Asian Schistosomiasis: Current Status and Prospects for Control Leading to Elimination

 , , , and
, , , and
Abstract
:1. Introduction
2. Parasite Features
2.1. Lifecycle
2.1.1. S. japonicum
2.1.2. S. mekongi
2.1.3. S. malayensis
2.2. Clinical Features
2.3. Diagnostics
2.4. Treatment
3. Epidemiology
3.1. Mammalian Definitive Hosts
3.2. Molluscan Intermediate Hosts
3.3. Environment
3.4. Transmission and Control
4. Current and Historical Status of Schistosomiasis in Asia
4.1. S. japonicum
4.1.1. Japan
4.1.2. P.R. China
4.1.3. Philippines
4.1.4. Indonesia
4.1.5. Myanmar
4.2. S. mekongi
4.2.1. Cambodia
4.2.2. Thailand
4.2.3. Lao PDR
4.2.4. Myanmar
4.3. S. malayensis
Malaysia
5. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.N. Schistosomiasis. Nat. Rev. Dis. Primers 2018, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Boissier, J.; Grech-Angelini, S.; Webster, B.L.; Allienne, J.F.; Huyse, T.; Mas-Coma, S.; Toulza, E.; Barre-Cardi, H.; Rollinson, D.; Kincaid-Smith, J.; et al. Outbreak of urogenital schistosomiasis in Corsica (France): An epidemiological case study. Lancet Infect. Dis. 2016, 16, 971–979. [Google Scholar] [CrossRef]
- Tchuem Tchuente, L.A.; Southgate, V.R.; Jourdane, J.; Webster, B.L.; Vercruysse, J. Schistosoma intercalatum: An endangered species in Cameroon? Trends Parasitol. 2003, 19, 389–393. [Google Scholar] [CrossRef]
- Greer, G.J.; Ow-Yang, C.K.; Yong, H.-S. Schistosoma malayensis n. sp.: A Schistosoma japonicum-complex schistosome from Peninsular Malaysia. J. Parasitol. 1988, 74, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Webster, B.L.; Diaw, O.T.; Seye, M.M.; Webster, J.P.; Rollinson, D. Introgressive hybridization of Schistosoma haematobium group species in Senegal: Species barrier break down between ruminant and human schistosomes. PLoS Negl. Trop. Dis. 2013, 7, e2110. [Google Scholar] [CrossRef] [PubMed]
- Leger, E.; Webster, J.P. Hybridizations within the genus Schistosoma: Implications for evolution, epidemiology and control. Parasitology 2017, 144, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Kebede, T.; Negash, Y.; Erko, B. Schistosoma mansoni infection in human and nonhuman primates in selected areas of Oromia Regional State, Ethiopia. J. Vector Borne Dis. 2018, 55, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Alarcón de Noya, B.; Pointier, J.P.; Colmenares, C.; Théron, A.; Balzan, C.; Cesari, I.M.; González, S.; Noya, O. Natural Schistosoma mansoni infection in wild rats from Guadeloupe: Parasitological and immunological aspects. Acta Trop. 1997, 68, 11–21. [Google Scholar] [CrossRef]
- Huyse, T.; Van den Broeck, F.; Hellemans, B.; Volckaert, F.A.M.; Polman, K. Hybridisation between the two major African schistosome species of humans. Int. J. Parasitol. 2013, 43, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Leger, E.; Garba, A.; Hamidou, A.A.; Webster, B.L.; Pennance, T.; Rollinson, D.; Webster, J.P. Introgressed animal schistosomes Schistosoma curassoni and S. bovis naturally infecting humans. Emerg. Infect. Dis. 2016, 22, 2212–2214. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.N.; Bergquist, R.; Leonardo, L.; Yang, G.J.; Yang, K.; Sudomo, M.; Olveda, R. Schistosomiasis japonica: Control and research needs. Adv. Parasitol. 2010, 72, 145–178. [Google Scholar] [PubMed]
- Kajihara, N.; Hirayama, K. The war against a regional disease in Japan: A history of the eradication of schistosomiasis japonica. Trop. Med. Health 2011, 39, 3–44. [Google Scholar] [PubMed]
- Ross, A.G.P.; Sleigh, A.C.; Li, Y.; Davis, G.M.; Williams, G.; Jiang, Z.; Feng, Z.; McManus, D.P. Schistosomiasis in the People’s Republic of China: Prospects and challenges for the 21st century. Clin. Microbiol. Rev. 2001, 14, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.P.; Shao, B.R. Schistosomiasis control in the People’s Republic of China. Am. J. Trop. Med. Hyg. 1982, 31, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Rollinson, D.; Knopp, S.; Levitz, S.; Stothard, J.R.; Tchuem Tchuenté, L.-A.; Garba, A.; Mohammed, K.A.; Schur, N.; Person, B.; Colley, D.G.; et al. Time to set the agenda for schistosomiasis elimination. Acta Trop. 2013, 128, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Soe, H.Z.; Oo, C.C.; Myat, T.O.; Maung, N.S. Detection of Schistosoma antibodies and exploration of associated factors among local residents around Inlay Lake, Southern Shan State, Myanmar. Infect. Dis. Poverty 2017, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- WHO. Expert consultation to accelerate elimination of Asian schistosomiasis. In Meeting Report WHO; WHO: Shanghai, China, 2017. [Google Scholar]
- Cameron, T.W.M. A new definitive host for Schistosoma mansoni. J. Helminthol. 1928, 6, 219–222. [Google Scholar] [CrossRef]
- He, Y.; Salafsky, B.; Ramaswamy, K. Host-parasite relationships of Schistosoma japonicum in mammalian hosts. Trends Parasitol. 2001, 17, 320–324. [Google Scholar] [CrossRef]
- Ho, Y.H.; He, Y.X. On the host specificity of Schistosoma japonicum. Chin. Med. J. 1963, 82, 403–414. [Google Scholar]
- Bustinduy, A.L.; King, C.H. 52—Schistosomiasis A2—Farrar, Jeremy. In Manson’s Tropical Infectious Diseases, 23th ed.; Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; W.B. Saunders: London, UK, 2014. [Google Scholar]
- Matsumoto, J.; Muth, S.; Socheat, D.; Matsuda, H. The first reported cases of canine schistosomiasis mekongi in Cambodia. Southeast Asian J. Trop. Med. Public Health 2002, 33, 458–461. [Google Scholar] [PubMed]
- Strandgaard, H.; Johansen, M.V.; Pholsena, K.; Teixayavong, K.; Christensen, N.O. The pig as a host for Schistosoma mekongi in Laos. J. Parasitol. 2001, 87, 708–709. [Google Scholar] [CrossRef]
- Blair, D.; van Herwerden, L.; Hirai, H.; Taguchi, T.; Habe, S.; Hirata, M.; Lai, K.; Upatham, S.; Agatsuma, T. Relationships between Schistosoma malayensis and other Asian schistosomes deduced from DNA sequences. Mol. Biochem. Parasitol. 1997, 85, 259–263. [Google Scholar] [CrossRef]
- Attwood, S.W.; Cottet, M. Malacological and parasitological surveys along the Xe Bangfai and its tributaries in Khammouane Province, Lao PDR. Hydroécol. Appl. 2016, 19, 245–270. [Google Scholar] [CrossRef]
- CDC. Schistosomiasis Infection. Available online: https://www.cdc.gov/dpdx/schistosomiasis/index.html (accessed on 2 March 2018).
