

Molecular Characterization of Staphylococcus aureus Complex Isolated from Free-Ranging Long-Tailed Macaques at Kosumpee Forest Park, Maha Sarakham, Thailand

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
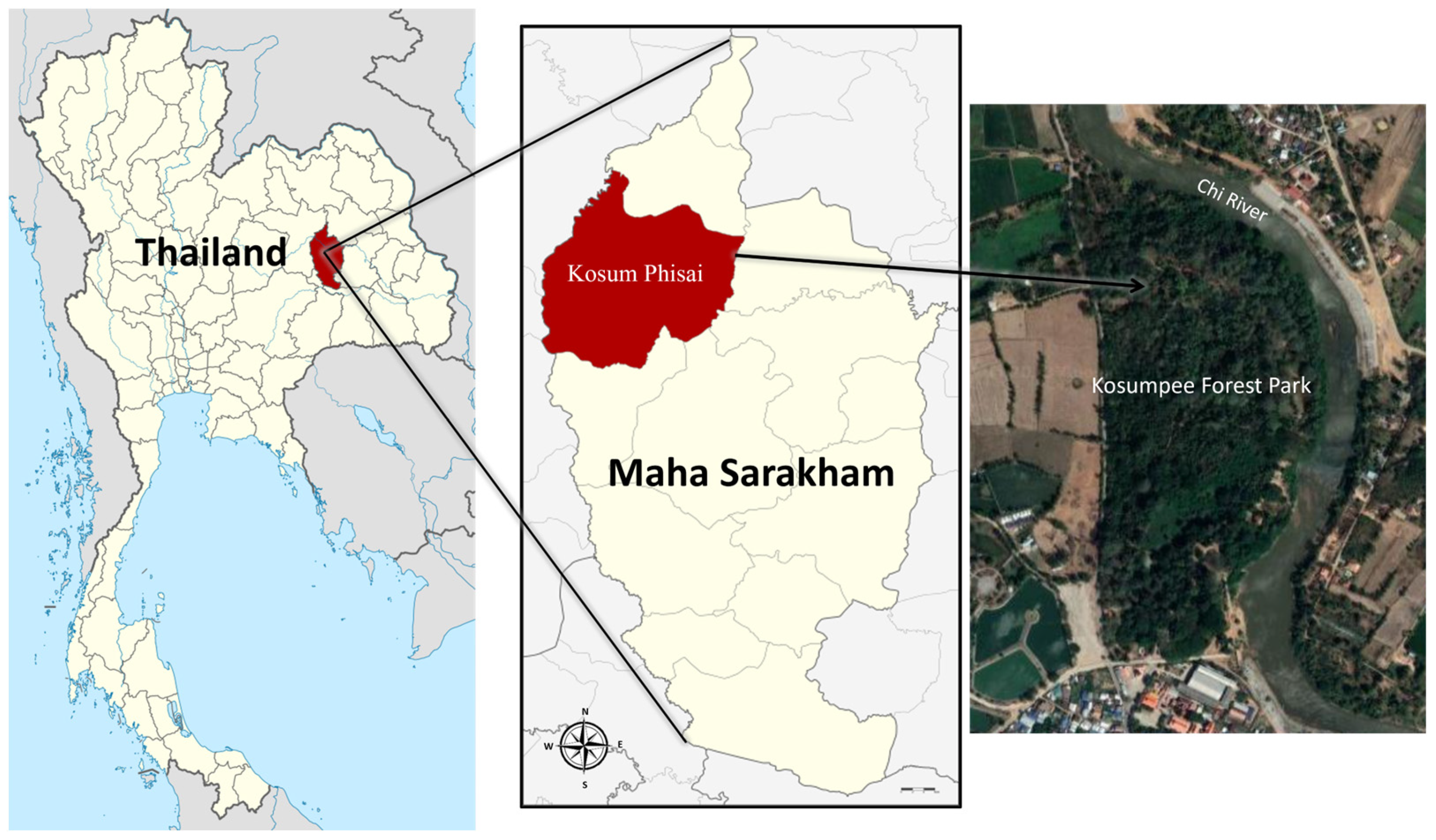
2.1. Sample Collection and Preparation
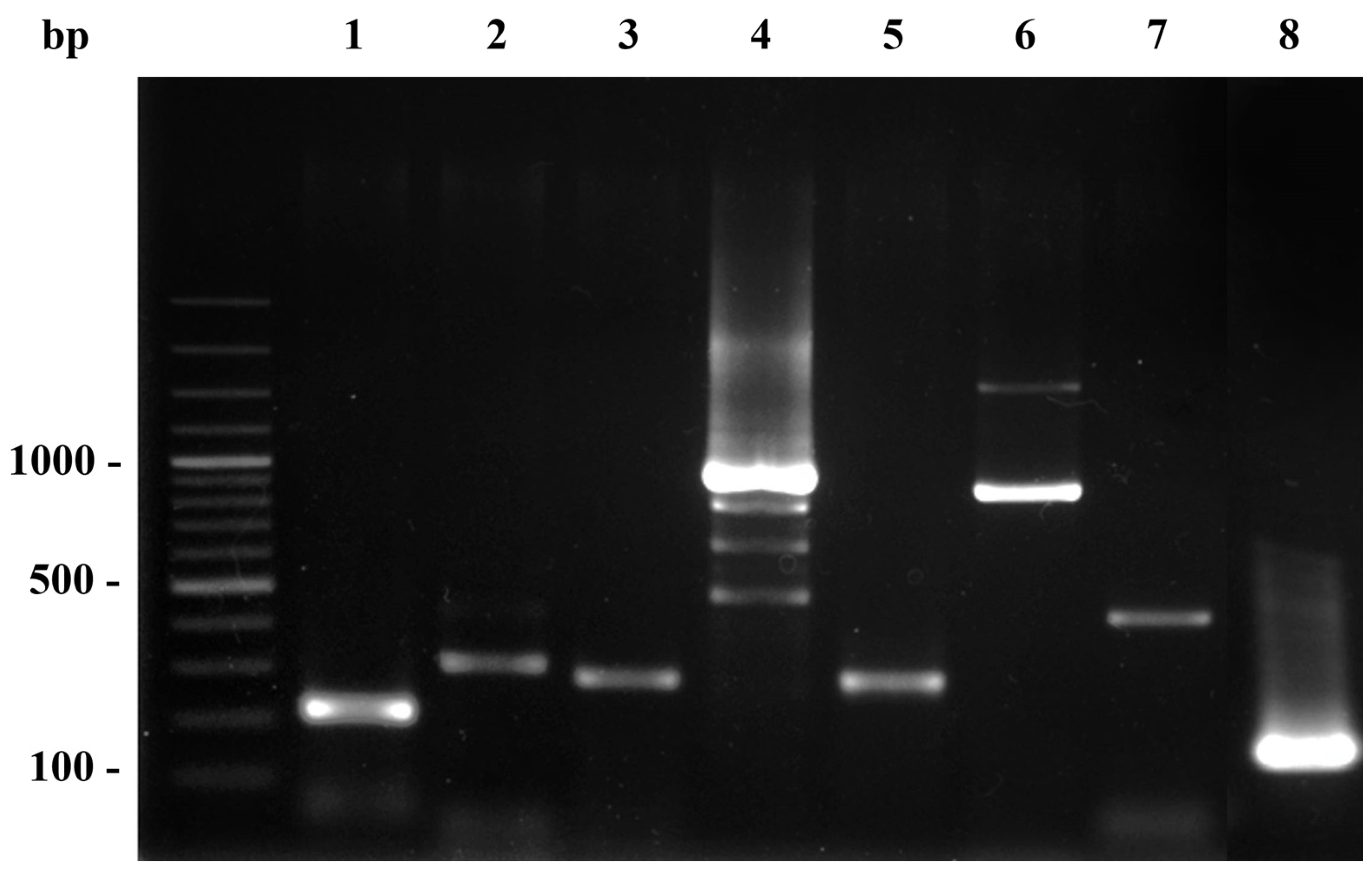
2.2. Detection of Staphylococcal Virulence Genes
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howden, B.P.; Giulieri, S.G.; Wong Fok Lung, T.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus host interactions and adaptation. Nat. Rev. Microbiol. 2023, 21, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Pumipuntu, N.; Chamnandee, T.; Saengthong, K.; Pathomthanasarn, S.; Tanee, T.; Kyes, P.; Thamsenanupap, P.; Karaket, A.; Roberts, M.C.; Kyes, R.C. Investigation of methicillin-resistant Staphylococcus aureus, methicillin-susceptible Staphylococcus aureus, and Staphylococcus argenteus from wild long-tailed macaques (Macaca fascicularis) at Kosumpee Forest Park, Maha Sarakham, Thailand. Vet. World. 2022, 15, 2693–2698. [Google Scholar] [CrossRef] [PubMed]
- Cihalova, K.; Chudobova, D.; Michalek, P.; Moulick, A.; Guran, R.; Kopel, P.; Adam, V.; Kizek, R. Staphylococcus aureus and MRSA Growth and Biofilm Formation after Treatment with Antibiotics and SeNPs. Int. J. Mol. Sci. 2015, 16, 24656–24672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus epidemiology in wildlife: A systematic review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [Green Version]
- Wielders, C.L.; Fluit, A.C.; Brisse, S.; Verhoef, J.; Schmitz, F.J. mecA gene is widely disseminated in Staphylococcus aureus population. J. Clin. Microbiol. 2002, 40, 3970–3975. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.K.; Ramaswamy, S.V.; Conradt, J.; Stemper, M.E.; Reich, R.; Reed, K.D.; Graviss, E.A. Novel polymorphisms in mec genes and a new mec complex type in methicillin-resistant Staphylococcus aureus isolates obtained in rural Wisconsin. Antimicrob. Agents Chemother. 2004, 48, 3080–3085. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Amin, K.B.; Rahman, S.M.M.; Khair, A.; Rahman, M.; Hossain, A.; Rahman, A.K.M.A.; Parvez, M.S.; Miura, N.; Alam, M.M. Investigation of methicillin-resistant Staphylococcus aureus among clinical isolates from humans and animals by culture methods and multiplex PCR. BMC Vet. Res. 2018, 14, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Schaumburg, F.; Alabi, A.S.; Köck, R.; Mellmann, A.; Kremsner, P.G.; Boesch, C.; Becker, K.; Leendertz, F.H.; Peters, G. Highly divergent Staphylococcus aureus isolates from African non-human primates. Environ. Microbiol. Rep. 2012, 4, 141–146. [Google Scholar] [CrossRef]
- Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Müller, E.; Gawlik, D.; Diezel, C.; Braun, S.D.; Paudel, S.; Acharya, M.; et al. Staphylococcus aureus and methicillin resistant S. aureus in Nepalese primates: Resistance to antimicrobials, virulence, and genetic lineages. Antibiotics 2020, 9, 689. [Google Scholar] [CrossRef]
- Nagel, M.; Dischinger, J.; Türck, M.; Verrier, D.; Oedenkoven, M.; Ngoubangoye, B.; Le Flohic, G.; Drexler, J.F.; Bierbaum, G.; Gonzalez, J.P. Human-associated Staphylococcus aureus strains within great ape populations in Central Africa (Gabon). Clin. Microbiol. Infect. 2013, 19, 1072–1077. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Müller, E.; Gawlik, D.; Paudel, S.; Acharya, M.; Bhattarai, S.; Pokharel, S.; et al. MRSA strains in Nepalese rhesus macaques (Macaca mulatta) and their environment. Front. Microbiol. 2019, 10, 2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaumburg, F.; Alabi, A.S.; Peters, G.; Becker, K. New epidemiology of Staphylococcus aureus infection in Africa. Clin. Microbiol. Infect. 2014, 20, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Indrawattana, N.; Sungkhachat, O.; Sookrung, N.; Chongsa-nguan, M.; Tungtrongchitr, A.; Voravuthikunchai, S.; Kong-ngoen, T.; Kurazono, H.; Chaicumpa, W. Staphylococcus aureus clinical isolates: Antibiotic susceptibility, molecular characteristics, and ability to form biofilm. Biomed. Res. Int. 2013, 2013, 314654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumipuntu, N.; Tunyong, W.; Chantratita, N.; Diraphat, P.; Pumirat, P.; Sookrung, N.; Chaicumpa, W.; Indrawattana, N. Staphylococcus spp. associated with subclinical bovine mastitis in central and northeast provinces of Thailand. PeerJ 2019, 7, e6587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Tan, X.; Zhang, X.; Xia, X.; Sun, H. The diversities of staphylococcal species, virulence, and antibiotic resistance genes in the subclinical mastitis milk from a single Chinese cow herd. Microb. Pathog. 2015, 88, 29–38. [Google Scholar] [CrossRef]
- Cheng, A.G.; McAdow, M.; Kim, H.K.; Bae, T.; Missiakas, D.M.; Schneewind, O. Contribution of coagulases towards Staphylococcus aureus disease and protective immunity. PLoS Pathog. 2010, 6, 1001036. [Google Scholar] [CrossRef] [Green Version]
