



Phenotype “Explosion” in Hercinothrips femoralis (O. M. Reuter 1891) (Thysanoptera: Thripidae): A Particular Phenomenon for Successful Introduction of Economic Species

 and
and
Abstract
:1. Introduction
2. Materials and Methods
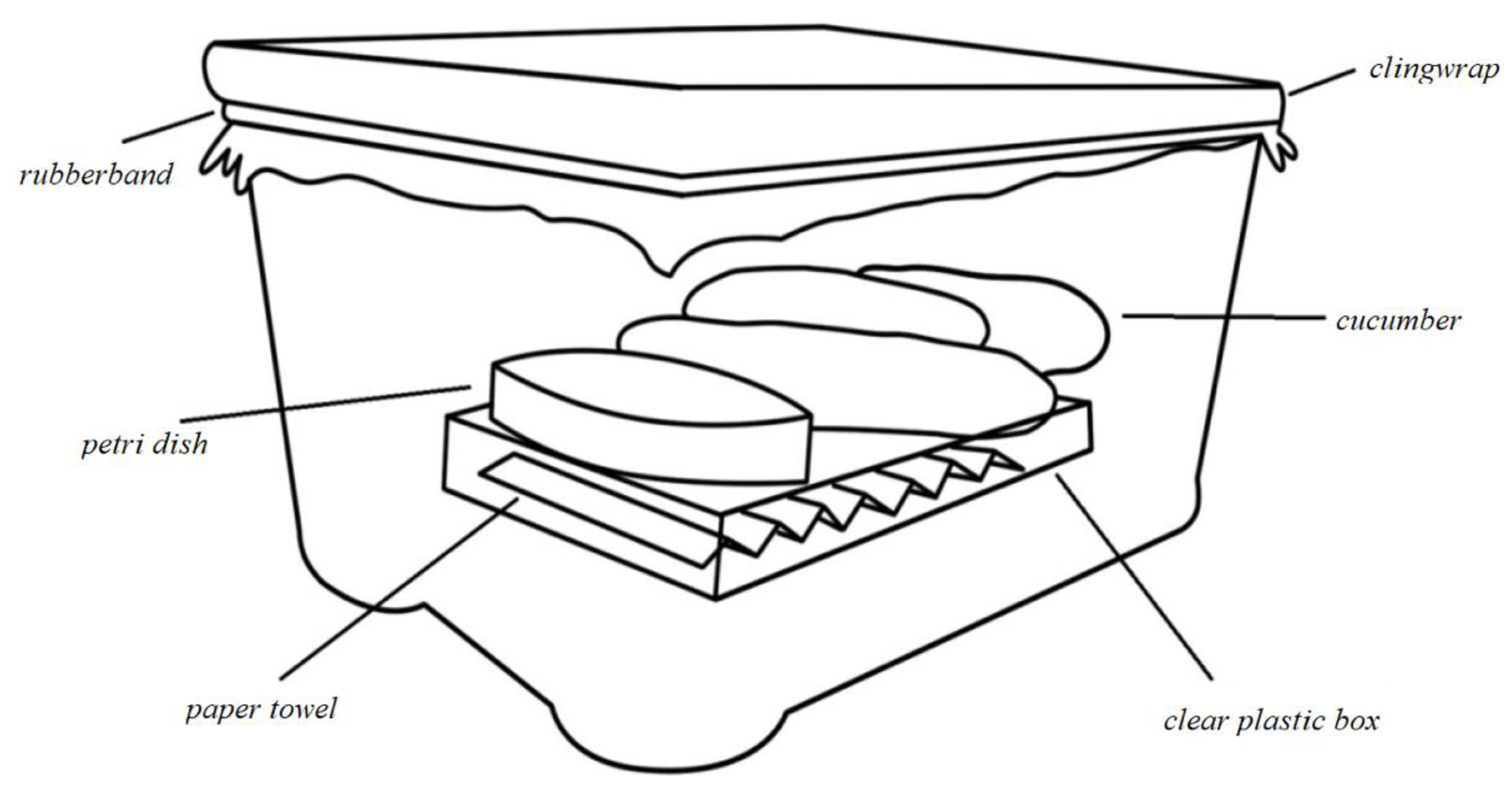
2.1. Experimental Design
2.2. Selection and Analysis of Morphological Variables of the Species Hercinothrips Femoralis
2.3. Statistical Analysis
3. Results
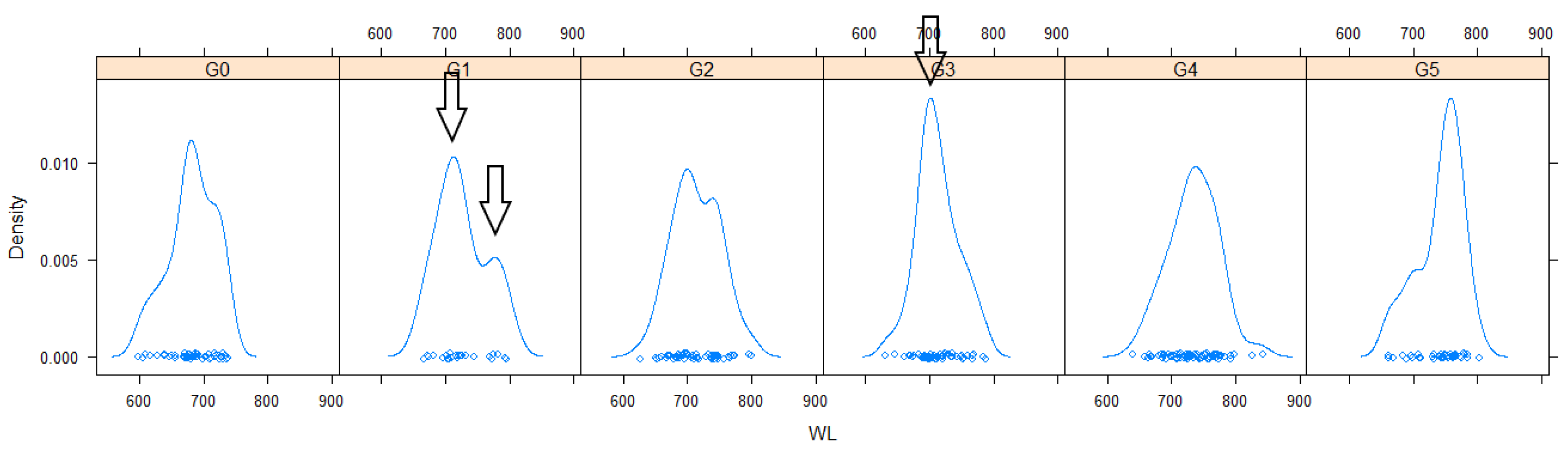
3.1. Phenotype “Explosion” during Primary Phase of Introduction
3.2. From Generalization (Phenotype “Explosion”) to Specialization (Selection of a Suitable Phenotype)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. App. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Rot, M.; Maistrello, L.; Costi, E.; Trdan, S. Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia. Insects 2022, 13, 956. [Google Scholar] [CrossRef] [PubMed]
- Whitman, D.W.; Agrawal, A.A. What is phenotypic plasticity and why is it important? In Phenotypic Plasticity of Insects: Mechanisms and Consequences; Whitman, D.W., Ananthakrishnan, T.N., Eds.; Science Publishers: Plymouth, NJ, USA; Enfield: Plymouth, NJ, USA, 2009; pp. 1–63. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Wennersten, L.; Forsman, A. Population-level consequences of polymorphism, plasticity and randomized phenotype switching: A review of predictions. Biol. Rev. 2012, 87, 756–776. [Google Scholar] [CrossRef]
- de Bello, F.; Carmona, C.P.; Dias, A.T.C.; Götzenberger, L.; Moretti, M.; Berg, M.P. Handbook of Trait-Based Ecology. From Theory to R Tools; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar] [CrossRef]
- Blanquart, F.; Kaltz, O.; Nuismer, S.L.; Gandon, S. A practival guide to measuring local adaptation. Ecol. Lett. 2013, 16, 1195–1205. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: New York, NY, USA, 2003; ISBN 9780195122350. [Google Scholar]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, T.J.; Scheiner, S.M. Phenotypic plasticity: Functional and Conceptual Approaches; Oxford University Press: Oxford, UK, 2004; p. 272. ISBN 0-19-513896-1. [Google Scholar]
- Gilbert, S.F.; Epel, D. Ecological Developmental Biology: The Environmental Regulation of Development, Health, and Evolution, 2nd ed.; Sinauer Assiociates: Sunderland, MA, USA, 2015; ISBN 978-1-60535-344-9. [Google Scholar]
