

Managing Water Stress in Olive (Olea europaea L.) Orchards Using Reference Equations for Midday Stem Water Potential

 , , and
, , and
Abstract
:1. Introduction
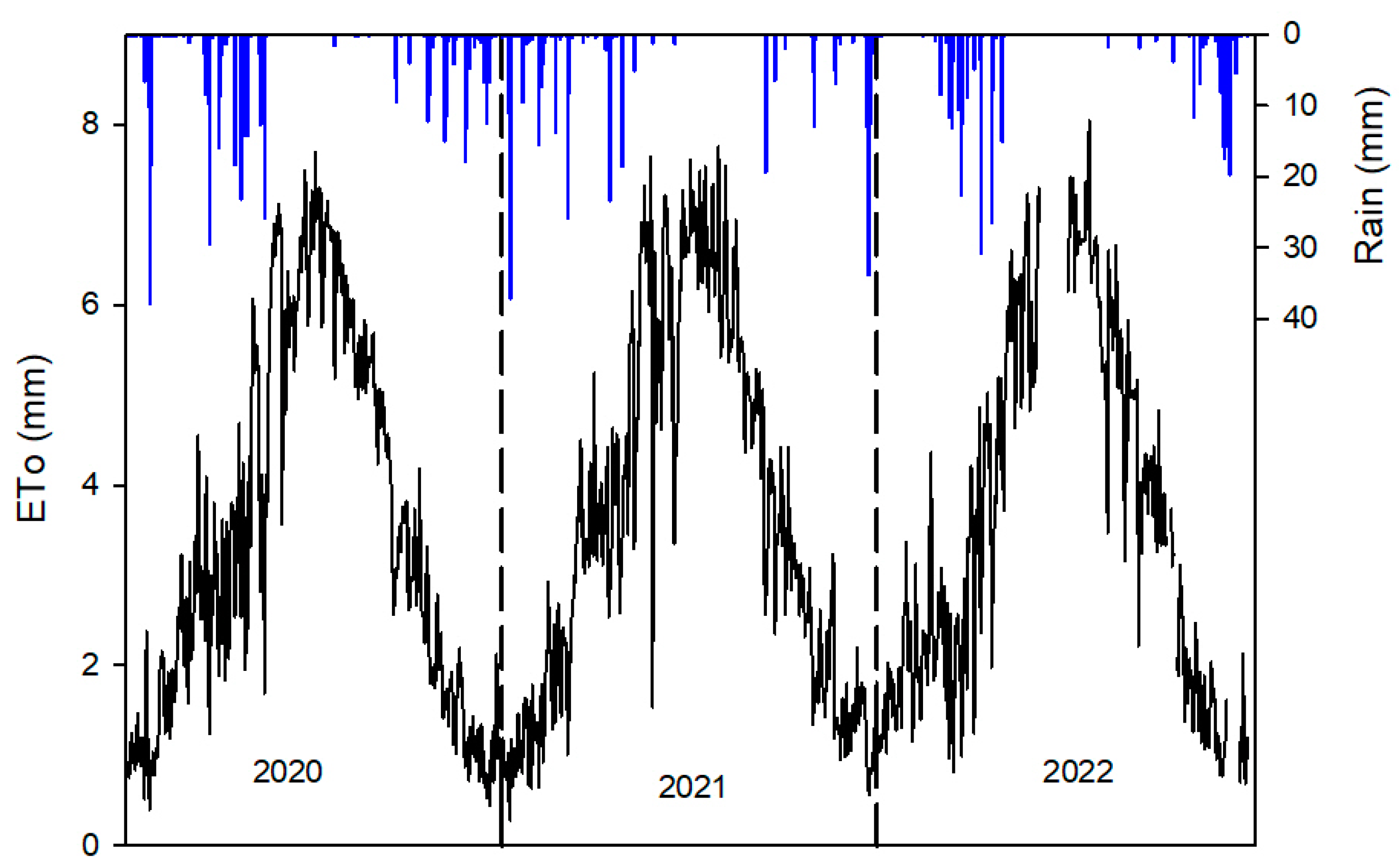
2. Materials and Methods
2.1. Description of the Site and the Treatments
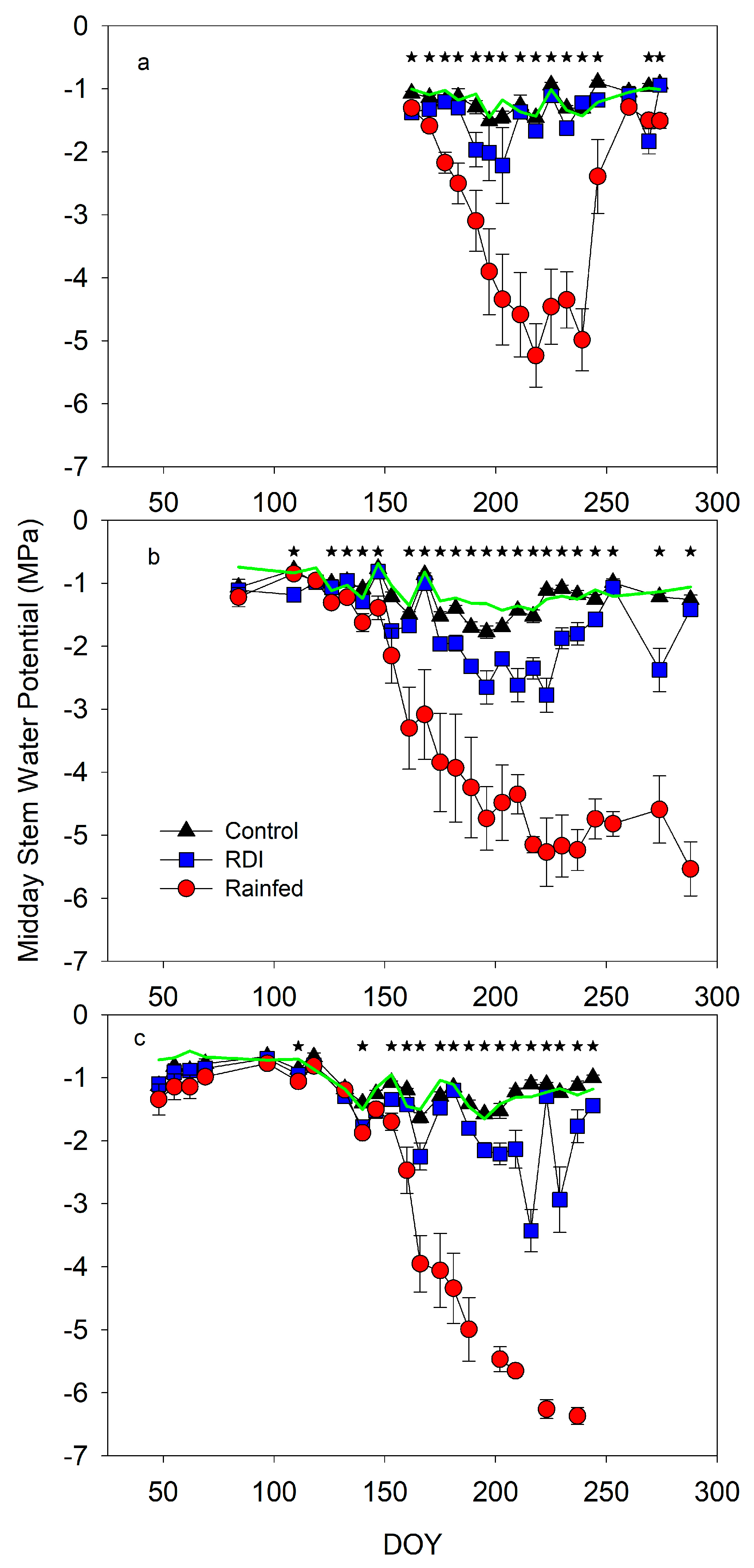
- Control. Irrigation scheduling was based on the midday stem water potential (SWP) measurements. During the 2020 and 2021 seasons, the target was to maximize vegetative growth. Then, the amount of irrigation was estimated using the FAO 56 approach (crop coefficients (Kc) 0.55, reduction coefficient (Kr) 0.7). However, when the SWP was more negative than −1.0 MPa, the water applied increased to 175% crop evapotranspiration (Etc). The harvest yield in the 2021 season was considered similar to that of a mature orchard (nearly 10,000 kg ha−1). Therefore, irrigation scheduling changed in order to control vegetative growth. In the 2022 season, irrigation scheduling was used considering the same approach, but the SWP threshold was estimated based on the [12] baseline and it was lowered in 0.5 Mpa from pit hardening until harvest. The water applied per season was 553, 772, and 727 mm in 2020, 2021, and 2022, respectively.
