. ulmoides are rich in rubber filaments. The rubber content in the fruit ranges from 12% to 17%, and the leaf rubber content is between 2% and 5% [15]. E. ulmoides rubber is a natural polymer compound with the molecular structure of trans-polyisoprene. It possesses a shape memory function [16], and a unique “rubber-plastic” duality [17]. This property makes it suitable for development into high-elasticity rubber materials, low-temperature plasticity materials, and thermoelastic memory materials. Accordingly, E. ulmoides is widely used in the rubber industry. For instance, it is blended with chloroprene rubber or polypropylene to enhance the characteristics of the composite materials and to prepare shape memory materials [18,19,20]. Rubber is one of the end products of the isoprene metabolic pathway in rubber-producing plants. IPP is a precursor for terpenoids, and rubber synthesis, with rubber synthesis primarily sourced from the MVA pathway and terpenoids, compounds from the MEP pathway [21]. In the MVA pathway, HMGR determines the direction of “carbon flow.” When HMGR activity increases, sesquiterpene products increase [22]. However, HMGR expression is affected by environmental factors, such as light, temperature, and humidity [4,23,24].
Global temperature extremes are becoming more frequent, impacting plant health via cold stress (0–18 °C), freezing (<0 °C), and heat stress (>25 °C), ultimately affecting crop production [25,26]. The growth, development, and accumulation of secondary metabolites in E. ulmoides are inevitably influenced by climate change [18]. Prior research has demonstrated that rubber trees yield more rubber within a temperature range of 18–28 °C [27]. To understand the effect of temperature on the growth and isoprene metabolic pathway of E. ulmoides seedlings, this study analyzed the transcriptomes of E. ulmoides seedlings after 40 days of treatment at the control temperature and high temperature (HT) using RNA sequencing (RNA-Seq). Differentially expressed genes were analyzed using GO functional classification and KEGG enrichment analysis to verify the expression levels of genes related to the isoprene metabolic pathway. This analysis is crucial for using terpenoids as secondary metabolites and rubber resources in E. ulmoides.
2. Materials and Methods
2.1. Experimental Material
The E. ulmoides seeds used were of the ‘Huazhong 15’ variety and procured from Yuncheng Huimin Seedling Seed Company, Shanxi Province, in November 2021. The seeds were stored in a 4 °C refrigerator.
2.2. Methods
2.2.1. Analysis of Phenotypic Changes
Fresh and plump E. ulmoides seeds were carefully selected in the spring season. These seeds were soaked in warm water overnight, sown in seedling pots, and placed outdoors under natural conditions to facilitate germination. Once the seedlings had grown to the six-leaf stage, they were carefully transferred to an artificial climate chamber. Three temperature treatments were set up to study how the growth of E. ulmoides is affected by low and high temperatures: 15 °C/10 °C (day/night) for low temperature (LT), 35 °C/30 °C (day/night) for high temperature (HT), and 25 °C/20 °C (day/night) for the control group. The photoperiod was set at 16/8 h (day/night), with a relative humidity of 65% ± 5% and a light intensity of 12,000 Lux. Phenotypic observations were conducted at regular intervals of 10 days, specifically on days 0, 10, 20, 30, and 40. Measurements were collected from the first fully extended leaves: leaf length (from tip to base, excluding the petiole) and leaf width (at the widest point in the middle). Plant height was measured from the tip of the terminal leaf to the soil tip. For each group of temperature treatments, a leaf disk of 6 mm diameter was carefully taken from a consistent position near the bottom of the leaf. These leaf disks were then mounted in tin foil and dried at 120 °C. The leaves were further dried in an oven at 80 °C until a constant weight was achieved. The dry weight of leaf disks was divided by the area of the disks to calculate the specific leaf weight. Three biological replicates were performed per group, with ten plants per replicate.
2.2.2. Determination of the Chlorophyll Content
For this procedure, 0.1 g of E. ulmoides leaves was accurately weighed using a precision scale and placed in a dry mortar. A small amount of 95% anhydrous ethanol was added incrementally to the morstar. The leaves were ground thoroughly in a light-avoiding area. The resulting mixture was transferred to 10 mL centrifugal tubes. The volume was adjusted to 10 mL by adding 95% anhydrous ethanol, ensuring the process was performed in the dark at room temperature [28]. The mixture was left in the dark overnight. Next, the optical density (OD) values were measured at wavelengths of 649, 665, and 470 nm using the SpectraMax®(Molecular Devices, Sunnyvale, CA, USA) ABS Plus microplate reader. Three biological replicates were set up for each group of treatment, including three mechanical replicates. The relevant calculation formula was as follows:
ChIa = (13.95 A665 − 6.88 A649)
ChIb = (24.96 A649 − 7.32 A665)
ChI = ChIa + ChIb
Car = (1000A470 − 2.05 × ChIa−114.8 × ChIb)/245
2.2.3. Determination of Total Triterpene Content
The total triterpene content was determined according to the method described by Tao et al. [29], with minor modifications. The ethanol extract was heated at 90–100 °C to evaporate the reagent, then 0.1 mL of 5% vanilla-glacial acetic acid solution and 0.8 mL of perchloric acid were added. The mixture was incubated in a water bath at 60 °C for 20 min to develop the color, then it was removed and cooled in an ice water bath for 3–5 min. After that, 5.0 mL of glacial acetic acid was added, and the mixture was left at room temperature for 10 min. The zero point was adjusted with a 1 cm cuvette, and the absorbance was measured at the wavelength of 550 nm. A standard curve was prepared using oleanolic acid, and the standard equation for oleanolic acid was Y = 0.0051X − 0.0025 (R2 = 0.9993), where Y is the absorbance value and X is the oleanolic acid fructose content (μg). The total triterpenes content in the samples was determined using the following formula:
where x is the amount of total triterpenes in the sample; m1 is the amount of total triterpenes in the sample reaction solution; m2 is the sample mass; V1 is the volume of anhydrous ethanol; V2 is the volume of the extract; and f is the dilution factor.
2.2.4. Determination of TPI Content
The extraction and analysis of E. ulmoides rubber followed the method of Lu et al. [30], with some modifications. For E. ulmoides rubber extraction, the plant’s leaf was subjected to the following steps: initially, it was fixed in an oven at 120 °C for 30 min, dried at 80 °C until it reached a constant weight, and then weighed to determine the dry weight of the tissue. The dried tissue was subsequently ground into a powder. Next, an appropriate amount of 10% NaOH solution was added to the powder. The mixture was placed in a water bath at 100 °C for 3 h, followed by further heating at 120 °C for 20 min under HT and high-pressure conditions. The residual NaOH solution was then washed off. The remaining material was subjected to a 60 °C water bath with distilled water for 3 h and then dried at 37 °C. To extract rubber, the dried material was introduced into a Soxhlet extractor and an appropriate amount of petroleum ether (with a boiling range of 60–90 °C) was added. Extraction was performed at 120 °C for 12 h. The resulting solid, representing E. ulmoides rubber, was obtained and weighed (referred to as dry rubber). Each group involved three biological replicates, and within each replicate, there were six plants. The following formula was used to calculate the rubber content (%): Rubber content (%) = fine rubber/tissue dry weight × 100%.