- Sultan, E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impact on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef]
- Stearns, S.C. The evolutionary significance of phenotypic plasticity. BioScience 1989, 39, 436–445. [Google Scholar] [CrossRef]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 547–556. [Google Scholar] [CrossRef]
- Forsman, A.; Ahnesjö, J.; Caesar, S. Fitness benefits of diverse offspring in pygmy grasshoppers. Evol. Ecol. Res. 2007, 9, 1305–1318. [Google Scholar]
- Forsman, A. Effects of genotypic and phenotypic variation on establishment are important for conservation, invasion, and infection biology. Proc. Natl. Acad. Sci. USA 2014, 11, 302–307. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef]
- Musolin, D.L.; Saulich, K. Responses of insects to the current climate changes: From physiology and behavior to range shifts. Entomol. Rev. 2012, 92, 715–740. [Google Scholar] [CrossRef]
- Manfredini, F.; Arbetman, M.; Toth, A.L. A potential role for phenotypic plasticity in Invy in invasions and declines of social insects. Front Ecol. Evol. 2019, 7, 1–17. [Google Scholar] [CrossRef]
- Trdan, S.; Kužnik, L.; Vidrih, M. First results concerning the efficacy of entomopathogenic nematodes against Hercinothrips femoralis (Reuter). Acta Agric. Slov. 2007, 89, 5–13. [Google Scholar] [CrossRef]
- Houston, K.J.; Mound, L.A.; Palmer, J.M. Two pest thrips (Thysanoptera) new to Australia, with notes on the distribution and structural variation of other species. Austral J. Entomol. 1991, 30, 231–232. [Google Scholar] [CrossRef]
- Roditakis, E.; Mound, L.A.; Roditakis, N.E. Note: First record in crete of Hercinothrips femoralis in greenhouse banana plantations. Phytoparasitica 2006, 34, 488–490. [Google Scholar] [CrossRef]
- Mound, L.A.; Morison, G.D.; Pitkin, B.R.; Palmer, J.M. Thysanoptera. In Handbooks for the Identification of British Insects; Mound, L.A., Ed.; Royal Entomological Society: St Albans, UK, 1976; Volume 1, p. 79. [Google Scholar]
- Lee, G.S.; Lee, W. Rediscovery of an exotic thrips Hercinothrips femoralis (Reuter), in Korea, severely damaging to ornamental plants. Korean Soc. Appl. Entomol. 2016, 1, 102. [Google Scholar]
- Masarovič, R.; Doričová, M.; Prokop, P.; Fedor, P. “Testing the limits”—An interesting record of the exotic banded greenhouse thrips Hercinothrips femoralis (Thysanoptera: Thripidae: Panchaetothripinae) at high Carpathian Mountain altitudes. Biologia 2014, 69, 1631–1634. [Google Scholar] [CrossRef]