- Koreen, L.S.; Ramaswamy, V.; Graviss, E.A.; Naidich, S.; Musser, J.M.; Kreiswirth, B.N. spa typing method for discriminating among Staphylococcus aureus isolates: Implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 2004, 42, 792–799. [Google Scholar] [CrossRef] [Green Version]
- Viau, M.; Longo, N.S.; Lipsky, P.E.; Zouali, M. Staphylococcal protein deletes B-1a and marginal zone B lymphocytes expressing human immunoglobulins: An immune evasion mechanism. J. Immunol. 2005, 175, 7719–7727. [Google Scholar] [CrossRef] [PubMed]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Bubeck Wardenburg, J.A. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, A.; Alber, J.; Limmler, C.; Bonner, B.; Huhn, A.; Kaleta, E.F.; Zschech, M. PCR-based detection of genes encoding virulence determinants in Staphylococcus aureus from birds. J. Vet. Med. B. Infect. Dis. Vet. Public Health 2005, 52, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Lina, G.; Piemont, Y.; Godail, G.F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of panton-valentine leucocidin producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyes, R.C.; Tanee, T.; Thamsenanupap, P.; Karaket, A.; Iskandar, E.; Kyes, P. Population status of the long-tailed macaques (Macaca fascicularis) at Kosumpee Forest Park, Maha Sarakham, Thailand. Am. J. Primatol. 2018, 80, 22. [Google Scholar]
- Schurer, J.M.; Ramirez, V.; Kyes, P.; Tanee, T.; Patarapadungkit, N.; Thamsenanupap, P.; Trufan, S.; Grant, E.T.; Garland-Lewis, G.; Kelley, S.; et al. Long-tailed macaques (Macaca fascicularis) in urban landscapes: Investigating gastrointestinal parasitism and barriers for healthy co-existence in northeast Thailand. Am. J. Trop. Med. Hyg. 2019, 100, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumipuntu, N.; Tanee, T.; Kyes, P.; Thamsenanupap, P.; Karaket, A.; Kyes, R.C. Leptospira Seroprevalence in free-ranging long-tailed macaques (Macaca fascicularis) at Kosumpee Forest Park, Maha Sarakham, Thailand. Infect. Dis. Rep. 2023, 15, 16–23. [Google Scholar] [CrossRef]
- Wu, D.; Li, X.; Yang, Y.; Zheng, Y.; Wang, C.; Deng, L.; Liu, L.; Li, C.; Shang, Y.; Zhao, C.; et al. Superantigen gene profiles and presence of exfoliative toxin genes in community-acquired meticillin-resistant Staphylococcus aureus isolated from Chinese children. J. Med. Microbiol. 2011, 60, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Aslantas, O.; Demir, C.; Turutoglu, H.; Cantekin, Z.; Ergun, Y.; Dogruer, G. Coagulase gene polymorphism of Staphylococcus aureus isolated form subclinical mastitis. Turk. J. Vet. Anim. Sci. 2007, 31, 253–257. [Google Scholar]