2.2.5. Total RNA Extraction, cDNA Library Construction, and Illumina Sequencing
The leaves from various plants of E. ulmoides, specifically the second true leaf from top to bottom, which were subjected to a 40-day temperature treatment, were promptly frozen in liquid nitrogen and stored at −80 °C for future use. To extract total RNA from E. ulmoides, the Trizol method was employed. The purity of the RNA was assessed using the NanoDrop method, calculating the OD260/OD280 ratio, and its quality was verified using 1% agarose gel electrophoresis. Subsequently, Oligo (dT) beads were utilized to isolate the mRNA, followed by fragmentation of the mRNA into short fragments using an interrupting reagent. These short fragments were employed as templates for the synthesis of the first cDNA strand via first-strand synthesis. Following this, the Second Strand Synthesis Mix was utilized to synthesize the second strand. The resulting product underwent end leveling and had “A” added to it. RNA-Seq Adapter was connected to the product after this process. After ligating the RNA-Seq Adapter, the ligated product was purified, followed by PCR amplification. The target product of 300–600 bp was purified and recovered using magnetic beads. Finally, the cDNA library was sequenced on the Illumina NovaSeq 6000 platform once the library construction was deemed qualified.
2.2.6. Transcriptome Data Analysis
A similarity comparison was performed for the clean data and reference genome (https://ngdc.cncb.ac.cn/gwh/Assembly/25206/show, accessed on 4 September 2023). A reference rate of ≥80–85% was required to proceed to the next step of bioinformatics analysis. The screening criteria for differentially expressed genes involved expression folds meeting |log2(fold change)| > 1 and p ≤ 0.5. For differentially expressed genes, GO and KEGG enrichment analyses were performed using the R package cluster profiler based on these databases.
2.2.7. RT-qPCR Validation
To validate the accuracy of the transcriptome sequencing results, samples stored at −80 °C were used. Total RNA was extracted using the RC411 kit (Vazyme Biotech Co., Ltd., Nanjing, China). Subsequently, the total RNA was reverse transcribed with the First Strand cDNA Synthesis Kit (Vazyme Biotech Co., Ltd., Nanjing, China). EuActin was employed as an internal reference gene. For RT-qPCR, primers were designed for seven genes related to E. ulmoides rubber synthesis (Table 1) and these primers were synthesized by Sangong Bioengineering Corporation (Shanghai, China). The relative expression of each gene was calculated using the 2−ΔΔCT method. This analysis was performed with three biological replicates and three technical replicates for each sample group. The RT-qPCR analysis employed the SYBR Green Dye Kit (Vazyme Biotech Co., Ltd., Nanjing, China).
2.2.8. Statistical Analysis of Data
Data were organized using Excel 2022. Statistical analysis was performed using SPSS 22. Significance was analyzed using one-way ANOVA. Data were graphed using the GraphPad Prism 8.0 software.
3. Results
3.1. Effect of Temperature on Phenotype
Treatment of E. ulmoides seedlings with varying temperatures led to a significant increase in leaf length and leaf width as the treatment duration increased. The plant height did not show significant changes in HT and LT treatments but significantly increased in the control (Figure 1A–E). On day 40, the plant height of LT and HT groups significantly decreased by 41.7% and 45.5%, respectively, compared with that of the control group. The leaf length of LT and HT groups significantly decreased by 38% and 32.3%, respectively, compared with that of the control group. The leaf width of LT and HT groups significantly decreased by 44.7% and 40.3%, respectively, compared with that of the control group. (Figure 1A–E). Additionally, the specific leaf weight of LT on day 40 was 4.0 mg/cm2, which was 1.8 times higher than that of the control (Figure 1F). These results suggest that 25 °C was the optimal ambient temperature for the growth of E. ulmoides seedlings, whereas 35 °C and 15 °C adversely affected their growth and development. Moreover, the specific leaf weight under LT was significantly higher than that of the control and HT treatment, indicating thicker leaf growth under LT conditions.
3.2. Effect of Temperature on Chlorophyll and Carotenoid Contents
Chlorophyll content serves as a crucial indicator for assessing plant photosynthetic capacity. In this study, chlorophyll and carotenoid contents after various temperature treatments were assessed. The results indicated that after 40 days of temperature treatment, leaves in the control group appeared green, LT leaves had a purple color, and HT leaves showed a yellowish color (Figure 2A). The contents of chlorophyll a, chlorophyll b, and total chlorophyll followed the pattern control > LT > HT (Figure 2B–F). On day 40, chlorophyll a content in the control was 1.372 mg.g−1, which was 1.8 times higher than that in HT, and chlorophyll b content was 0.594 mg.g−1, which was 1.9 times higher than that in HT. These findings suggested that excessively high temperatures (35 °C) were harmful to chlorophyll accumulation in the leaf blades of E. ulmoides seedlings. The carotenoid content in LT was 0.58 mg.g−1, which was twice that in HT. Furthermore, chlorophyll a/b values decreased with prolonged temperature treatment.
3.3. Effect of Temperature on Rubber and Total Triterpene Contents
After 40 days, rubber was extracted from the true leaves of plants in each treatment group, and the total triterpene content was analyzed. The results revealed that the rubber contents in leaves were 1.22%, 1.44%, and 0.90% after 40 days of LT, control, and HT treatments, respectively. The rubber content of LT and HT groups significantly decreased by 16.5% and 38%, respectively, compared with that of the control group. The differences among the three treatments were significant (p < 0.05). Total triterpene contents were 1.33%, 1.02%, and 1.23% for LT, control, and HT, respectively. Notably, total triterpene contents in LT and HT were 30% and 20% higher than those in the control, respectively (Figure 3B). This suggests that rubber accumulation is less favorable at extreme temperatures (either too low or too high), causing a redirection of precursors synthesized by the isoprene metabolic pathway toward terpene synthesis.
3.4. Transcriptome Data Analyses
3.4.1. Transcriptome Data Quality
Leaves of the plants treated at 25 °C (control) and 35 °C (HT) for 40 days were collected. A total of six samples were subjected to RNA-Seq, yielding raw reads of 40, 885, 940–43, 679, 536 (Table 2). Clean reads were in the range of 40, 082, 724–43, 233, 386 after the removal of low-quality bases (accession number: PRJNA1030736). The Q20 probability of the samples, indicating a base quality of >20, ranged from 96.46% to 97.59%, and the Q30 probability, indicating a base quality of >30, ranged from 90.87% to 93.19%. The comparison reference rate to the reference genome exceeded 95.28%, indicating the high sequencing quality of the E. ulmoides leaf samples and their suitability for further bioinformatics analysis.
