The History of Bancroftian Lymphatic Filariasis in Australasia and Oceania: Is There a Threat of Re-Occurrence in Mainland Australia?



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Lifecycle
3. Disease
4. Diagnosis
4.1. ICT Antigen Test
4.2. Filariasis Test Strip (FTS)
4.3. Blood Smears (mf Detection)
4.4. Other Diagnostic Methods
5. Prevention and Treatment
6. History of LF in Australia
6.1. Natural History in Australia
6.2. Discovery of the Adult Parasites
6.3. Discovering the Vector
7. Epidemiology
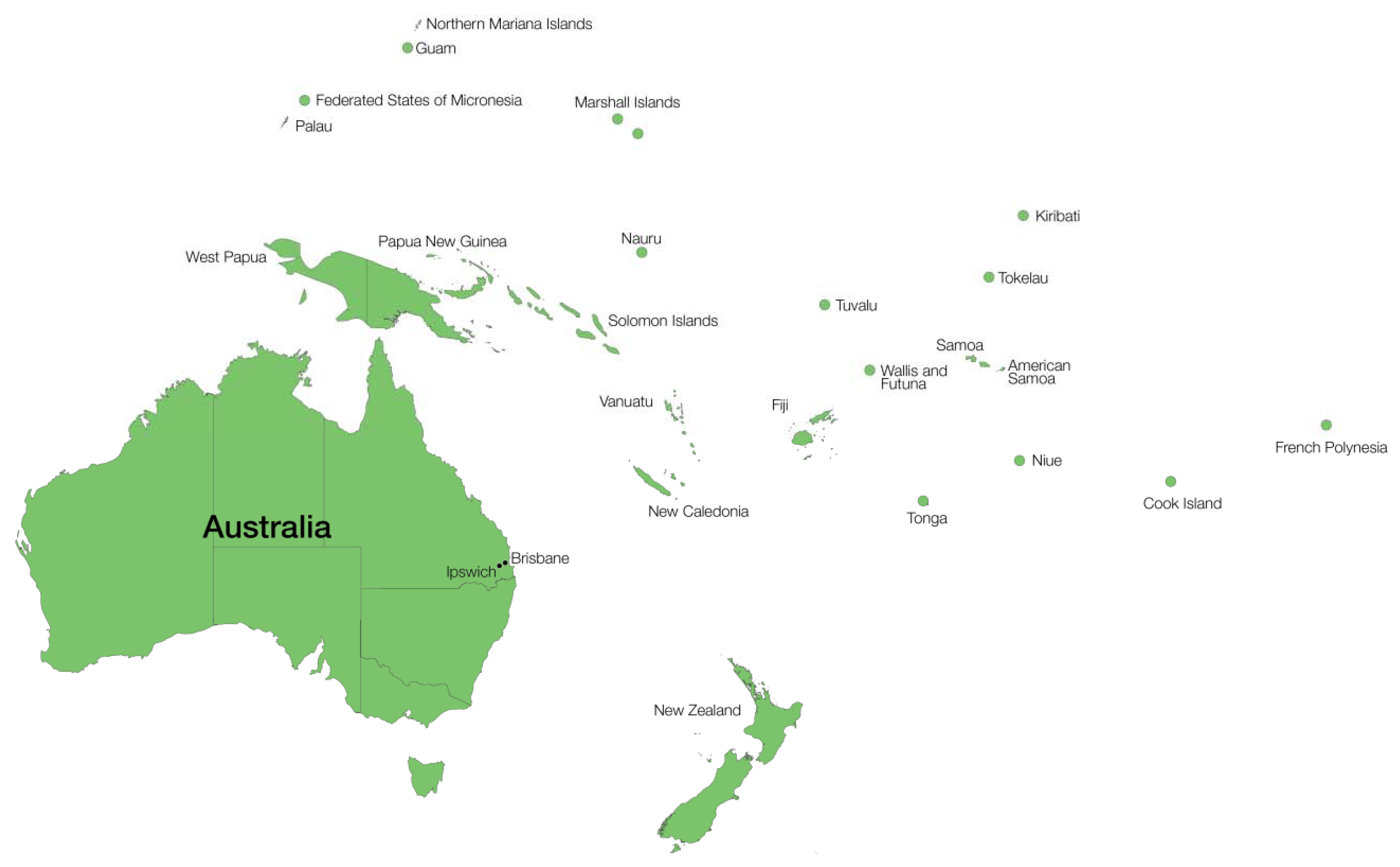
8. Current LF Prevalence in Oceania
8.1. Active Transmission
8.1.1. Papua New Guinea (PNG)
8.1.2. The Indonesian Province of Papua (Originally Irian Jaya)
8.1.3. Timor-Leste (East Timor)
8.1.4. Samoa (Formerly Western Samoa)
8.2. Active Surveillance
8.2.1. New Caledonia
8.2.2. Tuvalu (Formerly Ellice Islands)
8.2.3. Micronesia
8.2.4. Fiji
8.3. Elimination Achieved
8.3.1. Solomon Islands
8.3.2. Republic of Vanuatu
8.3.3. Tonga
8.3.4. Cook Islands
8.3.5. Niue
9. Immigration to Australia
10. Mosquito Hosts for LF
10.1. Host Species and Distribution in Australia (Including Islands of the Torres Strait)
10.2. Vector Competence—Are Some Species Better Than Others at Transmitting LF
Climate and Potential Spread of Mosquito Vectors in Australia
11. Conclusions
Author Contributions
Conflicts of Interest
References
- Angus, B.M.; Cannon, L.R.G.; Adlard, R.D. Parasitology and the Queensland Museum with Biographical Notes on Collectors; Queensland Museum: Queensland, Austrialia, 2007; Volume 53. [Google Scholar]
- Goel, T.C.; Goel, A. Lymphatic Filariasis; Springer Nature: Singapore, 2016. [Google Scholar]
- Hajdu, S.I. Elephantiasis. Ann. Clin. Lab. Sci. 2002, 32, 207–209. [Google Scholar] [PubMed]
- Small, S.T.; Ramesh, A.; Bun, K.; Reimer, L.; Thomsen, E.; Baea, M.; Bockarie, M.J.; Siba, P.; Kazura, J.W.; Tisch, D.J.; et al. Population genetics of the filarial worm Wuchereria bancrofti in a post-treatment region of Papua New Guinea: Insights into diversity and life history. PLoS Negl. Trop. Dis. 2013, 7, e2308. [Google Scholar] [CrossRef] [PubMed]
