
The Signaling of Neuregulin-Epidermal Growth Factor Receptors and Its Impact on the Nervous System
1
Department of Clinical Pathology, Santa Maria Goretti Hospital, 04100 Latina, Italy
2
Department of Medical-Surgical Sciences and Biotechnologies, University of Rome Sapienza, 04100 Latina, Italy
*
Author to whom correspondence should be addressed.
Neuroglia 2023, 4(4), 253-274; https://doi.org/10.3390/neuroglia4040018
Submission received: 7 July 2023
/
Revised: 27 September 2023
/
Accepted: 3 October 2023
/
Published: 13 October 2023
(This article belongs to the Special Issue Glioblastoma (GBM) Brain Tumor Invasion and Consequences on Diagnosis, Clinical Strategies and Therapy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The activation of members of the Epidermal Growth Factor Receptor (EGFR) family (including ErbB) triggers pathways that have significant effects on cellular processes and have profound consequences both in physiological and pathological conditions. Within the nervous system, the neuregulin (NRG)/ErbB3 signaling plays a crucial role in promoting the formation and maturation of excitatory synapses. Noteworthy is ErbB3, which is actively involved in the process of cerebellar lamination and myelination. All members of the ErbB-family, in particular ErbB3, have been observed within the nuclei of various cell types, including both full-length receptors and alternative variants. One of these variants was detected in Schwann cells and in glioblastoma primary cells where it showed a neuregulin-dependent expression. It binds to promoters’ chromatin associated with genes, like ezrin, involved in the formation of Ranvier’s node. Its nucleolar localization suggests that it may play a role in ribosome biogenesis and in cell proliferation. The regulation of ErbB3 expression is a complex and dynamic process that can be influenced by different factors, including miRNAs. This mechanism appears to play a significant role in glioblastoma and is often associated with a poor prognosis. Altogether, the targeting of ErbB3 has emerged as an active area of research in glioblastoma treatment. These findings highlight the underappreciated role of ErbB3 as a significant receptor that can potentially play a pivotal role in diverse pathologies, implying the existence of a shared and intricate mechanism that warrants further investigation.
Keywords:
ErbB receptors; ErbB3; glioblastoma; glioma; neuregulin; brain; nucleolus; nucleolin; miRNA; myelination; tyrosine kinase inhibitors1. The Family of ErbB Receptors
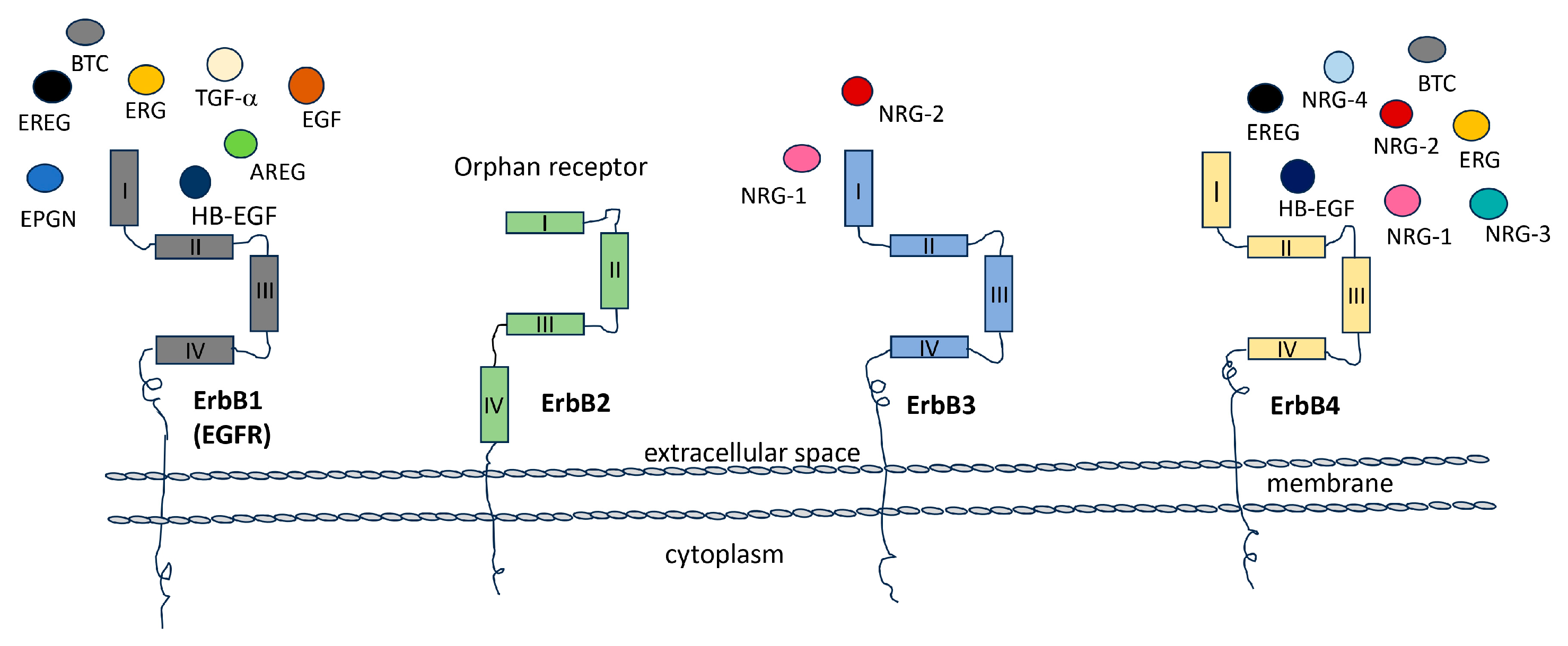
The Epidermal Growth Factor Receptors (EGFRs) are four tyrosine kinases that can establish a wide network of communication among them and between different cellular pathways. Thus, they regulate and control, in turn, either physiological or pathological conditions. The family of EGFRs consists of four members, ErbB1, ErbB2, ErbB3, and ErbB4 (ErbB: erythroblastic leukemia viral oncogene homolog), also called Her1, Her2, Her3, and Her4 respectively (Her: Human epidermal growth factor). These glycoproteins are transmembrane tyrosine kinase receptors that, once activated, regulate important intracellular pathways involved in cell division, proliferation, differentiation, migration, angiogenesis, apoptosis, and survival. They can also influence cellular behavior in response to extracellular signals [1]. Substantial evidence supports the involvement of ErbB family in the development and progression of various types of cancer. ErbBs are frequently found in elevated levels in tumors [2,3]. In numerous cells, multiple members of the ErbB receptor family are co-expressed. Upon stimulation with growth factor ligands, these receptors can form both homo- and heterodimers [4]. Specifically, ErbB receptors can arrange into homodimers (ErbB1 and ErbB4) or, predominantly, heterodimers (ErbB1/ErbB2, ErbB3/ErbB2, and ErbB4/ErbB2), initiating unique intracellular signaling pathways. There are more than ten distinct ligands that activate ErbB receptor family members. Among them, three have been categorized as EGF agonists due to their specific binding to ErbB1 (EGF, TGF-α, and amphiregulin), four are specific to ErbB3 and/or ErbB4 (neuregulins (NRGs) also known as heregulin (HRG), while another three are bispecific, capable of binding to both ErbB1 and ErbB4 (betacellulin, epiregulin, and heparin-binding EGF-like) [4,5,6] (Figure 1). The NRG gene family comprises four members: NRG-1, NRG-2, NRG-3, and NRG-4. Within this gene family, numerous isoforms are generated through alternative exon splicing. These isoforms exhibit diverse tissue distributions and have various biological activities [7]. In most cases, these NRG proteins are initially synthesized as precursors and subsequently undergo activation and release from the cell membrane through specific proteolytic processes, [8]. The subsequent activation of various pathways and crosstalk between them, either within the same family or among different receptors, remains an intriguing challenge [9,10].
They are functionally dependent and complementary, playing a significant role in heterodimer formation following ligand binding. This process is tightly controlled and is involved in various physiological conditions including embryonic development, as well as aggressive tumor scenarios like mammary lung cancer and glioblastoma [11]. Of particular interest is the relationship between ErbB2 and ErbB3, which, when functioning independently, are incomplete (with ErbB2 lacking the ligand binding site and ErbB3 lacking tyrosine kinase activity). However, when they bind with the NRG, they undergo heterodimerization and generate a potent activation signal. Furthermore, the interaction between ErbB3 and ErbB1 can have important implications for cancer biology and therapeutic strategies. Studies have shown that both ErbB3 and ErbB1 are frequently overexpressed in glioblastoma, the most common and aggressive form of glioma [11]. This receptor’s overexpression can lead to heterodimer formation and the activation of downstream signaling pathways, including the PI3K (phosphatidylinositol 3-kinases)-AKT (PKB—protein kinase B) and MAPK (mitogen-activated protein kinase) pathway, which can promote cell survival, proliferation, and invasion, and can contribute to glioma development and progression [12]. Overall, the interaction between ErbB3 and ErbB1 in glioblastoma is complex and multifaceted. Further research is needed to fully understand the nature and significance of this interaction and its potential as a therapeutic target.
To conclude, this family of receptors, along with its downstream pathway encompassing ligands and targets, holds significant relevance in numerous contexts, both physiological and pathological. Despite the multitude of studies dedicated to unraveling these intricate crosstalk interactions, further endeavors are required to fully comprehend the breadth of this mechanism.
2. ErbB Receptor Structure
Studies of the structures of the EGFR family allow us to understand conserved strategies of action, introducing strengths and weaknesses of the pathways and of targeting both. The general structure of the EGFRs is made of an endo-domain and an ecto-domain. The first domain is in turn composed of a transmembrane domain (23 aa), a juxta membrane domain (40 aa), and a tyrosine kinasic domain with the C-terminal tail (232 aa). The last domain is involved in the interaction between the receptor and the ligands and is composed of the N-terminal glycosylated extracellular domain (620 aa) [13]. After ligand binding, EGFRs interact with each other and form homo- or heterodimers, which then activate the intrinsic tyrosine kinase activity. This leads to the trans-phosphorylation of the receptors’ intracellular domains, which in turn recruit proteins involved in cytoplasmic signaling pathways associated with domain phosphorylation. Otherwise, with a partial homology sequence, the precise domain sequences vary in the EGFRs and lead them to acquire unique features. Higher analogy domain sequences are those that perform more similar functions in the four receptors, hence, less conserved domain sequences are those that perform different functions [10]. The highest grade of homology (64–67%) is verifiable in catalytic domains which form the essential structure for the tyrosine kinase activity [14]. This explains why several molecular inhibitors that bind to STP-binding cassettes target more than one ErbB-receptor [15]. The domains responsible for the interaction with the ligand show 43–45% homology between ErbB3, ErbB2; and ErbB1; in fact, they interact with similar ligands. The extracellular domains of ErbB3 show 50 conserved cysteine residues compared to ErbB2 and ErbB1, as well as ten partially conserved glycosylation sites. Interestingly, up to 30% of the entire molecular weight of ErbB3 is composed of the glycosidic groups. Just one of those glycosylation sites is conserved, which suggests that this modification contributes to the functional uniqueness of this receptor [16]. Glycosylation is important for ligand-dependent and -independent function of ErbB receptors, but also for protein stability and signaling. Mutation in the conserved glycosylation site (Asn to Gln) in a CHO cell line led to auto-dimerization and heterodimerization with ErbB2 even in the absence of the ligand and to the acquisition of neoplastic features by the cells [16]. Glycosylation in Asn-418 hampers tumor progression, stopping aberrant ErbB2-ErbB3 heterodimerization [10]. The proximal site of the transmembrane domain (21 aa) is very conserved. The cytoplasmic domain (277 aa) shows 60–62% sequence homology between ErbB3, ErbB1, and ErbB2 and overlaps with the tyrosine kinase catalytic domain. There is also one binding site for ATP (position 716–721) and a lysine residue with the C-terminal part conserved compared to the other members of the EGFR family. The most divergent region between ErbB3, ErbB2, and ErbB1 comprises 364 aa: Tyrosine residues (position 1197, 1199, and 1262) in proximity of phosphorylation sites and YEYMN repeated sequences (position 1260–1264). These hallmarks make this region the possible auto-phosphorylation site of ErbB3 protein [14]. ErbB4 is unique among ErbB receptors because it goes through alternative splicing. There are four different versions of ErbB4 mRNA obtained by two splicing locations: one in the outer part near the membrane and the other in the inner part, beyond the tyrosine kinase domain.
In summary, EGFRs share a common structural framework and a universal mechanism of activation and signaling, consisting of the following steps: ligand binding, homo- or heterodimerization, activation of the intrinsic tyrosine kinase activity, trans-phosphorylation of the intrinsic domain, and recruitment of associated proteins. It is worth noting that sequence variations lead the receptor to acquire distinctive characteristics alongside their shared functions. Among these shared features, certain targeting strategies can be advantageous or detrimental. These diversifications are prominently exemplified in the glycosylation sites of ErbB3, which can affect both ligand-dependent and -independent functions, as well as stability and signaling. Additionally, tyrosine residues near the phosphorylation site play a role as auto-phosphorylation sites, further illustrating the complexity of these receptors.
3. ErbB Receptors and Neural Development
Neuregulin/EGFR signaling is heavily involved in various aspects of neural development, from synapse formation to cellular differentiation, myelination, and establishment of pathways involved in psychiatric disorders. The members of the ErbB receptors family have been implicated in various aspects of neural development. NRG/ErbB signaling has been implicated in neural development, including circuit generation, axon lining, neurotransmission, and synaptic plasticity. Several members of this signaling network are encoded by genes involved in psychiatric disorders [17]. NRG/ErbB signaling also contributes to synapse formation and promotes the formation and maturation of excitatory synapses on GABAergic interneurons [18,19,20,21]. NRGs function by activating ErbB tyrosine kinases, such as ErbB2, ErbB3, and ErbB4 [22]. Specifically, the ErbB2 and ErbB4 receptors are involved in glial differentiation, which is necessary for the radial migration of neurons in the cortex and cerebellum, respectively [23]. Undifferentiated neural stem cells in the nervous system express high levels of ErbB1 or ErbB2 [24]. Following neural development and maturation, the level of ErbB1 decreases, and ErbB4 expression is elevated, both of which are limited to midbrain dopaminergic neurons and GABAergic cells [25]. In the glial lineage, ErbB1 expression is replaced by ErbB3 during development. The interactions of NRGs expressed on the neuronal surface with ErbB3 in oligodendrocytes promote their differentiation and myelination [26]. ErbB1 is distributed in dopaminergic neurons in the substantia nigra compacta, as well as in the ventral tegmental area [24]. Similarly, ErbB4 is expressed in the same regions but not always in the same dopaminergic cell population [25]. NRG1 has been linked to neural development and brain activity homeostasis. Nevertheless, the simultaneous knockout of ErbB2 and ErbB4 has no effect on cortical and cerebellar lamination [27,28]. Conversely, ErbB4 plays a critical role in the assembly of GABAergic circuitry [29,30,31,32], while ErbB3 is necessary for myelination in the central and peripheral nervous systems [33,34,35] (Figure 2). Although ErbB3 is expressed in the developing brain, its role in central nervous system development remains unknown due to the embryonic lethality caused by null mutation [36]. The NRG-ErbB signaling pathway appears to be common among mammals, functioning in both mice and humans. NRG plays a crucial role as a neuronally-released factor that activates microglia in the central nervous system (CNS). This activation is observed in response to experimental peripheral nerve injuries and degenerative diseases in both human and mouse models.