The analysis revealed 2309 DEGs after HT treatment compared with the control. Of these, 1608 genes were upregulated and 701 genes were downregulated (Figure 4). This suggests that DEGs were primarily enriched in the upregulated expression pathway after HT treatment.
3.4.2. GO and KEGG Enrichment Analysis of DEGs
The annotation of DEGs in E. ulmoides using the GO database revealed enrichment in three functional categories: molecular function, cellular components, and biological processes. In the biological process category, there was a notable focus on nitrogen compound metabolism (325), macromolecule metabolism (303), regulation of primary metabolism (165), and protein ubiquitination (23). In the molecular function category, emphasis was on catalytic activity (112), binding function (356), kinase activity (61), and protein ubiquitination transfer (23). In terms of cellular components, the major concentrations were in the cytoplasm (531) and plastids (142) (Figure 5A).
The annotation of DEGs into metabolic pathways and KEGG enrichment analysis revealed significant results. The DEGs were mainly enriched in five major categories of processes when comparing HT treatment with the control. These categories included cellular processes, environmental information processing, genetic information processing, metabolism, and organic systems. In terms of metabolic processes, the main enrichments were observed in carbohydrate metabolism (104), biosynthesis of secondary metabolites (87), and metabolism of terpenoids and ketones (31). Environmental information processing was enriched in signal transduction (127), and membrane transport (3). These findings suggest that HT treatment might have accelerated carbohydrate metabolism as well as terpene production and efflux (Figure 5B).
3.5. Effect of Temperature on Chlorophyll Metobolic Pathway
Given the close relationship between chlorophyll content and photosynthesis, the differential gene expression profiles of chlorophyll degradation metabolic pathways were analyzed. Two genes encoding chlorophyllases were identified, both of which were significantly upregulated after HT treatment. This suggests that the chlorophyllase gene is induced to express itself after HT treatment, promoting the degradation of chlorophyll into chlorophyllide a and chlorophyllide b. Consequently, this leads to a decrease in the total chlorophyll content (Figure 6).
3.6. Effect of Temperature on Isoprene Metabolic Pathway
To investigate the impact of temperature on the isoprene metabolic pathways, we examined the expression of relevant genes within the IPP biosynthetic pathway using transcriptome sequencing. In comparison with the control group, we observed alterations in gene expression. The 3-hydroxy3-methyl-coenzyme A reductase (HMGR) genes showed consistent upregulation. The 1-deoxyxylulose-5-phosphate synthetase (DXS) genes exhibited upregulation. Of the acetyl coenzyme A acetyltransferase (ACAT) genes, ACAT2 showed increased expression. The 3-hydroxy-3-methylglutarate monoacyl coenzyme A synthase (HMGS) genes, particularly HMGS2, displayed upregulated expression. In contrast, geranyl diphosphate synthase (GPS) genes, GPS2 and GPS3, demonstrated downregulated expression (Figure 7).
3.7. Validation of the Differential Gene Expression Level Using qRT-PCR
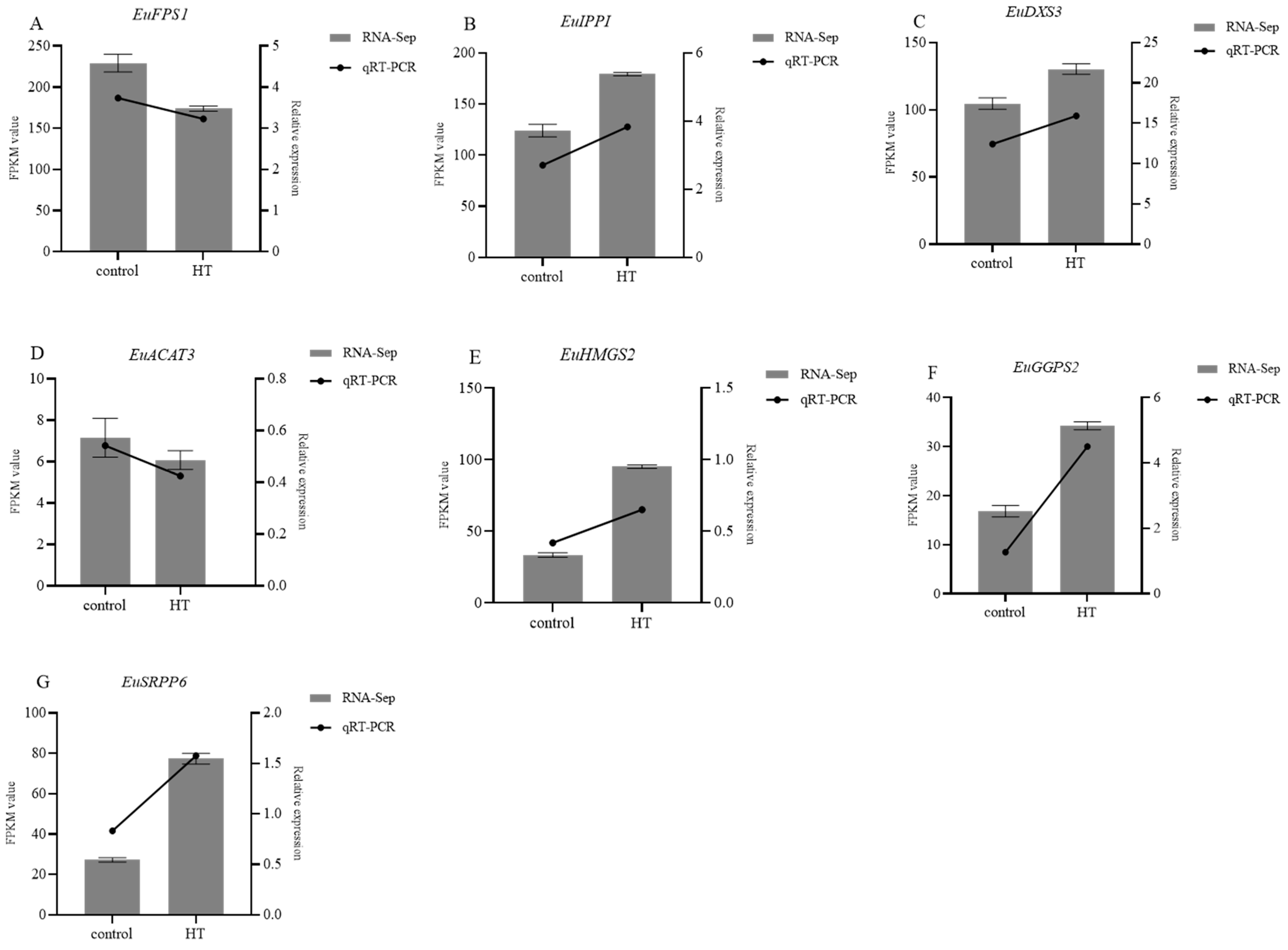
To further comprehend the effect of gene expression levels on the synthesis of secondary metabolites, seven genes with either upregulated or downregulated expression were selected under HT treatment. EuActin was used as an internal reference gene. The selected genes included ACAT3 and HMGS2 in the MVA pathway, DXS3 in the MEP pathway, and genes associated with rubber synthesis (IPPI, GGPS2, FPS1, and SRPP6). We utilized RT-qPCR to validate the accuracy of the transcriptome data analysis. The results of the qPCR amplification analyses were consistent with the expression trend of the differentiated genes observed in the transcriptome (Figure 8).