- WHO. Lymphatic Filariasis. Available online: http://www.who.int/mediacentre/factsheets/fs102/en/ (accessed on 25 January 2018).
- Gordon, C.A.; McManus, D.P.; Jones, M.K.; Gray, D.J.; Gobert, G.N. The increase of exotic zoonotic helminth infections: The impact of urbanization, climate change and globalization. Adv. Parasitol. 2016, 91, 311–397. [Google Scholar] [CrossRef] [PubMed]
- Boreham, P.F.L.; Marks, E.N. Human filariasis in Australia: Introduction, investigation and elimination. R. Soc. Qld. 1986, 97, 23–52. [Google Scholar]
- Sasa, M. Human Filariasis; Univeristy of Tokyo Press: Tokyo, Japan, 1976. [Google Scholar]
- Jeremiah, C.J.; Aboltins, C.A.; Stanley, P.A. Lymphatic filariasis in Australia: An update on presentation, diagnosis and treatment. Med. J. Aust. 2011, 194, 655–657. [Google Scholar] [PubMed]
- Kline, K.; McCarthy, J.S.; Pearson, M.; Loukas, A.; Hotez, P.J. Neglected tropical diseases of Oceania: Review of their prevalence, distribution, and opportunities for control. PLoS Negl. Trop. Dis. 2013, 7, e1755. [Google Scholar] [CrossRef] [PubMed]
- Graves, P.M.; Makita, L.; Susapu, M.; Brady, M.A.; Melrose, W.; Capuano, C.; Zhang, Z.; Dapeng, L.; Ozaki, M.; Reeve, D.; et al. Lymphatic filariasis in Papua New Guinea: Distribution at district level and impact of mass drug administration, 1980 to 2011. Parasit. Vectors 2013, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- WHO. Summary of global update on preventive chemotherapy implementation in 2016: Crossing the billion. Wkly. Epidemiol. Rec. 2017, 92, 589–609. [Google Scholar]
- WHO. Global Programme to Eliminate Lymphatic Filariasis. Available online: http://www.who.int/lymphatic_filariasis/elimination-programme/en/ (accessed on 25 January 2018).
- CDC. Biology—Life Cycle of Wuchereria bancrofti. Available online: https://www.cdc.gov/parasites/lymphaticfilariasis/biology_w_bancrofti.html (accessed on 1 February 2018).