In brief, the signaling pathways activated by NRGs and ErbB receptors play a crucial role in various aspects of neural development, maturation, and the maintenance of proper function and homeostasis, both in the central nervous system and the peripheral nervous system. These receptors are dedicated to distinct functions, leading to their expression in different locations and at various developmental stages, all highly coordinated. However, a comprehensive understanding of the precise roles that these receptors play in the brain remains a formidable challenge, as variations in their expression can potentially result in significant disorders.
4. Focusing on ErbB3
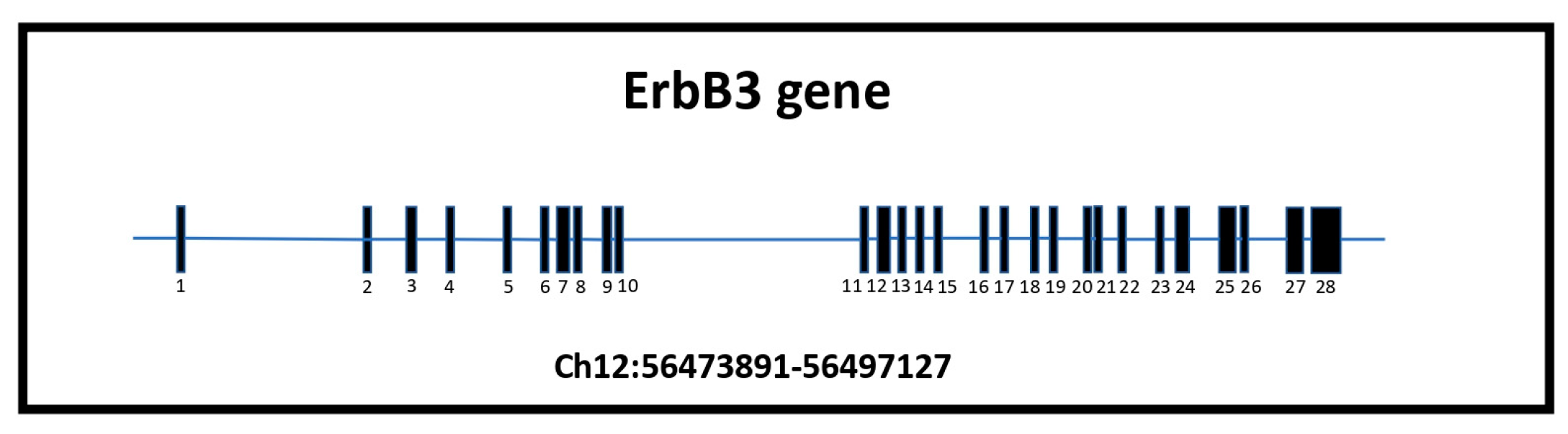
ErbB3 is the one receptor of the family that shows unique features that are worth focusing on. The ErbB3 gene is localized on human chromosome 12q13.2, a region of particular interest for specific translocation [37,38,39]. It is characterized by 28 exons and 28.2 kb, approximately 75 kb of genomic DNA (NCBI Gene ID2065) (Figure 3). The ErbB3 mRNA consists of 6.2 kb, 4080 nucleotides, and 1342 codons. The reading frame is from position 46 to 4126. The first ATG is in the 100th position contributing to the formation of a 148 kDa polypeptide that represents the full-length transmembrane receptor that is strictly correlated to ErbB1 and ErbB2. ErbB3 usually weighs around 185 kDa because it undergoes significant glycosylation at ten possible N-linked glycosylation sites [37,38,39,40].
Nevertheless, nucleotide analysis reveals three reading frames. ErbB3 gene expression is under control of its promotor characterized by high contents of GC nucleotides (about 65%), by the absence of a TATA box, and by the presence of more than one starting transcription site [41]. Furthermore, the promoter region of the ErbB3 gene contains multiple regulatory elements that control its transcriptional activity, including binding sites for various transcriptional factors such as AP-1 (Activator Protein-1), NF-KB (nuclear factor kappa-light-chain-enhancer of activated B cells), and E2F transcriptional factor. The promoter is more active in cells that show major ErbB3 expression that could be regulated by a variety of stimuli, including growth factors, hormones, and environmental stress [42,43]. The extracellular domain of ErbB3 (from aa 20 to 630) has some unique features that could be triggered for its different functions. It is made of four sub-domain b-helix right-handed or solenoid structures: I-L1, II-C1, III-L2, IV-C2. Sub-domain I and III are Leucine Rich Repeat members and are the deputies to interact with the ligands. Subdomains II and IV are rich in cysteine and are characterized by extended structures with disulfide bonds that set up the structure [13]. When the ligand is absent, domains I and IV interact with each other, making up a β-hairpin structure that confers a closed conformation that does not lead to ligand-binding and dimerization. Instead, when the ligand is present, the extracellular domain takes a more extended conformation that permits binding the ligand, and then heterodimerization with ErbB2, producing strongly mitogenic signals [13,44]. The wide conformation may require additional events or the simultaneous binding of two ligand molecules for its final action. It has been suggested that inactive ErbB3 exists in oligomerized pools that are built via ligand-independent mechanisms in the plasma membrane. NRG1β binding to ErbB3 can destabilize these oligomers, freeing ErbB3 to heterodimerize with a partner (typically ErbB2). The affinity of the binding between ErbB3’s extracellular domain and its favorite ligand NRG is very high and increases after heterodimerization with ErbB2. It remains high even at endosomal pH (pH 5.5) [3,45]. The transmembrane domain is characterized by a single a-helix chain which seems to interfere with the setting up of ligand binding and the induction of the receptor’s dimerization [46]. In this action, the GXXXG motifs appear important: they permit a wide interface area between adjacent helix, bring them near each other and create a plane surface that allows binding between the side chains. ErbB3 is the only one in the family that has a unique sequence like the one mentioned above, namely TXXXG, in the transmembrane domain. It seems to be the motif that reflects less aptitude to form oligomers as to promote heterodimerization [47]. Protein kinase enzymes show a catalytic domain with a great number of conserved amino acids suggesting their role in the catalytic function. Otherwise, ErbB3 presents some variation in three residues of this domain, suggesting a lower kinase activity but also new possibility of interaction with ErbB2 [48,49]. The catalytic domain of c-erbB3 (cloned form of ErbB3) shows a homology sequence of about 60–63% compared to the other ErbB family members [50]. The gene encoding c-erbB-3 was cloned almost simultaneously by two groups [50,51]. It encodes a 160–180 kDa glycoprotein which has all of the structural features of the EGF family of tyrosine kinase receptors [52]. The highest proportion of sequence identity with c-ErbB-3 is within the catalytic domains of EGFR (60%), c-erbB-2 (cloned form of ErbB2) (62%), and c-erbB-4 (cloned form of ErbB4) (63%). The substitution in E740N and D815N leads to a reduction in the auto-catalytic function as compared to the wild type of the receptor either in vitro or in vivo [53]. Mutations significantly increase the ability to interact with the SH2 (Src-Proto-oncogene tyrosine-protein kinase—Homology 2) domain of Syp and PI3K. Furthermore, ErbB3 escapes from Protein Kinase C (PKC) regulation because of the lack of regulatory threonine which is present in the other Ebbs’ members [51]. Based on these observations, it is suggested that ErbB3 is a pseudo-kinase that acts like a scaffold protein that mediates the assembly of signaling complexes or as an allosteric regulator of other proteins. Lacking tyrosine kinase activity, ErbB3’s cytoplasmic region is preserved, and it is thought to play a role in the interaction between ErbB3 with other proteins thanks to the consensus sequence for ATP binding (aa 716–721). c-ErbB3 interacts with SHC-transforming protein 1 [51], which is involved in apoptosis, drug resistance, mitogenesis, carcinogenesis and metastasis. Another possible interaction is between c-ErbB3 and the p85-PI3K subunit that is involved in cell growth, proliferation, differentiation, motility, and survival. c-ErbB3 contains six potential binding sites (“YMXM” or “YXXM” sequences) with p85-PI3K in the C-terminal domain and one of these in the tyrosine kinase domain [52]. Numerous tyrosine residues situated proximally to phosphorylation sites, along with recurring YEYMN sequences, potentially associated with auto-phosphorylation or uncharacterized functions, may initiate transduction pathways.
In summary, full-length ErbB3 represents a predicted 148 kDa polypeptide intimately linked to its family counterparts. The extracellular domain, entrusted with ligand binding, demonstrates robust affinity, and possesses distinctive features that can be harnessed for diverse functional roles. This can result in reduced kinase activity and affect interactions not only with ErbB2 but also with Syp and PI3K. Furthermore, the absence of regulatory threonine allows ErbB3 to elude PKC regulation. The interactions it orchestrates, thanks to these distinctive features, play pivotal roles in numerous critical cellular events.
5. Erbb3 and Glial Cells
The EGFRs are committed in different functions in the central nervous system (CNS) and peripheral nervous system (PNS), so that they are expressed in different localization, at different stages of development in a very coordinated way [54,55,56]. Rat models suggest that the Erbb3 receptor plays a critical role in brain development, and its expression begins as early as E16 in the brain, reaching a peak around P20. This temporal expression pattern correlates with the process of cerebellar lamination, which takes place from E15 to P21 [32,56]. However, as they age, the mutants show a smaller increase in brain weight compared to controls. Approximately 90% of mutant mice had significantly smaller brains than controls, indicating that Erbb3 mutant mice have impaired motor skills [57,58]. Interestingly, mice Erbb3 heterozygous mutants are healthy and fertile at birth. These findings suggest that brain regions involved in motor control and coordination might be altered by ErbB3 mutation. Motor impairments are often caused by disorders of the cerebellum [57,58,59,60]. The results suggest that the phenotypes of the Erbb3-mutants may be due to the lack of Erbb3 in glial cells in the cerebellum. Cerebellar glial cells are derived from glial precursors in the ventricular zone of the fourth ventricle and express glial fibrillary acidic protein (GFAP) [57,58]. Erbb3 was detected in GFAP + astroglial cells and was absent in astrocytes purified from the cerebella of GFAP: Cre; Erbb3F/F mice. The Erbb3 receptor is expressed in cerebellar astrocytes but not in granule neurons (GNs) [59,60]. Erbb3 ablation impairs the expansion of cerebellar glial cells. The number of mutant cerebellar astroglial cells is only 50% of the control at DIV12. The decrease in the number of mutant Erbb3 cerebellar astroglial cells is caused by impaired proliferation rather than increased apoptosis. The existence of multiple NRG ligands for ErbB3 suggests functional redundancy, which adds a layer of complexity in delineating the role of each NRG in neural development. In addition to NRG1, two other NRG proteins, namely NRG2 and NRG6, bind to ErbB3 and are expressed in the cerebellum [61,62,63]. Nevertheless, previous studies indicated that ErbB2 and ErbB4 are not essential for cerebellar lamination [27,28]. Therefore, it is possible that ErbB3’s role in regulating cerebellar lamination operates independently from the other ErbB receptors.
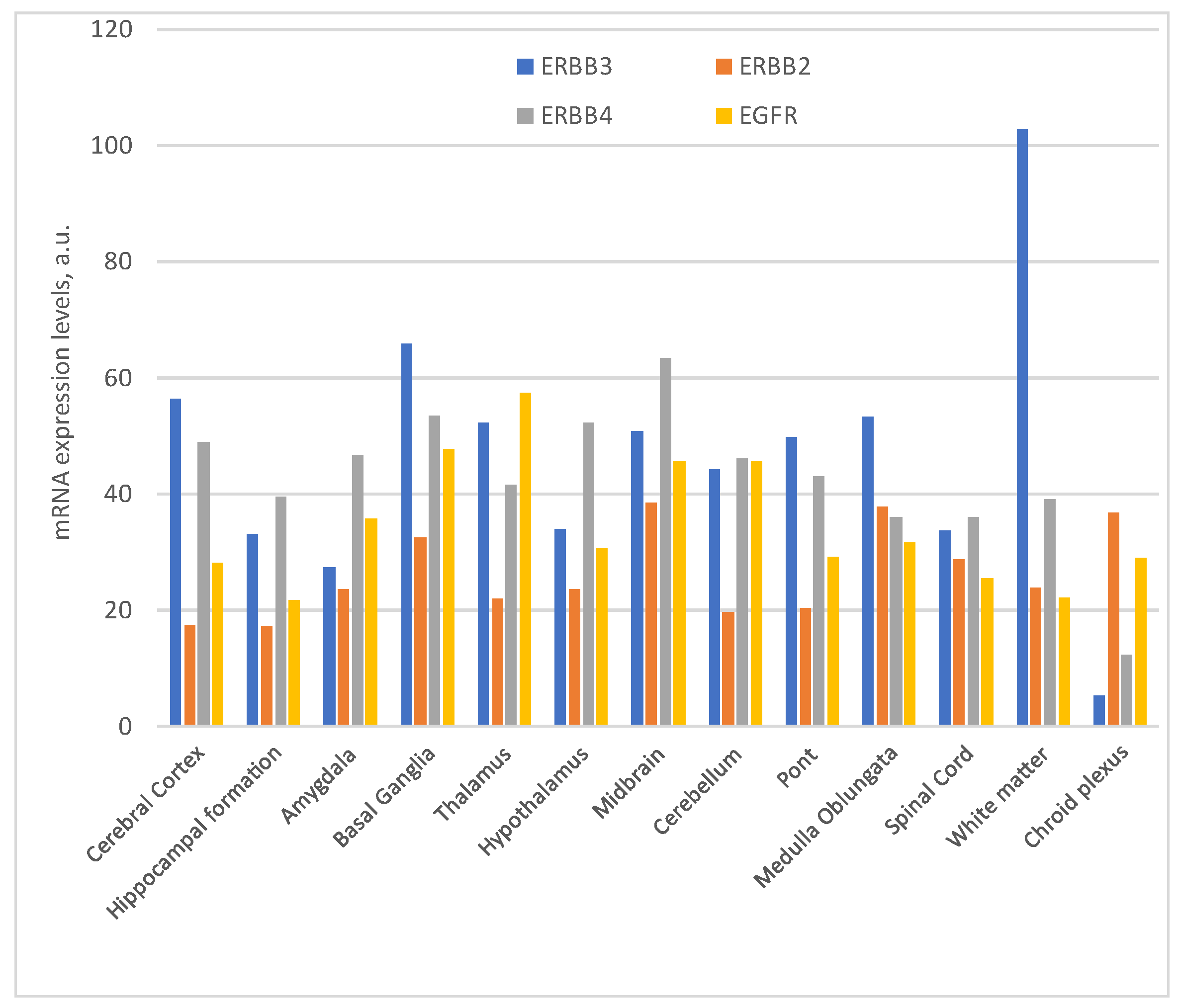
The distribution and significance of ErbB receptors have been extensively investigated within the avian and murine central nervous systems [63,64]. A recent study has extended this inquiry to the primate nervous system, elucidating the distribution of ErbB2, ErbB3, and ErbB4 mRNA and protein in the primate brain, uncovering significant variations in mRNA expression. Notably, that research unveiled the presence of ErbB proteins in both neurons and glial cells within the frontal cortex, with distinct ErbB antibodies strongly staining the neuronal nucleus. This distribution was widespread, encompassing cortical, hippocampal, and subcortical regions throughout the primate brain. The coexistence of ErbB2 and ErbB4 in multiple brain areas, including the cortex, hippocampus, and basal ganglia, suggests the potential for ErbB2 and ErbB4 dimerization across various brain regions [65]. In contrast to the pronounced ErbB2 mRNA levels in grey matter regions, ErbB3 mRNA expression exhibited a lower, more uniform distribution. Remarkably, unlike prior reports, several studies revealed comparable ErbB3 mRNA expression levels in both grey and white matter in juvenile and adult monkeys [65]. Nevertheless, they did observe a stronger ErbB3 mRNA signal in white matter, particularly in one infant monkey and several adult human brains. This hints at the possibility that ErbB3 mRNA may exist at reduced levels in the white matter of post-neonatal macaque brains relative to other species. Importantly, all three ErbB receptor proteins were detected in both pyramidal and non-pyramidal neuron types throughout the brain, aligning with previous observations in rats.