4. Discussion
Prolonged cold and heat stress during early growth can significantly hinder plant growth, leading to reduced plant heights and growth rates. Moreover, decreased photosynthesis and chlorophyll content have been documented in response to extreme temperatures. Factors such as leaf and stem scorching, as well as leaf abscission, further exacerbate the decline in yield [31,32,33]. Notably, chlorophyllase plays a pivotal role in catalyzing the hydrolysis of chlorophyll, forming chlorophyll lactones. A negative correlation exists between chlorophyll content and chlorophyllase activity [34]. Moreover, chlorophyllase is responsive to environmental stress, and environmental factors, such as strong light, pathogen infection, and leaf cell destruction, can accelerate chlorophyll degradation [35,36]. Our study revealed that growth parameters (e.g., plant height, leaf length, leaf width, specific leaf weight, and chlorophyll content) of E. ulmoides seedlings were affected by LT and HT treatments. Additionally, the expression of the CLH, responsible for chlorophyll degradation, was upregulated, suggesting that HT treatment accelerates chlorophyll decomposition and leads to reduced chlorophyll content. This is consistent with previous research.
Earlier studies have confirmed that HMGR is involved in rubber biosynthesis and associated with the biosynthesis of other terpenes [23,37]. A modest increase in geranylgeranyl pyrophosphate synthase (GPPS) activity boosts monoterpene production in transgenic tobacco [38]. Additionally, high temperature has been found to favor the expression of the Ginkgo GGPP gene [39]. Farnesyl pyrophosphate synthase (FPS) synthesizes FPP, which is conducive to the biosynthesis of rubber [40]. A study has demonstrated that TPS affects the concentration of IPP and DMAPP, as well as the flow direction, consequently impacting the rubber content [41], and its expression was mostly increased under osmotic stress and heat stress conditions [42].
Studies have indicated that cold and heat stress can affect the content of rubber and other terpenes [43,44,45], with rubber content decreasing and terpene content increasing under adversity. Chow noticed that IPP is used for the synthesis of carotenoids and rubber in Hevea brasiliensis [46]. In this study, the rubber content of LT and HT groups was significantly lower than that in the control group. However, the total triterpene content of LT and HT groups was significantly higher than that of control group. Analysis of the gene expression profiles related to the isoprene pathway revealed increased expression levels of HMGS2, HMGR, DXS, and IPPI under HT treatments. This enhanced expression facilitated the synthesis of IPP, a crucial precursor for the production of isoprene-like metabolites, ultimately laying the foundation for isoprene-like substance synthesis. However, while IPP precursor synthesis increased, GPS2 and GPS3 genes were downregulated at high temperatures, resulting in the reduced synthesis of GPP, the precursor for FPP synthesis. The majority of the synthesized GPP was directed toward monoterpene production. Furthermore, the expression of the FPS1 gene, responsible for FPP synthesis, was downregulated at high temperatures, leading to a decrease in FPP synthesis. Conversely, the expression of the GGPS gene, catalyzing GGPP synthesis using FPP, increased at high temperatures. Additionally, the expressions of most TPS genes were found to be upregulated. As a result, the synthesized FPP was primarily utilized for GGPP synthesis as well as the production of sesquiterpenes and triterpenes. It was speculated that in the control group, IPP was mainly used to synthesize rubber, whereas under HT conditions, despite an increase in IPP synthesis, IPP is predominantly diverted toward the synthesis of other terpenes and rubber; this may have contributed to the lower rubber content in the HT group than in the control group.
5. Conclusions
In this study, we confirmed that temperature conditions significantly influence rubber synthesis in E. ulmoides. Specifically, treatment at 25 °C was found to be conducive to the growth of E. ulmoides seedlings and rubber synthesis. Based on transcriptome and RT-qPCR results, our findings have led to the hypothesis that treatment at 35 °C upregulated the expression of CLH in the chlorophyll degradation pathway, leading to a reduction in the chlorophyll content and weakening of photosynthesis. Additionally, in the isoprene pathway, upregulation of GGPS and TPS genes, coupled with the downregulation of FPS1 and GPS, contributed to the diversion of IPP precursors toward the synthesis of terpenes, such as monoterpenes, diterpenes, and sesquiterpenes. This also led to the increased synthesis of GGPP while reducing the amount of FPP, the initiator of rubber synthesis in E. ulmoides, resulting in a decline in rubber content at high temperatures. Therefore, to achieve a high yield of E. ulmoides rubber, it is advisable to avoid HT conditions during cultivation. These results provide essential foundational data for understanding the regulatory mechanism of environmental factors in the biosynthesis of E. ulmoides rubber.
Author Contributions
W.Y. performed the experiment, analyzed the data, and wrote the initial draft of the paper. D.Z. conceived and oversaw the project, supervising the experimental design, data analysis, and paper revision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Guizhou Academy of Agricultural Sciences Talent Special Project, grant number((2022)02 and (2023)02), the project of the Talent Office of Guizhou Province(grant number RCJD2018-14), and the Guizhou Province High-level Innovative Talents Cultivation Project, grant number((2016) 4003).