- Joseph, H.; Speare, R.; Melrose, W. Laboratory diagnosis of lymphatic filariasis in Australia: Available tools and interpretation. Aust. J. Med. Sci. 2012, 33, 2–9. [Google Scholar]
- Shenoy, R.K.; Bockarie, M.J. Lymphatic filariasis in children: Clinical features, infection burdens and future prospects for elimination. Parasitology 2011, 138, 1559–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammie, P.J.; Cuenco, K.T.; Punkosdy, G.A. The pathogenesis of filarial lymphedema: Is it the worm or is it the host? Ann. N. Y. Acad. Sci. 2002, 979, 131–142; discussion 188–196. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, R.K. Clinical and pathological aspects of filarial lymphedema and its management. Korean J. Parasitol. 2008, 46, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Melrose, W.D. Lymphatic filariasis: New insights into an old disease. Int. J. Parasitol. 2002, 32, 947–960. [Google Scholar] [CrossRef]
- Stocks, M.E.; Freeman, M.C.; Addiss, D.G. The effect of hygiene-based lymphedema management in lymphatic filariasis-endemic areas: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2015, 9, e0004171. [Google Scholar] [CrossRef] [PubMed]
- Mullerpattan, J.B.; Udwadia, Z.F.; Udwadia, F.E. Tropical pulmonary eosinophilia—A review. Indian J. Med. Res. 2013, 138, 295–302. [Google Scholar] [PubMed]
- Melrose, W.; Rahmah, N. Use of Brugia rapid dipstick and ICT test to map distribution of lymphatic filariasis in the Democratic Republic of Timor-Leste. Southeast Asian J. Trop. Med. Public Health 2006, 37, 22–25. [Google Scholar] [PubMed]
- Knott, J. The periodicity of the microfilaria of Wuchereria bancrofti. Preliminary report of some injection experiments. Trans. R. Soc. Trop. Med. Hyg. 1935, 29, 59–64. [Google Scholar] [CrossRef]
- Jongthawin, J.; Intapan, P.M.; Lulitanond, V.; Sanpool, O.; Thanchomnang, T.; Sadaow, L.; Maleewong, W. Detection and quantification of Wuchereria bancrofti and Brugia malayi DNA in blood samples and mosquitoes using duplex droplet digital polymerase chain reaction. Parasitol. Res. 2016, 115, 2967–2972. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; McManus, D.P. Cell-free DNA as a diagnostic tool for human parasitic infections. Trends Parasitol. 2016, 32, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Weil, G.J.; Lammie, P.J.; Weiss, N. The ICT filariasis test: A rapid-format antigen test for diagnosis of bancroftian filariasis. Parasitol. Today 1997, 13, 401–404. [Google Scholar] [CrossRef]
- Weil, G.J.; Liftis, F. Identification and partial characterisation of a parasite antigen in sera from humans infected with Wuchereria bancrofti. J. Immunol. 1987, 138, 3035–3041. [Google Scholar] [PubMed]
- Wanji, S.; Amvongo-Adjia, N.; Njouendou, A.J.; Kengne-Ouafo, J.A.; Ndongmo, W.P.; Fombad, F.F.; Koudou, B.; Enyong, P.A.; Bockarie, M. Further evidence of the cross-reactivity of the Binax NOW filariasis ICT cards to non-Wuchereria bancrofti filariae: Experimental studies with Loa loa and Onchocerca ochengi. Parasit. Vectors 2016, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Wanji, S.; Amvongo-Adjia, N.; Koudou, B.; Njouendou, A.J.; Chounna Ndongmo, P.W.; Kengne-Ouafo, J.A.; Datchoua-Poutcheu, F.R.; Fovennso, B.A.; Tayong, D.B.; Fombad, F.F.; et al. Cross-Reactivity of filariasis ICT Cards in areas of contrasting endemicity of Loa loa and Mansonella perstans in Cameroon: Implications for shrinking of the lymphatic filariasis map in the central African region. PLoS Negl. Trop. Dis. 2015, 9, e0004184. [Google Scholar] [CrossRef] [PubMed]
- WHO. Improved Availability of New Test to Enhance Global Lymphatic Filariasis Elimination. Available online: http://www.who.int/neglected_diseases/news/new_test_enhance_global_lf_elimination/en/ (accessed on 25 May 2018).
- Weil, G.J.; Curtis, K.C.; Fakoli, L.; Fischer, K.; Gankpala, L.; Lammie, P.J.; Majewski, A.C.; Pelletreau, S.; Won, K.Y.; Bolay, F.K.; et al. Laboratory and field evaluation of a new rapid test for detecting Wuchereria bancrofti antigen in human blood. Am. J. Trop. Med. Hyg. 2013, 89, 11–15. [Google Scholar] [CrossRef] [PubMed]
- WHO. Strengthening the Assessment of Lymphatic Filariasis Transmission and Documenting the Achievement of Elimination; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Masson, J.; Douglass, J.; Roineau, M.; Aye, K.M.; Htwe, K.M.; Warner, J.; Graves, P.M. Relative performance and predictive values of plasma and dried blood spots with filter paper sampling techniques and dilutions of the lymphatic filariasis Og4C3 antigen ELISA for samples from Myanmar. Trop. Med. Infect. Dis. 2017, 2, 7. [Google Scholar] [CrossRef]