- Utzinger, J.; N’Goran, E.K.; Caffrey, C.R.; Keiser, J. From innovation to application: Social-ecological context, diagnostics, drugs and integrated control of schistosomiasis. Acta Trop. 2011, 120 (Suppl. 1), S121–S137. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Acosta, L.P.; Gobert, G.N.; Jiz, M.; Olveda, R.M.; Ross, A.G.; Gray, D.J.; Williams, G.M.; Harn, D.; Yuesheng, L.; et al. High prevalence of Schistosoma japonicum and Fasciola gigantica in bovines from Northern Samar, the Philippines. PLoS Negl. Trop. Dis. 2015, 9, e0003108. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Williams, G.M.; Li, Y.; McManus, D.P. Transmission dynamics of Schistosoma japonicum in the lakes and marshlands of China. PLoS ONE 2008, 3, e4058. [Google Scholar] [CrossRef] [PubMed]
- Rudge, J.W.; Carabin, H.; Balolong, E.; Tallo, V.; Shrivastava, J.; Lu, D.B.; Basanez, M.G.; Olveda, R.; McGarvey, S.T.; Webster, J.P. Population genetics of Schistosoma japonicum within the Philippines suggest high levels of transmission between humans and dogs. PLoS Negl. Trop. Dis. 2008, 2, e340. [Google Scholar] [CrossRef] [PubMed]
- Rudge, J.W.; Lu, D.-B.; Fang, G.-R.; Wang, T.-P.; Basáñez, M.-G.; Webster, J.P. Parasite genetic differentiation by habitat type and host species: Molecular epidemiology of Schistosoma japonicum in hilly and marshland areas of Anhui Province, China. Mol. Ecol. 2009, 18, 2134–2147. [Google Scholar] [CrossRef] [PubMed]
- Rudge, J.W.; Webster, J.P.; Lu, D.-B.; Wang, T.-P.; Fang, G.-R.; Basáñez, M.-G. Identifying host species driving transmission of schistosomiasis japonica, a multihost parasite system, in China. Proc. Natl. Acad. Sci. USA 2013, 110, 11457–11462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attwood, S.W.; Fatih, F.A.; Upatham, E.S. DNA-sequence variation among Schistosoma mekongi populations and related taxa: Phylogeography and the current distribution of Asian schistosomiasis. PLoS Negl. Trop. Dis. 2008, 2, e200. [Google Scholar] [CrossRef] [PubMed]
- Sornmani, S.; Kitikoon, V.; Thirachantra, S.; Harinasuta, C. Epidemiology of mekong schistosomiasis. Malacol. Rev. 1980, (Suppl. 2), 9–18. [Google Scholar]
- Kitikoon, V. Comparison of eggs and miracidia of Schistosoma mekongi and S. japonicum. Malacol. Rev. 1980, (Suppl. 2), 93–103. [Google Scholar]
- Fletcher, M.; Woodruff, D.S.; LoVerde, P.T. Genetic differentiation between Schistosoma mekongi and S. japonicum: An electrophoretic study. Malacol. Rev. 1980, 2, 113–122. [Google Scholar]
- Ross, A.G.; Vickers, D.; Olds, G.R.; Shah, S.M.; McManus, D.P. Katayama syndrome. Lancet Infect. Dis. 2007, 7, 218–224. [Google Scholar] [CrossRef]
- Gryseels, B. Schistosomiasis. Infect. Dis. Clin. N. Am. 2012, 26, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Gryseels, B.; Polman, K.; Clerinx, J.; Kestens, L. Human schistosomiasis. Lancet 2006, 368, 1106–1118. [Google Scholar] [CrossRef]
- Li, Y.; Ross, A.G.; Hou, X.; Lou, Z.; McManus, D.P. Oriental schistosomiasis with neurological complications: Case report. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Kane, C.A.; Most, H. Schistosomiasis of the central nervous system: Experiences in world war II and a review of the literature. Arch. Neurol. Psychiatry 1948, 59, 141–183. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, T.C.A.; Moreira, P.R.R. Neuroschistosomiasis: Clinical symptoms and pathogenesis. Lancet Neurol. 2011, 10, 853–864. [Google Scholar] [CrossRef]
- Liu, H.; Lim, C.C.; Feng, X.; Yao, Z.; Chen, Y.; Sun, H.; Chen, X. MRI in cerebral schistosomiasis: Characteristic nodular enhancement in 33 patients. Am. J. Roentgenol. 2008, 191, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Stecher, C.W.; Sacko, M.; Madsen, H.; Wilson, S.; Wejse, C.; Keita, A.D.; Landoure, A.; Traore, M.S.; Kallestrup, P.; Petersen, E.; et al. Anemia and growth retardation associated with Schistosoma haematobium infection in Mali: A possible subtle impact of a neglected tropical disease. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 144–153. [Google Scholar] [CrossRef] [PubMed]
- WHO. Schistosomiasis: Fact Sheet. Available online: http://www.who.int/mediacentre/factsheets/fs115/en/ (accessed on 20 February 2018).
- Glinz, D.; Silué, K.D.; Knopp, S.; Lohouringnon, L.K.; Yao, K.P.; Steinmann, P.; Rinaldi, L.; Cringoli, G.; N’Goran, E.K.; Utzinger, J. Comparing diagnostic accuracy of Kato-Katz, Koga agar plate, ether-concentration, and FLOTAC for Schistosoma mansoni and soil-transmitted helminths. PLoS Negl. Trop. Dis. 2010, 4, e754. [Google Scholar] [CrossRef] [PubMed]
- Habtamu, K.; Degarege, A.; Ye-Ebiyo, Y.; Erko, B. Comparison of the Kato-Katz and FLOTAC techniques for the diagnosis of soil-transmitted helminth infections. Parasitol. Int. 2011, 60, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Cringoli, G. FLOTAC, a novel apparatus for a multivalent faecal egg count technique. Parassitologica 2006, 48, 381–384. [Google Scholar]
- Cringoli, G.; Rinaldi, L.; Maurelli, M.P.; Utzinger, J. FLOTAC: New multivalent techniques for qualitative and quantitative copromicroscopic diagnosis of parasites in animals and humans. Nat. Protoc. 2010, 5, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gordon, C.A.; Hu, W.; McManus, D.P.; Chen, H.; Gray, D.J.; Ju, C.; Zeng, X.; Gobert, G.N.; Ge, J.; et al. A novel procedure for precise quantification of Schistosoma japonicum eggs in bovine feces. PLoS Negl. Trop. Dis. 2012, 6, e1885. [Google Scholar] [CrossRef]
- Lier, T.; Johansen, M.V.; Hjelmevoll, S.O.; Vennervald, B.J.; Simonsen, G.S. Real-time PCR for detection of low intensity Schistosoma japonicum infections in a pig model. Acta Trop. 2008, 105, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Anh, N.T.; Phuong, N.T.; Ha, G.H.; Thu, L.T.; Johansen, M.V.; Murrell, D.K.; Thamsborg, S.M. Evaluation of techniques for detection of small trematode eggs in faeces of domestic animals. Vet. Parasitol. 2008, 156, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.Q.; Xu, J.; Zhu, R.; Cao, C.L.; Bao, Z.P.; Yu, Q.; Zhang, L.J.; Xu, X.L.; Feng, Z.; Guo, J.G. Comparison of the miracidium hatching test and modified Kato-Katz method for detecting Schistosoma japonicum in low prevalence areas of China. Southeast Asian J. Trop. Med. Public Health 2014, 45, 20–25. [Google Scholar] [PubMed]