- Regulated deficit irrigation (RDI). The objective of this treatment was to apply mild or moderate water stress conditions, depending on the phenological stage. Irrigation scheduling was also based on the SWP measurements and on the comparison with the [12] baseline. Each plot was irrigated according to its SWP data. Trees were irrigated with 1 mm day−1 from the first date when the SWP was below the threshold, and the water applied increased by 2, 3, or 4 mm day−1 based on the distance to baseline. During the 2020 and 2021 seasons, in order to minimize the decrease in vegetative growth, the SWP threshold was the one obtained at the baseline and only during pit hardening it decreased to −2 MPa. In the 2022 season, the SWP threshold was the baseline value minus 0.5 MPa, but during pit hardening, it was minus 1.5 MPa. The water applied during the season was 267, 334, and 200 mm, respectively, in 2020, 2021, and 2022.
- Rainfed. Trees were not irrigated from June 2020. Only in September of this season they were irrigated with 45 mm before harvest. No irrigation was applied during 2021 and 2022.
2.2. Description of Measurements
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MAPA 2023. Yearbook of the Spanish Agriculture Ministry (Several Years). Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/default.aspx (accessed on 15 March 2023).
- Hsiao, T.C. Measurements of plant water status. In Irrigation of Agricultural Crops; Steward, B.A., Nielsen, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1990; pp. 243–280. [Google Scholar]
- Chalmers, D.J.; Wilson, I.B. Productivity of peach trees: Tree growth and water stress in relation to fruit growth and assimilate demand. Ann. Bot. 1978, 42, 285–294. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Fereres, E.; Raes, D. Crop Yield Response to Water; FAO Irrigation and Drainage Paper 66; FAO: Rome, Italy, 2012. [Google Scholar]
- Moriana, A.; Orgaz, F.; Fereres, E.; Pastor, M. Yield responses of a mature olive orchard to water deficits. J. Am. Soc. Hortic. Sci. 2003, 128, 425–431. [Google Scholar] [CrossRef]
- Ben-Gal, A.; Ron, Y.; Yermiyahu, U.; Zipori, I.; Naoum, S.; Dag, A. Evaluation of regulated deficit irrigation strategies for oil olives: A case study for two modern Israeli cultivars. Agric. Water Manag. 2021, 245, 106577. [Google Scholar] [CrossRef]
- Moriana, A.; Pérez-López, D.; Prieto, M.H.; Ramírez-Santa-Pau, M.; Pérez-Rodriguez, J.M. Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agric. Water Manag. 2012, 112, 43–54. [Google Scholar] [CrossRef]
- Girón, I.F.; Corell, M.; Martín-Palomo, M.J.; Galindo, A.; Torrecillas, A.; Moreno, F.; Moriana, A. Feasibility of trunk diameter fluctuations in the scheduling of regulated deficit irrigation for table olive trees without references trees. Agric. Water Manag. 2015, 161, 114–126. [Google Scholar] [CrossRef]
- McCutchan, H.; Shackel, K.A. Stem-water potential as a sensitive indicator of water stress in Prune trees (Prunus domestica L. cv French). J. Amer. Soc. Hort. Sci. 1992, 117, 607–611. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Fereres, E. Irrigation scheduling of almond trees with trunk diameter sensors. Irrig. Sci. 2004, 23, 11–19. [Google Scholar] [CrossRef]
- Ortuño, M.F.; Conejero, W.; Moreno, F.; Moriana, A.; Intrigliolo, D.S.; Biel, C.; Mellisho, C.D.; Pérez-Pastor, A.; Domingo, R.; Ruiz-Sánchez, M.C.; et al. Could trunk diameter sensors be used in woody crops for irrigation scheduling? A review of current knowledge and future perspectives. Agric. Water Manag. 2010, 97, 1–11. [Google Scholar] [CrossRef]
- Corell, M.; Pérez-López, D.; Martín-Palomo, M.J.; Centeno, A.; Girón, I.; Galindo, A.; Moreno, M.M.; Moreno, C.; Memmi, H.; Torrecillas, A.; et al. Comparison of the water potential baseline in different locations. Usefulness for irrigation scheduling of olive orchards. Agric. Water Manag. 2016, 177, 308–316. [Google Scholar] [CrossRef]