Data Availability Statement
The Eucommia ulmoides transcriptome database has been successfully uploaded to the NCBI database. All the data from this study can be find in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Guirimand, G.; Papon, N.; Courdavault, V.; Thabet, I.; Ginis, O.; Bouzid, S.; Giglioli-Guivarc, H.N.; Clastre, M. Peroxisomal localisation of the final steps of the mevalonic acid pathway in planta. Planta 2011, 234, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M.; Wildung, M.R.; McCaskill, D.; Croteau, R. A family of transketolases that directs isoprenoid biosynthesis via a mevalonateindependent pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 2100–2104. [Google Scholar] [CrossRef] [PubMed]
- Lois, L.M.; Rodrigue-Concepcion, M.; Gallego, F.; Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J. 2000, 22, 503–513. [Google Scholar] [CrossRef]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef]
- Hou, X.; Rivers, J.; León, P.; McQuinn, R.P.; Pogson, B.J. Synthesis and function of apocarotenoid signals in plants. Trends Plant Sci. 2016, 21, 792–803. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Wiberley, A.E.; Donohue, A.R. Isoprene emission from plants: Why and how. Ann. Bot. 2008, 101, 5–18. [Google Scholar] [CrossRef]
- Duhl, T.R.; Helmig, D.; Guenther, A. Sesquiterpene emissions from vegetation: A review. Biogeosciences 2008, 5, 761–777. [Google Scholar] [CrossRef]
- Jia, Q.; Köllner, T.G.; Gershenzon, J.; Chen, F. MTPSLs: New terpene synthases in nonseed plants. Trends Plant Sci. 2018, 23, 121–128. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Jin, J.; Sarojam, R.; Ramachandran, S. A comprehensive survey on the terpene synthase gene family provides new insight into its evolutionary patterns. Genome Biol. Evol. 2019, 11, 2078–2098. [Google Scholar] [CrossRef]
- Zi, J.; Mafu, S.; Peters, R.J. To gibberellins and beyond! surveying the evolution of (di)terpenoid metabolism. Annu. Rev. Plant Biol. 2014, 65, 259–286. [Google Scholar] [CrossRef] [PubMed]
- Swiezewska, E.; Danikiewicz, W. Polyisoprenoids: Structure, biosynthesis and function. Prog. Lipid Res. 2005, 44, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Call, V.; Dilcher, D. The fossil record of Eucommia (Eucommiaceae) in North America. Am. J. Bot. 1997, 84, 798. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.R.; Zhu, M.Q.; Liang, J.; Su, Y.Q. Antioxidant and antimicrobial activities and component analysis of crude pyroligneous acids of Eucommia ulmoides branch made at different temperatures. J. Northwest For. Univ. 2016, 31, 220–226. [Google Scholar]
- Sun, Z.; Li, F.; Du, H.; Zhu, J.; Wang, Y.P. A novel silvicultural model for increasing biopolymer production from Eucommia ulmoides Oliver trees. Ind. Crops Prod. 2013, 42, 216–222. [Google Scholar] [CrossRef]
- K Kang, H.L.; Gong, M.; Xu, M.; Wang, H.Y.; Li, Y.S.; Fang, Q.H.; Zhang, L.Q. Fabricated biobased Eucommia ulmoides gum/polyolefin elastomer thermoplastic vulcanizates into a Shape memory material. Ind. Eng. Chem. Res. 2019, 58, 6375–6384. [Google Scholar] [CrossRef]
- Yang, R.F. Prospects and research progress on Eucommia ulmoides gum. Prog. Chem. 1995, 7, 65–71. [Google Scholar]
- Qi, X.; Zhao, X.; Li, Y.X.; Zhang, J.C.; Yue, D.M. A high toughness elastomer based on natural Eucommia ulmoides gum. J. Appl. Polym. Sci. 2020, 11, 50007. [Google Scholar] [CrossRef]
- Zhang, J.C.; Xue, Z.H. Study on under-water sound absorption properties of Eucommia ulmoides gum and its blends. Polym. Bull. 2011, 67, 511–525. [Google Scholar] [CrossRef]
- Fang, Q.H.; Jin, X.; Yang, F.; Ma, C.; Gao, Y.; Wang, N. Preparation and characterizations of Eucommia ulmoides gum/polypropylene blend. Polym. Bull. 2016, 73, 357–367. [Google Scholar] [CrossRef]
- Sando, T.; Takaoka, C.; Mukai, Y.; Yamashita, A.; Hattori, M.; Ogasawara, N.; Fukusaki, E.; Kobayashi, A. Cloning and characterization of mevalonate pathway genes in a natural rubber producing plant, Hevea brasiliensis. Biosci. Biotechnol. Biochem. 2008, 72, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Stermer, B.A.; Bostock, R.M. Involvement of 3-hydroxy-3-methylglutaryl coenzyme A reductase in the regulation of sesquiterpenoid phytoalexin synthesis in potato. Plant Physiol. 1987, 84, 404–408. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Craker, L.E.; Salami, A.; Nazeri, V.; Sang, H.; Maggi, F. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiol. Biochem. 2017, 111, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, O.R.; Oh, J.Y.; Jang, M.G.; Yang, D.C. Functional analysis of 3-hydroxy-3-methylglutaryl coenzyme A reductase encoding genes in triterpene saponin-producing ginseng. Plant Physiol. 2014, 165, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, A.; Ullah, S.; Dar, A.A.; Sardar, M.F.; Mehmood, T.; Tufail, M.A.; Shakoor, A.; Haris, M. Nexus on climate change: Agriculture and possible solution to cope future climate change stresses. Environ. Sci. Pollut. Res. 2021, 28, 14211–14232. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.M.; Li, H.X.; Chong, K. Crop improvement through temperature resilience. Annu. Rev. Plant Biol. 2019, 70, 753–780. [Google Scholar] [CrossRef] [PubMed]
- Shuogang, A.; Yagang, G. Extrapolation of the high yield physiological regulation of Hevea brasiliensis in Xishuangbanna. Proceeding of the IRRDB Symposium on Physiology and Exploitation of Hevea brasiliensis, Kunming, China, 6–7 October 1990; The International Rubber Research and Development Board: Kuala Lumpur, Malaysia, 1990; pp. 83–92. [Google Scholar]
- Liu, S.X.; Huang, Y.Z.; Luo, Z.J.; Huang, Y.C.; Yang, X.W. Effects of exogenous melatonin on accumulation and chemical form of Cd in rice. J. Appl. Ecol. 2017, 28, 1588–1594. [Google Scholar]
- Tao, W.Y.; Xu, H.Y.; Ao, Z.H.; Xu, Z.H. Quantitative analysis of triterpenoids from Antrodia camphorata in submerged culture. Chin. Trad. Patent Med. 2008, 30, 402–405. [Google Scholar]