- Jurberg, A.D.; de Oliveira, A.A.; Lenzi, H.L.; Coelho, P.M.Z. A new miracidia hatching device for diagnosing schistosomiasis. Mem. Inst. Oswaldo Cruz 2008, 103, 112–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Rong, R.; Zhang, H.Q.; Shi, C.J.; Zhu, X.Q.; Xia, C.M. Sensitive and rapid detection of Schistosoma japonicum DNA by loop-mediated isothermal amplification (LAMP). Int. J. Parasitol. 2010, 40, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Furushima-Shimogawara, R.; Ohmae, H.; Wang, T.; Lu, S.; Chen, R.; Wen, L.; Ohta, N. Detection of early and single infections of Schistosoma japonicum in the intermediate host snail, Oncomelania hupensis, by PCR and loop-mediated isothermal amplification (LAMP) assay. Am. J. Trop. Med. Hyg. 2010, 83, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Gray, D.J.; Gobert, G.N.; McManus, D.P. DNA amplification approaches for the diagnosis of key parasitic helminth infections of humans. Mol. Cell. Probes 2011, 25, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gobert, G.N.; Cai, P.; McManus, D.P. Advances in the diagnosis of human schistosomiasis. Clin. Microbiol. Rev. 2015, 28, 939–967. [Google Scholar] [CrossRef] [PubMed]
- Gobert, G.N.; Chai, M.; Duke, M.; McManus, D.P. Copro-PCR based detection of Schistosoma eggs using mitochondrial DNA markers. Mol. Cell. Probes 2005, 19, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Acosta, L.P.; Gray, D.J.; Olveda, R.; Jarilla, B.; Gobert, G.N.; Ross, A.G.; McManus, D.P. High prevalence of Schistosoma japonicum infection in carabao from Samar province, the Philippines: Implications for transmission and control. PLoS Negl. Trop. Dis. 2012, 6, e1778. [Google Scholar] [CrossRef] [PubMed]
- Lier, T.; Simonsen, G.S.; Haaheim, H.; Hjelmevoll, S.O.; Vennervald, B.J.; Johansen, M.V. Novel real-time PCR for detection of Schistosoma japonicum in stool. Southeast Asian J. Trop. Med. Public Health 2006, 37, 257–264. [Google Scholar] [PubMed]
- Lier, T.; Simonsen, G.S.; Wang, T.; Lu, D.; Haukland, H.H.; Vennervald, B.J.; Hegstad, J.; Johansen, M.V. Real-time polymerase chain reaction for detection of low-intensity Schistosoma japonicum infections in China. Am. J. Trop. Med. Hyg. 2009, 81, 428–432. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Gordon, C.A.; Williams, G.M.; Yueshang, L.; Wang, Y.; Hu, J.; Gray, D.J.; Ross, A.G.; Harn, D.; McManus, D.P. Real-time PCR diagnosis of Schistosoma japonicum in low transmission areas of China. Infect. Dis. Poverty 2018, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; Acosta, L.P.; Gobert, G.N.; Olveda, R.M.; Ross, A.G.; Williams, G.M.; Gray, D.J.; Harn, D.; Li, Y.; McManus, D.P. Real-time PCR demonstrates high prevalence of Schistosoma japonicum in the Philippines: Implications for surveillance and control. PLoS Negl. Trop. Dis. 2015, 9, e0003483. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; Cai, P.; Gobert, G.N.; Duke, M.; Williams, G.M.; McManus, D.P. A novel duplex ddPCR assay for the diagnosis of schistosomiasis japonica: Proof of concept in an experimental mouse model. Parasitology 2017, 144, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; Gobert, G.N.; Cai, P.; McManus, D.P. Optimisation of a droplet digital PCR assay for the diagnosis of Schistosoma japonicum infection: A duplex approach with DNA binding dye chemistry. J. Microbiol. Methods 2016, 125, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; Williams, G.M.; Cai, P.; Gobert, G.N.; Olveda, R.M.; Ross, A.G.; Olveda, D.U.; McManus, D.P. Droplet digital PCR diagnosis of human schistosomiasis: Parasite cell-free DNA detection in diverse clinical samples. J. Infect. Dis. 2017, 216, 1611–1622. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Weerakoon, K.G.; Mu, Y.; Olveda, D.U.; Piao, X.; Liu, S.; Olveda, R.M.; Chen, Q.; Ross, A.G.; McManus, D.P. A parallel comparison of antigen candidates for development of an optimized serological diagnosis of chistosomiasis japonica in the Philippines. EBioMedicine 2017, 24, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Angeles, J.M.; Goto, Y.; Kirinoki, M.; Asada, M.; Leonardo, L.R.; Rivera, P.T.; Villacorte, E.A.; Inoue, N.; Chigusa, Y.; Kawazu, S. Utilization of ELISA using thioredoxin peroxidase-1 and tandem repeat proteins for diagnosis of Schistosoma japonicum infection among water buffaloes. PLoS Negl. Trop. Dis. 2012, 6, e1800. [Google Scholar] [CrossRef] [PubMed]
- Dawson, E.M.; Sousa-Figueiredo, J.C.; Kabatereine, N.B.; Doenhoff, M.J.; Stothard, J.R. Intestinal schistosomiasis in pre school-aged children of Lake Albert, Uganda: Diagnostic accuracy of a rapid test for detection of anti-schistosome antibodies. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xu, J.; Zhang, Y.; Liu, Y.; Hu, F.; Xu, X.; Li, J.; Gao, Z.; Wu, H.; Kurtis, J.; et al. Evaluation of IgG-ELISA for the diagnosis of Schistosoma japonicum in a high prevalence, low intensity endemic area of China. Acta Trop. 2008, 107, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Y.; Li, H.; Xing, Y.; Qu, G.; Dai, J.; Liang, Y. Immunodiagnostic efficacy of detection of Schistosoma japonicum human infections in China: A meta analysis. Asian Pac. J. Trop. Med. 2012, 5, 15–23. [Google Scholar] [CrossRef]
- Hinz, R.; Schwarz, N.G.; Hahn, A.; Frickmann, H. Serological approaches for the diagnosis of schistosomiasis—A review. Mol. Cell. Probes 2017, 31, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.B.; Zheng, H.M.; Jiang, Q.W. A diagnostic challenge for Schistosomiasis japonica in China: Consequences on praziquantel-based morbidity control. Parasit. Vectors 2011, 4, 194. [Google Scholar] [CrossRef] [PubMed]
- Utzinger, J.; Becker, S.L.; van Lieshout, L.; van Dam, G.J.; Knopp, S. New diagnostic tools in schistosomiasis. Clin. Microbiol. Infect. 2015, 21, 529–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardo, L.; Chigusa, Y.; Kikuchi, M.; Kato-Hayashi, N.; Kawazu, S.I.; Angeles, J.M.; Fontanilla, I.K.; Tabios, I.K.; Moendeg, K.; Goto, Y.; et al. Schistosomiasis in the Philippines: Challenges and some successes in control. Southeast Asian J. Trop. Med. Public Health 2016, 47, 651–666. [Google Scholar]
- Gray, D.J.; Ross, A.G.; Li, Y.; McManus, D.P. Diagnosis and management of schistosomiasis. Br. Med. J. 2011, 342, d2651. [Google Scholar] [CrossRef] [PubMed]
- Utzinger, J.; Keiser, J. Schistosomiasis and soil-transmitted helminthiasis: Common drugs for treatment and control. Expert Opin. Pharmacother. 2004, 5, 263–285. [Google Scholar] [CrossRef] [PubMed]
- Vale, N.; Gouveia, M.J.; Rinaldi, G.; Brindley, P.J.; Gärtner, F.; Correia da Costa, J.M. Praziquantel for schistosomiasis: Single-drug metabolism revisited, mode of action, and resistance. Antimicrob. Agents Chemother. 2017, 61, e02582-16. [Google Scholar] [CrossRef] [PubMed]