- Lewis, T. Thrips as Crop Pests; CABI Publishing: Wallingford, CT, USA, 1997; ISBN 9780851991788. [Google Scholar]
- Štefánik, M.; Zvaríková, M.; Masarovič, R.; Fedor, P. The significance of anthropochory in Hercinothrips femoralis (Thysanoptera: Thripidae)–Short Communication. Plant Prot. Sci. 2019, 55, 262–265. [Google Scholar] [CrossRef]
- Lewis, T. Thrips. Their Biology, Ecology, and Economic Importance; Academic Press: Cambridge, MA, USA, 1973; p. 349. ISBN 0124471609. [Google Scholar]
- Kumm, S.; Moritz, G. First detection of Wolbachia in arrhenotokous populations of thrips species (Thysanoptera: Thripidae and Phlaeothripidae) and its role in reproduction. Environ. Entomol. 2008, 37, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Spooner-Hart, R.N.; Riegler, M. Polyploidy versus endosymbionts in obligately thelytokous thrips. BMC Evol. Biol. 2015, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Štefánik, M. Morphometry of Hercinothrips Femoralis in Changing Environments; Comenius University: Bratislava, Slovakia, 2020; p. 122. [Google Scholar]
- Fedor, P.; Malenovský, I.; Vaňhara, J.; Sierka, W.; Havel, J. Thrips (Thysanoptera) identification using artificial neural networks. Bull. Entomol. Res. 2008, 98, 437–447. [Google Scholar] [CrossRef]
- Fedor, P.; Peña-Mendéz, E.M.; Kucharczyk, H.; Vaňhara, J.; Havel, J.; Doričová, M.; Prokop, P. Artificial neural networks in online semiautomated pest dicriminability—An applied case of 2 Thrips species. Turk. J. Agric. For. 2014, 38, 111–124. [Google Scholar] [CrossRef]
- Zvaríková, M.; Kiseľák, J.; Zvarík, M.; Masarovič, R.; Prokop, P.; Fedor, P. Ecological gradients and environmental impact in the forest dwelling Haplothrips subtilissimus (Thysanoptera: Phlaeothripidae) phenotypic variability. Ecol. Indic. 2016, 66, 313–320. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 31 October 2022).
- Basset, Y.; Springate, N.D.; Samuelson, G.A. Feeding habits and range of body size: A case study in Papua New Guinea using arboreal leaf-beetles and weevils (Coleoptera: Chrysomelidae, Curculionidae). Mitteilungen Der Schweiz. Ges. 1994, 67, 347–361. [Google Scholar]