- Tristan, A.; Ying, L.; Bes, M.; Etienne, J.; Vandenesch, F.; Lina, G. Use of multiplex PCR to identify Staphylococcus aureus adhesins involved in human hematogenous infections. J. Clin. Microbiol. 2003, 41, 4465–4467. [Google Scholar] [CrossRef] [Green Version]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frénay, H.M.; Bunschoten, A.E.; Schouls, L.M.; Van Leeuwen, W.J.; Vandenbrouke-Grauls, C.M.; Verhoef, J.; Mooi, F.R. Molecular typing of methicillin-resistant Staphylococcus aureus on the basis of protein a gene polymorphism. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 60–64. [Google Scholar] [CrossRef]
- Seki, K.; Sakurada, J.; Seong, H.K.; Murai, M.; Tachi, H.; Ishii, H.; Masuda, S. Occurrence of coagulase serotype among Staphylococcus aureus strains isolated from healthy individuals-special reference to correlation with size of protein-A gene. Microbiol. Immunol. 1998, 42, 407–409. [Google Scholar] [PubMed]
- Indrawattana, N.; Pumipuntu, N.; Suriyakhun, N.; Jangsangthong, A.; Kulpeanprasit, S.; Chantratita, N.; Sookrung, N.; Chaicumpa, W.; Buranasinsup, S. Staphylococcus argenteus from rabbits in Thailand. MicrobiologyOpen 2019, 8, e00665. [Google Scholar] [CrossRef] [Green Version]
- Rasmi, A.H.; Ahmed, E.F.; Darwish, A.M.A.; Gad, G.F.M. Virulence genes distributed among Staphylococcus aureus causing wound infections and their correlation to antibiotic resistance. BMC Infect. Dis. 2022, 22, 652. [Google Scholar] [CrossRef]
- Ballah, F.M.; Islam, M.S.; Rana, M.L.; Ullah, M.A.; Ferdous, F.B.; Neloy, F.H.; Ievy, S.; Sobur, M.A.; Rahman, A.T.; Khatun, M.M.; et al. Virulence determinants and methicillin resistance in biofilm-forming Staphylococcus aureus from various food sources in Bangladesh. Antibiotics 2022, 11, 1666. [Google Scholar] [CrossRef]
- Akineden, Ö.; Annemüller, C.; Hassan, A.A.; Lämmler, C.; Wolter, W.; Zschöck, M. Toxin genes and other characteristics of Staphylococcus aureus isolates from milk of cows with mastitis. Clin. Diagn. Lab. Immunol. 2001, 8, 959–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullahi, I.N.; Fernández-Fernández, R.; Juárez-Fernández, G.; Martínez-Álvarez, S.; Eguizábal, P.; Zarazaga, M.; Lozano, C.; Torres, C. Wild Animals Are Reservoirs and Sentinels of Staphylococcus aureus and MRSA Clones: A Problem with “One Health” Concern. Antibiotics 2021, 10, 1556. [Google Scholar] [CrossRef]