- Sabah, A.A.; Fletcher, C.; Webbe, G.; Doenhoff, M.J. Schistosoma mansoni: Chemotherapy of infections of different ages. Exp. Parasitol. 1986, 61, 294–303. [Google Scholar] [CrossRef]
- Utzinger, J.; Keiser, J.; Xiao, S.H.; Tanner, M.; Singer, B.H. Combination chemotherapy of schistosomiasis in laboratory studies and clinical trials. Antimicrob. Agents Chemother. 2003, 47, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Olveda, D.U.; Li, Y.; Olveda, R.M.; Lam, A.K.; McManus, D.P.; Chau, T.N.; Harn, D.A.; Williams, G.M.; Gray, D.J.; Ross, A.G. Bilharzia in the Philippines: Past, present, and future. Int. J. Infect. Dis. 2014, 18, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Shaoji, Z.; Pan, B.; Hu, L.; Wei, R.; Gao, Z.; Li, J.; Uwe, B. Reinfection with Schistosoma japonicum after treatment with praziquantel in Poyang Lake Region, China. Southeast Asian J. Trop. Med. Public Health 1994, 25, 163–169. [Google Scholar] [PubMed]
- Wang, W.; Wang, L.; Liang, Y. Susceptibility or resistance of praziquantel in human schistosomiasis: A review. Parasitol. Res. 2012, 111, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Cupit, P.M.; Cunningham, C. What is the mechanism of action of praziquantel and how might resistance strike? Future Med. Chem. 2015, 7, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, R.; Utzinger, J.; Keiser, J. Controlling schistosomiasis with praziquantel: How much longer without a viable alternative? Infect. Dis. Poverty 2017, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Soares Magalhães, R.J.; Salamat, M.S.; Leonardo, L.; Gray, D.J.; Carabin, H.; Halton, K.; McManus, D.P.; Williams, G.M.; Rivera, P.; Saniel, O.; et al. Geographical distribution of human Schistosoma japonicum infection in The Philippines: Tools to support disease control and further elimination. Int. J. Parasitol. 2014, 44, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, L.R.; Rivera, P.; Saniel, O.; Villacorte, E.; Crisostomo, B.; Hernandez, L.; Baquilod, M.; Erce, E.; Martinez, R.; Velayudhan, R. Prevalence survey of schistosomiasis in Mindanao and the Visayas, the Philippines. Parasitol. Int. 2008, 57, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Williams, G.M.; Li, Y.; Chen, H.; Forsyth, S.J.; Li, R.S.; Barnett, A.G.; Guo, J.; Ross, A.G.; Feng, Z.; et al. A cluster-randomised intervention trial against Schistosoma japonicum in the People’s Republic of China: Bovine and human transmission. PLoS ONE 2009, 4, e5900. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Williams, G.M.; Li, Y.; Chen, H.; Li, R.S.; Forsyth, S.J.; Barnett, A.G.; Guo, J.; Feng, Z.; McManus, D.P. A cluster-randomized bovine intervention trial against Schistosoma japonicum in the People’s Republic of China: Design and baseline results. Am. J. Trop. Med. Hyg. 2007, 77, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Williams, G.M.; Li, Y.S.; Chen, H.G.; Forsyth, S.; Li, R.; Barnett, A.; Guo, J.G.; Ross, A.; Feng, Z.; et al. The role of bovines in human Schistosoma japonicum infection in the People’s Republic China. Am. J. Trop. Med. Hyg. 2009, 81, 1046. [Google Scholar]
- Carlton, E.J.; Bates, M.N.; Zhong, B.; Seto, E.Y.; Spear, R.C. Evaluation of mammalian and intermediate host surveillance methods for detecting schistosomiasis reemergence in southwest China. PLoS Negl. Trop. Dis. 2011, 5, e987. [Google Scholar] [CrossRef] [PubMed]
- Van Dorssen, C.F.; Gordon, C.A.; Li, Y.; Williams, G.M.; Wang, Y.; Luo, Z.; Gobert, G.N.; You, H.; McManus, D.P.; Gray, D.J. Rodents, goats and dogs—Their potential roles in the transmission of schistosomiasis in China. Parasitology 2017, 144, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Angeles, J.M.; Leonardo, L.R.; Goto, Y.; Kirinoki, M.; Villacorte, E.A.; Hakimi, H.; Moendeg, K.J.; Lee, S.; Rivera, P.T.; Inoue, N.; et al. Water buffalo as sentinel animals for schistosomiasis surveillance. Bull. World Health Organ. 2015, 93, 511–512. [Google Scholar] [CrossRef] [PubMed]
- Satrija, F.; Ridwan, Y.; Jastal, S.; Rauf, A. Current status of schistosomiasis in Indonesia. Acta Trop. 2015, 141, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.G.P.; Olveda, R.M.; Acosta, L.; Harn, D.A.; Chy, D.; Yuesheng, L.; Gray, D.J.; Gordon, C.A.; McManus, D.P.; Williams, G.M. Road to the elimination of schistosomiasis from Asia: The journey is far from over. Microbes Infect. 2013, 15, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, S.; Wu, W.; Zhang, G.; Lu, D.; Ørnbjerg, N.; Johansen, M.V. Treatment and reinfection of water buffaloes and cattle infected with Schistosoma japonicum in Yangtze river valley, Anhui province, China. J. Parasitol. 2006, 92, 1088–1091. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fu, Z.; Feng, X.; Shi, Y.; Yuan, C.; Liu, J.; Hong, Y.; Li, H.; Lu, K.; Lin, J. Comparison of worm development and host immune responses in natural hosts of Schistosoma japonicum, yellow cattle and water buffalo. BMC Vet. Res. 2012, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-S.; McManus, D.P.; Lin, D.-D.; Williams, G.M.; Harn, D.A.; Ross, A.G.; Feng, Z.; Gray, D.J. The Schistosoma japonicum self-cure phenomenon in water buffaloes: Potential impact on the control and elimination of schistosomiasis in China. Int. J. Parasitol. 2014, 44, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, Y.; Gray, D.J.; Hu, G.; Chen, H.; Davis, G.M.; Sleigh, A.C.; Feng, Z.; McManus, D.P.; Williams, G.M. A drug-based intervention study on the importance of buffaloes for human Schistosoma japonicum infection around Poyang Lake, People’s Republic of China. Am. J. Trop. Med. Hyg. 2006, 74, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-B.; Liang, S.; Chen, G.-X.; Rea, C.; He, Z.-G.; Zhang, Z.-J.; Wei, J.-G.; Zhao, G.-M.; Jiang, Q.-W. An integrated strategy for transmission control of Schistosoma japonicum in a marshland area of China: Findings from a five-year longitudinal survey and mathematical modeling. Am. J. Trop. Med. Hyg. 2011, 85, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Yang, C.; Zhong, B.; Qiu, D. Re-emerging schistosomiasis in hilly and mountainous areas of Sichuan, China. Bull. World Health Organ. 2006, 84, 139–144. [Google Scholar] [CrossRef] [PubMed]
- WHO. NTD, WASH, Animal Health, Nutrition and Education Are Joining Forces to Eliminate Schistosomiasis in Mekong Region. Available online: http://www.wpro.who.int/mvp/ntd/ntd_wash/en/ (accessed on 12 September 2018).