- El-Moamly, A.A.; El-Sweify, M.A.; Hafez, M.A. Using the AD12-ICT rapid-format test to detect Wuchereria bancrofti circulating antigens in comparison to Og4C3-ELISA and nucleopore membrane filtration and microscopy techniques. Parasitol. Res. 2012, 111, 1379–1383. [Google Scholar] [CrossRef] [PubMed]
- Reeve, D.; Melrose, W. Evaluation of the Og34C filter paper technique in lymphatic filariasis prevalence studies. Lymphology 2014, 47, 65–72. [Google Scholar] [PubMed]
- Supali, T.; Ismid, I.S.; Wibowo, H.; Djuardi, Y.; Majawati, E.; Ginanjar, P.; Fischer, P. Estimation of the prevalence of lymphatic filariasis by a pool screen PCR assay using blood spots collected on filter paper. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Debrah, A.Y.; Mand, S.; Marfo-Debrekyei, Y.; Batsa, L.; Pfarr, K.; Buttner, M.; Adjei, O.; Buttner, D.; Hoerauf, A. Macrofilaricidal effect of 4 weeks of treatment with doxycycline on Wuchereria bancrofti. Trop. Med. Int. Health 2007, 12, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Turba, M.E.; Zambon, E.; Zannoni, A.; Russo, S.; Gentilini, F. Detection of Wolbachia DNA in blood for diagnosing filaria-associated syndromes in cats. J. Clin. Microbiol. 2012, 50, 2624–2630. [Google Scholar] [CrossRef] [PubMed]
- Bockarie, M.J.; Tavul, L.; Kastens, W.; Michael, E.; Kazura, J.W. Impact of untreated bednets on prevalence of Wuchereria bancrofti transmitted by Anopheles farauti in Papua New Guinea. Med. Vet. Entomol. 2002, 16, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Reimer, L.J.; Thomsen, E.K.; Tisch, D.J.; Henry-Halldin, C.N.; Zimmerman, P.A.; Baea, M.E.; Dagoro, H.; Susapu, M.; Hetzel, M.W.; Bockarie, M.J.; et al. Insecticidal bed nets and filariasis transmission in Papua New Guinea. N. Engl. J. Med. 2013, 369, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.W.; Klein, D.M.; Dornink, S.M.; Jespersen, D.J.; Kubofcik, J.; Nutman, T.B.; Merrigan, S.D.; Couturier, M.R.; Theel, E.S. Comparison of three immunoassays for detection of antibodies to Strongyloides stercoralis. Clin. Vaccine Immunol. 2014, 21, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Ichimori, K. MDA-lymphatic filariasis. Trop. Med. Health 2014, 42, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Debrah, A.Y.; Mand, S.; Marfo-Debrekyei, Y.; Batsa, L.; Albers, A.; Specht, S.; Klarmann, U.; Pfarr, K.; Adjei, O.; Hoerauf, A. Macrofilaricidal activity in Wuchereria bancrofti after 2 weeks treatment with a combination of rifampicin plus doxycycline. J. Parasitol. Res. 2011, 2011, 201617. [Google Scholar] [CrossRef] [PubMed]
- Noroes, J.; Dreyer, G.; Santos, A.; Mendes, V.G.; Medeiros, Z.; Addiss, D. Assessment of the efficacy of diethylcarbamazine on adult Wuchereria bancrofti in vivo. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 78–81. [Google Scholar] [CrossRef]
- Vanamail, P.; Ramaiah, K.D.; Pani, S.P.; Das, P.K.; Grenfell, B.T.; Bundy, D.A. Estimation of the fecund life span of Wuchereria bancrofti in an endemic area. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 119–121. [Google Scholar] [CrossRef]
- WHO. WHO Recommends Triple Drug Therapy to Accelerate Global Elimination of Lymphatic Filariasis. Available online: http://www.who.int/neglected_diseases/news/WHO_recommends_triple_medicine_therapy_for_LF_elimination/en/ (accessed on 25 May 2018).
- Lim, K.H.; Speare, R.; Thomas, G.; Graves, P. Surgical treatment of genital manifestations of lymphatic filariasis: A systematic review. World J. Surg. 2015, 39, 2885–2899. [Google Scholar] [CrossRef] [PubMed]
- Foster, W.D. A History of Parasitology; E. & S. Livingstone LTD.: Edinburgh/London, UK, 1965; pp. 89–104. [Google Scholar]
- Peard, J.G. Race, Place, and Medicine: The Idea of the Tropics in Nineteenth-Century Brazil: The Politics of Disease; Duke University Press: Durham, NC, USA, 1999. [Google Scholar]
- Cobbold, T.S. On the discovery of the intermediate host of Filaria sanguinis hominis. Lancet 1878, 1, 69. [Google Scholar]
- Bahiana. Heróis da Saúde na Bahia: Otto Edwar Heinrich Wucherer. Available online: http://www.bahiana.edu.br/herois/heroi.aspx?id=MQ== (accessed on 10 January 2018).