Collectively, these findings underscore the widespread and abundant expression of ErbB receptors, suggesting their ongoing involvement in modulating neurotransmission in the postnatal primate brain [66]. Of note is the pivotal role played by the ErbB3 receptor in brain development, with its expression pattern correlating closely with cerebellar lamination processes, where it appears to be the sole receptor within the ErbB family overseeing this crucial function. However, as they aged, a notable increase in brain weight and motor skill impairments emerged, potentially attributed to the absence of ErbB3 in cerebellar glial cells, leading to compromised cellular proliferation. Adding another layer of complexity, it is worth noting the variety of NRGs that can bind to ErbB3 receptors.
6. ErbB3 and Myelination
NRG/ErbBs signaling is also involved in the myelination process. An alteration at this level can have important impacts on neural development. In higher vertebrates, myelin sheaths play a crucial role in facilitating the transmission of nerve impulses, which are essential for normal motor and sensory functions. Myelin enables rapid propagation of impulses along axons, allowing for long-distance connectivity [66,67]. The ErbBs signaling pathways are important in facilitating communication between projection neurons and axon-associated glial cells to promote myelination. Schwann cells express both ErbB2 and ErbB3 but not ErbB4. During the myelination process, oligodendrocytes in the central nervous system (CNS) and Schwann cells in the peripheral nervous system (PNS) are engaged in complex interactions with axonal segments, leading to the formation of myelin sheaths around the axons [67]. This results in the formation of internodes that are regularly separated by nodes of Ranvier [68,69,70]. In the PNS, axonal NRG1 serves as a crucial signaling molecule that regulates the behavior of myelinating Schwann cells. When Schwann cells are cultured, they can be stimulated by purified axonal membranes, leading to their proliferation [71,72,73,74]. In vivo, the Schwann cell critical growth factor is NRG1 Type III, which remains associated with axonal membranes [74,75]. It is the most prominent isoform of NRG1 in motoneurons of the spinal cord and sensory neurons of the dorsal root ganglia. In fact, NRG1 null mutant mice die with developmental heart failure at E11 without significant nervous pathology. The mutant not only lacks Schwann cells but also exhibits a loss of ErbB3-negative sensory dorsal root ganglia (DRG) neurons and spinal motor neurons [76,77,78]. The first in vivo evidence for the critical function of NRG1/ErbB signaling in the PNS came from mutant mice lacking Erbb3 [36]. Some mutant embryos develop until birth but die shortly after, exhibiting lethal neuropathy characterized by breathing defects and defects in the regulation of functional Transient Receptor Potential Vanilloid 1—Receptor (TRPV1) along sensory neuron axons [78]. However, the most important role of Neuregulin 1 (NRG1) lies in controlling Schwann cell development and peripheral myelination. Mutant embryos lacking in specifically this membrane-associated isoform can develop to term [79,80,81,82]. However, perinatal death in these mice is caused by a significant absence of Schwann cells and a severe reduction in dorsal root ganglia (DRG) neurons and motor neurons, as observed in Erbb3 mutants. In a gene-targeting experiment, the floxed ErbB2 gene was selectively eliminated in Schwann cells expressing a Krox20-Cre (also called Early Growth Response Protein 2, EGR2) transgene, resulting in a severe neuropathy phenotype that confirmed the critical role of ErbB2 signaling during cell myelination [83]. Inactivation of the floxed Erbb3 gene in Cnp-Cre mice led to even more severe peripheral dysmyelination [32], suggesting that the absence of ErbB2 in Schwann cells can be partially compensated by the presence of ErbB3 [84]. NRG1 Type III is targeted to long axonal projections and serves multiple functions [84].
In summary, ErbB signaling pathways, particularly involving ErbB2 and ErbB3, play a crucial role in facilitating communication between projection neurons and axon-associated glial cells to support the process of myelination. A pivotal player in this intricate process is NRG1, which associates with axonal membranes and is deemed the critical growth factor in vivo. Its primary function is to regulate the development of Schwann cells and peripheral myelination. This isoform exhibits prominent expression in motor neurons of the spinal cord and sensory neurons within the dorsal root ganglia. The inactivation of ErbB3 leads to more severe peripheral dysmyelination compared to ErbB2, implying a compensatory role of ErbB3, but not ErbB2, in this vital signaling pathway.
7. ErbB3 and Alternative Variants
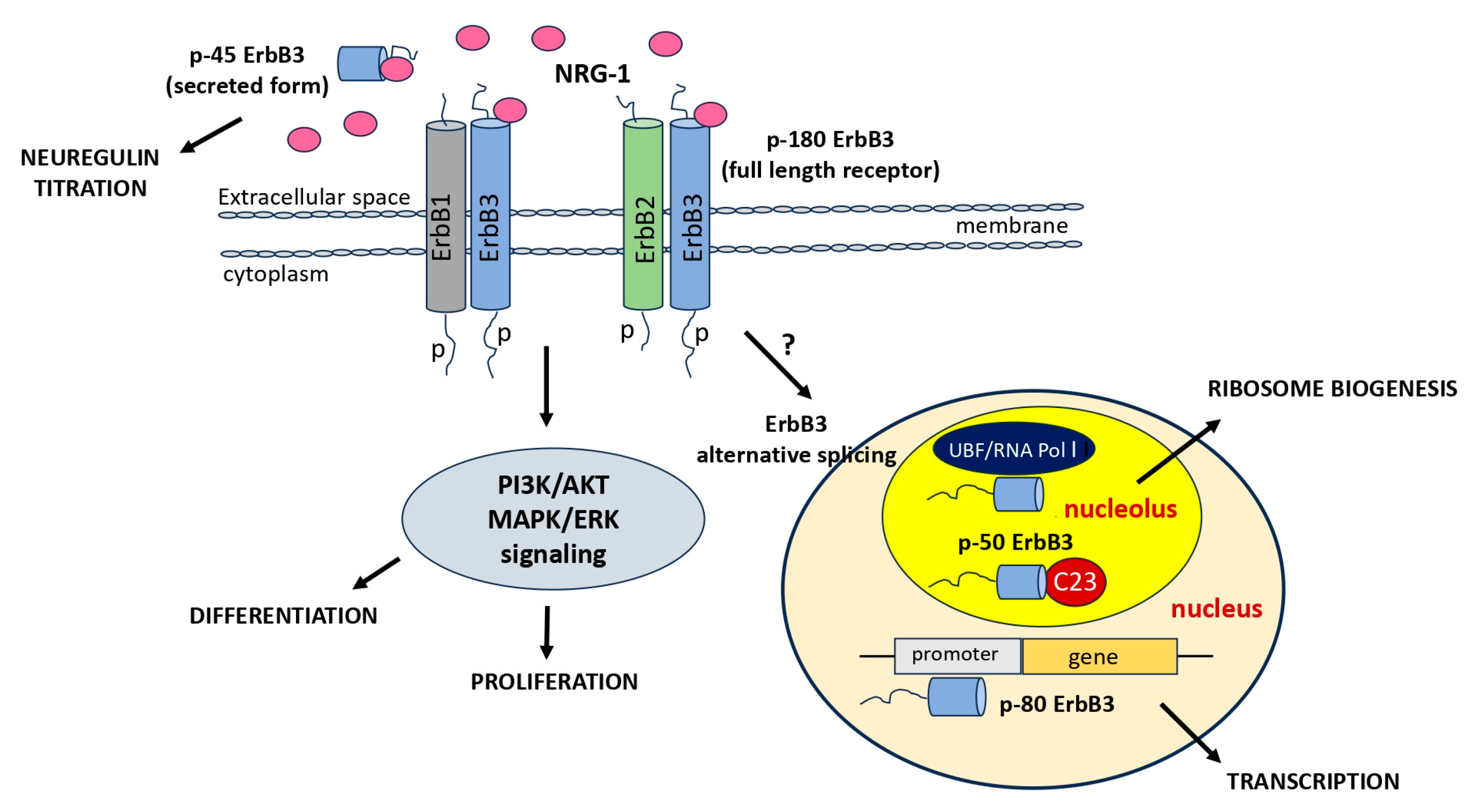
In addition to the specific features, there are additional layers of complexity presented by various ErbB isoforms and protein variants, each with unique cellular localizations. It is worth noting that all members of the ErbB family have been identified within the nuclei of various cell types, including both full-length receptors and alternative nuclear variants [85]. Nuclear variants of ErbB3 are generated through alternative transcription initiation and encode a portion of the cytoplasmic domain of the full-length ErbB3. ErbB3 demonstrates distinct bands of various molecular weights (120 kDa, 80 kDa, and 50 kDa) in addition to the full- length receptor (180 kDa) [85]. Specifically, one of these variants has been identified in Schwann cells, and its expression is dependent on neuregulin signaling [86]. Notably, the 180 kDa full-length ErbB3 band is exclusively detected in cytoplasmic extracts, while the 50 and 80 kDa band are exclusively present in nuclear extracts [86]. The nuclear ErbB3 binds to the chromatin of promoters associated with genes expressed in Schwann cells. Among these genes, ezrin has been identified as a component of Schwann cell microvilli and plays a role in the formation of the nodes of Ranvier [86,87,88,89,90]. The expression of ErbB3 can be regulated by a variety of stimuli, including growth factors, hormones, and environmental stress. It undergoes alternative splicing, to generate multiple isoforms of the protein. These isoforms have different functions and may play distinct roles in various cellular processes. ErbB3 is frequently overexpressed in various types of cancer, including breast, ovarian, and glioblastoma. This overexpression is thought to contribute to tumor growth and progression by activating signaling pathways that promote cell proliferation, survival, and invasion. The genomic analysis of the partial structure of c-ErbB3 (cloned form of ErbB3) shows that ErbB3 gene encodes the transmembrane receptor mentioned above and a secreted isoform by alternative splicing mechanism [15,91]. It is a 1.4 kb transcript that represents the secreted receptor. Its N-terminal is glycosylated (58 kDa) and is unable to bind NRG1b [92,93]. Four other cDNAs have been isolated (1.6, 1.7, 2.1, 2.3 kb) in ovarian cancer cell lines. Those four are synthesized from a non-codification region and use alternative polyadenylation signals [93].
Similar isoforms of ErbB3 have been identified in human non pathological placenta. The expression of these alternative isoforms seems to be tissue-specific and could play a specific role in cellular growth or in cell transformation [93]. In fact, some of these, such as p45-sErbB3 and especially p85-sErbB3 (secreted form of the human ErbB3 receptor), can bind NRG with high affinity, thereby inhibiting heterodimerization of ErbB2-ErbB3 and phosphorylation, and then cause a lack of pathway activation (PI3K, MAPK, Akt) [93]. One of these protein’s isoforms includes a truncated one known as ErbB3 50 kDa that lacks most of the extracellular domain. A recent study suggests that it may play a role in nucleolar function and ribosome biogenesis [94]. Here it could modulate ribosome biogenesis and cell proliferation, suggesting that it may contribute to tumor growth [94]. This variant of ErbB3 was found to interact with nucleolar proteins such as nucleolin (C23) and upstream binding factor (UBF), which are both involved in ribosome biogenesis [94]. Further research is needed to fully understand the functions of this short variant of ErbB3 in the nucleolus and its potential as a therapeutic target. Understanding the structure and function of this short variant of ErbB3 is therefore crucial for comprehending its role in downstream signaling pathways and for developing targeted therapies capable of inhibiting its activity (Figure 4).
In conclusion, all members of the EGFR family have been detected within the nuclei of various cell types, occurring either in the full-length isoform or as alternative variants. In the case of ErbB3, these variants are generated through alternative transcription initiation or splicing processes. These diverse isoforms are likely to serve distinct roles in cellular processes. For instance, one of these variants has been observed in Schwann cells, with its expression dependent on NRG signaling. Furthermore, the shorter isoforms exhibit a unique affinity for the nucleus, where they can interact with the chromatin promoters of genes such as ezrin, a protein involved in the formation of nodes of Ranvier. Other studies suggest their potential roles in cell growth, transformation, or as transcription factors. Within the nucleolus, these isoforms may influence ribosome biogenesis.
8. ErbB3 in Glioblastoma
When considering various diseases associated with abnormal EGFRs/NRG signaling, one particularly significant example is glioma. Glioma is a prevalent primary cancer occurring in the brain and central nervous system (CNS) [95,96]. Constituting nearly 80% of primary malignant brain tumors, gliomas originate from brain tissue and glial cells. The current World Health Organization (WHO) classification distinguishes four main glioma groups: adult diffuse glioma, pediatric diffuse low-grade glioma (LGG), pediatric diffuse high-grade glioma (HGG), and circumscribed astrocytic glioma. Based on proliferation extent, mitotic index, and necrotic levels observed histologically, gliomas are categorized into four grades: Grades I and II as LGGs, potentially manageable through resection based on location; Grades III and IV as HGGs, associated with an unfavorable prognosis and high malignancy [97,98]. Following the discovery of neurogenesis in the adult brain, neural stem cells have gained attention as potential glioma sources. Adult neurogenesis supports neuron and glial cell replacement, aiding cellular replenishment, remodeling, and injury response [96,99]. It is acknowledged that adult gliomas emerge from the neuroglial lineage postnatally, supported by evidence from glioma histology, molecular patterns, and murine glioma models targeting the neuroglial lineage. The mature mammalian brain houses two recognized neurogenic niches: the subventricular zone (SVZ) and the subgranular zone [96]. While an exact cause remains unclear for most malignant gliomas, exposure to ionizing radiation is an established risk factor [100]. Furthermore, suggestive data imply a link between immunologic factors and gliomas. Individuals with atopy show reduced glioma risk [101], and glioblastoma (Grade IV, the highest in the WHO brain tumor classification) patients with elevated IgE levels have extended survival [102]. Common molecular changes in glioblastoma include mutations in genes governing receptor tyrosine kinase (RTK)/rat sarcoma (RAS)/phosphoinositide 3-kinase (PI3K), p53, and retinoblastoma protein (RB) signaling [103]. Additionally, gene polymorphisms influencing detoxification, DNA repair, and cell-cycle regulation are implicated in glioma development [104]. Roughly 5% of malignant glioma patients have a familial glioma history, some linked to rare genetic syndromes like neurofibromatosis types 1 and 2, Li–Fraumeni syndrome, and Turcot’s syndrome [101,105]. Distinct mutations in isocitrate dehydrogenase (IDH1 or IDH2) genes seem to define glioma tumors, linked to younger age and better outcomes. Mutations in IDH1, a key Krebs cycle enzyme, occur in 12% of glioblastomas and predict a more favorable prognosis than IDH wildtype [101,106,107]. IDH2, a related enzyme in mitochondria, also correlates with longer survival [107]. Studies show IDH1 and IDH2 mutations are common in most low-grade gliomas. The underlying theory attributes the improved prognosis to reduced NADPH production, rendering mutated tumor cells more susceptible to reactive oxygen species (ROS) damage, compared to the resistance observed in non-mutated tumors, leading to a more aggressive behavior [107]. Glioblastoma (GBM) is the most common malignant brain tumor in adults, and among the most lethal of all cancers with a median survival of only 13–15 months [108,109]. GBMs are classified into primary tumors that arise de novo (90% of all cases) and secondary tumors that progress from low-grade gliomas [110]. According to differences in expression patterns, GBMs are divided into four subtypes: classical, proneural, mesenchymal, and neural, with intratumoral heterogeneity observed in single-cell RNA-sequencing studies [111]. In GBMs, the overexpression of ErbBs is often sustained by gene amplification, significantly contributes to tumorigenesis, and provides potential therapeutic targets [112]. ErbB1 (EGFR) amplification is enriched in the classical subtype [104]. Amplification of the ErbB1 (EGFR) gene occurs in 57.4% of primary GBM patients compared to 8% of secondary GBM patients and is associated with high levels of ErbB1 (EGFR) protein [113]. GBMs exhibit a distinct set of ErbB1 (EGFR) deletions and point mutations. These GBM-associated ErbB1 (EGFR) deletions include EGFRvI (N-terminal deletion), vII (deletion of exons 14–15), vIII (deletion of exons 2–7), vIV (deletion of exons 25–27), and vV (deletion of exons 25–28). Among these, vII and vIII are known to be oncogenic. Additionally, point mutations in the extracellular region of ErbB1 (EGFR), such as R108K, A289V/D/T, G598D, and other extracellular domain mutations, have been detected in 24% of GBM samples. These point mutations sustain ErbB1 (EGFR) in an active conformation. Notably, among the ErbB1 (EGFR) mutants found in GBM, EGFRvIII is the most prevalent. EGFRvIII lacks amino acids 6–273, with the deletion of these 268 amino acids leading to a junction site with a new glycine residue positioned between amino acids 5 and 274. Analysis of other members of the EGFR family has highlighted the significance of ErbB4 as the second most prevalent member within GBMs. ErbB4 is notably expressed in neuron-like elements and, interestingly, in some GBM cell lines as well. Furthermore, the ErbB2 receptor is detectable in the tissue of primary patients affected by glioblastoma, with a high ErbB2 expression rate of 21.5% among samples, while moderate and absent ErbB2 expression is observed in 19.6% and 58.9% of cases, respectively [114,115,116,117] Recent discoveries have also shed light on the involvement of ErbB3, yet another member of the EGFR family, in the progression of GBM [118]. Using a model that utilizes patient-derived stem-like cells, researchers identified a specific subgroup of human GBMs, characterized by excessive ErbB3 expression and its accumulation in the cell nucleus.