- Chen, S.B.; Ai, L.; Hu, W.; Xu, J.; Bergquist, R.; Qin, Z.Q.; Chen, J.H. New anti-Schistosoma approaches in the People’s Republic of China: Development of diagnostics, vaccines and other new techniques belonging to the ‘omics’ group. Adv. Parasitol. 2016, 92, 385–408. [Google Scholar] [PubMed]
- Gray, D.J.; Li, Y.S.; Williams, G.M.; Zhao, Z.Y.; Harn, D.A.; Li, S.M.; Ren, M.Y.; Feng, Z.; Guo, F.Y.; Guo, J.G.; et al. A multi-component integrated approach for the elimination of schistosomiasis in the People’s Republic of China: Design and baseline results of a 4-year cluster-randomised intervention trial. Int. J. Parasitol. 2014, 44, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Inobaya, M.T.; Olveda, R.M.; Chau, T.N.P.; Olveda, D.U.; Ross, A.G.P. Prevention and control of schistosomiasis: A current perspective. Res. Rep. Trop. Med. 2014, 2014, 65–75. [Google Scholar] [PubMed]
- McManus, D.P.; Loukas, A. Current status of vaccines for schistosomiasis. Clin. Microbiol. Rev. 2008, 21, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Tebeje, B.M.; Harvie, M.; You, H.; Loukas, A.; McManus, D.P. Schistosomiasis vaccines: Where do we stand? Parasit. Vectors 2016, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Da’dara, A.A.; Li, Y.S.; Xiong, T.; Zhou, J.; Williams, G.M.; McManus, D.P.; Feng, Z.; Yu, X.L.; Gray, D.J.; Harn, D.A. DNA-based vaccines protect against zoonotic schistosomiasis in water buffalo. Vaccine 2008, 26, 3617–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.; Sleigh, A.C.; Li, Y.; Feng, Z.; Davis, G.M.; Chen, H.; Ross, A.G.P.; Bergquist, R.; McManus, D.P. Mathematical modelling of schistosomiasis japonica: Comparison of control strategies in the People’s Republic of China. Acta Trop. 2002, 82, 253–262. [Google Scholar] [CrossRef]
- He, Y.; Guo, Y.; Ni, C.; Xia, F.; Liu, H.; Yu, Q. Compatibility between Onomelania hupensis and different isolates of Schistosoma japonicum in China. Southeast Asian J. Trop. Med. Public Health 1991, 22, 245–254. [Google Scholar] [PubMed]
- Wang, X.Q.; Mao, S.P. Comparison of morphology, pathogenicity and drug response among three isolates of Schistosoma japonicum in the mainland of China. Ann. Parasitol. Hum. Comp. 1989, 64, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Kirinoki, M.; Hu, M.; Yokoi, H.; Kawai, S.; Terrado, R.; Ilagan, E.; Chigusa, Y.; Sasaki, Y.; Matsuda, H. Comparative studies on susceptibilities of two different Japanese isolates of Oncomelania nosophora to three strains of Schistosoma japonicum originating from Japan, China, and the Philippines. Parasitology 2005, 130, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.P.; Jiang, M.S.; Littlewood, D.T.; Nie, P. Distinct genetic diversity of Oncomelania hupensis, intermediate host of Schistosoma japonicum in mainland China as revealed by ITS sequences. PLoS Negl. Trop. Dis. 2010, 4, e611. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Li, S.Z.; Abe, E.M.; Webster, B.L.; Rollinson, D.; Zhou, X.N. The genetic diversity and geographical separation study of Oncomelania hupensis populations in mainland China using microsatellite loci. Parasit. Vectors 2016, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Z.; Wang, Y.X.; Yang, K.; Liu, Q.; Wang, Q.; Zhang, Y.; Wu, X.H.; Guo, J.G.; Bergquist, R.; Zhou, X.N. Landscape genetics: The correlation of spatial and genetic distances of Oncomelania hupensis, the intermediate host snail of Schistosoma japonicum in mainland China. Geospat. Health 2009, 3, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Wang, W.; Zuo, Y.P.; Zhang, Z.Q.; Hong, Q.B.; Yang, G.J.; Zhu, H.R.; Liang, Y.S.; Yang, H.T. An integrated environmental improvement of marshlands: Impact on control and elimination of schistosomiasis in marshland regions along the Yangtze River, China. Infect. Dis. Poverty 2017, 6, 72. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Xue, J.B.; Zhang, X.; Hu, H.H.; Abe, E.M.; Rollinson, D.; Bergquist, R.; Zhou, Y.; Li, S.Z.; Zhou, X.N. Pattern analysis of schistosomiasis prevalence by exploring predictive modeling in Jiangling County, Hubei Province, P.R. China. Infect. Dis. Poverty 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- WHO. Expert Consultation to Accelerate Elimination of Asian Schistosomiasis, Shanghai, People’s Republic of China, 22–23 May 2017: Meeting Report; WHO Regional Office for the Western Pacific: Manila, Philippines, 2017; p. 18. [Google Scholar]
- Knopp, S.; Corstjens, P.L.; Koukounari, A.; Cercamondi, C.I.; Ame, S.M.; Ali, S.M.; de Dood, C.J.; Mohammed, K.A.; Utzinger, J.; Rollinson, D.; et al. Sensitivity and specificity of a urine circulating anodic antigen test for the diagnosis of Schistosoma haematobium in low endemic settings. PLoS Negl. Trop. Dis. 2015, 9, e0003752. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Carabin, H.; Balolong, D.; Tallo, V.L.; Olveda, R.; Yuan, M.; McGarvey, S.T. Prevalence of Schistosoma japonicum infection of Oncomelania quadrasi snail colonies in 50 irrigated and rain-fed villages of Samar Province, the Philippines. Acta Trop. 2008, 105, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Sumarni, S. The new endemic area of Schistosoma japonicum in Bada Highland Western Lore subdistrict, District of Poso, Central Sulawesi Province. Trop. Med. J. 2011, 1, 1–12. [Google Scholar]
- Vonghachack, Y.; Odermatt, P.; Taisayyavong, K.; Phounsavath, S.; Akkhavong, K.; Sayasone, S. Transmission of Opisthorchis viverrini, Schistosoma mekongi and soil-transmitted helminthes on the Mekong Islands, Southern Lao PDR. Infect. Dis. Poverty 2017, 6, 131. [Google Scholar] [CrossRef] [PubMed]
- Attwood, S.W. Schistosomiasis in the Mekong Region: Epidemiology and phylogeography. Adv. Parasitol. 2001, 50, 87–152. [Google Scholar] [PubMed]
- Attwood, S.W.; Upatham, E.S. Observations on Neotricula aperta (Gastropoda: Pomatiopsidae) population densities in Thailand and central Laos: Implications for the spread of Mekong schistosomiasis. Parasit. Vectors 2012, 5, 126. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Zhou, Y.B.; Chen, Y.; Liang, S.; Li, L.H.; Zheng, S.B.; Zhu, S.P.; Ren, G.H.; Song, X.X.; Jiang, Q.W. Three Gorges Dam: Impact of water level changes on the density of schistosome-transmitting snail Oncomelania hupensis in Dongting Lake area, China. PLoS Negl. Trop. Dis. 2015, 9, e0003882. [Google Scholar] [CrossRef] [PubMed]
- Pesigan, T.P.; Hairston, N.G.; Jauregui, J.J.; Garcia, E.G.; Santos, A.T.; Santos, B.C.; Besa, A.A. Studies on Schistosoma japonicum infection in the Philippines. 2. The molluscan host. Bull. World Health Organ. 1958, 18, 481–578. [Google Scholar] [PubMed]
- Garjito, T.A.; Sudomo, M.; Dahlan, M.; Nurwidayati, A. Schistosomiasis in Indonesia: Past and present. Parasitol. Int. 2008, 57, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Minai, M.; Hosaka, Y.; Ohta, N. Historical view of schistosomiasis japonica in Japan: Implementation and evaluation of disease-control strategies in Yamanashi Prefecture. Parasitol. Int. 2003, 52, 321–326. [Google Scholar] [CrossRef]