- Bancroft, P. Discussion of a paper by E. S. Jackson. Aust. Med. Gaz. 1893, 12, 261–262. [Google Scholar]
- Bancroft, J. Diseases of animals and plants that interfere with colonial progress. Divin. Hall Rec. 1879, 1, 1–14. [Google Scholar]
- Sweet, W.C. Report on malaria and filaria survey of Australia and on mosquito surveys in Queensland, Western Australia and Northern Territory. In Final Report Australian Hookworm Campaign; Australian Hookworm Campaign: Brisbane, Australia, 1924; Part II; pp. 1–37. [Google Scholar]
- Mackerras, I.M.; Marks, E.N. The Bancrofts: A century of scientific endeavour. Proc. R. Soc. Qld. 1973, 84, 1–34. [Google Scholar]
- Cobbold, T.S. Discovery of the adult representative of microscopic filariae. Lancet 1877, 2, 70–71. [Google Scholar]
- Cobbold, T.S. On filaria bancrofti. Lancet 1877, 2, 495–496. [Google Scholar] [CrossRef]
- Cobbold, T.S. Verification of recent haematozoal discoveries in Australia and Egypt. Br. Med. J. 1876, 1, 780. [Google Scholar] [CrossRef]
- Bourne, A.G. A note on Filaria sanguinis hominis: With a description of a male specimen. Br. Med. J. 1888, 1, 1050. [Google Scholar] [CrossRef] [PubMed]
- Trewin, B.J.; Darbro, J.M.; Jansen, C.C.; Schellhorn, N.A.; Zalucki, M.P.; Hurst, T.P.; Devine, G.J. The elimination of the dengue vector, Aedes aegypti, from Brisbane, Australia: The role of surveillance, larval habitat removal and policy. PLoS Negl. Trop. Dis. 2017, 11, e0005848. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, T.L. Notes on filaria in Queensland. Aust. Med. Gaz. 1901, 29, 233–234. [Google Scholar]
- Bancroft, T.L. Filarial metamorphosis in the mosquito. Aust. Med. Gaz. 1899, 18, 120. [Google Scholar]
- Mackerras, I.M. Metamorphosis of Filaria bancrofti Cobbold. Br. Med. J. 1933, 2, 36. [Google Scholar] [CrossRef]
- Manson, P. On the development of Filaria sanguinis hominis, and on the mosquito considered as a nurse. J. Linn. Soc. Lond. Zool. 1878, 14, 304–311. [Google Scholar] [CrossRef]
- Bancroft, T.L.; Edin, M.B. On a proposed technique for the prevention of dengue fever and filariasis. Aust. Med. Gaz. 11912, 31, 80–81. [Google Scholar]
- Turner, P.; Copeman, B.; Gerisi, D.; Speare, R. A comparison of the Og4C3 antigen capture ELISA, the Knott test, an IgG4 assay and clinical signs, in the diagnosis of Bancroftian filariasis. Trop. Med. Parasitol. 1993, 44, 45–48. [Google Scholar] [PubMed]
- Burkot, T.R.; Durrheim, D.N.; Melrose, W.D.; Speare, R.; Ichimori, K. The argument for integrating vector control with multiple drug administration campaigns to ensure elimination of lymphatic filariasis. Filaria J. 2006, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Zeldenryk, L.M.; Gray, M.; Speare, R.; Gordon, S.; Melrose, W. The emerging story of disability associated with lymphatic filariasis: A critical review. PLoS Negl. Trop. Dis. 2011, 5, e1366. [Google Scholar] [CrossRef] [PubMed]
- Zeldenryk, L.; Gordon, S.; Gray, M.; Speare, R.; Melrose, W. Disability Measurement for Lymphatic Filariasis: A Review of Generic Tools Used within Morbidity Management Programs. PLoS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Durrheim, D.N.; Nelesone, T.; Speare, R.; Melrose, W. Certifying lymphatic filariasis elimination in the Pacific—The need for new tools. Pac. Health Dialog 2003, 10, 149–154. [Google Scholar] [PubMed]
- Harrington, H.; Asugeni, J.; Jimuru, C.; Gwalaa, J.; Ribeyro, E.; Bradbury, R.; Joseph, H.; Melrose, W.; MacLaren, D.; Speare, R. A practical strategy for responding to a case of lymphatic filariasis post-elimination in Pacific islands. Parasit. Vectors 2013, 6, 218. [Google Scholar] [CrossRef] [PubMed]
- Michael, E.; Bundy, D.A. Global mapping of lymphatic filariasis. Parasitol. Today 1997, 13, 472–476. [Google Scholar] [CrossRef]
- JCU. James Cook University: WHO Collaborating Centres. Available online: https://research.jcu.edu.au/who-collaborating-centres (accessed on 20 January 2018).
