
Microbiome-Glia Crosstalk: Bridging the Communication Divide in the Central Nervous System

Department of Biology, Faculty of Science, University of the Fraser Valley, Abbotsford, BC V2S 7M8, Canada
Neuroglia 2024, 5(2), 89-104; https://doi.org/10.3390/neuroglia5020007
Submission received: 2 March 2024
/
Revised: 17 April 2024
/
Accepted: 19 April 2024
/
Published: 21 April 2024
Abstract
:The traditional neuron-centric view of the central nervous system (CNS) is shifting toward recognizing the importance of communication between the neurons and the network of glial cells. This shift is leading to a more comprehensive understanding of how glial cells contribute to CNS function. Alongside this shift, recent discoveries have illuminated the significant role of the human microbiome, comprising trillions of microorganisms, mirroring the number of human cells in an individual. This paper delves into the multifaceted functions of neuroglia, or glial cells, which extend far beyond their traditional roles of supporting and protecting neurons. Neuroglia modulate synaptic activity, insulate axons, support neurogenesis and synaptic plasticity, respond to injury and inflammation, and engage in phagocytosis. Meanwhile, the microbiome, long overlooked, emerges as a crucial player in brain functionality akin to glial cells. This review aims to underscore the importance of the interaction between glial cells and resident microorganisms in shaping the development and function of the human brain, a concept that has been less studied. Through a comprehensive examination of existing literature, we discuss the mechanisms by which glial cells interface with the microbiome, offering insights into the contribution of this relationship to neural homeostasis and health. Furthermore, we discuss the implications of dysbiosis within this interaction, highlighting its potential contribution to neurological disorders and paving the way for novel therapeutic interventions targeting both glial cells and the microbiome.
{kind=link}
{kind=link}
1. Introduction
For a long time, neurons have been considered the key players in brain function, while glial cells have been overlooked. However, a growing body of studies has been shedding light on the crucial role of glial cells in regulating neural activity and preserving brain health. This paradigm shift in neuroscience is gradually revealing the true complexity of the brain and challenging our traditional understanding of its workings [1,2,3,4,5]. Emerging evidence is now unveiling the intricate interplay between the composition of our microbiome and the functionality of our central nervous system as well [6,7,8,9,10,11,12]. These findings not only underscore the complexity of brain physiology, but also underscore the profound influence of gut microbiome on cognitive well-being [12,13,14]. As our understanding deepens, the comprehensive view of brain function evolves, highlighting the collaborative interactions of neurons, glial cells, and the microbiome in shaping the functions of our central nervous system, opening new avenues for therapeutical interventions in neurological disorders.
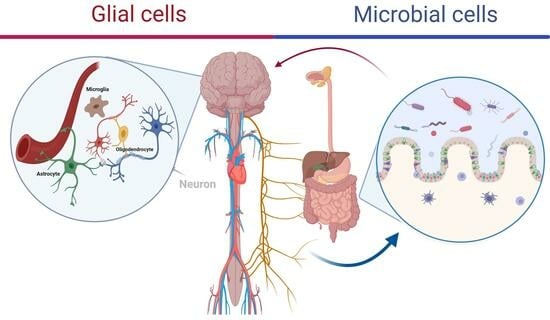
Glia, also known as glial cells or neuroglia, are non-neuronal cells in the central nervous system that include astrocytes, microglia, and oligodendrocytes. They emerge as crucial mediators in this complex dialogue (Figure 1) [15,16,17]. These cells, once viewed primarily as support structures, are now recognized for their active participation in the microbial brain axis, a relatively recent concept that underscores the bidirectional interactions between the brain and the microbiota [18,19,20].
Astrocytes, radial glia, and oligodendrocytes originate from neural cells, while microglia differentiate from blood monocytes during ontogeny. Microglia, the parenchymal macrophages of the central nervous system, make their first appearance in the neuroepithelium during embryonic day 9.5 and then proceed to spread throughout the parenchyma of the CNS. They undergo a gradual process of differentiation, culminating in their final maturation state during the second week of postnatal development [21,22].