- Shackel, K.; Moriana, A.; Marino, G.; Corell, M.; Perez-Lopez, D.; Martin-Palomo, M.J.; Caruso, T.; Marra, F.; Agüero-Alcaras, L.M.; Milliron, L.; et al. Establishing a reference baseline for midday stem water potential in olive and its use for plant-based irrigation management. Frontiers 2021, 12, 791711. [Google Scholar] [CrossRef]
- Marino, G.; Caruso, T.; Ferguson, L.; Marra, F.P. Gas exchanges and stem water potential define stress thresholds for efficient irrigation management in Olive (Olea europaea L.). Water 2018, 10, 342. [Google Scholar] [CrossRef]
- Ahumada-Orellana, L.; Ortega-Farias, S.; Poblete-Echeverría, C.; Searles, P.S. Estimation of stomatal conductance and stem water potential threshold values for water stress in olive trees (cv Arbequina). Irrig. Sci. 2019, 37, 461–467. [Google Scholar] [CrossRef]
- Hueso, A.; Trentacoste, E.R.; Junquera, P.; Gómez-Miguel, V.; Gómez del Campo, M. Differences in stem water potential during oil synthesis determine fruit characteristics and production but not vegetative growth or return bloom in an olive hedgerow orchard (cv Arbequina). Agric. Water Manag. 2019, 223, 105589. [Google Scholar] [CrossRef]
- Fernández, J.E. Understanding olive adaptation to abiotic stresses as a tool to increase crop performance. Environ. Exp. Bot. 2014, 103, 158–179. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Diaz-Espejo, A.; Morales-Sillero, A.; Martín-Palomo, M.J.; Mayr, S.; Beikircher, B.; Fernández, J.E. Shoot hydraulic characteristics, plant water status and stomatal response in olive trees under different soil water conditions. Plant Soil 2013, 373, 77–87. [Google Scholar] [CrossRef]
- Perez-Martin, A.; Flexas, J.; Ribas-Carbó, M.; Bota, J.; Tomás, M.; Infante, J.M.; Diaz-Espejo, A. Interactive effects of soil waterdeficit and air vapour pressure deficit on mesophyll conductance to CO2 in Vitis vinifera and Olea europea. J. Exp. Bot. 2009, 60, 2391–2405. [Google Scholar] [CrossRef]
- Fernández, J.E.; Moreno, F. Water use by the olive tree. J. Crop Production 1999, 2, 101–162. [Google Scholar] [CrossRef]
- Moriana, A.; Fereres, E. Plant indicators for scheduling irrigation for young olive trees. Irrig. Sci. 2002, 21, 83–90. [Google Scholar] [CrossRef]
- Díaz-Espejo, A.; Walcroft, A.; Fernandez, J.E.; Hafidi, B.; Palomo, M.J.; Giron, I.F. Modelling photosynthesis in olive leaves under drought conditions. Tree Physiol. 2006, 26, 1445–1456. [Google Scholar] [CrossRef]
- Cuevas, M.V.; Torres-Ruiz, J.M.; Alvarez, R.; Jimenez, M.D.; Cuerva, J.; Fernandez, J.E. Assessment of trunk diameter variations derived índices as water stress indicators in mature olive tres. Agric. Water Manag. 2010, 97, 1293–1302. [Google Scholar] [CrossRef]
- Moriana, A.; Villalobos, F.J.; Fereres, E. Stomatal and photosynthetic responses of olive (Olea europaea L.) leaves to water deficit. Plant Cell Environ. 2002, 25, 395–405. [Google Scholar] [CrossRef]
- Díaz-Espejo, A.; Fernández, J.E.; Torres-Ruíz, J.M.; Rodríguez-Domínguez, C.M.; Pérez-Martin, A.; Hernández-Santana, V. The olive tree under water stress: Fitting the pieces of response mechanisms in the crop performance puzzle. In Water Scarcity and Sustainable Agriculture in Semiarid Environment. Tools, Strategies and Challenges for Woody Crops; García-Tejero, I., Martin-Zuago, V.H., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 439–480. [Google Scholar]
- SIAR 2023. Climatic Station Data in Spain. Available online: http://eportal.mapa.gob.es/websiar/SeleccionParametrosMap.aspx?dst=1 (accessed on 15 March 2023).
- AEMET 2023. Spanish Agency of Meteorology. Available online: http://www.aemet.es/es/serviciosclimaticos/datosclimatologicos/valoresclimatologicos?l=5783&k=undefined (accessed on 15 March 2023).
