
A Human Microglial Cell Line Expresses γ-Aminobutyric Acid (GABA) Receptors and Responds to GABA and Muscimol by Increasing Production of IL-8

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Human Microglia Cell (HMC3) Culture
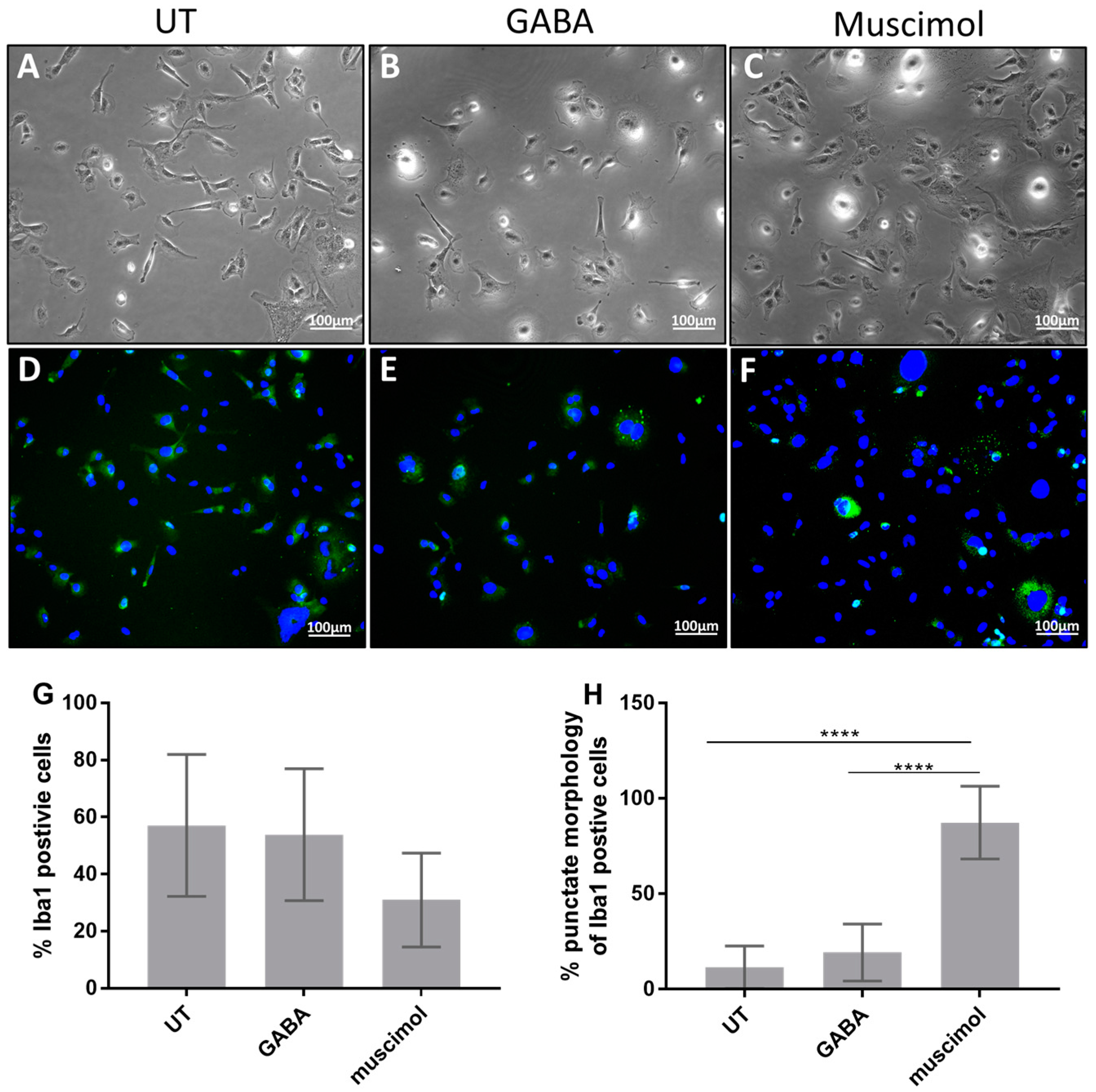
2.3. Fluorescent Microscopy and Iba1 Expression Analysis
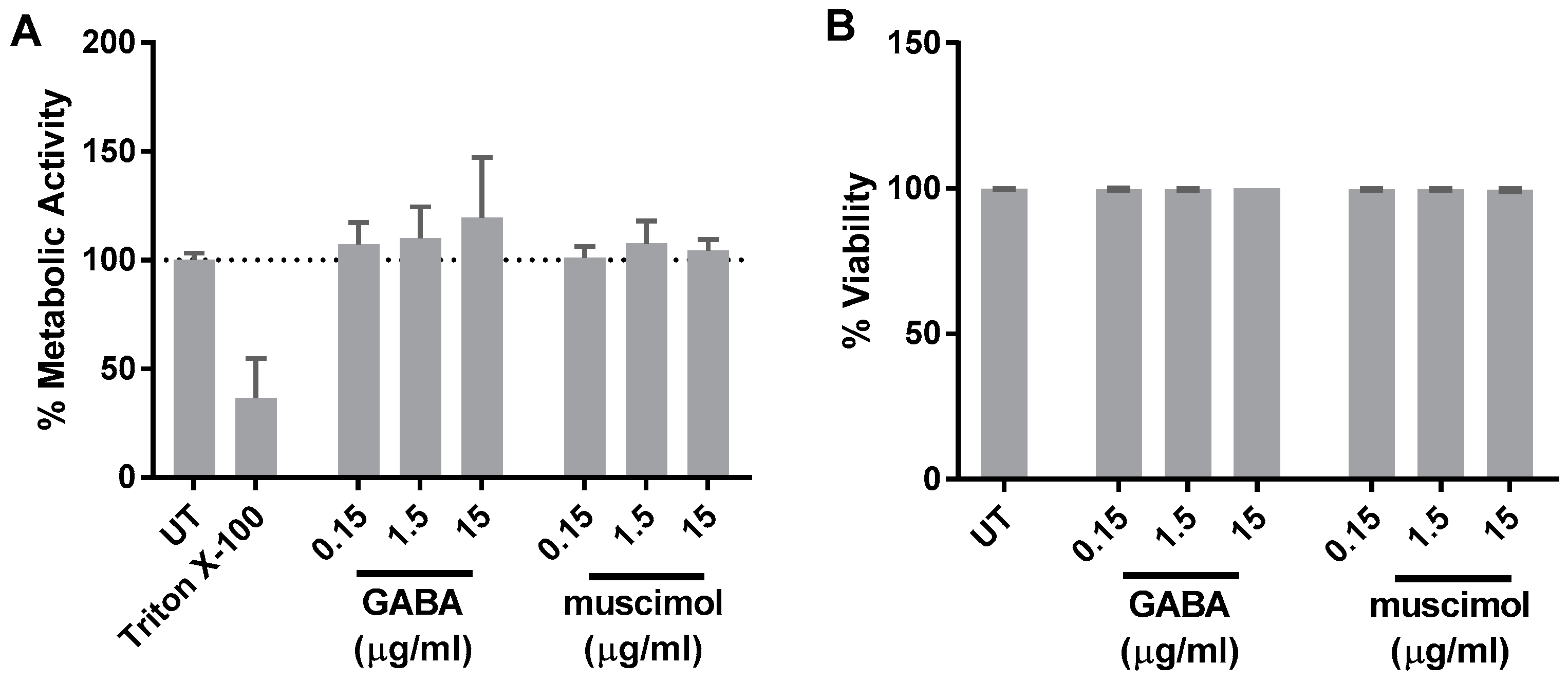
2.4. Metabolic Activity Analysis
Trypan Blue Exclusion Assay
2.5. Reactive Oxygen Species (ROS) Analysis
2.6. qPCR Array Analysis of Genes Associated with Neurotransmitter Receptors, Neurotransmitter Synthesis, and Exocytosis
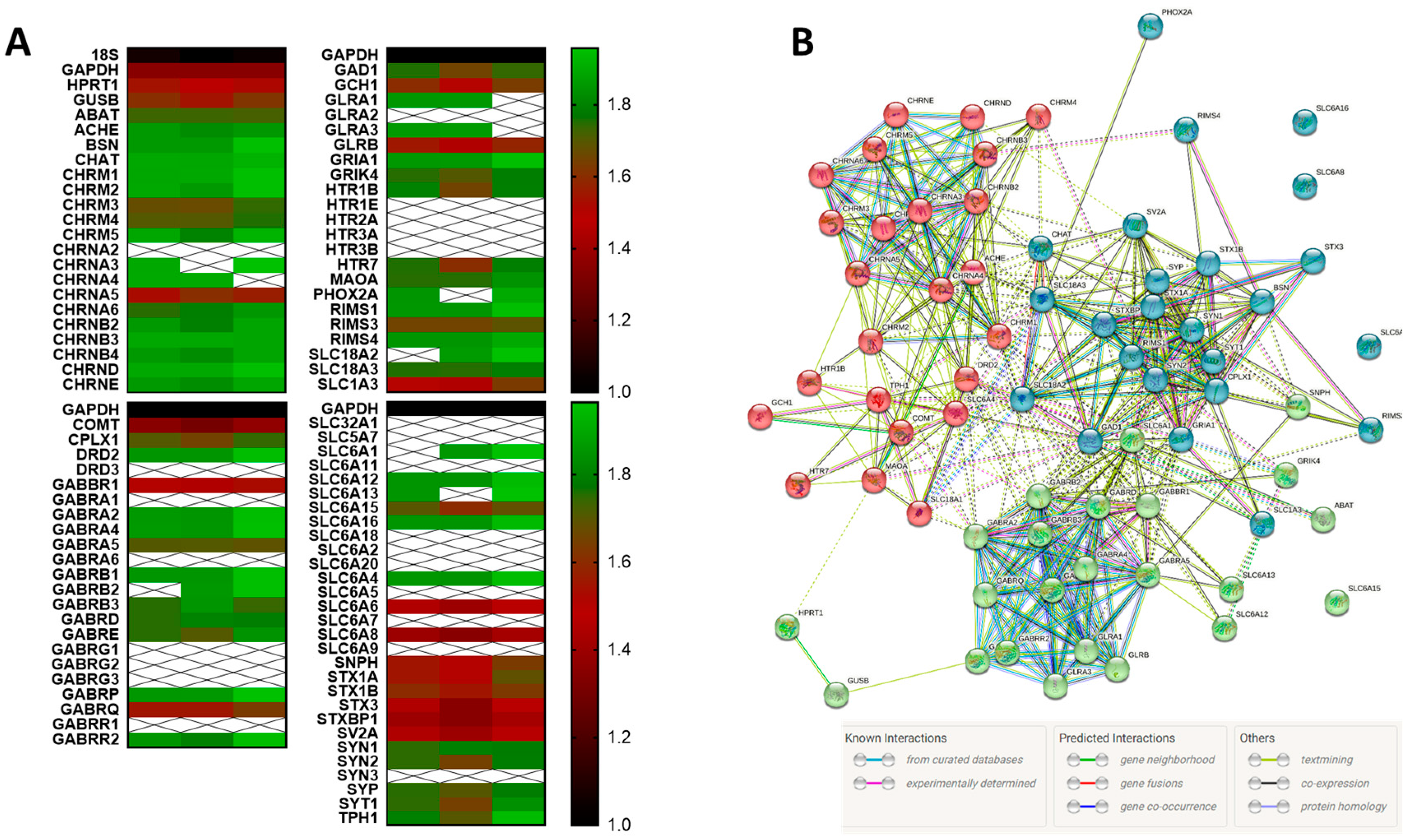
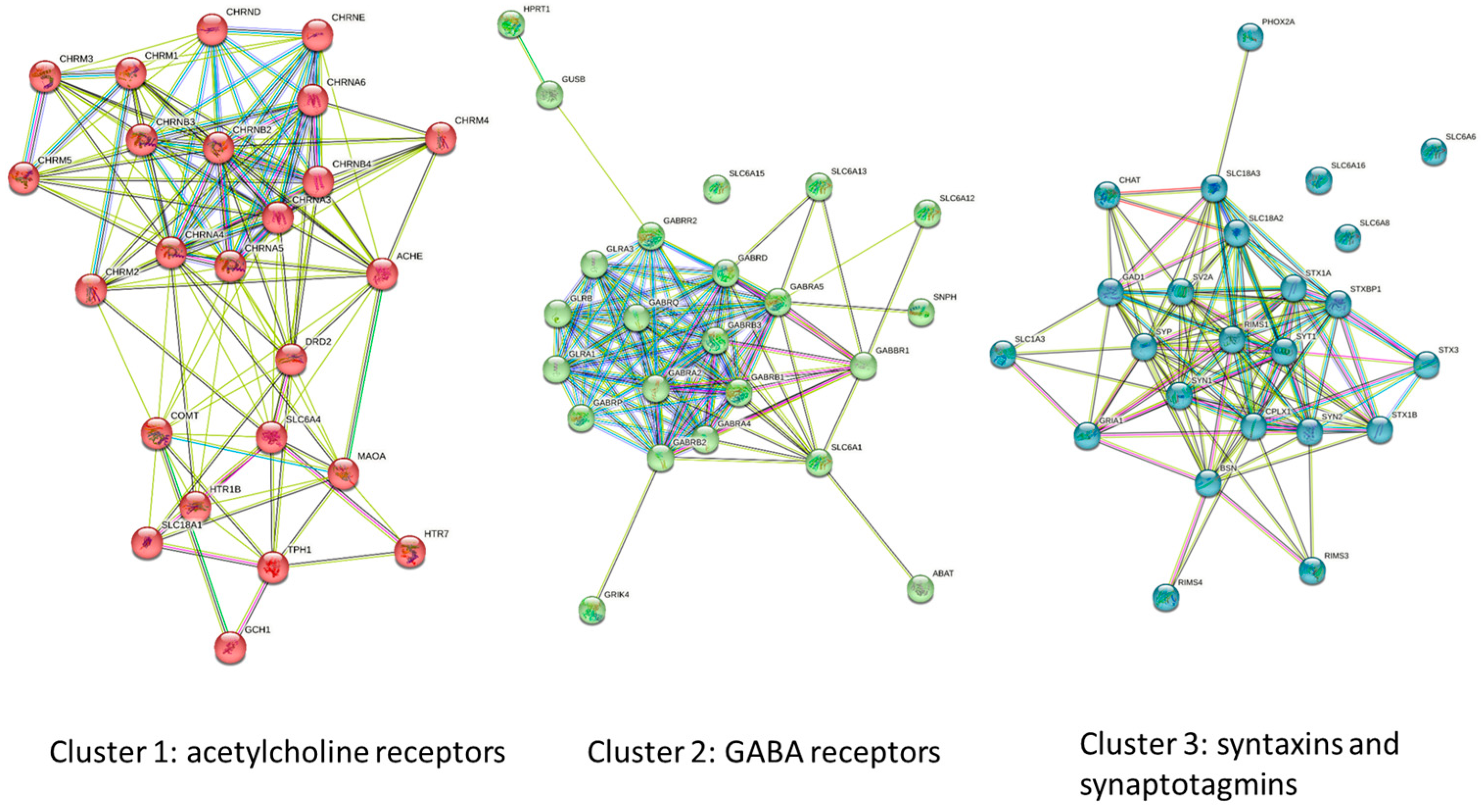
2.7. STRING Analysis and Protein Modeling
2.8. Metabolomic Profiling Using NMR
2.9. Chromatographic Conditions for HPLC-MS Analysis
2.10. Statistical Analysis
3. Results
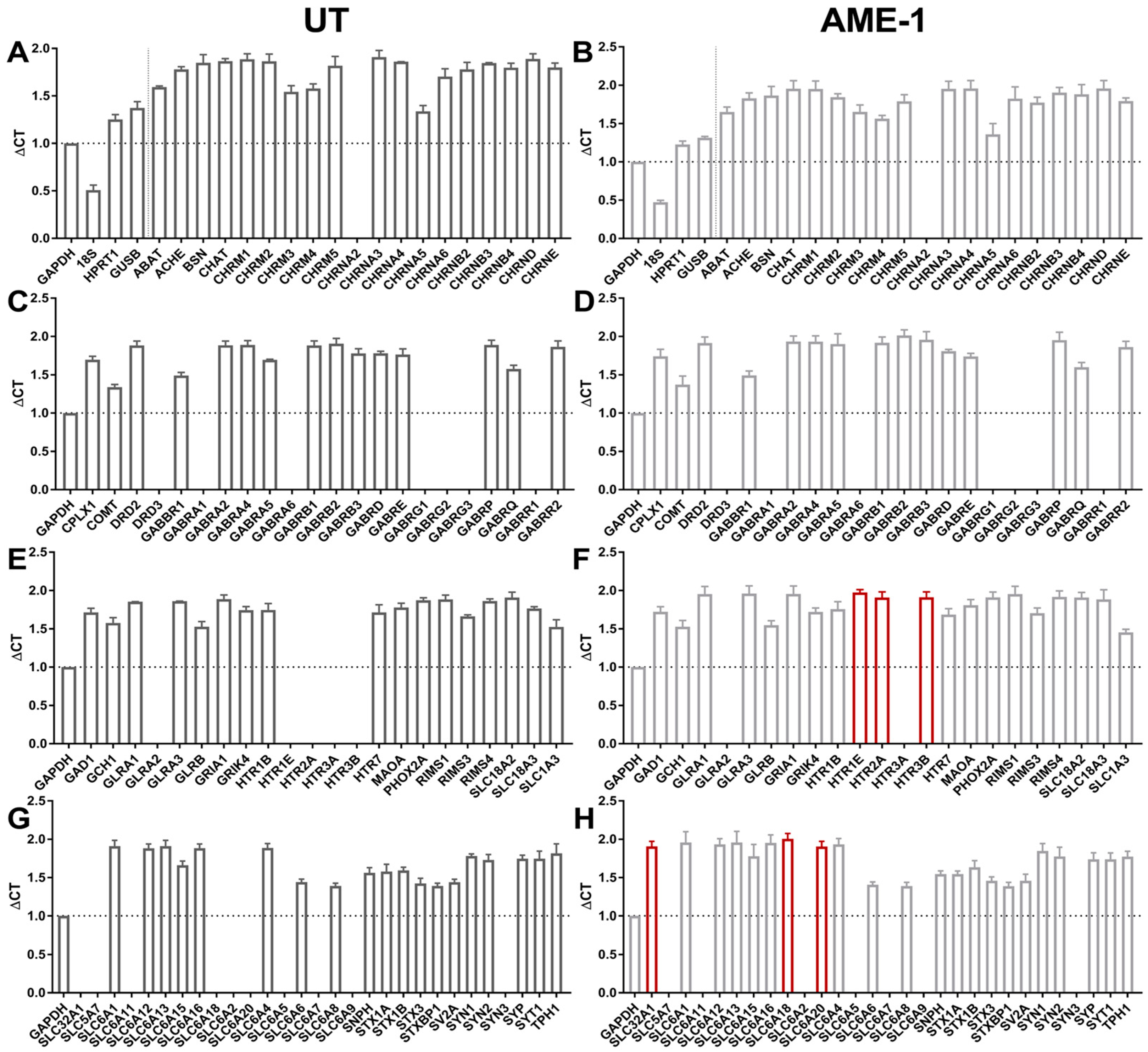
3.1. HMC3 Express Neuroreceptors
3.2. GABA and Muscimol Cause Changes in HMC3 Morphology and Iba1 Localization
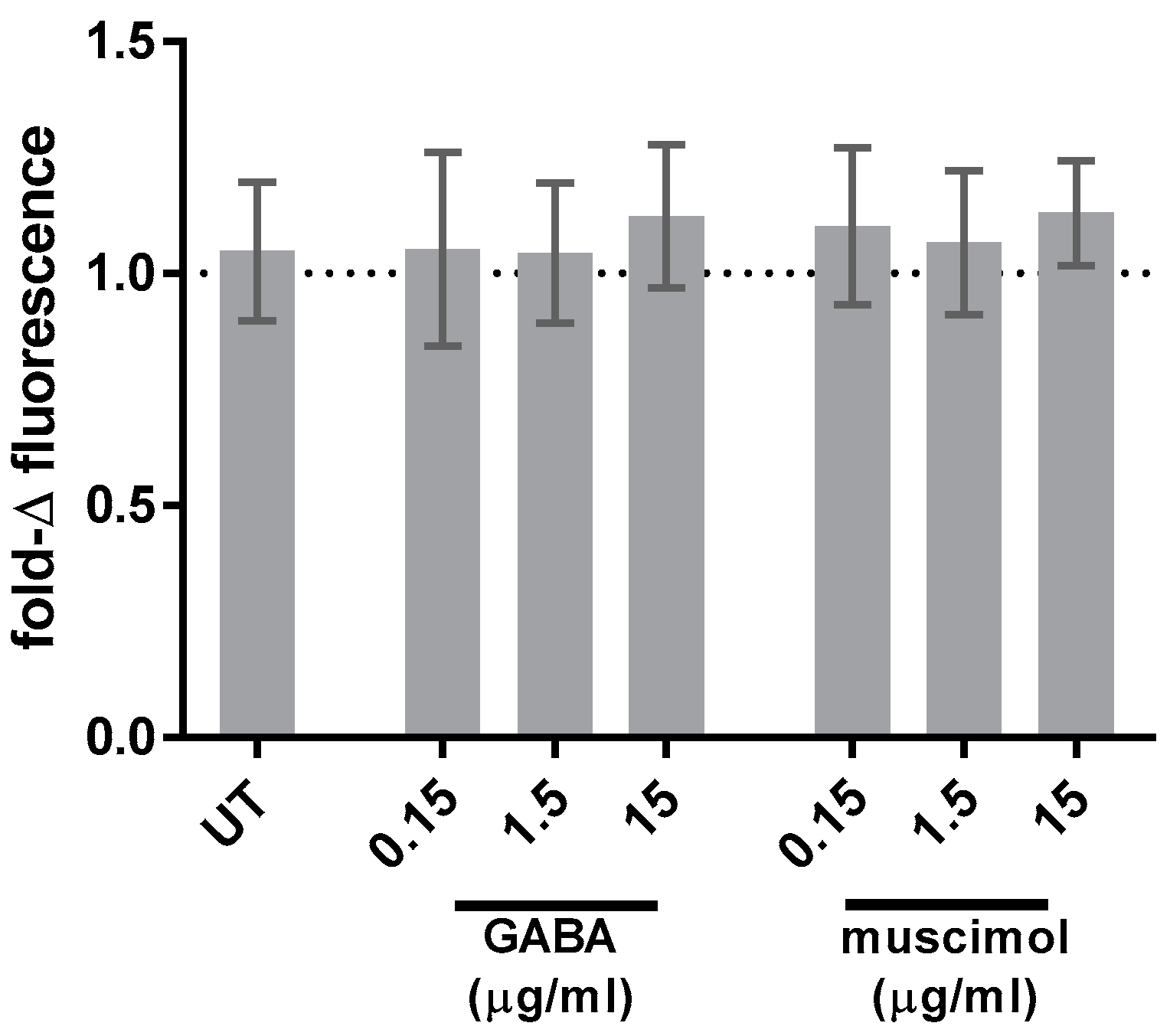
3.3. GABA and Muscimol Have No Effect on HMC3 ROS Generation
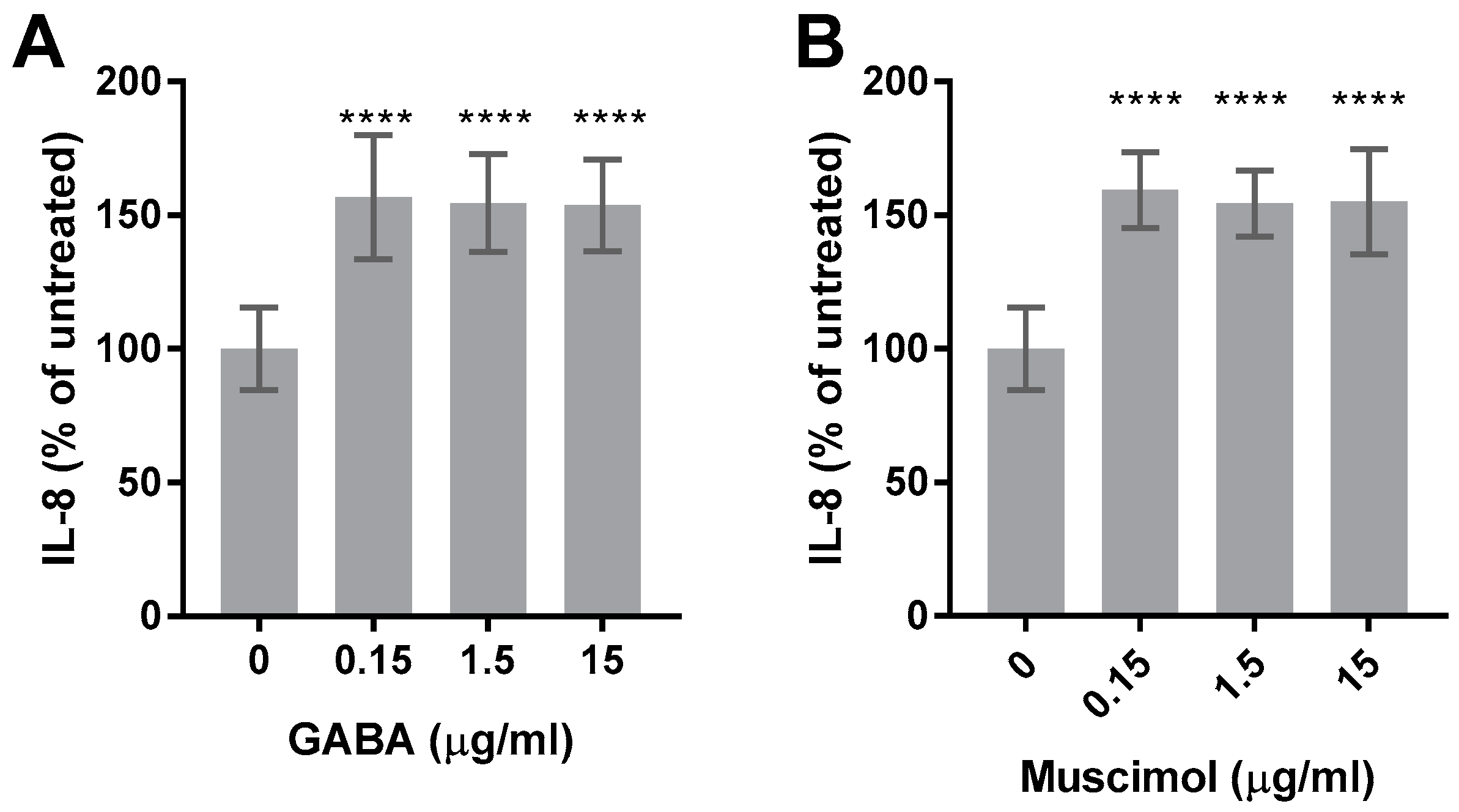
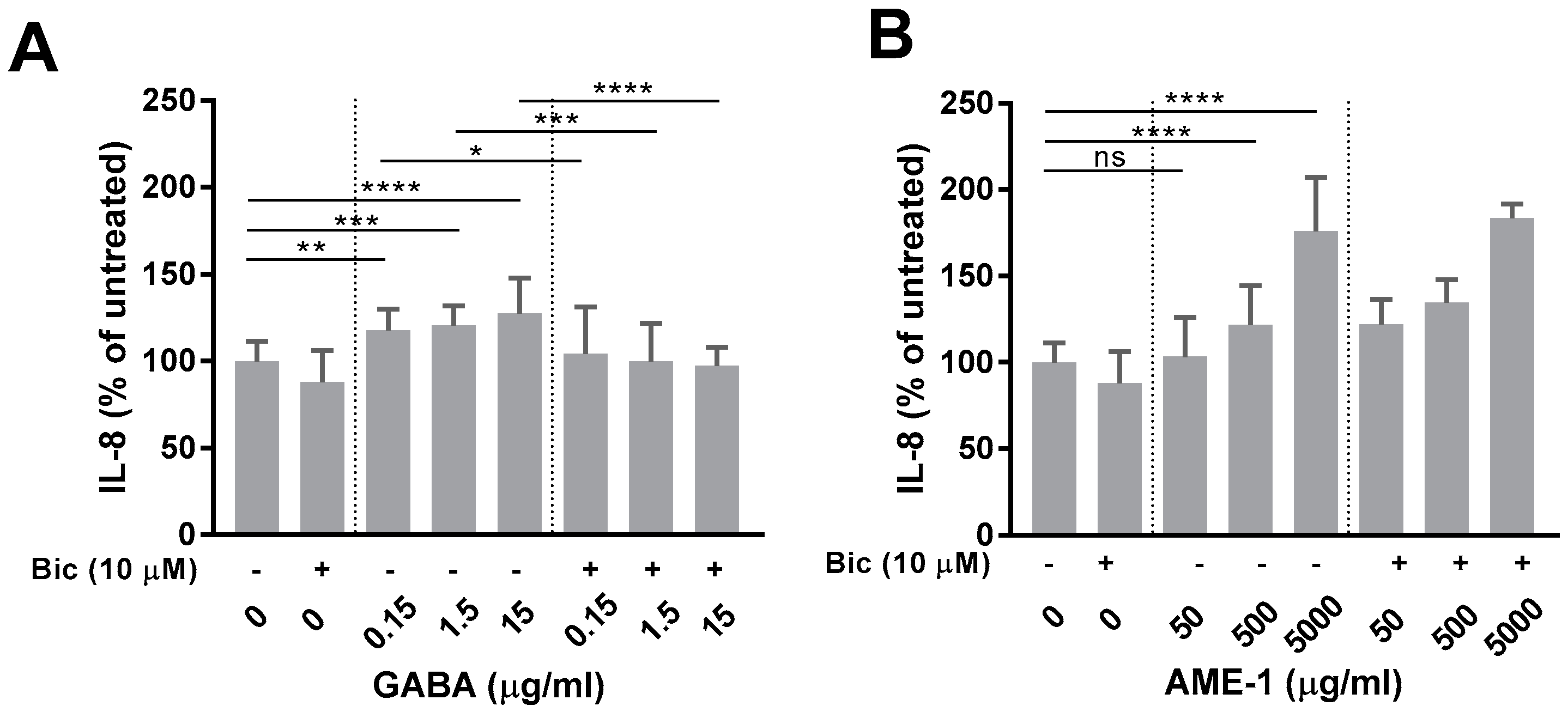
3.4. Muscimol and GABA Stimulate HMC3
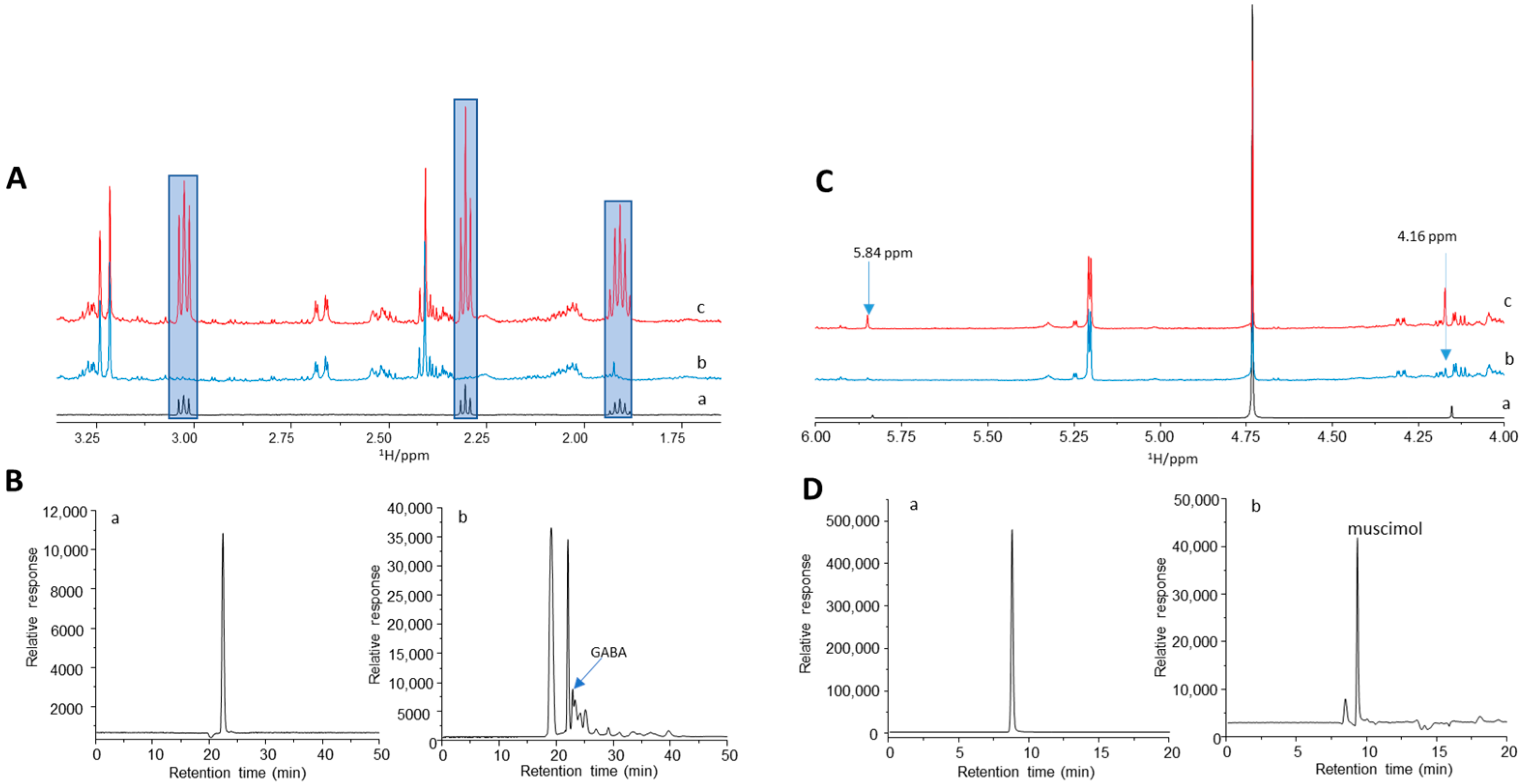
3.5. AME-1 Contains GABA and Muscimol
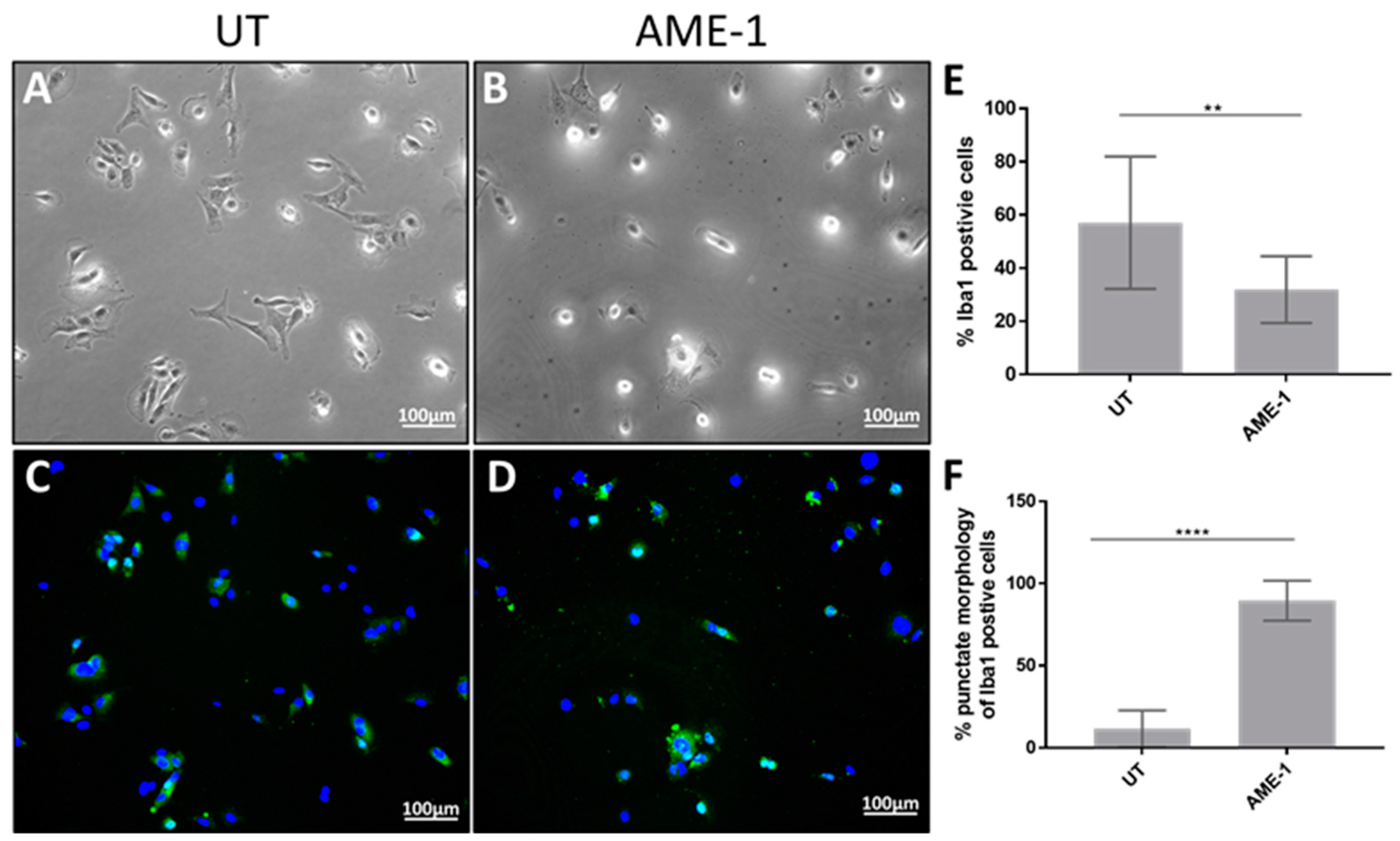
3.6. AME-1 Modifies HMC3 Morphology and IBa1 Expression Localization
3.7. AME-1 Modifies Neuroreceptor and Neurotransmitter Gene Expression by HMC3
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Batsukh, S.; Oh, S.; Rheu, K.; Lee, B.J.; Park, C.H.; Son, K.H.; Byun, K. Rice Germ Ameliorated Chronic Unpredictable Mild Stress-Induced Depressive-like Behavior by Reducing Neuroinflammation. Nutrients 2022, 14, 5382. [Google Scholar] [CrossRef]