- Dmitriew, C.; Rowe, C. The effect of larval nutrition on reproductive performance in a food limited adult environment. PLoS ONE 2011, 6, e17399. [Google Scholar] [CrossRef]
- Fedor, P.; Štefánik, M.; Zvaríková, M.; Masarovič, R.; Sigmund, J. Phenotypic “explosion” as a morphometric phenomenon in successful introduction and biological invasion. In 5th Symposium on Palaearctic Thysanoptera; University of Agriculture: Krakow, Poland, 2017; pp. 20–21. [Google Scholar]
- Parker, J.; Johnston, L.A. The proximate determinants of insect size. J. Biol. 2006, 5, 15. [Google Scholar] [CrossRef]
- Gilpin, M.E.; Soulé, M.E. Minimum viable populations: Processes of species extinction. In Conservation Biology: The Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer: Sunderland, MA, USA, 1986; pp. 19–34. ISBN 0878937951. [Google Scholar]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1983; ISBN 0-521-24684-9. [Google Scholar]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Lenaerts, A.W.; Coulter, A.A.; Feiner, Z.S.; Goforth, R.R. Egg size variability in an establishing population of invasive silver carp Hypophthalmichthys molitrix (Valenciennes, 1844). Aquat. Invasions 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Distribution, abundance and population dynamics. In Thrips as Crop Pest; Lewis, T., Ed.; CAB International: Oxon, UK, 1997. [Google Scholar]
- Masarovič, R.; Zvaríková, M.; Fedorová, J.; Fedor, P. Thigmotactic behavior of Limothrips cerealium (Thysanoptera: Thripidae) leads to laboratory equipment damage in the Czech Republic. J. Entomol. Sci 2017, 52, 308–310. [Google Scholar] [CrossRef]
- Berrigan, D.; Charnov, E.L. Reaction norms for age and size at maturity in response to temperature: A puzzle for life historians. Oikos 1994, 70, 474–478. [Google Scholar] [CrossRef]
- Berg, M.P.; Kiers, E.T.; Driessen, G.; van der Heijden, M.; Kooi, B.W.; Kuenen, F.; Liefting, M.; Verhoef, H.A.; Ellers, J. Adapt or disperse: Understanding of species persistence in a changing world. Glob. Chang. Biol. 2010, 16, 587–598. [Google Scholar] [CrossRef]
- Schloss, C.A.; Nunez, T.A.; Lawler, J.J. Dispersal will limit ability of mammals to track climate change in the Western Hemisphere. Proc. Natl. Acad. Sci. USA 2012, 109, 8606–8611. [Google Scholar] [CrossRef] [PubMed]
- Chuang, A.; Peterson, C.R. Expanding population edges: Theories, traits, and trait-offs. Glob. Chang. Biol. 2016, 22, 494–512. [Google Scholar] [CrossRef]