- Lu, Z.K.; Xie, B.X.; Du, H.Y. Study on extraction method of Eucommia rubber. J. Fujian Coll. 2004, 24, 353–356. [Google Scholar]
- Al-Deeb, T.; Abo Gamar, M.; Khaleel, S.; Al-Ghzawi, A.L.; Al Khateeb, W.; Jawarneh, M.; Jahmani, M.Y.; Al-Zoubi, O.; Habeeb, T. Individual and interactive ecophysiological effect of temperature, watering regime and abscisic acid on the growth and development of tomato seedlings. Agronomy 2023, 13, 930. [Google Scholar] [CrossRef]
- Kim, S.; Hwang, G.; Lee, S.; Zhu, J.Y.; Paik, I.; Nguyen, T.T.; Kim, J.; Oh, E. High ambient temperature represses anthocyanin biosynthesis through degradation of HY5. Front. Plant Sci. 2017, 8, 1787. [Google Scholar] [CrossRef] [PubMed]
- Maulana, F.; Tesso, T.T. Cold temperature episode at seedling and flowering stages reduces growth and yield components in sorghum. Crop Sci. 2013, 53, 355–734. [Google Scholar] [CrossRef]
- Azoulay, S.T.; Harpaz, S.; Belausov, E.; Lovat, N.; Krokhin, O.; Spicer, V.; Standing, K.G.; Goldschmidt, E.E.; Eyal, Y. Citrus chlorophyllase dynamics at ethylene-induced fruit color-break: A study of chlorophyllase expression, posttranslational processing kinetics, and in situ intracellular localization. Plant Physiol. 2008, 148, 108–118. [Google Scholar] [CrossRef]
- Hu, X.; Makita, S.; Schelbert, S.; Sano, S.; Ochiai, M.; Tsuchiya, T.; Hasegawa, S.F.; Hörtensteiner, S.; Tanaka, A.; Tanaka, R. Reexamination of chlorophyllase function implies its involvement in defense against chewing herbivores. Plant Physiol. 2015, 167, 660–670. [Google Scholar] [CrossRef]
- Kariola, T.; Brader, G.; Li, J.; Palva, E.T. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell 2005, 17, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Chye, M.L.; Tan, C.T.; Chua, N.H. Three genes encode 3-hydroxy-3-methylglutaryl-coenzyme A reductase in Hevea brasiliensis: HMG1 and HMG3 are differentially expressed. Plant Mol. Biol. 1992, 19, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.L.; Wong, W.S.; Jang, I.C.; Chua, N.H. Co-expression of peppermint geranyl diphosphate synthase small subunit enhances monoterpene production in transgenic tobacco plants. New Phytol. 2017, 213, 1133–1144. [Google Scholar] [CrossRef]
- Hua, W.; Song, J.; Li, C.; Wang, Z. Molecular cloning and characterization of the promoter of SmGGPPs and its expression pattern in Salvia miltiorrhiza. Mol. Biol. Rep. 2012, 39, 5775–5783. [Google Scholar] [CrossRef]
- Deng, X.M.; Guo, D.; Yang, S.J.; Shi, M.J.; Chao, J.Q.; Li, H.L.; Peng, S.H.; Tian, W.M. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree, Hevea brasiliensis. J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef]
- Liang, J.; Wang, D.; Li, X.; Huang, W.; Xie, C.; Fu, M.; Zhang, H.; Meng, Q. In silico genome-wide mining and analysis of terpene synthase gene family in Hevea brasiliensis. Biochem. Genet. 2023, 61, 1185–1209. [Google Scholar] [CrossRef]
- Yan, Y.; Li, M.; Zhang, X.; Kong, W.; Bendahmane, M.; Bao, M.; Fu, X. Tissue-specific expression of the terpene synthase family genes in Rosa chinensis and effect of abiotic stress conditions. Genes 2022, 13, 547. [Google Scholar] [CrossRef] [PubMed]
- Blanch, J.S.; Peñuelas, J.; Sardans, J.; Lusia, J. Drought, warming and soil fertilization effects on leaf volatile terpene concentrations in Pinus halepensis and Quercus ilex. Acta Physiol. Plant. 2009, 31, 207–218. [Google Scholar] [CrossRef]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, U. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Omokhafe, K.O.O.; Emuedo, O.A.E. Evaluation of influence of five weather characters on latex yield in Hevea brasiliensis. Int. J. Agric. Res. 2006, 1, 234–239. [Google Scholar] [CrossRef]
- Chow, K.S.; Mat-Isa, M.N.; Bahari, A.; Ghazali, A.K.; Alias, H.; Mohd-Zainuddin, Z.; Hoh, C.C.; Wan, K.L. Metabolic routes affecting rubber biosynthesis in Hevea brasiliensis latex. J. Exp. Bot. 2012, 63, 1863–1871. [Google Scholar] [CrossRef]
Figure 1.
Effect of temperature on seedling growth: (A) plants before treatment; (B) treated plants; (C) plant height; (D) leaf length; (E) leaf width; (F) specific leaf weight. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).
Figure 1.
Effect of temperature on seedling growth: (A) plants before treatment; (B) treated plants; (C) plant height; (D) leaf length; (E) leaf width; (F) specific leaf weight. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).

Figure 2.
Effect of temperature on appearance as well as the chlorophyll and carotenoid contents of E. ulmoides. (A) leaf phenotype after 40 days of treatment; (B) chlorophyll a content; (C) chlorophyll b content; (D) chlorophyll content; (E) chlorophyll a/b values; (F) carotenoid content. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).
Figure 2.
Effect of temperature on appearance as well as the chlorophyll and carotenoid contents of E. ulmoides. (A) leaf phenotype after 40 days of treatment; (B) chlorophyll a content; (C) chlorophyll b content; (D) chlorophyll content; (E) chlorophyll a/b values; (F) carotenoid content. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).

Figure 3.
Impact of temperature on rubber and total triterpene contents in E. ulmoides seedling leaves: (A) change in rubber content; (B) change in total triterpene content. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).
Figure 3.
Impact of temperature on rubber and total triterpene contents in E. ulmoides seedling leaves: (A) change in rubber content; (B) change in total triterpene content. Note: “NS” and different letters above the bar represent insignificant and significant differences, respectively, between the control and treatment (p < 0.05).

Figure 4.
The number of differentially expressed genes in HT-vs-control groups.

Figure 5.
GO and KEGG classification of differentially expressed genes between the control and HT groups: (A) GO enrichment analysis; (B) KEGG enrichment analysis.
Figure 5.
GO and KEGG classification of differentially expressed genes between the control and HT groups: (A) GO enrichment analysis; (B) KEGG enrichment analysis.