- Liu, Y.U.; Ying, Y.; Li, Y.; Eyo, U.B.; Chen, T.; Zheng, J.; Umpierre, A.D.; Zhu, J.; Bosco, D.B.; Dong, H.; et al. Neuronal network activity controls microglial process surveillance in awake mice via norepinephrine signaling. Nat. Neurosci. 2019, 22, 1771–1781. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Shi, X.J.; Qi, L.; Wang, C.; Mamtilahun, M.; Zhang, Z.J.; Chung, W.S.; Yang, G.Y.; Tang, Y.H. Microglia and astrocytes mediate synapse engulfment in a MER tyrosine kinase-dependent manner after traumatic brain injury. Neural Regen. Res. 2023, 18, 1770–1776. [Google Scholar] [CrossRef]
- Maurya, S.K.; Gupta, S.; Mishra, R. Transcriptional and epigenetic regulation of microglia in maintenance of brain homeostasis and neurodegeneration. Front. Mol. Neurosci. 2022, 15, 1072046. [Google Scholar] [CrossRef]
- Dossi, E.; Huberfeld, G. GABAergic circuits drive focal seizures. Neurobiol. Dis. 2023, 180, 106102. [Google Scholar] [CrossRef]
- Tian, J.; Kaufman, D.L. The GABA and GABA-Receptor System in Inflammation, Anti-Tumor Immune Responses, and COVID-19. Biomedicines 2023, 11, 254. [Google Scholar] [CrossRef]
- Cheung, G.; Kann, O.; Kohsaka, S.; Faerber, K.; Kettenmann, H. GABAergic activities enhance macrophage inflammatory protein-1alpha release from microglia (brain macrophages) in postnatal mouse brain. J. Physiol. 2009, 587 Pt 4, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Kosarmadar, N.; Ghasemzadeh, Z.; Rezayof, A. Inhibition of microglia in the basolateral amygdala enhanced morphine-induced antinociception: Possible role of GABAA receptors. Eur. J. Pharmacol. 2015, 765, 157–163. [Google Scholar] [CrossRef]
- Silva, M.; Martins, D.; Tavares, I.; Morgado, C. Inhibition of spinal 5-HT3R reverted diabetes-induced mechanical hypersensitivity in a GABAAR-mediated neurotransmission-dependent manner. Neuroscience 2015, 304, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Schwab, C.; McGeer, P.L. Astrocytes are GABAergic cells that modulate microglial activity. Glia 2011, 59, 152–165. [Google Scholar] [CrossRef]