Finally, glioma, brain, and central nervous system cancers are significant examples of diseases associated with abnormal EGFRs/NRG signaling. Gliomas constitute nearly 80% of primary malignant brain tumors. They are classified into four groups based on the World Health Organization (WHO) classification and categorized into four grades based on histological features, with Grades III and IV (HGGs) having an unfavorable prognosis. In GBM (Grade IV of glioma), the overexpression of ErbBs, often due to gene amplification, significantly contributes to tumorigenesis and presents potential therapeutic targets. EGFR amplification is particularly enriched in the classical subtype and occurs in a majority of primary GBM patients compared to secondary GBM patients. GBM also exhibits distinct EGFR deletions and point mutations. Among these mutations, EGFRvIII is the most prevalent. Recent research has highlighted the role of ErbB3, a member of the EGFR family, in GBM progression. A specific subgroup of GBMs characterized by excessive ErbB3 expression and its accumulation in the cell nucleus has been identified.
9. ErbB3 and miRNA in Glioblastoma
MicroRNAs are small non-coding RNAs that regulate gene expression at the post-transcriptional level by binding to the 3′ untranslated region (UTR) of target mRNAs. It has been proven that miRNAs play a significant role in various biological processes, including the cell cycle, apoptosis, proliferation, and differentiation [119,120]. Aberrant miRNA expression has been implicated in various diseases, including cancer [121,122]. There is growing evidence that miRNAs can regulate ErbB3 expression and function. Several miRNAs have been identified as regulators of ErbB3 expression in cancer cells, including miR-125a, miR-125b, miR-205, miR-708, and miR-1224 [118,123]. These miRNAs have been shown to inhibit ErbB3 expression by binding to its 3′ UTR and reducing mRNA stability and/or translation. Overall, the regulation of ErbB3 expression and function by miRNAs is a complex and dynamic process that is likely to play a significant role in cancer pathogenesis, including glioma cancer. Notably, miRNAs have emerged as significant players in cancer and their association with gliomas has gained increasing attention [118,123,124,125]. In gliomas, miRNAs have also been found to regulate ErbB3 expression and function. For example, miR-34a has been shown to target ErbB3 and inhibit glioma cell proliferation and invasion [126]. In addition, miR-125b has been found to regulate ErbB3-mediated PI3K-AKT signaling in glioblastoma cells, and its downregulation is associated with glioma progression and poor prognosis [127]. Interestingly, ErbB3 has been found to regulate miRNA expression in turn. Overall, the regulation of ErbB3 expression and function by miRNAs is a complex and dynamic process that is likely to play a significant role in cancer pathogenesis, including gliomas. Further research is needed to fully understand the nature and significance of these interactions and to develop effective miRNA-based therapies for glioma patients. The overexpression of ErbB3 can be driven by hereditary promoter methylation or post-transcriptional silencing of miR-205, thereby sustaining the malignant phenotype in glioblastoma [118]. Furthermore, ErbB3 is recognized as a driver of drug resistance against EGFR inhibitors in cancer through various mechanisms [128,129].
In sum, these studies offer important avenues for exploration to gain a comprehensive understanding of the pathogenesis and mechanisms of drug resistance. They also present a challenge in identifying patients with specific molecular characteristics and offer prospects for new therapeutic strategies that are yet to be developed. In gliomas, miR-34a inhibits glioma cell growth and invasion by targeting ErbB3. MiR-125b regulates ErbB3-mediated signaling and its downregulation is linked to glioma progression. Interestingly, ErbB3 can reciprocally affect miRNA expression. Overall, understanding these interactions is crucial for cancer pathogenesis, particularly in gliomas, and for developing miRNA-based therapies. ErbB3 overexpression can result from hereditary promoter methylation or miR-205 silencing, sustaining the malignant phenotype. Further research is needed to unravel the significance of these interactions.
10. ErbB3 Receptor and Nucleolus in Glioblastoma
The nucleolus is a vital subcompartment that plays a central role in maintaining cell homeostasis and facilitating ribosome biogenesis. Mounting evidence suggests its function as a stress sensor, with changes in its morphology linked to aggressive cancers [130]. Tumor suppressors, such as retinoblastoma (pRB) and p53, exert negative regulation on RNA polymerase I and disrupt the assembly of transcriptional machinery at the rDNA promoter [131]. The nucleolus, a subnuclear compartment responsible for ribosome biogenesis, not only plays a critical role in cellular homeostasis. Proper coordination of ribosome production with cell-cycle progression relies on cell cycle checkpoints [132,133]. Additionally, several tumor suppressor agents, including p53, PTEN, pRB, and ARF (alternative reading frame), localize within the nucleolus in cancer, including glioblastoma [134]. In recent studies, the presence of ErbB family members has been detected within the nucleus and nucleolus of cells [85,95,135,136,137].
This intriguing localization of ErbBs holds the promise of uncovering fresh insights into their functions in pathological contexts, potentially offering valuable markers and targets for innovative interventions. In physiological conditions, all members of the EGFR family, along with their ligands, have been observed in the nucleoplasm or nucleolus of cells. However, this localization persists even in cancer cells [135]. The nucleolar presence of ErbB3 has been demonstrated in cell lines and in primary glioblastoma cells obtained from biopsies of affected patients [94]. In high-grade glioblastoma, where the concentration of NRG1 increases, ErbB3 and the nucleolar protein nucleolin (C23) can be released from the nucleolus and translocate to the cytoplasm, thereby positively influencing cell proliferation. The binding of actinomycin D disrupts this mechanism, causing the nuclear accumulation of ErbB3 and C23. This disruption leads to a decrease in ribosome biogenesis and subsequently inhibits cell proliferation. It is noteworthy that in glioblastoma, the overexpression of C23 is linked to its cytoplasmic localization, and this localization increases proportionally with the tumor grade [94,95]. Thus, the interaction between ErbB3 and C23 might play a role in tumor growth. Earlier studies hinted at C23’s direct influence on ErbB receptors [138]. Recent research has revealed the presence of additional ErbB family members within the nucleus and nucleolus of cancer cells. Specifically, ErbB1 has been observed to migrate into the nucleus in response to stress, during cell proliferation, or under tumor conditions. In contrast, the unique localization pattern of ErbB4 stems from proteolytic cleavage, a mechanism commonly shared among receptors in this family [137]. Regardless of the scenario, it is important to highlight that these receptors possess unique nuclear and nucleolar localization motifs, which strongly suggests their involvement in authentic cellular functions within these compartments. ErbB3, even more intriguingly, demonstrates localization within the nucleolus across various cell types, including those afflicted by cancers like breast cancer and cervical cancer, and in both cell lines and primary cells of glioblastoma [94]. This variant of ErbB3 exhibits a molecular weight of 50 kDa and has been found to engage with nucleolar proteins, notably C23 and UBF, both of which play pivotal roles in ribosome biogenesis [94]. Intriguingly, silencing ErbB3 has been shown to block the cell cycle and impact cell proliferation. Recently, mitogenic growth factors, which are typically cell-surface or secreted proteins, have also been discovered in the nucleus and potentially influence transcription. Neuregulin1 is found in the nuclei of human breast cancer cells. Furthermore, an isoform of this ligand, neuregulin1-β3, is found in the nucleoli of the cells [139].
In conclusion, despite these compelling findings, the precise functions of ErbB3 within the nuclear domain remain incompletely understood, necessitating further in-depth investigations. ErbB receptors have been detected in both the nucleoplasm and nucleolus of both normal and cancer cells, indicating their potential roles in diverse cellular contexts. This observation is reinforced by the presence of nuclear and nucleolar localization sequences within these receptors. Notably, in high-grade glioblastoma (GBM), a variant of ErbB3 with a molecular weight of 50 kDa has been identified within the nucleolus, potentially influencing ribosome biogenesis and cell proliferation.
11. ErbB3 Signaling and Target Therapies
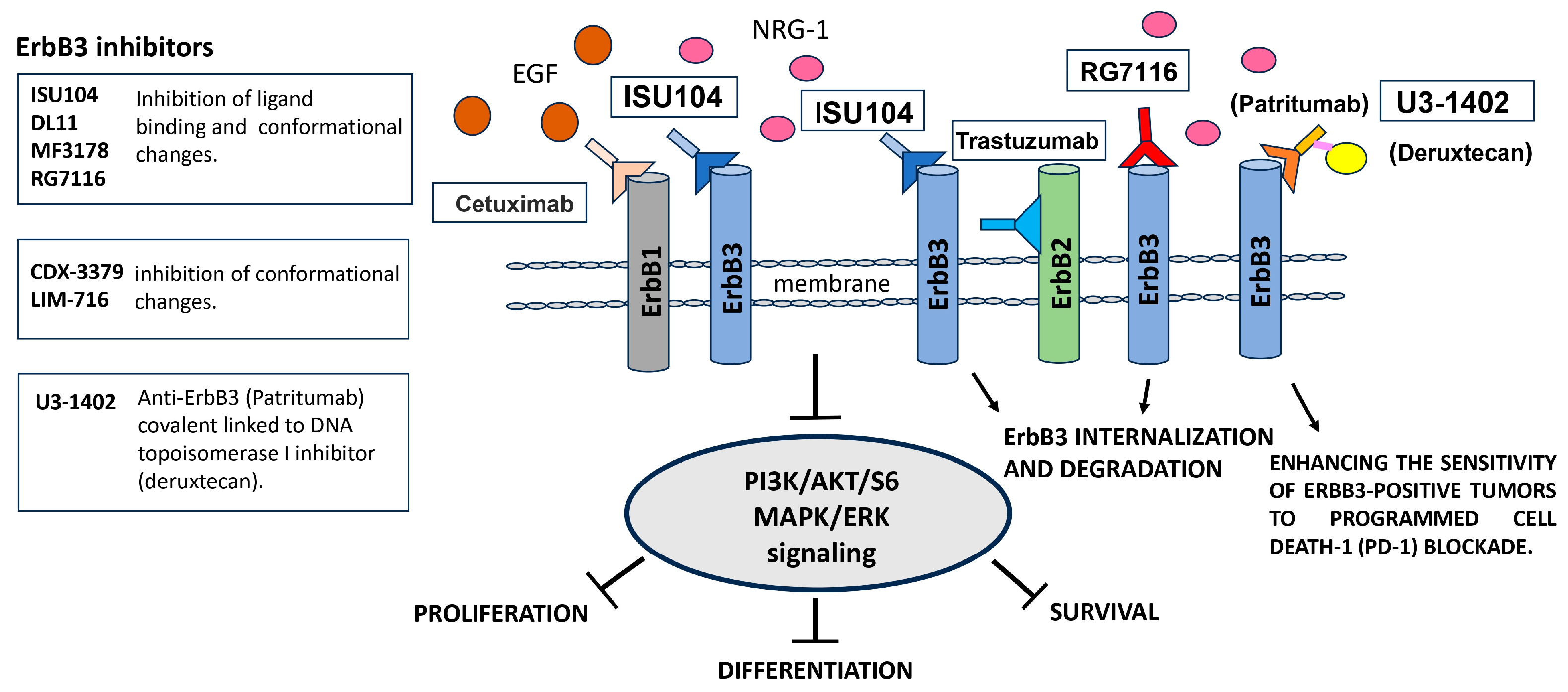
Aberrant signaling by ErbB family members plays a pivotal role in initiating tumors and facilitating the evasion of anti-tumor immune responses across a spectrum of malignancies. Studies have demonstrated the strong therapeutic potential of drugs designed to target these signaling pathways. Nevertheless, a recurring challenge arises as patients frequently develop acquired resistance to these treatments over time [140]. The current therapeutic approach for glioblastoma (GBM), a WHO grade IV glioma, still relies on a standard protocol established in 2005. This protocol involves surgery which is often limited due to the infiltrative nature of the tumor, followed by radiotherapy and chemotherapy with temozolomide [141,142,143,144]. Glioblastoma commonly exhibits genetic alterations, including changes in the EGFR gene, such as amplification and EGFRvIII mutations [112,113]. Amplification of the EGFR gene leads to the activation of downstream signaling pathways that play a significant role in driving tumorigenesis. Additionally, ErbB3 is frequently overexpressed in glioma, particularly in glioblastoma [118]. Notably, the expression of ErbB3 is correlated with a bleak prognosis in glioma patients [143]. In fact, ErbB3 signaling in gliomas is believed to trigger numerous downstream signaling pathways that enhance tumor growth, invasion, and resistance to therapy [143]. Additionally, studies have demonstrated that ErbB3 expression in glioblastoma cell lines can occur in two forms: the full-length receptor and a shorter variant (50 kDa), which is associated with the presence of NRG [94]. Moreover, this variant is also observed in the nucleolus of primary cultures of patients affected by glioblastoma. It is thought to translocate to the nucleolus, where its presence is linked to altered expression of pre-rRNA 47S. Furthermore, silencing this variant of ErbB3 leads to cancer cell death and cell cycle arrest. Given its role in glioma progression and resistance to therapy, ErbB3 has emerged as a promising therapeutic target in glioma. Several drugs that target ErbB3 are currently in development, including monoclonal antibodies and small molecule inhibitors with inhibitor effect on different cancers [144,145]. A monoclonal antibody called lumretuzumab (also known as RG7116, RO5479599, and GEhuMabHER3) is currently under clinical development. Two clinical trials hinge on it: NCT01918254 and NCT01482377, in combination with other drugs it targets HER2+, HER2+ metastatic breast cancer or HER3+ solid tumor of epithelial origin (completed phase I—clinicaltrial.gov) [146]. Lumretuzumab targets the ErbB3 receptor and belongs to the IgG1 class of humanized glycoengineered antibodies. It specifically binds to subdomain I of the extracellular domain of ErbB3 [145]. When used alone or in combination with other anti-ErbB therapies, this antibody has demonstrated significant inhibition of tumor growth [147]. In addition, lumretuzumab demonstrates a great affinity for human FcγRIIIa receptors on immune effector cells. Another promising antibody currently in early clinical development is ISU104, a human anti-ErbB3 antibody. ISU104 specifically targets subdomain III of ErbB3. Its effect was studied in vitro and in vivo. In the first case, the experiment was performed in BxPC3 and SKBR3 cell lines, which are models for NRG1-dependent and -independent cell proliferation, respectively. In vivo studies have shown that ISU104 achieves more than 70% inhibition of tumor growth in various cancer models. In vivo efficacy of ISU104 was investigated using different cancer cell–derived xenograft models, including head and neck squamous cell carcinomas (HNSCC), lung, pancreatic, colon, skin, and breast cancers, which are known to be related to ErbB3 oncogenic signaling [148]. It is being evaluated in a Phase I, open-label, dose-finding study to assess its safety, tolerability, and pharmacokinetics in patients with advanced solid tumors. It functions by downregulating ErbB3 and inhibiting NRG binding, blocking dimerization with other ErbB partners, and inactivating downstream signaling. Antibody-drug conjugates (ADCs) represent a novel class of antitumoral agents that combine the selectivity of monoclonal antibodies (mAbs) with the cell-killing properties of cytotoxic drugs. These drugs are attached to the mAbs through a linker. One example of an ADC is U3-1402, also known as patritumab deruxtecan or ErbB3-DXd. U3-1402 is composed of patritumab, which is covalently conjugated to a drug-linker containing deruxtecan, a topoisomerase I inhibitor. A second example is RG7116 bound potently and selectively to domain 1 of human ErbB3. Heregulin binding was prevented by RG7116 at concentrations of more than 1 nmol/L as was nearly complete inhibition of ErbB3 heterodimerization and phosphorylation, thereby preventing downstream AKT phosphorylation. In vivo, RG7116 treatment inhibited xenograft tumor growth up to 90% relative to controls in a manner accompanied by downregulation of cell surface ErbB3. RG7116 efficacy was further enhanced in combination with anti-EGFR (RG7160) or anti-ErbB2 (pertuzumab) mAbs. Furthermore, the ADCC potency of RG7116 was enhanced compared with the non-glycoengineered parental antibody, both in vitro and in orthotopic tumor xenograft models, where an increased median survival was documented [149]. U3-1402 efficiently internalized, induced ErbB3 degradation and showed growth inhibition activity in HER3+ breast, gastric, and colorectal cancers. Recent studies have shown that U3-1402 can sensitize ErbB3+ tumors to programmed cell death-1 (PD-1) blockade [150,151,152]. Data are available under clinical trial NCT03260491 phase I for metastatic or unresectable NSCLC and NCT02980341 phase II and III for breast cancer. This combination therapy works by blocking the activation of ErbB3 and downstream signaling pathways, effectively inhibiting cancer cell growth and promoting cell death (Figure 5).