- Thomas, C.D.; Bodsworth, E.J.; Wilson, R.J.; Simmons, A.D.; Davies, Z.G.; Musche, M.; Conradt, L. Ecological and evolutionary processes at expanding range margins. Nature 2001, 411, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Gutowsky, L.F.G.; Fox, M.G. Occupation, body size and sex ratio of round goby (Neogobius melanostomus) in established and newly invaded areas of an Ontario river. Hydrobiologia 2011, 671, 27–37. [Google Scholar] [CrossRef]
- Brown, G.P.; Kelehear, C.; Shine, R. The early toad gets the worm: Cane toads at an invasion front benefit from higher prey availability. J. Anim. Ecol. 2013, 82, 854–862. [Google Scholar] [CrossRef]
- Betts, C.R.; Wootton, R.J. Wing shape and flight behaviour in butterflies (Lepidoptera: Papilionoidea and Hesperioidea): A preliminary analysis. J. Exp. Biol. 1988, 138, 271–288. [Google Scholar] [CrossRef]
- DeVries, P.J.; Penz, C.M.; Hill, R.I. Vertical distribution, flight behaviour and evolution of wing morphology in Morpho butterflies. J. Anim. Ecol. 2010, 79, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. The role of propagule pressure in biological invasions. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge Univ Press: Cambridge, UK, 2010; ISBN 978-0-521-70271-3. [Google Scholar]
- Forsman, A.; AhnesjÖ, J.; Caeasar, S.; Karlsson, M. A model of ecological and evolutionary consequences of color polymorphism. Ecology 2008, 89, 34–40. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Källström, B. Increasing intraspecific diversity increases predictability in population survival in the face of perturbations. Oikos 2007, 116, 700–705. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SD | HW | PL | PW | WL | CL | OW | BL |
|---|---|---|---|---|---|---|---|
| G0 | 5.41 | 7.25 | 8.11 | 36.68 | 5.94 | 5.02 | 75.33 |
| G1 | 4.58 | 8.25 | 7.92 | 38.91 | 6.01 | 5.77 | 116.01 |
| G2 | 5.01 | 8.06 | 8.16 | 37.40 | 5.87 | 6.23 | 97.80 |
| G3 | 4.09 | 5.70 | 7.14 | 33.09 | 4.35 | 5.08 | 84.62 |
| G4 | 3.70 | 9.77 | 6.83 | 39.39 | 5.15 | 5.24 | 121.42 |
| G5 | 3.73 | 7.43 | 6.75 | 35.87 | 3.94 | 7.07 | 64.13 |
| s2 | HW | PL | PW | WL | CL | OW | BL |
|---|---|---|---|---|---|---|---|
| G0 | 29.28 | 52.63 | 65.79 | 1345.42 | 35.33 | 25.25 | 5674.08 |
| G1 | 21.00 | 68.07 | 62.80 | 1514.05 | 36.16 | 33.31 | 13,458.03 |