- Synowitz, M.; Ahmann, P.; Matyash, M.; Kuhn, S.A.; Hofmann, B.; Zimmer, C.; Kirchhoff, F.; Kiwit, J.C.; Kettenmann, H. GABA(A)-receptor expression in glioma cells is triggered by contact with neuronal cells. Eur. J. Neurosci. 2001, 14, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Mason, E.R.; Soni, D.M.; Chu, S. Microglial Phagocytosis/Cell Health High-Content Assay. Curr. Protoc. 2023, 3, e724. [Google Scholar] [CrossRef]
- Dello Russo, C.; Cappoli, N.; Coletta, I.; Mezzogori, D.; Paciello, F.; Pozzoli, G.; Navarra, P.; Battaglia, A. The human microglial HMC3 cell line: Where do we stand? A systematic literature review. J. Neuroinflamm. 2018, 15, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumbs, S.B.H.; Kubler, R.; Gharu, L.; Schipper, P.J.; Borst, A.L.; Snijders, G.; Ormel, P.R.; van Berlekom, A.B.; Wensing, A.M.J.; de Witte, L.D.; et al. Human microglial models to study HIV infection and neuropathogenesis: A literature overview and comparative analyses. J. Neurovirol 2022, 28, 64–91. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Pehar, M.; Yan, Z.; Kulka, M. Amanita muscaria extract potentiates production of proinflammatory cytokines by dsRNA-activated human microglia. Front. Pharmacol. 2023, 14, 1102465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.N.; Bast, T.; Xu, Y.; Feldon, J. Temporary inhibition of dorsal or ventral hippocampus by muscimol: Distinct effects on measures of innate anxiety on the elevated plus maze, but similar disruption of contextual fear conditioning. Behav. Brain Res. 2014, 262, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Sienkiewicz-Jarosz, H.; Szyndler, J.; Czlonkowska, A.I.; Siemiatkowski, M.; Maciejak, P.; Wislowska, A.; Zienowicz, M.; Lehner, M.; Turzynska, D.; Bidzinski, A.; et al. Rat behavior in two models of anxiety and brain [3H]muscimol binding: Pharmacological, correlation, and multifactor analysis. Behav. Brain Res. 2003, 145, 17–22. [Google Scholar] [CrossRef]
- Lee, M.R.; Dukan, E.; Milne, I. Amanita muscaria (fly agaric): From a shamanistic hallucinogen to the search for acetylcholine. J. R. Coll. Physicians Edinb. 2018, 48, 85–91. [Google Scholar] [CrossRef]
- Michelot, D.; Melendez-Howell, L.M. Amanita muscaria: Chemistry, biology, toxicology, and ethnomycology. Mycol. Res. 2003, 107, 131–146. [Google Scholar] [CrossRef]
- Rho, J.M.; Donevan, S.D.; Rogawski, M.A. Direct activation of GABAA receptors by barbiturates in cultured rat hippocampal neurons. J. Physiol. 1996, 497, 509–522. [Google Scholar] [CrossRef]
- Olsen, R.W.; Yang, J.; King, R.G.; Dilber, A.; Stauber, G.B.; Ransom, R.W. Barbiturate and benzodiazepine modulation of GABA receptor binding and function. Life Sci. 1986, 39, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Mercier, P.; Lewis, M.J.; Chang, D.; Baker, D.; Wishart, D.S. Towards automatic metabolomic profiling of high-resolution one-dimensional proton NMR spectra. J. Biomol. NMR 2011, 49, 307–323. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, M.K.; Simoncicova, E.; Carrier, M.; Tremblay, M.E. Microglia in Human Postmortem Brain Samples: Quantitative Ultrastructural Analysis of Scanning Electron Microscopy Images. Methods Mol. Biol. 2023, 2561, 63–85. [Google Scholar] [CrossRef] [PubMed]