In sum, the conventional therapeutic approach for glioblastoma, established in 2005, has inherent limitations. Multiple monoclonal antibodies targeting ErbB3 have been developed and demonstrated to effectively block ErBR3’s downstream signaling. This blocking action is achieved by preventing ligand binding, inhibiting heterodimerization, maintaining ErbB3 in an inactive state, and potentially inducing ErbR3 internalization.
12. Discussion
There is mounting evidence highlighting the crucial role of ErbB receptor family members in the development of the nervous system. Furthermore, they exert significant influence on the neoplastic transformation and progression of various tumors, including brain tumors like glioblastoma. ErbB3 is commonly observed to be overexpressed in cancer cells. Despite its functional deficiency in the tyrosine kinase activity, it can activate crucial signaling pathways that significantly contribute to various mechanisms driving cancer progression. Given its role in cancer progression and resistance to therapy, ErbB3 has emerged as a promising target for cancer therapy. Several drugs that target ErbB3 are currently in clinical development, including small molecule inhibitors and monoclonal antibodies such us ISU104 and U3-1402. U3-1402 is a potential anti-ErbB3 under development that could be safely administered and has demonstrated promising antitumor efficacy in treated ErbB3-expressing metastatic breast cancer. These drugs work by blocking the activation of ErbB3 and downstream signaling pathways, thereby inhibiting cancer cell growth. Additionally, ErbB3 is recognized as a key driver of drug resistance in various cancers, including glioblastoma [140,141,142,143]. Its activation becomes particularly pronounced in response to targeted therapies aimed at inhibiting other members of the EGFR family, such as ErbB2. There are several mechanisms by which cancer cells can develop resistance to drugs that target ErbB3, and researchers have been developing strategies that target alternative signaling pathways. One approach involves the development of drugs that specifically target ErbB3-independent signaling complexes. These drugs may disrupt the formation of homodimers or heterodimers between EGFR family members or inhibit downstream signaling pathways that are activated independently of ErbB2. In addition, strategies such as the use of immunotherapy or combination therapies that target multiple cancer cell pathways simultaneously are being explored to overcome ErbB3 drug resistance.
13. Conclusions
The signaling of NRG-ErbB3 and its impact on the nervous system has attracted substantial attention, acknowledging the profound implications it possesses. Delving into the intricate mechanisms and outcomes of NRG-ErbB signaling within the context of the nervous system holds potential for advancing our comprehension of neural development, function, and pathologies, including brain tumors.
14. Future Directions
The direction of future research on the NRG-ErbBs signaling pathway in the nervous system holds promise for advancing our understanding of neural development and gaining insights into brain cancers like glioblastoma. This knowledge paves the way for the development of targeted therapies that can improve the treatment of neurological disorders and cancer. In this context, the targeting of ErbB3 is an active area of research for glioblastoma treatment. Various drugs, including monoclonal antibodies and small molecule inhibitors, are currently being developed to specifically inhibit ErbB3 and its signaling pathways. Furthermore, the combination of ErbB3 targeting with other treatment modalities such as chemotherapy or radiation therapy will enhance treatment effectiveness and improve patient outcomes.
Author Contributions
D.P. conceived and designed the manuscript, created the figures, and wrote the manuscript. M.T. designed and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Academic Funding Sapienza 2020 RM11916B410542D4 (to D.P.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burgess, A.W. Regulation of Signaling from the Epidermal Growth Factor Family. J. Phys. Chem. B 2022, 126, 7475–7485. [Google Scholar] [CrossRef] [PubMed]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; De Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Ferguson, K.M.; Darling, P.J.; Mohan, M.J.; Macatee, T.L.; Lemmon, M.A. Extracellular domains drive homo- but not hetero-dimerization of erbB receptors. EMBO J. 2000, 19, 4632–4643. [Google Scholar] [CrossRef]
- Jones, J.T.; Akita, R.W.; Sliwkowski, M.X. Binding specificities and affinities of egf domains for ErbB receptors. FEBS Lett. 1999, 447, 227–231. [Google Scholar] [CrossRef]
- Landgraf, R.; Eisenberg, D. Heregulin reverses the oligomerization of HER3. Biochemistry 2000, 39, 8503–8511. [Google Scholar] [CrossRef]
- Breuleux, M. Role of heregulin in human cancer. Cell. Mol. Life Sci. 2007, 64, 2358–2377. [Google Scholar] [CrossRef]
- Willem, M. Proteolytic processing of Neuregulin-1. Brain Res. Bull. 2016, 126 Pt 2, 178–182. [Google Scholar] [CrossRef]
- Hu, C.; Leche, C.A., 2nd; Kiyatkin, A.; Yu, Z.; Stayrook, S.E.; Ferguson, K.M.; Lemmon, M.A. Glioblastoma mutations alter EGFR dimer structure to prevent ligand bias. Nature 2022, 602, 518–522. [Google Scholar] [CrossRef]
- Black, L.E.; Longo, J.F.; Carroll, S.L. Mechanisms of Receptor Tyrosine-Protein Kinase ErbB-3 (ERBB3) Action in Human Neoplasia. Am. J. Pathol. 2019, 189, 1898–1912. [Google Scholar] [CrossRef]
- Gimple, R.C.; Bhargava, S.; Dixit, D.; Rich, J.N. Glioblastoma stem cells: Lessons from the tumor hierarchy in a lethal cancer. Genes Dev. 2019, 33, 591–609. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Li, D.; Yang, T.; Liu, F.; Kong, J.; Zhou, Y. PTPN1 promotes the progression of glioma by activating the MAPK/ERK and PI3K/AKT pathways and is associated with poor patient survival. Oncol. Rep. 2019, 42, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.W.; Cho, H.S.; Eigenbrot, C.; Ferguson, K.M.; Garrett, T.P.; Leahy, D.J.; Lemmon, M.A.; Sliwkowski, M.X.; Ward, C.W.; Yokoyama, S. An open-and-shut case? Recent insights into the activation of EGF/ErbB receptors. Mol. Cell 2003, 12, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Kraus, M.H.; Issing, W.; Miki, T.; Popescu, N.C.; Aaronson, S.A. Isolation and characterization of ERBB3, a third member of the ERBB/epidermal growth factor receptor family: Evidence for overexpression in a subset of human mammary tumors. Proc. Natl. Acad. Sci. USA 1989, 86, 9193–9197. [Google Scholar] [CrossRef] [PubMed]
- Bose, R.; Zhang, X. The ErbB kinase domain: Structural perspectives into kinase activation and inhibition. Exp. Cell Res. 2009, 315, 649–658. [Google Scholar] [CrossRef]
- Yokoe, S.; Takahashi, M.; Asahi, M.; Lee, S.H.; Li, W.; Osumi, D.; Miyoshi, E.; Taniguchi, N. The Asn418-linked-N-glycan of ErbB3 plays a crucial role in preventing spontaneous heterodimerization and tumor promotion. Cancer Res. 2007, 67, 1935–1942. [Google Scholar] [CrossRef]
- Sotoyama, H.; Namba, H.; Tohmi, M.; Nawa, H. Schizophrenia Animal Modeling with Epidermal Growth Factor and Its Homologs: Their Connections to the Inflammatory Pathway and the Dopamine System. Biomolecules 2023, 13, 372. [Google Scholar] [CrossRef]
- Lemmetyinen, T.T.; Viitala, E.W.; Wartiovaara, L.; Kaprio, T.; Hagström, J.; Haglund, C.; Katajisto, P.; Wang, T.C.; Domènech-Moreno, E.; Ollila, S. Fibroblast derived EGF ligand neuregulin 1 induces fetal-like reprogramming of the intestinal epithelium without supporting tumorigenic growth. Dis. Models Mech. 2023, 16, dmm049692. [Google Scholar] [CrossRef]
- Ting, A.K.; Chen, Y.; Wen, L.; Yin, D.M.; Shen, C.; Tao, Y.; Liu, X.; Xiong, W.C.; Mei, L. Neuregulin 1 Promotes Excitatory Synapse Development and Function in GABAergic Interneurons. J. Neurosci. 2011, 31, 15–25. [Google Scholar] [CrossRef]
- Müller, T.; Braud, S.; Jüttner, R.; Voigt, B.C.; Paulick, K.; Sheean, M.E.; Klisch, C.; Gueneykaya, D.; Rathjen, F.G.; Geiger, J.R.; et al. Neuregulin 3 promotes excitatory synapse formation on hippocampal interneurons. EMBO J. 2018, 37, e98858. [Google Scholar] [CrossRef]
- Vullhorst, D.; Bloom, M.S.; Akella, N.; Buonanno, A. ER-PM Junctions on GABAergic Interneurons Are Organized by Neuregulin 2/VAP Interactions and Regulated by NMDA Receptors. Int. J. Mol. Sci. 2023, 24, 2908. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Nave, K.-A. Neuregulin-ERBB Signaling in the Nervous System and Neuropsychiatric Diseases. Neuron 2014, 83, 27–49. [Google Scholar] [CrossRef] [PubMed]
- Rio, C.; Rieff, H.I.; Qi, P.; Corfas, G. Neuregulin and erbB Receptors Play a Critical Role in Neuronal Migration. Neuron 1997, 19, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Namba, H.; Zheng, Y.; Nawa, H. In situ hybridization reveals developmental regulation of ErbB1-4 mRNA expression in mouse midbrain: Implication of ErbB receptors for dopaminergic neurons. Neuroscience 2009, 161, 95–110. [Google Scholar] [CrossRef]
- Iwakura, Y.; Zheng, Y.; Sibilia, M.; Abe, Y.; Piao, Y.S.; Yokomaku, D.; Wang, R.; Ishizuka, Y.; Takei, N.; Nawa, H. qualitative and quantitative re-evaluation of epidermal growth factor-ErbB1 action on developin midbrain dopaminergig action in vivo and in vitro: Target-derived neurotrophic signaling. J. Neurochem. 2011, 118, 45–56. [Google Scholar] [CrossRef]
- Makinodan, M.; Rosen, K.M.; Ito, S.; Corfas, G. A critical period for social experience-dependent oligodendrocyte maturation and myelination. Science 2012, 337, 1357–1360. [Google Scholar] [CrossRef]
- Sathyamurthy, A.; Yin, D.M.; Barik, A.; Shen, C.; Bean, J.C.; Figueiredo, D.; She, J.X.; Xiong, W.C.; Mei, L. ERBB3-mediated regulation of Bergmann glia proliferation in cerebellar lamination. Development 2015, 142, 522–532. [Google Scholar] [CrossRef]
- Wang, H.; Liu, F.; Chen, W.; Sun, X.; Cui, W.; Dong, Z.; Zhao, K.; Zhang, H.; Li, H.; Xing, G.; et al. Genetic recovery of ErbB4 in adulthood partially restores brain functions in null mice. Proc. Natl. Acad. Sci. USA 2018, 115, 13105–13110. [Google Scholar] [CrossRef]
- Luo, B.; Liu, Z.; Lin, D.; Chen, W.; Ren, D.; Yu, Z.; Xiong, M.; Zhao, C.; Fei, E.; Li, B. ErbB4 promotes inhibitory synapse formation by cell adhesion, independent of its kinase activity. Transl. Psychiatry 2021, 11, 361. [Google Scholar] [CrossRef]
- Pitcher, J.-L.; Alexander, N.; Miranda, P.J.; Johns, T.G. ErbB4 in the brain: Focus on high grade glioma. Front. Oncol. 2022, 12, 983514. [Google Scholar] [CrossRef]
- Exposito-Alonso, D.; Osório, C.; Bernard, C.; Pascual-García, S.; Del Pino, I.; Marín, O.; Rico, B. Subcellular sorting of neuregulins controls the assembly of excitatory-inhibitory cortical circuits. eLife 2020, 9, e57000. [Google Scholar] [CrossRef] [PubMed]
- Riethmacher, D.; Sonnenberg-Riethmacher, E.; Brinkmann, V.; Yamaai, T.; Lewin, G.R.; Birchmeier, C. Severe neuropathies in mice with targeted mutations in the ErbB3 receptor. Nature 1997, 389, 725–730. [Google Scholar] [CrossRef]
- Hu, X.; Xiao, G.; He, L.; Zhu, Q.; Niu, X.; Li, H.; Xu, Q.; Wei, Z.; Huang, H.; Luan, Y.; et al. ErbB dysregulation impairs cognition via myelination-dependent and-independent oligodendropathy. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lyons, D.A.; Pogoda, H.M.; Voas, M.G.; Woods, I.G.; Diamond, B.; Nix, R.; Arana, N.; Jacobs, J.; Talbot, W.S. erbb3 and erbb2 are essential for schwann cell migration and myelination in zebrafish. Curr. Biol. 2005, 15, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Makinodan, M.; Yamauchi, T.; Tatsumi, K.; Okuda, H.; Takeda, T.; Kiuchi, K.; Sadamatsu, M.; Wanaka, A.; Kishimoto, T. Demyelination in the juvenile period, but not in adulthood, leads to long-lasting cognitive impairment and deficient social interaction in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 978–985. [Google Scholar] [CrossRef]
- Senger, K.; Yuan, W.; Sagolla, M.; Doerr, J.; Bolon, B.; Ziai, J.; Sun, K.H.; Warming, S.; Roose-Girma, M.; Zhang, N.; et al. Embryonic lethality, and defective mammary gland development of activator-function impaired conditional knock-in Erbb3V943R mice. Adv. Genet. 2020, 2, e10036. [Google Scholar] [CrossRef]
- Limon, J.; Turc-Carel, C.; Dal Cin, P.; Rao, U.; Sandberg, A.A. Recurrent chromosome translocation in liposarcoma. Cancer Genet. Cytogenet. 1986, 22, 93–94. [Google Scholar] [CrossRef]
- Mrozek, K.; Karakousis, C.P.; Bloomfield, C.D. Chromosome 12 breakpoints are cytogenetically different in benign and malignant lipogenic tumors: Localization of breakpoints in lipoma to 12q15 and in myxoid liposarcoma to 12.q13.1. Cancer Res. 1993, 53, 1670–1675. [Google Scholar]