- Bockarie, M.J.; Tisch, D.J.; Kastens, W.; Alexander, N.D.; Dimber, Z.; Bockarie, F.; Ibam, E.; Alpers, M.P.; Kazura, J.W. Mass treatment to eliminate filariasis in Papua New Guinea. N. Engl. J. Med. 2002, 347, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Erickson, S.M.; Thomsen, E.K.; Keven, J.B.; Vincent, N.; Koimbu, G.; Siba, P.M.; Christensen, B.M.; Reimer, L.J. Mosquito-parasite interactions can shape filariasis transmission dynamics and impact elimination programs. PLoS Negl. Trop. Dis. 2013, 7, e2433. [Google Scholar] [CrossRef] [PubMed]
- Tisch, D.J.; Alexander, N.D.; Kiniboro, B.; Dagoro, H.; Siba, P.M.; Bockarie, M.J.; Alpers, M.P.; Kazura, J.W. Reduction in acute filariasis morbidity during a mass drug administration trial to eliminate lymphatic filariasis in Papua New Guinea. PLoS Negl. Trop. Dis. 2011, 5, e1241. [Google Scholar] [CrossRef] [PubMed]
- Tisch, D.J.; Bockarie, M.J.; Dimber, Z.; Kiniboro, B.; Tarongka, N.; Hazlett, F.E.; Kastens, W.; Alpers, M.P.; Kazura, J.W. Mass drug administration trial to eliminate lymphatic filariasis in Papua New Guinea: Changes in microfilaremia, filarial antigen, and Bm14 antibody after cessation. Am. J. Trop. Med. Hyg. 2008, 78, 289–293. [Google Scholar] [PubMed]
- Weil, G.J.; Kastens, W.; Susapu, M.; Laney, S.J.; Williams, S.A.; King, C.L.; Kazura, J.W.; Bockarie, M.J. The impact of repeated rounds of mass drug administration with diethylcarbamazine plus albendazole on bancroftian filariasis in Papua New Guinea. PLoS Negl. Trop. Dis. 2008, 2, e344. [Google Scholar] [CrossRef] [PubMed]
- WHO. Pacific Program to Eliminate Lymphatic Filariasis: Country Programs. Available online: http://www.wpro.who.int/southpacific/pacelf/countries/en/ (accessed on 25 January 2018).
- Hotez, P.J.; Bottazzi, M.E.; Strych, U.; Chang, L.Y.; Lim, Y.A.; Goodenow, M.M.; AbuBakar, S. Neglected tropical diseases among the Association of Southeast Asian Nations (ASEAN): Overview and update. PLoS Negl. Trop. Dis. 2015, 9, e0003575. [Google Scholar] [CrossRef] [PubMed]
- Wibawa, T.; Satoto, T.B.T. Magnitude of neglected tropical diseases in Indonesia at postmillennium Development Goals Era. J. Trop. Med. 2016, 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Oemijati, S. Current status of filariasis in Indonesia. Southeast Asian J. Trop. Med. Public Health 1993, 24 (Suppl. 2), 2–4. [Google Scholar] [PubMed]
- Wahyono, T.Y.M.; Purwantyastuti; Supali, T. Filariasis di Indonesia; Buletin Jendela Epidemiologi: Jakarta, Indonesia, 2010. [Google Scholar]
- 2015 Indonesian Health Profile; Ministry of Health RI: Jakarta, Indonesia, 2016.
- Krentel, A.; Damayanti, R.; Titaley, C.R.; Suharno, N.; Bradley, M.; Lynam, T. Improving coverage and compliance in mass drug administration for the elimination of LF in two ‘endgame’ districts in Indonesia using micronarrative surveys. PLoS Negl. Trop. Dis. 2016, 10, e0005027. [Google Scholar] [CrossRef] [PubMed]
- Krentel, A.; Fischer, P.; Manoempil, P.; Supali, T.; Servais, G.; Ruckert, P. Using knowledge, attitudes and practice (KAP) surveys on lymphatic filariasis to prepare a health promotion campaign for mass drug administration in Alor District, Indonesia. Trop. Med. Int. Health 2006, 11, 1731–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, N.; McMinn, P.; de Jesus Gomes, M.S.; Freitas, L.T.; Counahan, M.; Freitas, C. Timor-Leste National Parasite Survey: Report and Recommendations; Ministerio da Saude: Dili, Timor-Leste, 2012.
- Frances, S.P.; Baade, L.M.; Kubofcik, J.; Nutman, T.B.; Melrose, W.D.; McCarthy, J.S.; Nissen, M.D. Seroconversion to filarial antigens in Australian defence force personnel in Timor-Leste. Am. J. Trop. Med. Hyg. 2008, 78, 560–563. [Google Scholar] [PubMed]
- Lau, C.L.; Won, K.Y.; Becker, L.; Soares Magalhaes, R.J.; Fuimaono, S.; Melrose, W.; Lammie, P.J.; Graves, P.M. Seroprevalence and spatial epidemiology of lymphatic filariasis in American Samoa after successful mass drug administration. PLoS Negl. Trop. Dis. 2014, 8, e3297. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.L.; Sheridan, S.; Ryan, S.; Roineau, M.; Andreosso, A.; Fuimaono, S.; Tufa, J.; Graves, P.M. Detecting and confirming residual hotspots of lymphatic filariasis transmission in American Samoa 8 years after stopping mass drug administration. PLoS Negl. Trop. Dis. 2017, 11, e0005914. [Google Scholar] [CrossRef] [PubMed]
- Monchy, D.; Barny, S.; Rougier, Y.; Baudet, J.M.; Gentile, B. Survey of lymphatic filariasis on Ouvea Island in New Caledonia. Med. Trop. 1999, 59, 146–150. [Google Scholar]