- Lituma, P.J.; Woo, E.; O’Hara, B.F.; Castillo, P.E.; Sibinga, N.E.S.; Nandi, S. Altered synaptic connectivity and brain function in mice lacking microglial adapter protein Iba1. Proc. Natl. Acad. Sci. USA 2021, 118, e2115539118. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Bedard, K.; Sorce, S.; Hinz, B.; Dubois-Dauphin, M.; Krause, K.H. NOX4 expression in human microglia leads to constitutive generation of reactive oxygen species and to constitutive IL-6 expression. J. Innate Immun. 2009, 1, 570–581. [Google Scholar] [CrossRef]
- Sanjay, A.S.; Lee, H.-J. Honeyberry-derived carbon quantum dots ameliorate LPS-induced neuroinflammation and oxidative stress through Nrf2/HO-1 signalling in HMC3 cells. Artif. Cells Nanomed. Biotechnol. 2023, 51, 95–107. [Google Scholar] [CrossRef]
- Fernandez-Arjona, M.D.M.; Grondona, J.M.; Fernandez-Llebrez, P.; Lopez-Avalos, M.D. Microglial Morphometric Parameters Correlate With the Expression Level of IL-1beta, and Allow Identifying Different Activated Morphotypes. Front. Cell. Neurosci. 2019, 13, 472. [Google Scholar] [CrossRef] [Green Version]
- abd-el-Basset, E.; Fedoroff, S. Effect of bacterial wall lipopolysaccharide (LPS) on morphology, motility, and cytoskeletal organization of microglia in cultures. J. Neurosci. Res. 1995, 41, 222–237. [Google Scholar] [CrossRef]
- Jiang, X.; He, H.; Mo, L.; Liu, Q.; Yang, F.; Zhou, Y.; Li, L.; Su, D.; Yi, S.; Zhang, J. Mapping the Plasticity of Morphology, Molecular Properties and Function in Mouse Primary Microglia. Front. Cell. Neurosci. 2021, 15, 811061. [Google Scholar] [CrossRef]
- Wong, W.T.; Wang, M.; Li, W. Regulation of microglia by ionotropic glutamatergic and GABAergic neurotransmission. Neuron Glia Biol. 2011, 7, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, D.; Xu, J.; Tong, J.; Wang, Z.; Huang, L.; Yang, Y.; Bai, X.; Wang, P.; Suo, H.; et al. Tiagabine Protects Dopaminergic Neurons against Neurotoxins by Inhibiting Microglial Activation. Sci. Rep. 2015, 5, 