- Yang, G.-J.; Li, W.; Sun, L.-P.; Wu, F.; Yang, K.; Huang, Y.-X.; Zhou, X.-N. Molluscicidal efficacies of different formulations of niclosamide: Result of meta-analysis of Chinese literature. Parasit. Vectors 2010, 3, 84. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.R.; Liu, L.; Zhou, X.N.; Yang, G.J. Ecological model to predict potential habitats of Oncomelania hupensis, the intermediate host of Schistosoma japonicum in the mountainous regions, China. PLoS Negl. Trop. Dis. 2015, 9, e0004028. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhou, X.N.; Yang, K.; Wang, X.H.; Yao, Z.Q.; Wang, T.P.; Yang, G.J.; Yang, Y.J.; Zhang, S.Q.; Wang, J.; et al. Strategy formulation for schistosomiasis japonica control in different environmental settings supported by spatial analysis: A case study from China. Geospat. Health 2007, 1, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, Z.; Chen, Y.; Wang, Z.; Gao, J.; Tao, B.; Jiang, Q.; Jiang, Q. Spatial pattern of schistosomiasis in Xingzi, Jiangxi Province, China: The effects of environmental factors. Parasit. Vectors 2013, 6, 214. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, R.; Bergquist, R.; Lynn, H.; Gao, F.; Wang, Q.; Zhang, S.; Sun, L.; Zhang, Z.; Jiang, Q. Spatio-temporal transmission and environmental determinants of schistosomiasis japonica in Anhui Province, China. PLoS Negl. Trop. Dis. 2015, 9, e0003470. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Sleigh, A.C.; Ross, A.G.P.; Williams, G.; Tanner, M.; McManus, D.P. Epidemiology of Schistosoma japonicum in China: Morbidity and strategies for control in the Dongting Lake region. Int. J. Parasitol. 2000, 30, 273–281. [Google Scholar] [CrossRef]
- Yang, K.; Li, L.; Huang, Y.; Zhang, J.; Wu, F.; Hang, D.; Steinmann, P.; Liang, Y. Spatio-temporal analysis to identify determinants of Oncomelania hupensis infection with Schistosoma japonicum in Jiangsu province, China. Parasit. Vectors 2013, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.N.; Yang, G.J.; Yang, K.; Wang, X.H.; Hong, Q.B.; Sun, L.P.; Malone, J.B.; Kristensen, T.K.; Bergquist, N.R.; Utzinger, J. Potential impact of climate change on schistosomiasis transmission in China. Am. J. Trop. Med. Hyg. 2008, 78, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Raso, G.; Zhao, Z.Y.; He, Y.K.; Ellis, M.K.; McManus, D.P. Large water management projects and schistosomiasis control, Dongting Lake region, China. Emerg. Infect. Dis. 2007, 13, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Thrift, A.P.; Williams, G.M.; Zheng, F.; Li, Y.-S.; Guo, J.; Chen, H.; Wang, T.; Xu, X.J.; Zhu, R.; et al. Five-year longitudinal assessment of the downstream impact on schistosomiasis transmission following closure of the Three Gorges Dam. PLoS Negl. Trop. Dis. 2012, 6, e1588. [Google Scholar] [CrossRef]
- McManus, D.P.; Gray, D.J.; Li, Y.; Feng, Z.; Williams, G.M.; Stewart, D.; Rey-Ladino, J.; Ross, A.G. Schistosomiasis in the People’s Republic of China: The era of the Three Gorges Dam. Clin. Microbiol. Rev. 2010, 23, 442–466. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Fan, J.; Peterson, A.T. Schistosoma japonicum transmission risk maps at present and under climate change in mainland China. PLoS Negl. Trop. Dis. 2017, 11, e0006021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Bergquist, R.; Leonardo, L.R.; Olveda, R. Schistosomiasis: The Disease and Its Control. 2008. Available online: https://www.researchgate.net/publication/242285494_Schistosomiasis_The_Disease_and_its_Control (accessed on 7 January 2019).
- Nyein, N. WHO’s Field Visit Reports over 400 Schistosomiasis Cases in Rakhine State. Available online: http://www.moi.gov.mm/moi:eng/?q=news/14/11/2018/id-14720 (accessed on 7 January 2019).
- Raso, G.; Li, Y.S.; Zhao, Z.Y.; Balen, J.; Williams, G.M.; McManus, D.P. Spatial distribution of human Schistosoma japonicum infections in the Dongting Lake Region, China. PLoS ONE 2009, 4, e6947. [Google Scholar] [CrossRef] [PubMed]
- Ziegelbauer, K.; Speich, B.; Mäusezahl, D.; Bos, R.; Keiser, J.; Utzinger, J. Effect of sanitation on soil-transmitted helminth infection: Systematic review and meta-analysis. PLoS Med. 2012, 9, e1001162. [Google Scholar] [CrossRef] [PubMed]
- Strunz, E.C.; Addiss, D.G.; Stocks, M.E.; Ogden, S.; Utzinger, J.; Freeman, M.C. Water, sanitation, hygiene, and soil-transmitted helminth infection: A systematic review and meta-analysis. PLoS Med. 2014, 11, e1001620. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.J.; Savage, G.B.; Gray, D.J.; Atkinson, J.M.; Magalhães, R.J.S.; Nery, S.V.; McCarthy, J.S.; Velleman, Y.; Wicken, J.H.; Traub, R.J.; et al. Water, sanitation, and hygiene (WASH): A critical component for sustainable soil-transmitted helminth and schistosomiasis control. PLoS Negl. Trop. Dis. 2014, 8, e2651. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.J.; Biritwum, N.K.; Woods, G.; Velleman, Y.; Fleming, F.; Stothard, J.R. Tailoring water, sanitation, and hygiene (WASH) targets for soil-transmitted helminthiasis and schistosomiasis control. Trends Parasitol. 2018, 34, 53–63. [Google Scholar] [CrossRef] [PubMed]
- WHO. Sixty Fifth World Health Assembly: Elimination of Schistosimasis; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Secor, W.E. Water-based interventions for schistosomiasis control. Pathog. Glob. Health 2014, 108, 246–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimes, J.E.T.; Croll, D.; Harrison, W.E.; Utzinger, J.; Freeman, M.C.; Templeton, M.R. The roles of water, sanitation and hygiene in reducing schistosomiasis: A review. Parasit. Vectors 2015, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- WHO. Sanitation, Drinking-Water and Health: Achievements and Challenges Ahead. Available online: http://www.who.int/iris/handle/10665/260222 (accessed on 19 October 2018).
- USAID. Water, Sanitation and Hygiene in Indonesia. Available online: https://www.usaid.gov/actingonthecall/stories/indonesia-wash (accessed on 19 October 2018).
- UNICEF. Water, Sanitation and Hygiene: Philippines. Available online: https://www.unicef.org/philippines/wes_21334.html (accessed on 19 October 2018).
- WGF. Goal Wash Project in the Philippines. Available online: http://www.watergovernance.org/resources/goal-wash-project-philippines/ (accessed on 19 October 2018).
- UN. Sanitation, Drinking-Water and Hygiene Status Overview: Indonesia; UN-Water Global Analysis and Assessment of Sanitation and Drinking-Water: Geneva, Switzerland, 2015. [Google Scholar]
- Camon, C.B. Implementation of Water, Sanitation and Hygiene (WASH) in Public Schools: A Citizen Participatory Audit; Department of Education: Cagayan de Oro City, Philippines, 2017.