- Daures, M.; Champagnat, J.; Pfannstiel, A.; Ringuenoire, F.; Grangeon, J.P.; Musso, D. Filariasis serosurvey, New Caledonia, South Pacific, 2013. Parasit. Vectors 2015, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Pretrick, M.; Melrose, W.; Chaine, J.P.; Canyon, D.; Carron, J.; Graves, P.M.; Bradbury, R.S. Identification and control of an isolated, but intense focus of lymphatic filariasis on Satawal Island, Federated States of Micronesia, in 2003. Trop. Med. Health 2017, 45, 17. [Google Scholar] [CrossRef] [PubMed]
- WHO. Weekly Epidemiological Record; WHO: Geneva, Switzerland, 2010; pp. 365–372. [Google Scholar]
- Fraser, M.; Taleo, G.; Taleo, F.; Yaviong, J.; Amos, M.; Babu, M.; Kalkoa, M. Evaluation of the program to eliminate lymphatic filariasis in Vanuatu following two years of mass drug administration implementation: Results and methodologic approach. Am. J. Trop. Med. Hyg. 2005, 73, 753–758. [Google Scholar] [PubMed]
- Allen, T.; Taleo, F.; Graves, P.M.; Wood, P.; Taleo, G.; Baker, M.C.; Bradley, M.; Ichimori, K. Impact of the Lymphatic Filariasis Control Program towards elimination of filariasis in Vanuatu, 1997–2006. Trop. Med. Health 2017, 45, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taleo, F.; Taleo, G.; Graves, P.M.; Wood, P.; Kim, S.H.; Ozaki, M.; Joseph, H.; Chu, B.; Pavluck, A.; Yajima, A.; et al. Surveillance efforts after mass drug administration to validate elimination of lymphatic filariasis as a public health problem in Vanuatu. Trop. Med. Health 2017, 45, 18. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cambodia, Cook Islands, Niue and Vanuatu Eliminate Lymphatic Filariasis as a Public Health Problem; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Australian Bureau of Statistics. Migration, Australia, 2015–16. Available online: http://www.abs.gov.au/ausstats/[email protected]/mf/3412.0 (accessed on 4 July 2017).
- Lowichik, A.; Lowrie, R.C., Jr. Uptake and development of Wuchereria bancrofti in Aedes aegypti and Haitian Culex quinquefasciatus that were fed on a monkey with low-density microfilaremia. Trop. Med. Parasitol. 1988, 39, 227–229. [Google Scholar] [PubMed]
- Queensland Government. Dengue Outbreaks. Available online: https://www.health.qld.gov.au/clinical-practice/guidelines-procedures/diseases-infection/diseases/mosquito-borne/dengue/dengue-outbreaks (accessed on 28 January 2018).
- Queensland Government. Queensland Dengue Management Plan 2015–2020; State of Queensland (Queensland Health): Brisbane, Australia, 2015.
- Queensland Government. Mosquito Control. Available online: https://www.qld.gov.au/health/conditions/all/prevention/mosquito-borne/control/breeding-sites (accessed on 28 January 2018).
- Ciota, A.; Kramer, L. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef] [PubMed]
- Kovendan, K.; Murugan, K.; Vincent, S.; Kamalakannan, S. Larvicidal efficacy of Jatropha curcas and bacterial insecticide, Bacillus thuringiensis, against lymphatic filarial vector, Culex quinquefasciatus Say (Diptera: culicidae). Parasitol. Res. 2011, 109, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Muzari, M.O.; Devine, G.; Davis, J.; Crunkhorn, B.; van den Hurk, A.; Whelan, P.; Russell, R.; Walker, J.; Horne, P.; Ehlers, G.; et al. Holding back the tiger: Successful control program protects Australia from Aedes albopictus expansion. PLoS Negl. Trop. Dis. 2017, 11, e0005286. [Google Scholar] [CrossRef] [PubMed]
- Proestos, Y.; Christophides, G.K.; Erguler, K.; Tanarhte, M.; Waldock, J.; Lelieveld, J. Present and future projections of habitat suitability of the Asian tiger mosquito, a vector of viral pathogens, from global climate simulation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef]
- Southgate, B.A. Intensity and efficiency of transmission and the development of microfilaraemia and disease: Their relationship in lymphatic filariasis. J. Trop. Med. Hyg. 1992, 95, 1–12. [Google Scholar] [PubMed]
- Subramanian, S.; Krishnamoorthy, K.; Ramaiah, K.D.; Habbema, J.D.; Das, P.K.; Plaisier, A.P. The relationship between microfilarial load in the human host and uptake and development of Wuchereria bancrofti microfilariae by Culex quinquefasciatus: A study under natural conditions. Parasitology 1998, 116 Pt 3, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Bryan, J.H.; McMahon, P.; Barnes, A. Factors affecting transmission of Wuchereria bancrofti by anopheline mosquitoes. 3. Uptake and damage to ingested microfilariae by Anopheles gambiae, An. arabiensis, An. merus and An. funestus in east Africa. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 265–268. [Google Scholar] [CrossRef]