15720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Tang, B.; Wang, L.; Huo, Q.; Tan, S.; Misrani, A.; Han, Y.; Li, H.; Hu, H.; Wang, J.; et al. Systemic LPS-induced microglial activation results in increased GABAergic tone: A mechanism of protection against neuroinflammation in the medial prefrontal cortex in mice. Brain Behav. Immun. 2022, 99, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Lang, L.; Xu, B.; Yuan, J.; Li, S.; Lian, S.; Chen, Y.; Guo, J.; Yang, H. GABA-mediated activated microglia induce neuroinflammation in the hippocampus of mice following cold exposure through the NLRP3 inflammasome and NF-kappaB signaling pathways. Int. Immunopharmacol. 2020, 89, 106908. [Google Scholar] [CrossRef]
- Hoerbelt, P.; Ramerstorfer, J.; Ernst, M.; Sieghart, W.; Thomson, J.L.; Hough, L.B.; Fleck, M.W. Mutagenesis and computational docking studies support the existence of a histamine binding site at the extracellular beta3+beta3- interface of homooligomeric beta3 GABAA receptors. Neuropharmacology 2016, 108, 252–263. [Google Scholar] [CrossRef]
- Yanovsky, Y.; Schubring, S.; Fleischer, W.; Gisselmann, G.; Zhu, X.R.; Lubbert, H.; Hatt, H.; Rudolph, U.; Haas, H.L.; Sergeeva, O.A. GABAA receptors involved in sleep and anaesthesia: Beta1- versus beta3-containing assemblies. Pflugers Arch. 2012, 463, 187–199. [Google Scholar] [CrossRef]
- Sente, A.; Desai, R.; Naydenova, K.; Malinauskas, T.; Jounaidi, Y.; Miehling, J.; Zhou, X.; Masiulis, S.; Hardwick, S.W.; Chirgadze, D.Y.; et al. Differential assembly diversifies GABA(A) receptor structures and signalling. Nature 2022, 604, 190–194. [Google Scholar] [CrossRef]
- Caruso, G.; Privitera, A.; Saab, M.W.; Musso, N.; Maugeri, S.; Fidilio, A.; Privitera, A.P.; Pittala, A.; Jolivet, R.B.; Lanzano, L.; et al. Characterization of Carnosine Effect on Human Microglial Cells under Basal Conditions. Biomedicines 2023, 11, 474. [Google Scholar] [CrossRef]