- Rapoport, H.F.; Pérez-López, D.; Hammami, S.B.M.; Aguera, J.; Moriana, A. Fruit pit hardening: Physical measurements during olive growth. Ann. Appl. Biol. 2013, 163, 200–208. [Google Scholar] [CrossRef]
- Gomez-del-Campo, M. Physiological and growth responses to irrigation of a newly established hedgerow olive orchard. HortScience 2010, 45, 809–814. [Google Scholar] [CrossRef]
- Moriana, A.; Pérez-López, D.; Gómez-Rico, A.; Salvador, M.D.; Olmedilla, N.; Ribas, F.; Fregapane, G. Irrigation scheduling for traditional, low density olive orchards: Water relations and influence on oil characteristics. Agric. Water Manag. 2007, 87, 171–179. [Google Scholar] [CrossRef]
- Agüero-Alcaras, L.M.; Rousseaux, M.C.; Searles, P.S. Responses of several soil and plant idicators to postharvestregulated deficit irrigation in olive trees and their potential for irrigation scheduling. Agric. Water Manag. 2016, 171, 10–20. [Google Scholar] [CrossRef]
- Padilla-Díaz, C.M.; Rodriguez-Dominguez, C.M.; Hernández-Santana, V.; Perez-Martin, A.; Fernandes, R.D.M.; Montero, A.; Garcia, J.M.; Fernández, J.E. Water status, gas Exchange and crop performance in a super high density olive orchard under deficit irrigation scheduled from leaf turgor measurements. Agric. Water Manag. 2018, 202, 241–252. [Google Scholar] [CrossRef]
- Corell, M.; Martín-Palomo, M.J.; Girón, I.; Andreu, L.; Galindo, A.; Centeno, A.; Pérez-López, D.; Moriana, A. Stem water potential-based regulated deficit irrigation scheduling for olive table trees. Agric. Water Manag. 2020, 242, 106418. [Google Scholar] [CrossRef]
- Pérez-López, D.; Gijón, M.C.; Moriana, A. Influence of irrigation rate on the rehydration of olive tree plantlets. Agric. Water Manag. 2008, 95, 1161. [Google Scholar] [CrossRef]
- Morales-Sillero, A.; García, J.M.; Torres-Ruiz, J.M.; Montero, A.; Sánchez-Ortiz, A.; Fernández, J.E. Is the productive performance of olive trees under localized irrigation affected by leaving some roots in drying soil? Agric. Water Manag. 2013, 123, 79–92. [Google Scholar] [CrossRef]
- Fereres, E. Variability in Adaptive Mechanisms to Water Deficits in Annual and Perennial Crop Plants. Actual. Bot. 1984, 131, 17–32. [Google Scholar] [CrossRef]