- Takahashi, M.; Hasegawa, Y.; Ikeda, Y.; Wada, Y.; Tajiri, M.; Ariki, S.; Takamiya, R.; Nishitani, C.; Araki, M.; Yamaguchi, Y.; et al. Suppression of heregulin β signaling by the single N-glycan deletion mutant of soluble ErbB3 protein. J. Biol. Chem. 2013, 288, 32910–32921. [Google Scholar] [CrossRef]
- Sithandam, G.; Anderson, L.M. The ERBB3 receptor in cancer and cancer gene therapy. Cancer Gene Ther. 2008, 15, 413–448. [Google Scholar] [CrossRef]
- Skinner, A.; Hurst, H.C. Transcriptional regulation of the c-erbB-3 gene in human breast carcinoma cell lines. Oncogene 1993, 8, 3393–3401. [Google Scholar]
- Zhu, C.H.; Domman, F.E. Dominant negative interference of transcription factor AP-2 causus inhibition of ErbB-3 expression and suppresses malignant cell growth. Breast Cancer Res. Treat. 2001, 71, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Kani, K.; Park, E.; Landgraf, R. The extracellular domains of ErbB3 retain high ligand binding affinty at endosome pH and in the locked conformation. Biochemistry 2005, 44, 1842–15857. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Patel, D.; Ellis, N.; Brown, S.P.; Lewandowski, J.R.; Dixon, A.M. Modulation of Transmembrane Domain Interactions in Neu Receptor Tyrosine Kinase by Membrane Fluidity and Cholesterol. J. Membr. Biol. 2019, 252, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Mendrola, J.M.; Berger, M.B.; King, M.C.; Lemmon, M.A. The single transmembrane domains of ErbB receptors self-associate in cell membranes. J. Biol. Chem. 2002, 277, 4704–4712. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.A.; Staros, J.V. Evolutionary analysis of the ErbB receptor and ligand families. J. Mol. Evol. 2000, 50, 397–412. [Google Scholar] [CrossRef]
- Guy, P.M.; Platko, J.V.; Cantley, L.C.; Cerione, R.A.; Carraway, K.L. Insect cell-expressed p180erbB3 possesses an impaired tyrosine kinase activity. Proc. Natl. Acad. Sci. USA 1994, 91, 8132–8136. [Google Scholar] [CrossRef]
- Plowman, G.D.; Whitney, G.S.; Neubauer, M.G.; Green, J.M.; McDonald, V.L.; Todaro, G.J.; Shoyab, M. Molecular cloning and expression of an additional epidermal growth factor receptor-related gene. Proc. Natl. Acad. Sci. USA 1990, 87, 4905–4909. [Google Scholar] [CrossRef]
- Plowman, G.D.; Green, J.M.; Culouscou, J.M.; Carlton, G.W.; Rothwell, V.M.; Buckley, S. Heregulin induces tyrosine phosphorylation of HER4/p180erbB4. Nature 1993, 366, 473–475. [Google Scholar] [CrossRef]
- Prigent, S.A.; Gullick, W.J. Identification of c-erbB-3 binding sites for phosphaidylinositol 3′kinase and SHC using an EGF receptor/c-erbB3 chimera. EMBO J. 1994, 13, 2831–2841. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Hamburger, A.W. The use of the yeast two hybrid system to evaluate ErbB-3 interactions with SH2 domain containing proteins. Biochem. Biophys. Res. Commun. 1998, 251, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Waterman, H.; Alroy, I.; Strano, S.; Seger, R.; Yarden, Y. The C-terminus of the kinase-defective neuregulin receptor ErbB-3 confers mitogenic superiority and dictates endocytic routing. EMBO J. 1999, 18, 3348–3358. [Google Scholar] [CrossRef] [PubMed]
- Kataria, H.; Alizadeh, A.; Karimi-Abdolrezaee, S. Neuregulin-1/ErbB network: An emerging modulator of nervous system injury and repair. Prog. Neurobiol. 2019, 180, 101643. [Google Scholar] [CrossRef] [PubMed]
- Goldowitz, D.; Hamre, K. The cells and molecules that make a cerebellum. Trends Neurosci. 1998, 21, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Manni, E.; Petrosini, L. A century of cerebellar somatotopy: A debated representation. Nat. Rev. Neurosci. 2004, 5, 241–249. [Google Scholar] [CrossRef]
- Bignami, A.; Dahl, D. Differentiation of astrocytes in the cerebellar cortex and the pyramidal tracts of the newborn rat. An immunofluorescence study with antibodies to a protein specific to astrocytes. Brain Res. 1973, 49, 393–402. [Google Scholar] [CrossRef]
- Yamada, K.; Watanabe, M. Cytodifferentiation of Bergmann glia and its relationship with Purkinje cells. Anat. Sci. Int. 2002, 77, 94–108. [Google Scholar] [CrossRef]
- Ledonne, A.; Mercuri, N.B. On the Modulatory Roles of Neuregulins/ErbB Signaling on Synaptic Plasticity. Int. J. Mol. Sci. 2019, 21, 275. [Google Scholar] [CrossRef]
- Busfield, S.J.; Michnick, D.A.; Chickering, T.W.; Revett, T.L.; Ma, J.; Woolf, E.A.; Comrack, C.A.; Dussault, B.J.; Woolf, J.; Goodearl, A.D.; et al. Characterization of a neuregulin-related gene, Don-1, that is highly expressed in restricted regions of the cerebellum and hippocampus. Mol. Cell. Biol. 1997, 17, 4007–4014. [Google Scholar] [CrossRef]
- Carraway, K.L., III; Weber, J.L.; Unger, M.J.; Ledesma, J.; Yu, N.; Gassmann, M.; Lai, C. Neuregulin-2, a new ligand of ErbB3/ErbB4-receptor tyrosine kinases. Nature 1997, 387, 512–516. [Google Scholar] [CrossRef]
- Jüttner, R.; Rathjen, F.G. Molecular analysis of axonal target specificity and synapse formation. Cell. Mol. Life Sci. 2005, 62, 2811–2827. [Google Scholar] [CrossRef] [PubMed]
- Kinugasa, Y.; Ishiguro, H.; Tokita, Y.; Oohira, A.; Ohmoto, H.; Higashiyama, S. Neuroglycan C, a novel member of the neuregulin family. Biochem. Biophys. Res. Commun. 2004, 321, 1045–1049. [Google Scholar] [CrossRef] [PubMed]
- Fox, I.J.; Kornblum, H.I. Developmental Profiling of ErbB Receptors in the Murine Central Nervous System: Implications for Functional Interactions. J. Neurosci. Res. 2005, 79, 584–597. [Google Scholar] [CrossRef]
- Francoeur, J.R.; Richardson, P.M.; Dunn, R.J.; Carbonetto, S. Distribution of Erb-B2, Erb-B3, and Erb-B4 in the Developing Avian Nervous System. J. Neurosci. Res. 1995, 41, 836–845. [Google Scholar] [CrossRef]
- Thompson, M.; Lauderdale, S.; Webster, M.J.; Chong, V.Z.; McClintock, B.; Saunders, R.; Weickert, C.S. Widespread expression of ErbB2, ErbB3 and ErbB4 in non-human primate brain. Brain Res. 2007, 1139, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Grider, M.H.; Belcea, C.Q.; Covington, B.P.; Reddy, V.; Sharma, S. Neuroanatomy, Nodes of Ranvier; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Snaidero, N.; Möbius, W.; Czopka, T.; Hekking, L.H.; Mathisen, C.; Verkleij, D.; Goebbels, S.; Edgar, J.; Merkler, D.; Lyons, D.A.; et al. Myelin membrane wrapping of CNS axons by PI (3,4,5) P3-dependent polarized growth at the inner tongue. Cell 2014, 156, 277–290. [Google Scholar] [CrossRef]
- Emery, B. Transcriptional and post-transcriptional control of CNS myelination. Curr. Opin. Neurobiol. 2010, 20, 601–607. [Google Scholar] [CrossRef]
- Emery, B. Regulation of oligodendrocyte differentiation and myelination. Science 2010, 330, 779–782. [Google Scholar] [CrossRef]
- Nave, K.A. Myelination, and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef]
- Lubetzki, C.; Sol-Foulon, N.; Desmazières, A. Nodes of Ranvier during development and repair in the CNS. Nat. Rev. Neurol. 2020, 16, 426–439. [Google Scholar] [CrossRef]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Ronzano, R.; Roux, T.; Thetiot, M.; Aigrot, M.S.; Richard, L.; Lejeune, F.X.; Mazuir, E.; Vallat, J.M.; Lubetzki, C.; Desmazières, A. Microglia-neuron interaction at nodes of Ranvier depends on neuronal activity through potassium release and contributes to remyelination. Nat. Commun. 2021, 12, 5219. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.D.; Partlow, L.M. Preparation of pure neuronal and non-neuronal cultures from embryonic chick sympathetic ganglia: A new method based on both differential cell adhesiveness and the formation of homotypic neuronal aggregates. Brain Res. 1976, 114, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.L.; Williams, A.K.; Glaser, L.; Bunge, R.P. Studies of Schwann cell proliferation. II. Characterization of the stimulation and specificity of the response to a neurite membrane fraction. J. Cell Biol. 1980, 84, 753–766. [Google Scholar] [CrossRef]
- Wood, P.M.; Bunge, R.P. Evidence that sensory axons are mitogenic for Schwann cells. Nature 1975, 256, 662–664. [Google Scholar] [CrossRef]
- Lemke, G.E.; Brockes, J.P. Identification and purification of glial growth factor. J. Neurosci. 1984, 4, 75–83. [Google Scholar] [CrossRef]
- Meyer, D.; Birchmeier, C. Multiple essential functions of neuregulin in development. Nature 1995, 378, 386–390. [Google Scholar] [CrossRef]
- Kim, K.; Lee, D. ERBB3-dependent AKT and ERK pathways are essential for atrioventricular cushion development in mouse embryos. PLoS ONE 2021, 16, e0259426. [Google Scholar] [CrossRef]
- Wolpowitz, D.; Mason, T.B.; Dietrich, P.; Mendelsohn, M.; Talmage, D.A.; Role, L.W. Cysteine-rich domain isoforms of the neuregulin-1 gene are required for maintenance of peripheral synapses. Neuron 2000, 25, 79–91. [Google Scholar] [CrossRef]
- McCarthy, K.D.; Partlow, L.M. Neuronal stimulation of (3H) thymidine incorporation by primary cultures of highly purified non-neuronal cells. Brain Res. 1976, 114, 415–426. [Google Scholar] [CrossRef]
- Salzer, J.L.; Bunge, R.P. Studies of Schwann cell proliferation. I. An analysis in tissue culture of proliferation during development, Wallerian degeneration, and direct injury. J. Cell Biol. 1980, 84, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Honjo, Y.; Kniss, J.; Eisen, J.S. Neuregulin-mediated ErbB3 signaling is required for formation of zebrafish dorsal root ganglion neurons. Development 2008, 135, 2615–2625. [Google Scholar] [CrossRef] [PubMed]
- Garratt, A.N.; Voiculescu, O.; Topilko, P.; Charnay, P.; Birchmeier, C. A dual role of erbB2 in myelination and in expansion of the schwann cell precursor pool. J. Cell Biol. 2000, 148, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Akmentin, W.; Du, C.; Role, L.W.; Talmage, D.A. Axonal Type III Nrg1 Controls Glutamate Synapse Formation and GluA2 Trafficking in Hippocampal-Accumbens Connections. eNeuro 2017, 4, ENEURO.0232-16.2017. [Google Scholar] [CrossRef]
- Andrique, L.; Fauvin, D.; El Maassarani, M.; Colasson, H.; Vannier, B.; Séité, P. ErbB3 80 kDa, a nuclear variant of the ErbB3 receptor, binds to the Cyclin D1 promoter to activate cell proliferation but is negatively controlled by p14ARF. Cell. Signal. 2012, 24, 1074–1085. [Google Scholar] [CrossRef] [PubMed]
- Adilakshmi, T.; Ness-Myers, J.; Madrid-Aliste, C.; Fiser, A.; Tapinos, N. A nuclear variant of ErbB3 receptor tyrosine kinase regulates ezrin distribution and Schwann cell myelination. J. Neurosci. 2011, 31, 5106–5119. [Google Scholar] [CrossRef] [PubMed]
- Jathal, M.K.; Siddiqui, S.; Vasilatis, D.M.; Durbin Johnson, B.P.; Drake, C.; Mooso, B.A.; D’Abronzo, L.S.; Batra, N.; Mudryj, M.; Ghosh, P.M. Androgen receptor transcriptional activity is required for heregulin-1β-mediated nuclear localization of the HER3/ErbB3 receptor tyrosine kinase. J. Biol. Chem. 2023, 299, 104973. [Google Scholar] [CrossRef]
- Melendez-Vasquez, C.V.; Rios, J.C.; Zanazzi, G.; Lambert, S.; Bretscher, A.; Salzer, J.L. Nodes of Ranvier form in association with ezrin-radixin-moesin (ERM)-positive Schwann cell processes. Proc. Natl. Acad. Sci. USA 2001, 98, 1235–1240. [Google Scholar] [CrossRef]
- Rasband, M.N.; Peles, E. Mechanisms of node of Ranvier assembly. Nat. Rev. Neurosci. 2021, 22, 7–20. [Google Scholar] [CrossRef]
- Katoh, M.; Yazaki, Y.; Sugimura, T.; Terada, M. c-erbB3 gene encodes secreted as well transmembrane receptor tyrosine kinase. Biochem. Biophys. Res. Commun. 1993, 192, 1189–1197. [Google Scholar] [CrossRef]
- Srinivasan, R.; Leverton, K.E.; Sheldon, H.; Hurst, H.C.; Sarraf, C.; Gullick, W.J. Intracellular expression of the truncated extracellular domain of c-erbB3/HER3. Cell. Signal. 2001, 13, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Maihle, N.J. Isolation and characterization of four alternate c-erbB3 transcripts expressed in ovarian carcinoma-derived cell lines and normal human tissues. Oncogene 1998, 16, 3243–3252. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Akita, R.W.; Sliwkowski, M.X.; Maihle, N.J. A naturally occurring secreted human ErbB3 receptor isoform inhibits heregulin-stimulated activation of ErbB2, ErbB3 and ErbB4. Cancer Res. 2001, 61, 4467–4473. [Google Scholar]
- Tagliaferro, M.; Rosa, P.; Bellenchi, G.C.; Bastianelli, D.; Trotta, R.; Tito, C.; Fazi, F.; Calogero, A.; Ponti, D. Nucleolar localization of the ErbB3 receptor as a new target in glioblastoma. BMC Mol. Cell Biol. 2022, 23, 13. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, L.C.; Obernier, K.; Alvarez-Buylla, A. Adult neural stem cells bridge their niche. Cell Stem Cell 2012, 10, 698–708. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Bauchet, L.; Davis, F.G.; Deltour, I.; Fisher, J.L.; Langer, C.E.; Pekmezci, M.; Schwartzbaum, J.A.; Turner, M.C.; Walsh, K.M.; et al. The epidemiology of glioma in adults: A “state of the science” review. Neuro-Oncology 2014, 16, 896–913. [Google Scholar] [CrossRef]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109, Erratum in Acta Neuropathol. 2007, 114, 547. [Google Scholar] [CrossRef]