- Zheng, Y.; Qin, C.; Zhang, X.; Zhu, Y.; Li, A.; Wang, M.; Tang, Y.; Kreiswirth, B.N.; Chen, L.; Zhang, H.; et al. The tst gene associated Staphylococcus aureus pathogenicity island facilitates its pathogenesis by promoting the secretion of inflammatory cytokines and inducing immune suppression. Microb. Pathog. 2019, 138, 103797. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Pauly, M.; Anoh, E.; Mossoun, A.; Wiersma, L.; Schubert, G.; Flammen, A.; Alabi, A.S.; Muyembe-Tamfum, J.J.; Grobusch, M.P.; et al. Staphylococcus aureus complex from animals and humans in three remote African regions. Clin. Microbiol. Infect. 2015, 21, 345.e1–345.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, S.; Roberts, S.; O’Shea, K.; Maloney, D.; Lorenzo, M.; Benson, C.E. Panton, valentine leukocidin (PVL) toxin positive MRSA strains isolated from companion animals. Vet. Microbiol. 2005, 108, 145–148. [Google Scholar] [CrossRef]
- Prashanth, K.; Rao, K.R.; Reddy, P.V.; Saranathan, R.; Makki, A.R. Genotypic characterization of Staphylococcus aureus obtained from humans and bovine mastitis samples in India. J. Glob. Infect. Dis. 2011, 3, 115–122. [Google Scholar] [CrossRef]
- Falugi, F.; Kim, H.K.; Missiakas, D.M.; Schneewind, O. Role of protein A in the evasion of host adaptive immune responses by Staphylococcus aureus. mBio 2013, 4, e00575-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman-Bausier, P.; Labate, C.; Towell, A.M.; Derclaye, S.; Geoghegan, J.A.; Dufrêne, Y.F. Staphylococcus aureus clumping factor A is a force-sensitive molecular switch that activates bacterial adhesion. Proc. Natl. Acad. Sci. USA 2018, 115, 5564–5569. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Target Gene | Sequence (5′–3′) | Amplicon Size (bp) | Ref. |
|---|---|---|---|
| tst | F: TTCACTATTTGTAAAAGTGTCAGACCCACT R: TACTAATGAATTTTTTTATCGTAAGCCCTT | 180 | [26] |
| coa | F: CGAGACCAAGATTCAACAAG R: AAAGAAAACCACTCACATCA | 410, 740, 910, 970 | [27] |
| clfA | F: ATTGGCGTGGCTTCAGTGCT R: CGTTTCTTCCGTAGTTGCATTTG | 292 | [28] |
| hla | F: CTGATTACTATCCAAGAAATTCGATTG R: CTTTCCAGCCTACTTTTTTATCAGT | 209 | [29] |
| hlb | F: GTGCACTTACTGACAATAGTGC R: GTTGATGAGTAGCTACCTTCAGT | 309 | [29] |
| spa (x-region) | F: CAAGCACCA AAAGAGGAA R: CACCAGGTTTAACGACAT | 320 | [30] |
| spa (IgG-biding region) | F: CACCTGCTGCAAATGCTGCG R: GGCTTGTTGTTGTCTTCCTC | 920 | [31] |
| pvl | F: ATCATTAGGTAAAATGTCTGGACATGATCCA R: GCATCAASTGTATTGGATAGCAAAAGC | 433 | [32] |
| Macaque ID | Isolate No. | Bacterial Identification | Virulence Genes Detection |
|---|---|---|---|