- Hristovska, I.; Pascual, O. Deciphering Resting Microglial Morphology and Process Motility from a Synaptic Prospect. Front. Integr. Neurosci. 2015, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogn, C.J.; Puchades, M.; Gundersen, V. Rare contacts between synapses and microglial processes containing high levels of Iba1 and actin--a postembedding immunogold study in the healthy rat brain. Eur. J. Neurosci. 2013, 38, 2030–2040. [Google Scholar] [CrossRef]
- Zhou, J.M.; Gu, S.S.; Mei, W.H.; Zhou, J.; Wang, Z.Z.; Xiao, W. Ginkgolides and bilobalide protect BV2 microglia cells against OGD/reoxygenation injury by inhibiting TLR2/4 signaling pathways. Cell. Stress. Chaperones 2016, 21, 1037–1053. [Google Scholar] [CrossRef] [Green Version]
- Lipovsky, M.M.; Gekker, G.; Hu, S.; Ehrlich, L.C.; Hoepelman, A.I.; Peterson, P.K. Cryptococcal glucuronoxylomannan induces interleukin (IL)-8 production by human microglia but inhibits neutrophil migration toward IL-8. J. Infect. Dis. 1998, 177, 260–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhter, R.; Shao, Y.; Formica, S.; Khrestian, M.; Bekris, L.M. TREM2 alters the phagocytic, apoptotic and inflammatory response to Abeta(42) in HMC3 cells. Mol. Immunol. 2021, 131, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, L.C.; Hu, S.; Sheng, W.S.; Sutton, R.L.; Rockswold, G.L.; Peterson, P.K.; Chao, C.C. Cytokine regulation of human microglial cell IL-8 production. J. Immunol. 1998, 160, 1944–1948. [Google Scholar] [CrossRef] [PubMed]
- Valekova, I.; Jarkovska, K.; Kotrcova, E.; Bucci, J.; Ellederova, Z.; Juhas, S.; Motlik, J.; Gadher, S.J.; Kovarova, H. Revelation of the IFNalpha, IL-10, IL-8 and IL-1beta as promising biomarkers reflecting immuno-pathological mechanisms in porcine Huntington’s disease model. J. Neuroimmunol. 2016, 293, 71–81. [Google Scholar] [CrossRef]