- Bai, J.; Varghese, J.; Jain, R. Adult Glioma WHO Classification Update, Genomics, and Imaging: What the Radiologists Need to Know. Top. Magn. Reson. Imaging 2020, 29, 71–82. [Google Scholar] [CrossRef]
- Riquelme, P.A.; Drapeau, E.; Doetsch, F. Brain micro-ecologies: Neural stem cell niches in the adult mammalian brain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 123–137. [Google Scholar] [CrossRef]
- Fisher, J.L.; Schwartzbaum, J.A.; Wrensch, M.; Wiemels, J.L. Epidemiology of brain tumors. Neurol. Clin. 2007, 25, 867–890. [Google Scholar] [CrossRef]
- Linos, E.; Raine, T.; Alonso, A.; Michaud, D. Atopy and risk of brain tumors: A meta-analysis. J. Natl. Cancer Inst. 2007, 99, 1544–1550. [Google Scholar] [CrossRef] [PubMed]
- Wrensch, M.; Wiencke, J.K.; Wiemels, J.; Miike, R.; Patoka, J.; Moghadassi, M.; McMillan, A.; Kelsey, K.T.; Aldape, K.; Lamborn, K.R.; et al. Serum IgE, tumor epidermal growth factor receptor expression, and inherited polymorphisms associated with glioma survival. Cancer Res. 2006, 66, 4531–4541. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, H.G.; Galanis, E.; Weller, M. Glioblastoma. Handb. Clin. Neurol. 2016, 134, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477, Erratum in Cell 2014, 157, 753. [Google Scholar] [CrossRef]
- Farrell, C.J.; Plotkin, S.R. Genetic causes of brain tumors: Neurofibromatosis, tuberous sclerosis, von Hippel-Lindau, and other syndromes. Neurol. Clin. 2007, 25, 925–946. [Google Scholar] [CrossRef]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef]
- Ransom, D.T.; Ritland, S.R.; Kimmel, D.W.; Moertel, C.A.; Dahl, R.J.; Scheithauer, B.W.; Kelly, P.J.; Jenkins, R.B. Cytogenetic and loss of heterozygosity studies in ependymomas, pilocytic astrocytomas, and oligodendrogliomas. Genes Chromosomes Cancer 1992, 5, 348–356. [Google Scholar] [CrossRef]
- Cloughesy, T.F.; Cavenee, W.K.; Mischel, P.S. Glioblastoma: From molecular pathology to targeted treatment. Annu. Rev. Pathol. 2014, 9, 1–25. [Google Scholar] [CrossRef]
- Kleihues, P.; Ohgaki, H. Primary and secondary glioblastomas: From concept to clinical diagnosis. Neuro-Oncology 1999, 1, 44–51. [Google Scholar] [CrossRef]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Chen, X.; Zhang, S.; Shi, H.; Ye, Y.; Shi, H.; Zou, Z.; Li, P.; Guo, Q.; Ma, L.; et al. EGFR/SRC/ERK-stabilized YTHDF2 promotes cholesterol dysregulation and invasive growth of glioblastoma. Nat. Commun. 2021, 12, 177. [Google Scholar] [CrossRef] [PubMed]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Furnari, F.B.; Cloughesy, T.F.; Cavenee, W.K.; Mischel, P.S. Heterogeneity of epidermal growth factor receptor signalling networks in glioblastoma. Nat. Rev. Cancer 2015, 15, 302–310. [Google Scholar] [CrossRef]
- Andersson, U.; Guo, D.; Malmer, B.; Bergenheim, A.T.; Brännström, T.; Hedman, H.; Henriksson, R. Epidermal growth factor receptor family (EGFR, ErbB2-4) in gliomas and meningiomas. Acta Neuropathol. 2004, 108, 135–142. [Google Scholar] [CrossRef]
- Stommel, J.M.; Kimmelman, A.C.; Ying, H.; Nabioullin, R.; Ponugoti, A.H.; Wiedemeyer, R.; Stegh, A.H.; Bradner, J.E.; Ligon, K.L.; Brennan, C.; et al. Coactivation of receptor tyrosine kinases affects the response of tumor cells to targeted therapies. Science 2007, 318, 287–290. [Google Scholar] [CrossRef]
- Zhang, C.; Burger, M.C.; Jennewein, L.; Genßler, S.; Schönfeld, K.; Zeiner, P.; Hattingen, E.; Harter, P.N.; Mittelbronn, M.; Tonn, T.; et al. ErbB2/HER2-Specific NK Cells for Targeted Therapy of Glioblastoma. J. Natl. Cancer Inst. 2015, 108, djv375. [Google Scholar] [CrossRef]
- De Bacco, F.; Orzan, F.; Erriquez, J.; Casanova, E.; Barault, L.; Albano, R.; D’Ambrosio, A.; Bigatto, V.; Reato, G.; Patanè, M.; et al. ERBB3 overexpression due to miR-205 inactivation confers sensitivity to FGF, metabolic activation, and liability to ERBB3 targeting in glioblastoma. Cell Rep. 2021, 36, 109455. [Google Scholar] [CrossRef]
- Shang, R.; Lee, S.; Senavirathne, G.; Lai, E.C. microRNAs in action: Biogenesis, function and regulation. Nat. Rev. Genet. 2023. [Google Scholar] [CrossRef]
- Lukiw, W.J. MicroRNA (miRNA) Complexity in Alzheimer’s Disease (AD). Biology 2023, 12, 788. [Google Scholar] [CrossRef]
- Ishida, T.; Ueyama, T.; Ihara, D.; Harada, Y.; Nakagawa, S.; Saito, K.; Nakao, S.; Kawamura, T. c-Myc/microRNA-17-92 Axis Phase-Dependently Regulates PTEN and p21 Expression via ceRNA during Reprogramming to Mouse Pluripotent Stem Cells. Biomedicines 2023, 11, 1737. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Mafi, A.; Rahmati, A.; Babaei Aghdam, Z.; Salami, R.; Salami, M.; Vakili, O.; Aghadavod, E. Recent insights into the microRNA-dependent modulation of gliomas from pathogenesis to diagnosis and treatment. Cell. Mol. Biol. Lett. 2022, 27, 65. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs-microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Asadi-Moghaddam, K.; Chiocca, E.A.; Lawler, S.E. Potential role of miRNAs and their inhibitors in glioma treatment. Expert Rev. Anticancer Ther. 2010, 10, 1753–1762. [Google Scholar] [CrossRef]
- Guessous, F.; Zhang, Y.; Kofman, A.; Catania, A.; Li, Y.; Schiff, D.; Purow, B.; Abounader, R. microRNA-34a is tumor suppressive in brain tumors and glioma stem cells. Cell Cycle 2010, 9, 1031–1036. [Google Scholar] [CrossRef]
- Yuan, M.; Da Silva, A.C.A.L.; Arnold, A.; Okeke, L.; Ames, H.; Correa-Cerro, L.S.; Vizcaino, M.A.; Ho, C.Y.; Eberhart, C.G.; Rodriguez, F.J. MicroRNA (miR) 125b regulates cell growth and invasion in pediatric low grade glioma. Sci. Rep. 2018, 8, 12506. [Google Scholar] [CrossRef]
- Wang, D.D.; Ma, L.; Wong, M.P.; Lee, V.H.; Yan, H. Contribution of EGFR and ErbB-3 Heterodimerization to the EGFR Mutation-Induced Gefitinib and Erlotinib-Resistance in Non-Small-Cell Lung Carcinoma Treatments. PLoS ONE 2015, 10, e0128360. [Google Scholar] [CrossRef]
- Koganemaru, S.; Kuboki, Y.; Koga, Y.; Kojima, T.; Yamauchi, M.; Maeda, N.; Kagari, T.; Hirotani, K.; Yasunaga, M.; Matsumura, Y.; et al. U3-1402, a Novel HER3-Targeting Antibody-Drug Conjugate, for the Treatment of Colorectal Cancer. Mol. Cancer Ther. 2019, 18, 2043–2050. [Google Scholar] [CrossRef]
- Elhamamsy, A.R.; Metge, B.J.; Alsheikh, H.A.; Shevde, L.A.; Samant, R.S. Ribosome Biogenesis: A Central Player in Cancer Metastasis and Therapeutic Resistance. Cancer Res. 2022, 82, 2344–2353. [Google Scholar] [CrossRef]
- Zhai, W.; Comai, L. Repression of RNA polymerase I transcription by the tumor suppressor p53. Mol. Cell. Biol. 2000, 20, 5930–5938. [Google Scholar] [CrossRef] [PubMed]
- Voit, R.; Schäfer, K.; Grummt, I. Mechanism of repression of RNA polymerase I transcription by the retinoblastoma protein. Mol. Cell. Biol. 1997, 17, 4230–4237. [Google Scholar] [CrossRef] [PubMed]
- Ponti, D.; Bellenchi, G.C.; Puca, R.; Bastianelli, D.; Maroder, M.; Ragona, G.; Roussel, P.; Thiry, M.; Mercola, D.; Calogero, A. The transcription factor EGR1 localizes to the nucleolus and is linked to suppression of ribosomal precursor synthesis. PLoS ONE 2014, 9, e96037. [Google Scholar] [CrossRef] [PubMed]
- Ponti, D.; Bastianelli, D.; Rosa, P.; Pacini, L.; Ibrahim, M.; Rendina, E.A.; Ragona, G.; Calogero, A. The expression of B23 and EGR1 proteins is functionally linked in tumor cells under stress conditions. BMC Cell Biol. 2015, 16, 27. [Google Scholar] [CrossRef]
- Madera, S.; Izzo, F.; Chervo, M.F.; Dupont, A.; Chiauzzi, V.A.; Bruni, S.; Petrillo, E.; Merin, S.S.; De Martino, M.; Montero, D.; et al. Halting ErbB-2 isoforms retrograde transport to the nucleus as a new theragnostic approach for triple-negative breast cancer. Cell Death Dis. 2022, 13, 447. [Google Scholar] [CrossRef]
- Wang, Y.N.; Yamaguchi, H.; Hsu, J.M.; Hung, M.C. Nuclear trafficking of the epidermal growth factor receptor family membrane proteins. Oncogene 2010, 29, 3997–4006. [Google Scholar] [CrossRef]
- Sundvall, M.; Peri, L.; Määttä, J.A.; Tvorogov, D.; Paatero, I.; Savisalo, M.; Silvennoinen, O.; Yarden, Y.; Elenius, K. Differential nuclear localization and kinase activity of alternative ErbB4 intracellular domains. Oncogene 2007, 26, 6905–6914. [Google Scholar] [CrossRef]
- Farin, K.; Di Segni, A.; Mor, A.; Pinkas-Kramarski, R. Structure-function analysis of nucleolin and ErbB receptors interactions. PLoS ONE 2009, 4, e6128. [Google Scholar] [CrossRef]
- Wang, M.; Trim, C.M.; Gullick, W.J. Localisation of Neuregulin 1-β3 to different sub-nuclear structures alters gene expression. Exp. Cell Res. 2011, 317, 423–432. [Google Scholar] [CrossRef]
- Kumagai, S.; Koyama, S.; Nishikawa, H. Antitumour immunity regulated by aberrant ERBB family signalling. Nat. Rev. Cancer 2021, 21, 181–197. [Google Scholar] [CrossRef]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- De Bacco, F.; Boccaccio, C. ERBB3 as a therapeutic target in glioblastoma: Overexpression can make the difference. Mol. Cell. Oncol. 2021, 8, 1990677. [Google Scholar] [CrossRef] [PubMed]
- Ocana, A.; Vera-Badillo, F.; Seruga, B.; Templeton, A.; Pandiella, A.; Amir, E. HER3 overexpression and survival in solid tumors: A meta-analysis. J. Natl. Cancer Inst. 2013, 105, 266–273, Erratum in J. Natl. Cancer Inst. 2013, 105, 944. [Google Scholar] [CrossRef] [PubMed]
- Haratani, K.; Yonesaka, K.; Takamura, S.; Maenishi, O.; Kato, R.; Takegawa, N.; Kawakami, H.; Tanaka, K.; Hayashi, H.; Takeda, M.; et al. U3-1402 sensitizes HER3-expressing tumors to PD-1 blockade by immune activation. J. Clin. Investig. 2020, 130, 374–388. [Google Scholar] [CrossRef]
- Hong, M.; Yoo, Y.; Kim, M.; Kim, J.Y.; Cha, J.S.; Choi, M.K.; Kim, U.; Kim, K.; Sohn, Y.; Bae, D.; et al. A Novel Therapeutic Anti-ErbB3, ISU104 Exhibits Potent Antitumorigenic Activity by Inhibiting Ligand Binding and ErbB3 Heterodimerization. Mol. Cancer Ther. 2021, 20, 1142–1152. [Google Scholar] [CrossRef]
- Cejalvo, J.M.; Jacob, W.; Fleitas Kanonnikoff, T.; Felip, E.; Navarro Mendivil, A.; Martinez Garcia, M.; Taus Garcia, A.; Leighl, N.; Lassen, U.; Mau-Soerensen, M.; et al. A phase Ib/II study of HER3-targeting lumretuzumab in combination with carboplatin and paclitaxel as first-line treatment in patients with advanced or metastatic squamous non-small cell lung cancer. ESMO Open 2019, 4, e000532. [Google Scholar] [CrossRef]
- Mirschberger, C.; Schiller, C.B.; Schräml, M.; Dimoudis, N.; Friess, T.; Gerdes, C.A.; Reiff, U.; Lifke, V.; Hoelzlwimmer, G.; Kolm, I.; et al. RG7116, a therapeutic antibody that binds the inactive HER3 receptor and is optimized for immune effector activation. Cancer Res. 2013, 73, 5183–5194. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Koyama, K.; Kamai, Y.; Hirotani, K.; Ogitani, Y.; Zembutsu, A.; Abe, M.; Kaneda, Y.; Maeda, N.; Shiose, Y.; et al. A Novel HER3-Targeting Antibody-Drug Conjugate, U3-1402, Exhibits Potent Therapeutic Efficacy through the Delivery of Cytotoxic Payload by Efficient Internalization. Clin. Cancer Res. 2019, 25, 7151–7161. [Google Scholar] [CrossRef]
- Gandullo-Sánchez, L.; Ocaña, A.; Pandiella, A. HER3 in cancer: From the bench to the bedside. J. Exp. Clin. Cancer Res. 2022, 41, 310. [Google Scholar] [CrossRef]
- Pan, P.C.; Magge, R.S. Mechanisms of EGFR Resistance in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 8471. [Google Scholar] [CrossRef]
- Hafeez, U.; Parslow, A.C.; Gan, H.K.; Scott, A.M. New insights into ErbB3 function and therapeutic targeting in cancer. Expert Rev. Anticancer Ther. 2020, 20, 1057–1074. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The receptor tyrosine kinases family (RTKs). The members of the Receptor Tyrosine Kinase (RTK) family (ErbB1, ERbB2, ERbB3 and ErbB4), play pivotal roles in a wide array of biological processes, encompassing apoptosis, cell cycle regulation, cell division, cytoskeletal organization, cell differentiation, developmental processes, immune responses, and dynamics within the nervous system. These receptors achieve their functions by interacting with specific ligands, making them some of the most extensively studied receptors in signal transduction and their involvement in oncogenesis. In the diagram, the Epidermal Growth Factor Receptor (ErbB1/EGFR, depicted in grey) is surrounded by its potential ligands, including amphiregulin (AREG), epiregulin (EREG), epigen (EPGN), epidermal growth factor (EGF), heparin-binding epidermal growth factor (HB-EGF), betacellulin (BTC), and transforming growth factor-alpha (TGF-a). ErbB2 (shown in green) is in its active conformation. ErbB3 (colored blue) is presented in conjunction with its ligands neuregulin (NRG-1) and neuregulin-2 (NRG-2), while ErbB4 (in yellow) is surrounded by its specific ligands, which include EREG, HB-EGF, NRG-1, NRG-2, NRG-3, and NRG-4.