| M 1 | 1.1 | MRSA | clfA, coa, spa (Ig), spa(X), hlb, tst, pvl |
| 1.2 | MSSA | clfA, coa, spa (Ig), spa(X), hla, hlb, pvl | |
| M 2 | 2.2 | MRSA | clfA, coa, spa (Ig), spa(X), hla, tst, pvl |
| M 4 | 4.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, tst |
| M 7 | 7.3 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst |
| M 9 | 9.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst |
| M 15 | 15.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst |
| M 16 | 16.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb |
| M 17 | 17.1 | S. argenteus | clfA, coa, spa (Ig), spa(X), hla, pvl |
| M 19 | 19.2 | MSSA | clfA, coa, spa (Ig), spa(X), hla, pvl |
| M 21 | 21.3 | MSSA | clfA, coa, spa (Ig), spa(X), |
| 21.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst, pvl | |
| M 22 | 22.1 | MSSA | clfA, coa, spa (Ig), spa(X), hla, tst |
| 22.4 | MRSA | clfA, coa, spa (Ig), hla | |
| M 24 | 24.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, pvl |
| M 25 | 25.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst, pvl |
| M 26 | 26.5 | MSSA | clfA, coa, spa (Ig), spa(X), hla, hlb, pvl |
| M 27 | 27.3 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst, pvl |
| M 28 | 28.1 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, tst, pvl |
| 28.4 | S. argenteus | clfA, coa, spa (Ig), spa(X), hlb, pvl | |
| M 29 | 29.5 | MRSA | clfA, coa, spa (Ig), spa(X), hla, hlb, pvl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pumipuntu, N.; Tanee, T.; Thamsenanupap, P.; Kyes, P.; Karaket, A.; Kyes, R.C. Molecular Characterization of Staphylococcus aureus Complex Isolated from Free-Ranging Long-Tailed Macaques at Kosumpee Forest Park, Maha Sarakham, Thailand. Trop. Med. Infect. Dis. 2023, 8, 374. https://doi.org/10.3390/tropicalmed8070374
Pumipuntu N, Tanee T, Thamsenanupap P, Kyes P, Karaket A, Kyes RC. Molecular Characterization of Staphylococcus aureus Complex Isolated from Free-Ranging Long-Tailed Macaques at Kosumpee Forest Park, Maha Sarakham, Thailand. Tropical Medicine and Infectious Disease. 2023; 8(7):374. https://doi.org/10.3390/tropicalmed8070374
Chicago/Turabian StylePumipuntu, Natapol, Tawatchai Tanee, Penkhae Thamsenanupap, Pensri Kyes, Apichat Karaket, and Randall C. Kyes. 2023. "Molecular Characterization of Staphylococcus aureus Complex Isolated from Free-Ranging Long-Tailed Macaques at Kosumpee Forest Park, Maha Sarakham, Thailand" Tropical Medicine and Infectious Disease 8, no. 7: 374. https://doi.org/10.3390/tropicalmed8070374