- Bryan, J.H.; Southgate, B.A. Factors affecting transmission of Wuchereria bancrofti by anopheline mosquitoes. 1. Uptake of microfilariae. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 128–137. [Google Scholar] [CrossRef]
- Bryan, J.H.; Southgate, B.A. Factors affecting transmission of Wuchereria bancrofti by anopheline mosquitoes. 2. Damage to ingested microfilariae by mosquito foregut armatures and development of filarial larvae in mosquitoes. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 138–145. [Google Scholar] [CrossRef]
- Southgate, B.A.; Bryan, J.H. Factors affecting transmission of Wuchereria bancrofti by anopheline mosquitoes. 4. Facilitation, limitation, proportionality and their epidemiological significance. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 523–530. [Google Scholar] [CrossRef]
- Boakye, D.A.; Wilson, M.D.; Appawu, M.A.; Gyapong, J. Vector competence, for Wuchereria bancrofti, of the Anopheles populations in the Bongo district of Ghana. Ann. Trop. Med. Parasitol. 2004, 98, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Manson, P. The Filaria Sanguinis Hominis and Certain Forms of Parasitic Disease in India, China and Warm Countries; H. K. Lewis: London, UK, 1883. [Google Scholar]
- Shriram, A.N.; Ramaiah, K.D.; Krishnamoorthy, K.; Sehgal, S.C. Diurnal pattern of human-biting activity and transmission of subperiodic Wuchereria bancrofti (Filariidea: Dipetalonematidae) by Ochlerotatus niveus (Diptera: Culicidae) on the Andaman and Nicobar islands of India. Am. J. Trop. Med. Hyg. 2005, 72, 273–277. [Google Scholar] [PubMed]
- Samarawickrema, W.A.; Kimura, E.; Spears, G.F.; Penaia, L.; Sone, F.; Paulson, G.S.; Cummings, R.F. Distribution of vectors, transmission indices and microfilaria rates of subperiodic Wuchereria bancrofti in relation to village ecotypes in Samoa. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 129–135. [Google Scholar] [CrossRef]
- Russell, R.C.; Webb, C.E.; Davies, N. Aedes aegypti (L.) and Aedes polynesiensis Marks (Diptera: Culicidae) in Moorea, French Polynesia: A Study of adult population structures and pathogen (Wuchereria bancrofti and Dirofilaria immitis) infection rates to indicate regional and seasonal epidemiological risk for dengue and filariasis. J. Med. Entomol. 2005, 42, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Ariani, C.V.; Juneja, P.; Smith, S.; Tinsley, M.C.; Jiggins, F.M. Vector competence of Aedes aegypti mosquitoes for filarial nematodes is affected by age and nutrient limitation. Exp. Gerontol. 2015, 61, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Beebe, N.W.; Cooper, R.D.; Mottram, P.; Sweeney, A.W. Australia’s dengue risk driven by human adaptation to climate change. PLoS Negl. Trop. Dis. 2009, 3, e429. [Google Scholar] [CrossRef] [PubMed]
- Richard, V.; Paoaafaite, T.; Cao-Lormeau, V.M. Vector competence of Aedes aegypti and Aedes polynesiensis populations from French Polynesia for chikungunya virus. PLoS Negl. Trop. Dis. 2016, 10, e0004694. [Google Scholar] [CrossRef] [PubMed]
- Gilotra, S.K.; Shah, K.V. Laboratory studies on transmission of chikungunya virus by mosquitoes. Am. J. Epidemiol. 1967, 86, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Transmission of Ross River virus by Aedes polynesiensis and Aedes aegypti. Am. J. Trop. Med. Hyg. 1981, 30, 1303–1306. [Google Scholar] [CrossRef] [PubMed]
- Rollins, A. TB Superbug Makes Landfall in Australia. Available online: https://ama.com.au/ausmed/tb-superbug-makes-landfall-australia (accessed on 29 March 2018).



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, C.A.; Jones, M.K.; McManus, D.P. The History of Bancroftian Lymphatic Filariasis in Australasia and Oceania: Is There a Threat of Re-Occurrence in Mainland Australia? Trop. Med. Infect. Dis. 2018, 3, 58. https://doi.org/10.3390/tropicalmed3020058
Gordon CA, Jones MK, McManus DP. The History of Bancroftian Lymphatic Filariasis in Australasia and Oceania: Is There a Threat of Re-Occurrence in Mainland Australia? Tropical Medicine and Infectious Disease. 2018; 3(2):58. https://doi.org/10.3390/tropicalmed3020058
Chicago/Turabian StyleGordon, Catherine A., Malcolm K. Jones, and Donald P. McManus. 2018. "The History of Bancroftian Lymphatic Filariasis in Australasia and Oceania: Is There a Threat of Re-Occurrence in Mainland Australia?" Tropical Medicine and Infectious Disease 3, no. 2: 58. https://doi.org/10.3390/tropicalmed3020058
APA StyleGordon, C. A., Jones, M. K., & McManus, D. P. (2018). The History of Bancroftian Lymphatic Filariasis in Australasia and Oceania: Is There a Threat of Re-Occurrence in Mainland Australia? Tropical Medicine and Infectious Disease, 3(2), 58. https://doi.org/10.3390/tropicalmed3020058