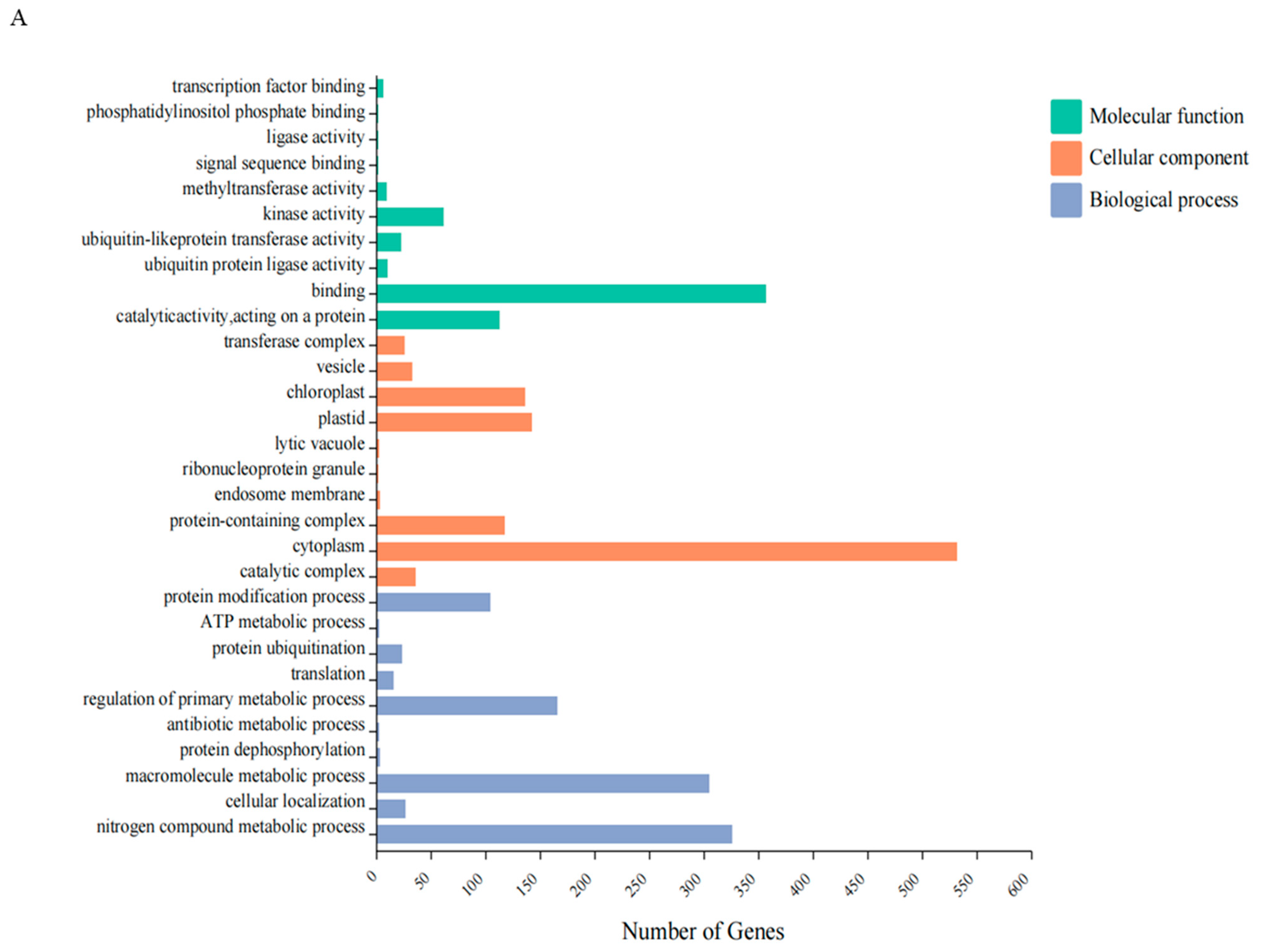
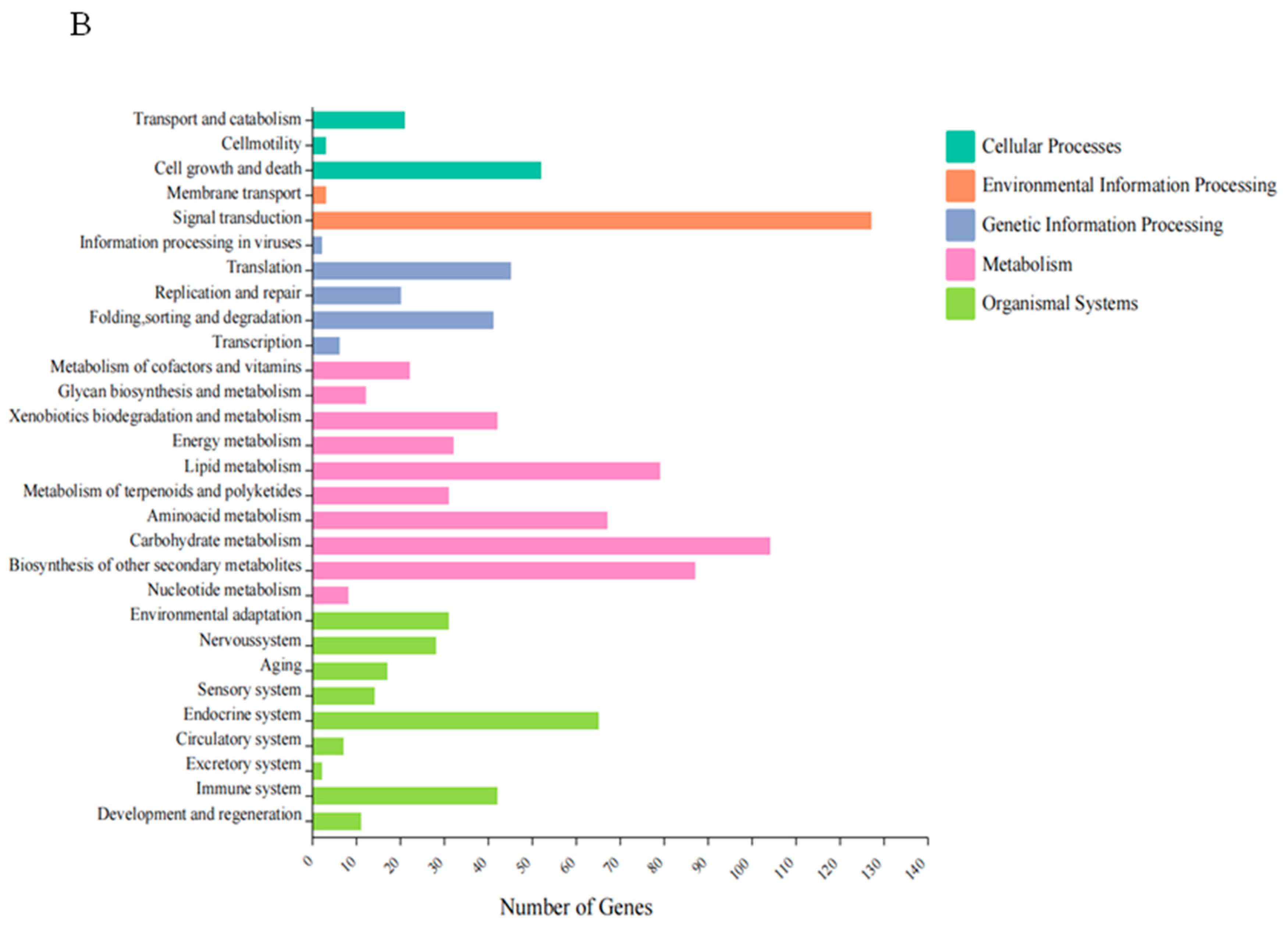
Figure 6.
Differential expression profile of the chlorophyll degradation pathway.

Figure 7.
HT-vs-control gene differential expression profile.