- Pérez-López, D.; Ribas, F.; Moriana, A.; Olmedilla, N.; De Juan, A. The effect of irrigation schedules on the water relations and growth of a young olive (Olea europaea L.) orchard. Agric. Water Manag. 2007, 89, 297–304. [Google Scholar] [CrossRef]
- López-Bernal, A.; Fernandes-Silva, A.A.; Vega, V.A.; Hidalgo, J.C.; León, L.; Testi, L.; Villalobos, F.J. A fruit growth approach to estimate oil content in olives. Eur. J. Agron. 2021, 123, 126206. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
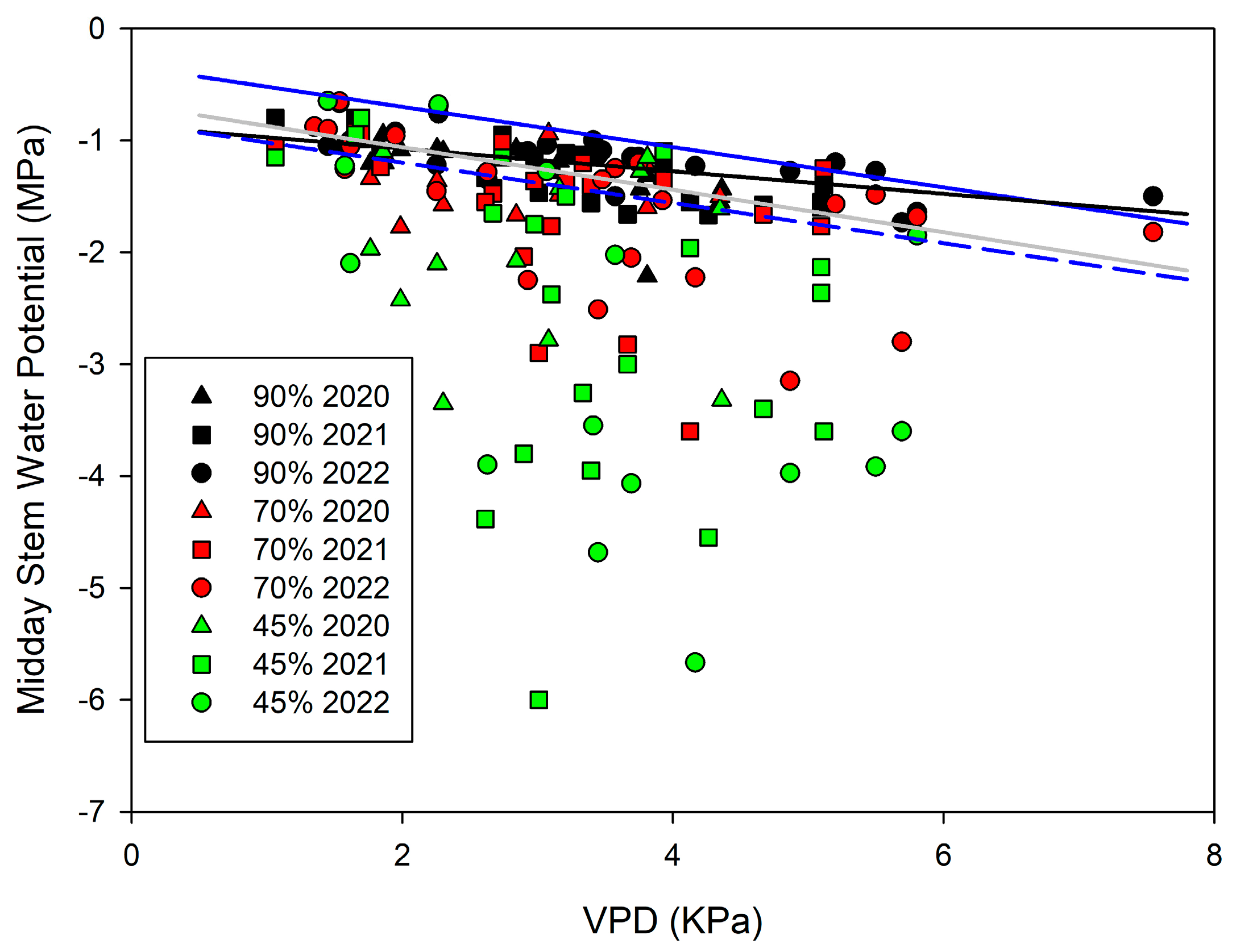
| Dataset | Equation | Data 1 | R2 | SD | MSE | Sig |
|---|---|---|---|---|---|---|
| 90% Tmax | SWP= −0.039 Tmax | 61 | 0.44 *** | 0.21 | 0.04 | No |
| 70% Tmax | SWP= −0.050 Tmax | 56 | 0.27 *** | 0.52 | 0.27 | No |
| 45% Tmax | SWP = −0.081 Tmax | 48 | 0.16 *** | 1.24 | 1.54 | No |
| ISO Tmax | SWP= −0.468-0.023 Tmax | 84 | 0.39 *** | 0.16 | 0.03 | Yes |
| 90% VPD | SWP= −0.785-0.14 VPD | 61 | 0.43 *** | 0.21 | 0.04 | Yes |
| 70% VPD | SWP= −0.859-0.21 VPD | 56 | 0.21 *** | 0.54 | 0.30 | No |
| 45% VPD | SWP = −1.119-0.43 VPD | 48 | 0.15 ** | 1.26 | 1.59 | No |
| ISO VPD | SWP= −0.871-0.10 VPD | 84 | 0.34 *** | 0.17 | 0.03 | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Piñero, M.; Corell, M.; Moriana, A.; Castro-Valdecantos, P.; Martin-Palomo, M.-J. Managing Water Stress in Olive (Olea europaea L.) Orchards Using Reference Equations for Midday Stem Water Potential. Horticulturae 2023, 9, 563. https://doi.org/10.3390/horticulturae9050563
Sánchez-Piñero M, Corell M, Moriana A, Castro-Valdecantos P, Martin-Palomo M-J. Managing Water Stress in Olive (Olea europaea L.) Orchards Using Reference Equations for Midday Stem Water Potential. Horticulturae. 2023; 9(5):563. https://doi.org/10.3390/horticulturae9050563
Chicago/Turabian StyleSánchez-Piñero, Marta, Mireia Corell, Alfonso Moriana, Pedro Castro-Valdecantos, and María-José Martin-Palomo. 2023. "Managing Water Stress in Olive (Olea europaea L.) Orchards Using Reference Equations for Midday Stem Water Potential" Horticulturae 9, no. 5: 563. https://doi.org/10.3390/horticulturae9050563