- Tsai, S.J. Role of interleukin 8 in depression and other psychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110173. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Count in Network 1 | Strength 2/False Recovery Rate 3 |
|---|---|---|
| Synaptic transmission involved in micturition | 3 of 3 | 2.44/8.19 × 10−5 |
| GABA biosynthesis process | 3 of 3 | 2.44/8.19 × 10−5 |
| Negative regulation of GABA secretion | 2 of 2 | 2.44/4.70 × 10−3 |
| Behavioral response to nicotine | 4 of 7 | 2.29/6.44 × 10−8 |
| GABA import | ||
| Phospholipase C-activating G protein-coupled acetylcholine receptor signaling | 2 of 3 | 2.26/7.50 × 10−3 |
| Adenylate cyclase-inhibiting G protein-coupled acetylcholine signaling | 5 of 8 | 2.24/1.02 × 10−7 |
| Exocytic insertion of neurotransmitter receptor to postsynaptic membrane | 3 of 5 | 2.22/2.00 × 10−4 |
| Regulation of GABA secretion | 4 of 7 | 2.2/5.99 × 10−6 |
| Synaptic transmission, glycinergic | 3 of 6 | 2.14/2.90 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, A.; Yan, Z.; Kulka, M. A Human Microglial Cell Line Expresses γ-Aminobutyric Acid (GABA) Receptors and Responds to GABA and Muscimol by Increasing Production of IL-8. Neuroglia 2023, 4, 172-187. https://doi.org/10.3390/neuroglia4030012
Wagner A, Yan Z, Kulka M. A Human Microglial Cell Line Expresses γ-Aminobutyric Acid (GABA) Receptors and Responds to GABA and Muscimol by Increasing Production of IL-8. Neuroglia. 2023; 4(3):172-187. https://doi.org/10.3390/neuroglia4030012
Chicago/Turabian StyleWagner, Ashley, Zhimin Yan, and Marianna Kulka. 2023. "A Human Microglial Cell Line Expresses γ-Aminobutyric Acid (GABA) Receptors and Responds to GABA and Muscimol by Increasing Production of IL-8" Neuroglia 4, no. 3: 172-187. https://doi.org/10.3390/neuroglia4030012