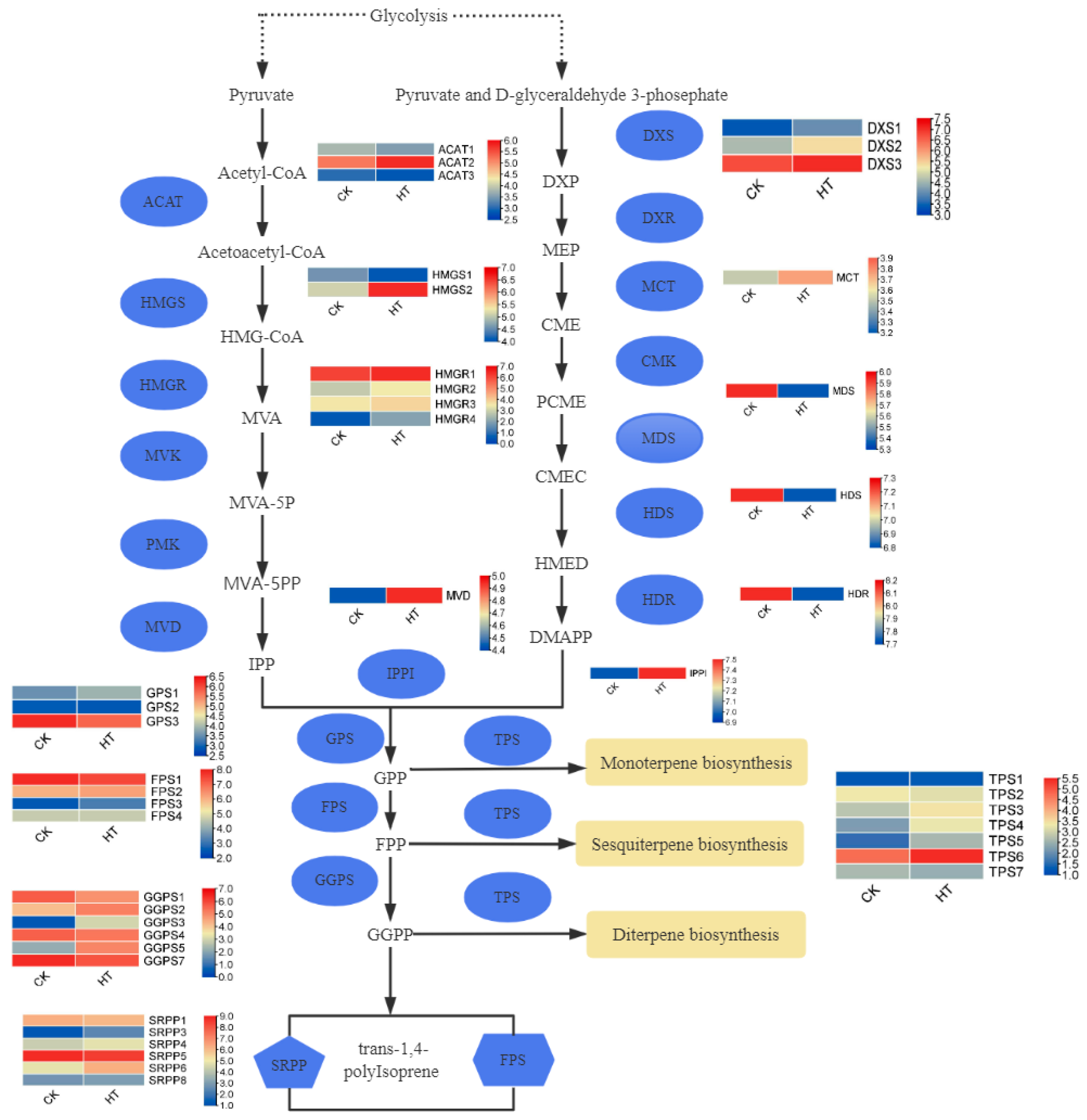
Figure 8.
The differentially expressed genes between HT and control verified using RT-qPCR: (A) EuFPS1; (B) EuIPPI; (C) EuDXS3; (D) EuACAT3; (E) EuHMGS2; (F) EuGGPS2; (G) EuSRPP6.
Figure 8.
The differentially expressed genes between HT and control verified using RT-qPCR: (A) EuFPS1; (B) EuIPPI; (C) EuDXS3; (D) EuACAT3; (E) EuHMGS2; (F) EuGGPS2; (G) EuSRPP6.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
RT-qPCR primer sequence.
| Gene Name | Primer Sequence (5′→3′) | Sequence Length |
|---|---|---|
| HMGS2 | F:GGAGAGCAGTTCGTGGGATGG R:GCAGCAGCTCCACCAGTCG | 99 |
| IPPI | F:ACTCGGTCCAGCTTTCGTCTTC R:AGAATCAGCGACGGCGGTATC | 81 |
| DXS3 | F:AGCAGGGCATAGTTCCACAAGC R:CATGGCTCCATCTCCGATCACC | 109 |
| SRPP6 | F:CGTCCGCTATGTCCGATCCATG R:GCGAACAGAGTCCAGGCAAGAG | 117 |
| ACAT3 | F:TCCAGCAGCAGCAACAGAGTG R:CCATTGGTGTTCGTGCAACTCC | 80 |
| GGPS2 | F:ACGACGACCTCCCCTGTATGG R:CCGTGGTGGAAGTGGCGATG | 144 |
| FPS1 | F:GGTTGGTGCATTGAGTGGCTTC R:TTGACCTCGGCGTGTGTGTG | 81 |
| Actin | F:TTGTTAGCAACTGGGATGATATGG R:CAGGGTGTTCTTCAGGAGCAA |
Table 2.
Data quality assessment statistics sheet.
| Sample Name | Raw Reads | Clean Reads | Q20 (%) | Q30 (%) | Mapped Read (%) | GC Content (%) |
|---|---|---|---|---|---|---|
| control-1 | 40,885,940 | 40,082,724 | 96.46 | 90.87 | 95.28 | 47.16 |
| control-2 | 41,794,830 | 41,135,962 | 96.80 | 91.52 | 95.55 | 46.97 |
| control-3 | 41,940,464 | 41,265,186 | 97.34 | 92.71 | 95.93 | 47.23 |
| HT-1 | 43,679,536 | 43,233,386 | 97.38 | 92.70 | 95.65 | 47.12 |
| HT-2 | 42,241,678 | 43,233,386 | 97.46 | 92.89 | 95.75 | 47.05 |
| HT-3 | 43,511,804 | 43,079,342 | 97.59 | 93.19 | 95.81 | 47.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yao, W.; Zhao, D. Effects of Temperature on Growth and Isoprene Metabolism Pathway in Eucommia ulmoides Oliv. Horticulturae 2023, 9, 1298. https://doi.org/10.3390/horticulturae9121298
AMA Style
Yao W, Zhao D. Effects of Temperature on Growth and Isoprene Metabolism Pathway in Eucommia ulmoides Oliv. Horticulturae. 2023; 9(12):1298. https://doi.org/10.3390/horticulturae9121298
Chicago/Turabian StyleYao, Wenqin, and Degang Zhao. 2023. "Effects of Temperature on Growth and Isoprene Metabolism Pathway in Eucommia ulmoides Oliv" Horticulturae 9, no. 12: 1298. https://doi.org/10.3390/horticulturae9121298
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.