- Chen, L.; Zhong, B.; Xu, J.; Li, R.Z.; Cao, C.L. Health education as an important component in the national schistosomiasis control programme in the People’s Republic of China. Adv. Parasitol. 2016, 92, 307–339. [Google Scholar] [PubMed]
- Wang, L.; Chen, H.; Guo, J.; Zeng, X.; Hong, X.; Xiong, J.; Wu, X.; Wang, X.; Wang, L.; Xia, G.; et al. A strategy to control transmission of Schistosoma japonicum in China. N. Engl. J. Med. 2009, 360, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.G.; Cao, C.L.; Hu, G.H.; Lin, H.; Li, D.; Zhu, R.; Xu, J. The role of ‘passive chemotherapy’ plus health education for schistosomiasis control in China during maintenance and consolidation phase. Acta Trop. 2005, 96, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.H.; Hu, J.; Song, K.Y.; Lin, D.D.; Zhang, J.; Cao, C.L.; Xu, J.; Li, D.; Jiang, W.S. The role of health education and health promotion in the control of schistosomiasis: Experiences from a 12-year intervention study in the Poyang Lake area. Acta Trop. 2005, 96, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Kloos, H. Human behavior, health education and schistosomiasis control: A review. Soc. Sci. Med. 1995, 40, 1497–1511. [Google Scholar] [CrossRef]
- McManus, D.P.; Bieri, F.A.; Li, Y.S.; Williams, G.M.; Yuan, L.P.; Henglin, Y.; Du, Z.W.; Clements, A.C.A.; Steinmann, P.; Raso, G.; et al. Health education and the control of intestinal worm infections in China: A new vision. Parasit. Vectors 2014, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Y.; Deng, Y.; Steinmann, P.; Yang, K. The effects of health education on schistosomiasis japonica prevalence and relevant knowledge in the People’s Republic of China: A systematic review and meta-analysis. Parasitol. Int. 2013, 62, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Muth, S.; Sayasone, S.; Odermatt-Biays, S.; Phompida, S.; Duong, S.; Odermatt, P. Schistosoma mekongi in Cambodia and Lao People’s Democratic Republic. Adv. Parasitol. 2010, 72, 179–203. [Google Scholar] [PubMed]
- McManus, D.P.; Gray, D.J.; Ross, A.G.; Williams, G.M.; He, H.B.; Li, Y.S. Schistosomiasis research in the Dongting Lake region and its impact on local and national treatment and control in China. PLoS Negl. Trop. Dis. 2011, 5, e1053. [Google Scholar] [CrossRef] [PubMed]
- Molehin, A.J.; Rojo, J.U.; Siddiqui, S.Z.; Gray, S.A.; Carter, D.; Siddiqui, A.A. Development of a schistosomiasis vaccine. Expert Rev. Vac. 2016, 15, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Rim, H.-J.; Chai, J.-Y.; Min, D.-Y.; Cho, S.; Eom, K.S.; Hong, S.; Sohn, W.; Yong, T.; Deodato, G.; Standgaard, H.; et al. Prevalence of intestinal parasite infections on a national scale among primary schoolchildren in Laos. Parasitol. Res. 2003, 91, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, S.; Xia, C.; Chen, Y.; Lynn, H.; Zhang, T.; Xiong, C.; Chen, G.; He, Z.; Zhang, Z. Assessment of the national schistosomiasis control program in a typical region along the Yangtze River, China. Int. J. Parasitol. 2017, 47, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Qin, Y.; Chu, K.; Meng, R.; Liu, Y.; McGarvey, S.T.; Olveda, R.; Acosta, L.P.; Ji, M.; Fernandez, T.; et al. High prevalence of Schistosoma japonicum infection in water buffaloes in the Philippines assessed by real-time polymerase chain reaction. Am. J. Trop. Med. Hyg. 2010, 82, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Logan, O.T. A case of dysentery in Hunan province, caused by the trematode, Schistosoma japonicum. Chin. Med. J. 1905, 19, 243–245. [Google Scholar]
- Wooley, P.G. The occurrence of Schistosoma japonicum Vel Cattol in the Philippines. Philipp. J. Sci. 1906, 1, 83–90. [Google Scholar]
- Okabe, K. Progress of Medical Parasitology in Japan; Meguro Parasitology Museum: Tokyo, Japan, 1964; Volume 1. [Google Scholar]
- Lei, Z.L.; Zheng, H.; Zhang, L.J.; Zhu, R.; Xu, Z.M.; Xu, J.; Fu, Q.; Wang, Q.; Li, S.Z.; Zhou, X.N. Endemic status of schistosomiasis in People’s Republic of China in 2013. Chin. J. Schistosomiasis Control 2014, 26, 591–597. [Google Scholar]
- WHO. Elimination of Schistosomiasis from Low-Transmission Areas: Report of a WHO Informal Consultation; WHO: Geneva, Switzerland, 2008. [Google Scholar]
- Yuan, H.; Jiagang, G.; Bergquist, R.; Tanner, M.; Xianyi, C.; Huanzeng, W. The 1992–1999 World Bank Schistosomiasis research initiative in China: Outcome and perspectives. Parasitol. Int. 2000, 49, 195–207. [Google Scholar] [CrossRef]
- Collins, C.; Xu, J.; Tang, S. Schistosomiasis control and the health system in P.R. China. Infect. Dis. Poverty 2012, 1, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.P.; Wang, W.; Hong, Q.B.; Li, S.Z.; Liang, Y.S.; Yang, H.T.; Zhou, X.N. Approaches being used in the national schistosomiasis elimination programme in China: A review. Infect. Dis. Poverty 2017, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.D.; Guo, J.G.; Wu, X.H.; Chen, H.G.; Wang, T.P.; Zhu, S.P.; Zhang, Z.H.; Steinmann, P.; Yang, G.J.; Wang, S.P.; et al. China’s new strategy to block Schistosoma japonicum transmission: Experiences and impact beyond schistosomiasis. Trop. Med. Int. Health 2009, 14, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, L.R.; Rivera, P.; Saniel, O.; Villacorte, E.; Labanan, M.A.; Crisostomo, B.; Hernandez, L.; Baquilod, M.; Erce, E.; Martinez, R.; et al. A national baseline prevalence survey of schistosomiasis in the Philippines using stratified two-step systematic cluster sampling design. J. Trop. Med. 2012, 2012, 936128. [Google Scholar] [CrossRef] [PubMed]
- Acosta, L.P.; Aligui, G.D.L.; Tiu, W.U.; McManus, D.P.; Olveda, R.M. Immune correlate study on human Schistosoma japonicum in a well-defined population in Leyte, Philippines: I. Assessment of ‘resistance’ versus ‘susceptibility’ to S. japonicum infection. Acta Trop. 2002, 84, 127–136. [Google Scholar] [CrossRef]
- Tarafder, M.R.; Balolong, E., Jr.; Carabin, H.; Bélisle, P.; Tallo, V.; Joseph, L.; Alday, P.; Gonzales, R.O.; Riley, S.; Olveda, R.; et al. A cross-sectional study of the prevalence of intensity of infection with Schistosoma japonicum in 50 irrigated and rain-fed villages in Samar Province, the Philippines. BMC Public Health 2006, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Inobaya, M.T.; Olveda, R.M.; Tallo, V.; McManus, D.P.; Williams, G.M.; Harn, D.A.; Li, Y.; Chau, T.N.P.; Olveda, D.U.; Ross, A.G. Schistosomiasis mass drug administration in the Philippines: Lessons learnt and the global implications. Microbes Infect. 2015, 17, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Steinman, P.; Maybe, D.; Zhou, X.N.; Lv, S.; Li, S.Z.; Peeling, R. Evolution of the national schistosomiasis control programmes in the People’s Republic of China. Adv. Parasitol. 2016, 92, 1–38. [Google Scholar] [PubMed]
- Pesigan, T.P.; Farooq, M.; Hairston, N.G.; Jauregui, J.J.; Garcia, E.G.; Santos, A.T.; Santos, B.C.; Besa, A.A. Studies on Schistosoma japonicum infection in the Philippines: 3. Preliminary control experiments. Bull. World Health Organ. 1958, 19, 223–261. [Google Scholar] [PubMed]
- Pesigan, T.P.; Hairston, N.G. The effect of snail control on the prevalence of Schistosoma japonicum infection in the Philippines. Bull. World Health Organ. 1961, 25, 479–482. [Google Scholar] [PubMed]
- DOH. Schistosomiasis Control Program. Available online: http://www.doh.gov.ph/content/schistosomiasis-control-program.html (accessed on 1 September 2014).