Figure 1.
The receptor tyrosine kinases family (RTKs). The members of the Receptor Tyrosine Kinase (RTK) family (ErbB1, ERbB2, ERbB3 and ErbB4), play pivotal roles in a wide array of biological processes, encompassing apoptosis, cell cycle regulation, cell division, cytoskeletal organization, cell differentiation, developmental processes, immune responses, and dynamics within the nervous system. These receptors achieve their functions by interacting with specific ligands, making them some of the most extensively studied receptors in signal transduction and their involvement in oncogenesis. In the diagram, the Epidermal Growth Factor Receptor (ErbB1/EGFR, depicted in grey) is surrounded by its potential ligands, including amphiregulin (AREG), epiregulin (EREG), epigen (EPGN), epidermal growth factor (EGF), heparin-binding epidermal growth factor (HB-EGF), betacellulin (BTC), and transforming growth factor-alpha (TGF-a). ErbB2 (shown in green) is in its active conformation. ErbB3 (colored blue) is presented in conjunction with its ligands neuregulin (NRG-1) and neuregulin-2 (NRG-2), while ErbB4 (in yellow) is surrounded by its specific ligands, which include EREG, HB-EGF, NRG-1, NRG-2, NRG-3, and NRG-4.

Figure 2.
Expression of ErbBs mRNA in the Central Nervous System (nTPM: normalized transcript expression) https://www.proteinatlas.org/ (access on 2 October 2023).
Figure 2.
Expression of ErbBs mRNA in the Central Nervous System (nTPM: normalized transcript expression) https://www.proteinatlas.org/ (access on 2 October 2023).

Figure 3.
Genomic location of human ErbB3 gene. The exons are in black and the introns in blue.
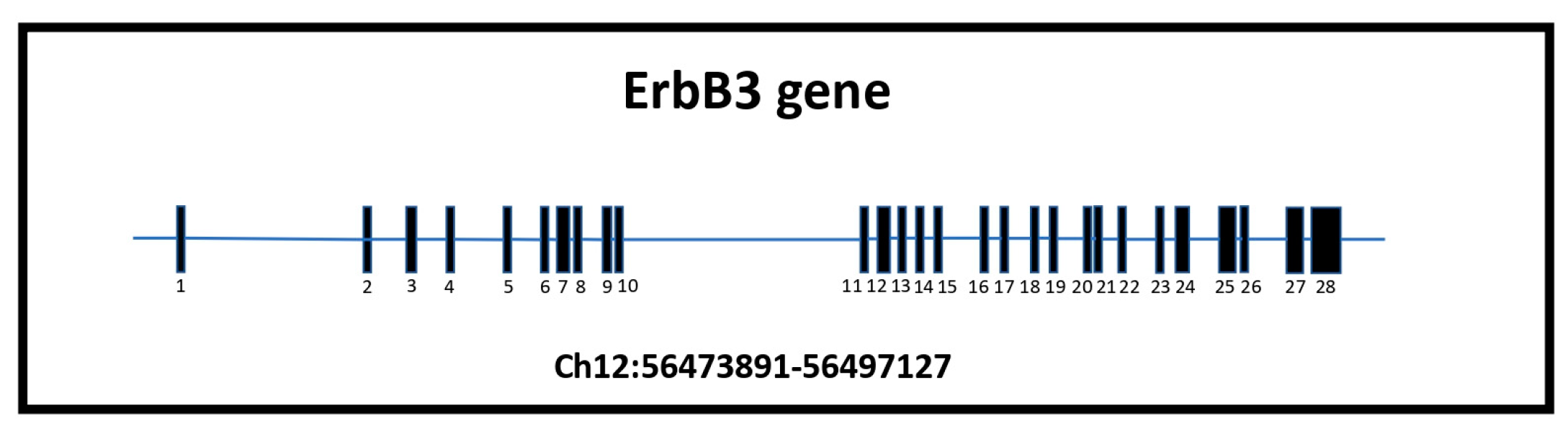
Figure 4.
Schematic diagram showing ErbB3 Variants’ locations and functions inside and outside cells.
Figure 4.
Schematic diagram showing ErbB3 Variants’ locations and functions inside and outside cells.
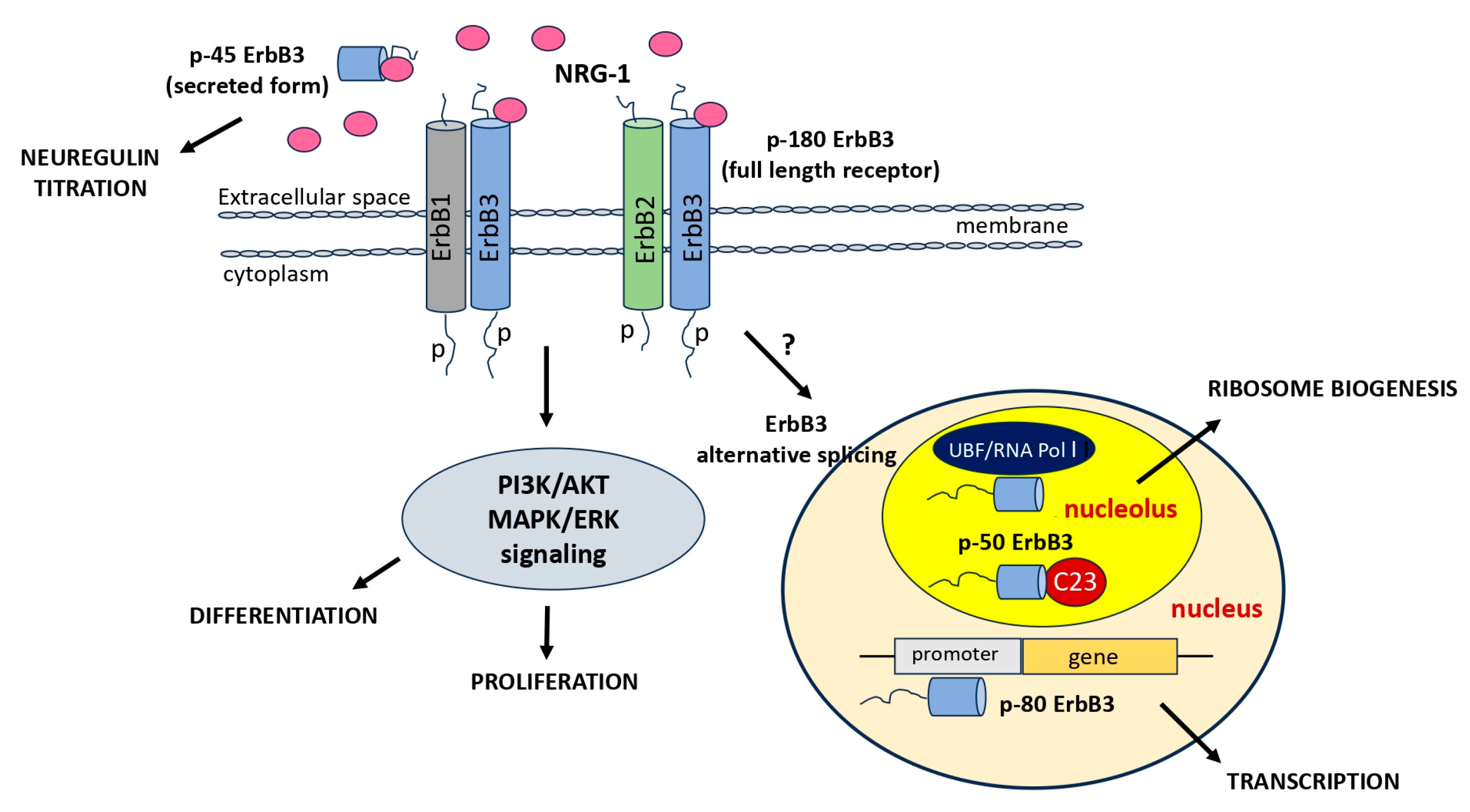
Figure 5.
Schematic diagram of the ErbB3 inhibition in cancer cells (BxPC3 and SKBR3 cell lines, which are models for NRG1-dependent and -independent cell proliferation, respectively). The ErbB3 inhibitors directly bind the receptor and prevent conformational changes necessary for heterodimerization. Following NRG-1 binding, ErbB3 forms heterodimers with other members of the EGFR family, initiating phosphorylation cascades that stimulate cell proliferation, differentiation, and survival. Among these dimers, the ErbB2–ErbB3 complex is the most active. ErbB3 plays a pivotal role in cancer progression by mediating resistance to EGFR inhibitors and promoting metastasis. Two anticancer drugs, Cetuximab targeting ErbB1(EGFR) and Trastuzumab targeting ErbB2, have been utilized in cancer treatment. These drugs have shown clinical benefits, but a substantial number of patients do not respond or develop resistance to them.
Figure 5.
Schematic diagram of the ErbB3 inhibition in cancer cells (BxPC3 and SKBR3 cell lines, which are models for NRG1-dependent and -independent cell proliferation, respectively). The ErbB3 inhibitors directly bind the receptor and prevent conformational changes necessary for heterodimerization. Following NRG-1 binding, ErbB3 forms heterodimers with other members of the EGFR family, initiating phosphorylation cascades that stimulate cell proliferation, differentiation, and survival. Among these dimers, the ErbB2–ErbB3 complex is the most active. ErbB3 plays a pivotal role in cancer progression by mediating resistance to EGFR inhibitors and promoting metastasis. Two anticancer drugs, Cetuximab targeting ErbB1(EGFR) and Trastuzumab targeting ErbB2, have been utilized in cancer treatment. These drugs have shown clinical benefits, but a substantial number of patients do not respond or develop resistance to them.
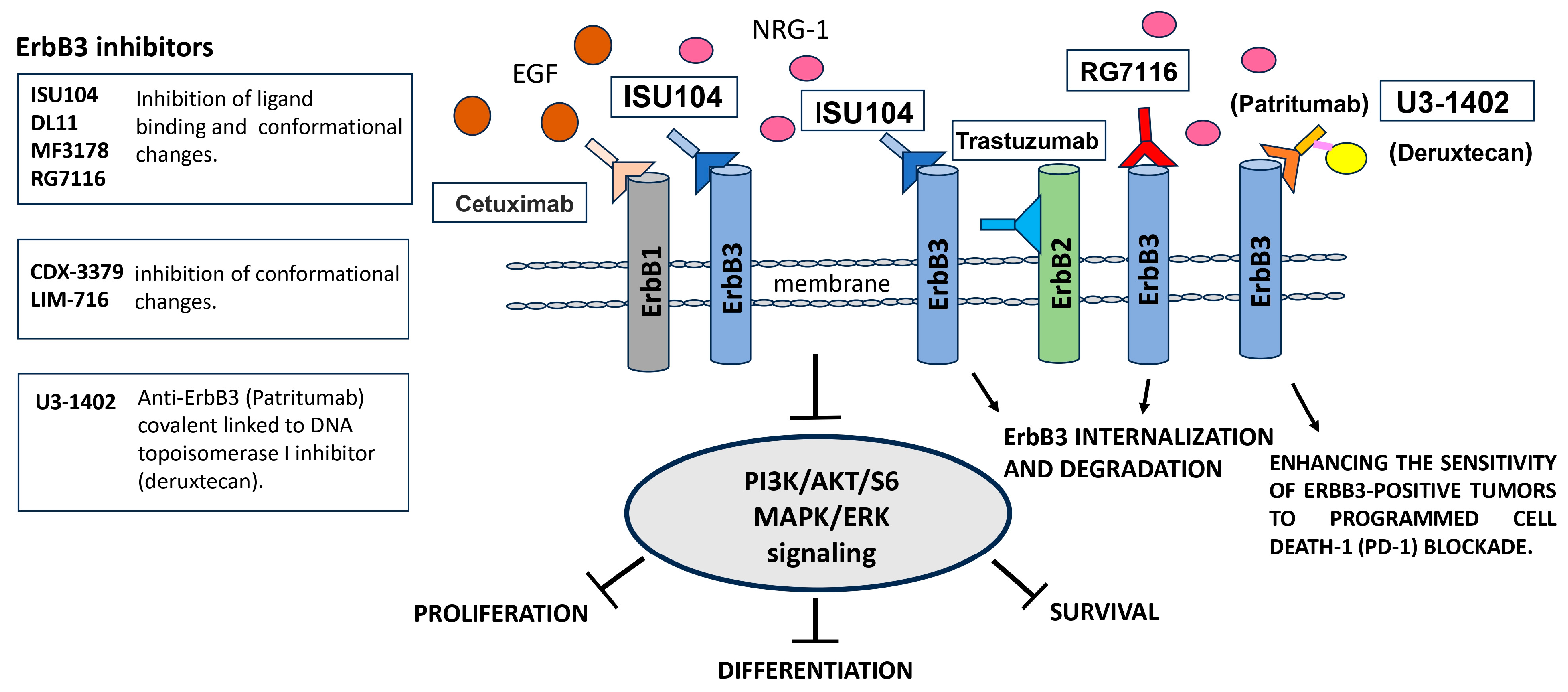
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tagliaferro, M.; Ponti, D. The Signaling of Neuregulin-Epidermal Growth Factor Receptors and Its Impact on the Nervous System. Neuroglia 2023, 4, 253-274. https://doi.org/10.3390/neuroglia4040018
AMA Style
Tagliaferro M, Ponti D. The Signaling of Neuregulin-Epidermal Growth Factor Receptors and Its Impact on the Nervous System. Neuroglia. 2023; 4(4):253-274. https://doi.org/10.3390/neuroglia4040018
Chicago/Turabian StyleTagliaferro, Marzia, and Donatella Ponti. 2023. "The Signaling of Neuregulin-Epidermal Growth Factor Receptors and Its Impact on the Nervous System" Neuroglia 4, no. 4: 253-274. https://doi.org/10.3390/neuroglia4040018