| G2 | 25.06 | 64.98 | 66.50 | 1398.66 | 34.46 | 38.81 | 9565.71 |
| G3 | 16.73 | 32.51 | 50.96 | 1095.11 | 18.96 | 25.77 | 7159.71 |
| G4 | 13.70 | 95.49 | 46.69 | 1551.62 | 26.51 | 27.49 | 14,742.25 |
| G5 | 13.95 | 55.26 | 45.53 | 1286.63 | 15.56 | 49.99 | 4112.21 |
| HW | PL | PW | WL | CL | OW | BL | |
|---|---|---|---|---|---|---|---|
| G1-G0 | ** | ||||||
| G2-G1 | |||||||
| G3-G2 | . | . | * | ||||
| G4-G3 | *** | * | |||||
| G5-G4 | * | . | ** | ||||
| Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1 | |||||||
| IQR | HW | PL | PW | WL | CL | OW | BL |
|---|---|---|---|---|---|---|---|
| G0 | 7.83 | 11.54 | 11.19 | 45.35 | 9.57 | 6.67 | 95.39 |
| G1 | 4.74 | 10.23 | 12.85 | 53.88 | 8.60 | 7.66 | 190.05 |
| G2 | 5.90 | 9.18 | 9.40 | 54.05 | 9.02 | 9.82 | 149.41 |
| G3 | 4.44 | 8.04 | 10.79 | 40.80 | 4.54 | 5.85 | 94.71 |
| G4 | 5.79 | 17.22 | 9.14 | 55.53 | 7.55 | 6.74 | 177.59 |
| G5 | 5.58 | 9.91 | 7.87 | 44.22 | 5.44 | 11.51 | 89.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masarovič, R.; Zvaríková, M.; Marcišová, M.; Provaznik, Z.J.; Prokop, P.; Fedor, P. Phenotype “Explosion” in Hercinothrips femoralis (O. M. Reuter 1891) (Thysanoptera: Thripidae): A Particular Phenomenon for Successful Introduction of Economic Species. Horticulturae 2023, 9, 1327. https://doi.org/10.3390/horticulturae9121327
Masarovič R, Zvaríková M, Marcišová M, Provaznik ZJ, Prokop P, Fedor P. Phenotype “Explosion” in Hercinothrips femoralis (O. M. Reuter 1891) (Thysanoptera: Thripidae): A Particular Phenomenon for Successful Introduction of Economic Species. Horticulturae. 2023; 9(12):1327. https://doi.org/10.3390/horticulturae9121327
Chicago/Turabian StyleMasarovič, Rudolf, Martina Zvaríková, Michaela Marcišová, Zuzana Ježová Provaznik, Pavol Prokop, and Peter Fedor. 2023. "Phenotype “Explosion” in Hercinothrips femoralis (O. M. Reuter 1891) (Thysanoptera: Thripidae): A Particular Phenomenon for Successful Introduction of Economic Species" Horticulturae 9, no. 12: 1327. https://doi.org/10.3390/horticulturae9121327
APA StyleMasarovič, R., Zvaríková, M., Marcišová, M., Provaznik, Z. J., Prokop, P., & Fedor, P. (2023). Phenotype “Explosion” in Hercinothrips femoralis (O. M. Reuter 1891) (Thysanoptera: Thripidae): A Particular Phenomenon for Successful Introduction of Economic Species. Horticulturae, 9(12), 1327. https://doi.org/10.3390/horticulturae9121327