- Leonardo, L.R.; Acosta, L.P.; Olveda, R.M.; Aligui, G.D.L. Difficulties and strategies in the control of schistosomiasis in the Philippines. Acta Trop. 2002, 82, 295–299. [Google Scholar] [CrossRef]
- Tallo, V.L.; Carabian, H.; Alday, P.P.; Balolong, E., Jr.; Olveda, R.M.; McGarvey, S.T. Is mass treatment the appropriate schistosomiasis elimination strategy? Bull. World Health Organ. 2008, 86, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Inobaya, M.T.; Chau, T.N.; Ng, S.-K.; MacDougall, C.; Olveda, R.M.; Tallo, V.L.; Landicho, J.M.; Malacad, C.M.; Aligato, M.F.; Guevarra, J.B.; et al. Mass drug administration and the sustainable control of schistosomiasis: An evaluation of treatment compliance in the rural Philippines. Parasit. Vectors 2018, 11, 441. [Google Scholar] [CrossRef] [PubMed]
- Inobaya, M.T.; Chau, T.N.; Ng, S.K.; MacDougall, C.; Olveda, R.M.; Tallo, V.L.; Landicho, J.M.; Malacad, C.M.; Aligato, M.F.; Guevarra, J.R.; et al. Mass drug administration the sustainable control of schistosomiasis: Community health workers are vital for global elimination efforts. Int. J. Infect. Dis. 2018, 66, 14–21. [Google Scholar] [CrossRef] [PubMed]
- WHO. Neglected Tropical Diseases in Indonesia: An Integrated Plan of Action 2011–2015; Ministry of Health Indonesia: Jakarta, Indonesia, 2010.
- Nasution, A. Government Decentralization Program in Indonesia. Available online: https://www.adb.org/publications/government-decentralization-program-indonesia (accessed on 17 September 2018).
- WHO. Schistosomiasis: An Emerging Public Health Problem; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- MoH. Freshwater Snail Parasite that Infects about Schistosomiasis Cases Attended the Professional Union. Available online: http://mohs.gov.mm/ (accessed on 12 December 2018).
- Han, K.; Wai, K.; Htun, M.W.; Aye, K.; Latt, A.; Chi Aung San, N.; Phyo, A.; Xu, J.; Li, S.-Z.; Zhou, X.-N.; et al. Molecular Verification of Schistosoma mekongi Infection in ShweKyin Township; Myanmar, 2017. In Proceedings of the 16th Annual Workshop of Regional Network on Asian Schistosomiasis and other Zoonotic Helminths (RNAS+), Yangon, Myanmar, 26–28 October 2016. [Google Scholar]
- Wai, K.T.; Han, K.T.; Oo, T. Intensifying responsiveness towards neglected intestinal helminth infections in a resource-constrained setting. Trop. Med. Health 2017, 45, 12. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S. Development in Times of Transition: The Socio-Econommic Status of Rakhine State; Livelihoods and Food Security Trust (LIFT) Fund: Rakhine, Myanmar, 2018. [Google Scholar]
- Harinasuta, C. Introduction: Schistosomiasis in Asia. Malacol. Rev. 1980, 2, 431–438. [Google Scholar]
- Urbani, C.; Sinoun, M.; Socheat, D.; Pholsena, K.; Strandgaard, H.; Odermatt, P.; Hatz, C. Epidemiology and control of mekongi schistosomiasis. Acta Trop. 2002, 82, 157–168. [Google Scholar] [CrossRef]
- WHO. Communities Drive Elimination of Schistosomiasis through Improving Water, Sanitation and Hygiene in Cambodia and the Lao People’s Democratic Republic. Available online: http://www.wpro.who.int/mvp/ntd/cl_swash/en/ (accessed on 12 September 2018).
- WHO. Seventeenth Meeting of the Regional Programme Review Group on Neglected Tropical Diseases in the Western Pacific; WHO: Manila, Philippines, 2017. [Google Scholar]
- IAMAT. Thailand General Health Risks: Schistosomiasis. Available online: https://www.iamat.org/country/thailand/risk/schistosomiasis (accessed on 29 January 2019).
- Sornmani, S.; Vivatanasesth, P.; Thirachantra, S. Clinical study of Mekong schistosomiasis at Khong Island, Southern Laos. Malacol. Rev. 1980, 7, 270–281. [Google Scholar]
- Campa, P.; Develoux, M.; Belkadi, G.; Magne, D.; Lame, C.; Carayon, M.-J.; Girard, P.-M. Chronic Schistosoma mekongi in a traveler—A case report and review of the literature. J. Travel Med. 2014, 21, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Clerinx, J.; Cnops, L.; Huyse, T.; Tannich, E.; Van Esbroeck, M. Diagnostic issues of acute schistosomiasis with Schistosoma mekongi in a traveler: A case report. J. Travel Med. 2013, 20, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Chuah, C.; Gobert, G.N.; Latif, B.; Heo, C.C.; Leow, C.Y. Schistosomiasis in Malaysia: A review. Acta Trop. 2019, 190, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Murugasu, R.; Wang, F.; Dissanaike, A.S. Schistosoma japonicum-type infection in Malaysia—Report of the first living case. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 389–391. [Google Scholar] [CrossRef]
- Latif, B.; Heo, C.C.; Razuin, R.; Shamalaa, D.V.; Tappe, D. Autochthonous human schistosomiasis, Malaysia. Emerg. Infect. Dis. 2013, 19, 1340–1341. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; McManus, D.P.; Jones, M.K.; Gray, D.J.; Gobert, G.N. The increase of exotic zoonotic helminth infections: The impact of urbanization, climate change and globalization. Adv. Parasitol. 2016, 91, 311–397. [Google Scholar] [PubMed]
- McCreesh, N.; Nikulin, G.; Booth, M. Predicting the effects of climate change on Schistosoma mansoni transmission in eastern Africa. Parasit. Vectors 2015, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Stensgaard, A.S.; Vounatsou, P.; Sengupta, M.E.; Utzinger, J. Schistosomes, snails and climate change: Current trends and future expectations. Acta Trop. 2019, 190, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Boissier, J.; Moné, H.; Mitta, G.; Bargues, M.D.; Molyneux, D.; Mas-Coma, S. Schistosomiasis reaches Europe. Lancet Infect. Dis. 2015, 15, 757–758. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Distribution | Animal Definitive Hosts | Intermediate Hosts | Eggs | |
|---|---|---|---|---|
| S. japonicum | Indonesia, the Philippines, P. R. China | 46 known mammalian hosts including water buffalo and cattle, dogs, pigs, and rodents | Oncomelania spp. | 70–100 × 55–64 µm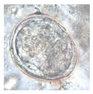 |
| S. mekongi | Cambodia, Lao PDR, Thailand | Dogs and pigs | Neotricula spp. | 50–80 × 40–65 µm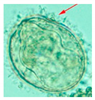 |
| S. malayensis | Malaysia | Rodents | Robertsiella spp. | 53–90 × 33–62 µm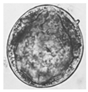 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, C.A.; Kurscheid, J.; Williams, G.M.; Clements, A.C.A.; Li, Y.; Zhou, X.-N.; Utzinger, J.; McManus, D.P.; Gray, D.J. Asian Schistosomiasis: Current Status and Prospects for Control Leading to Elimination. Trop. Med. Infect. Dis. 2019, 4, 40. https://doi.org/10.3390/tropicalmed4010040
Gordon CA, Kurscheid J, Williams GM, Clements ACA, Li Y, Zhou X-N, Utzinger J, McManus DP, Gray DJ. Asian Schistosomiasis: Current Status and Prospects for Control Leading to Elimination. Tropical Medicine and Infectious Disease. 2019; 4(1):40. https://doi.org/10.3390/tropicalmed4010040
Chicago/Turabian StyleGordon, Catherine A., Johanna Kurscheid, Gail M. Williams, Archie C. A. Clements, Yuesheng Li, Xiao-Nong Zhou, Jürg Utzinger, Donald P. McManus, and Darren J. Gray. 2019. "Asian Schistosomiasis: Current Status and Prospects for Control Leading to Elimination" Tropical Medicine and Infectious Disease 4, no. 1: 40. https://doi.org/10.3390/tropicalmed4010040