In the developmental timeline, neurons typically precede glial cells. Glial cells play a crucial role in neuronal development by offering trophic support vital for neuronal survival. They are also involved in processes such as neuronal migration, axon and dendrite outgrowth, and synaptogenesis [23]. We should acknowledge that there are other types of neuroglia within the CNS such as ependymal cells that line the ventricles of the brain and the central canal of the spinal cord and contribute to the production and circulation of cerebrospinal fluid or radial glial cells, specialized cells in the developing nervous system of all vertebrates [24,25,26]. However, because our understanding of their involvement in microbial CNS communication is very limited, they will not be discussed in this review.
Recent findings have illuminated the substantial contribution of neuroglial cells in orchestrating this crosstalk [20,27]. Astrocytes, for instance, play a crucial role in modulating the neuroinflammatory response and maintaining homeostasis in the brain environment in response to microbial signals from the gut microbiome [7,18,20,28,29]. Microglia, the resident immune cells of the CNS, are now acknowledged for their role in surveilling and responding to microbial threats [30]. In addition, they contribute to modulating the gut microbiome in orchestration with oligodendrocytes [31,32]. Oligodendrocytes, traditionally associated with myelin production, are implicated in shaping the neural environment and influencing microbial interactions [33]. These insights into neuroglial cells’ multifaceted functions challenge traditional perspectives and open new avenues for understanding the intricate relationships within the microbial brain axis. The once-clear boundaries between the central nervous system and the microbial world are now giving way to a more nuanced understanding of the dynamic interplay that influences both neurological health and microbial communities.
2. Microglia: Guardians of Neural Health
Microglia, the resident immune cells of the CNS, play a pivotal role in detecting and responding to microbial activities [30,34]. They are the innate immune cells of the central nervous system (CNS), function as brain macrophages, and are unevenly distributed throughout the brain [23]. Notably, regions such as the hippocampal dentate gyrus with the largest population (12%), substantia nigra, basal ganglia, and olfactory telencephalon in mice harbor significant microglial populations. These cells self-renew over an organism’s lifespan and display variations in size and branching patterns across different layers of the cerebellar cortex due to interactions with neurons and intrinsic mechanisms [23,35,36]. Crucially, microglia play a pivotal role in regulating the neuronal network by facilitating brain development, maintenance, and repair through debris clearance and phagocytosis of pathogens. They undergo distinct morphological and functional stages, with pro-inflammatory microglia (M1 state) releasing cytokines associated with dysfunction upon chronic activation, while neuroprotective microglia (M2 state) engage in tissue repair, extracellular matrix reconstruction and support neuron survival via neurotrophic factors [37,38,39,40,41,42,43,44].
Their dynamic functions extend beyond immune surveillance to include the modulation of neuroinflammation, a critical process in the CNS’s response to microbial activities. For example, through pattern recognition receptors (PRRs), microglia recognize pathogen-associated molecular patterns (PAMPs), initiating immune responses [45,46]. Microglial activation contributes to the orchestration of neuroinflammatory responses and the clearance of invading pathogens as well as controlling the composition of the gut microbiota [34]. Let us delve deeper into how microglia, as the resident immune cells of the central nervous system (CNS), contribute to modulating neuroinflammation through their ability to detect and respond to microbial life.
2.1. Microorganisms and Microglial States
Microglial activation is a dynamic process that can manifest in different states, ranging from a pro-inflammatory (M1) phenotype to an anti-inflammatory (M2) phenotype [47]. M1 microglia are associated with the release of pro-inflammatory cytokines and reactive oxygen species, contributing to neuroinflammation. In response to microbial challenges, microglia may undergo M1 activation to mount an immune response aimed at clearing pathogens. However, dysregulated or prolonged M1 activation can contribute to chronic neuroinflammation and neuronal damage [30,39,48].
However, the significance of these microglial responses is not restricted to the pathological state of the central nervous system. Research has demonstrated that the normal microbiome plays a crucial role in controlling the maturation and activation of microglia, the guardians of neural immunity under homeostatic conditions. Erny et al. [49], in a study published in Nature Neuroscience, highlighted the significant contributions of the host microbiota to microglial homeostasis. They observed that germ-free (GF) mice exhibited widespread defects in microglia, including altered cell proportions and an immature phenotype, resulting in impaired innate immune responses. Temporarily eliminating the host microbiota drastically altered microglia properties, while a limited microbiota complexity also led to defective microglia. Conversely, recolonization with a complex microbiota partially restored microglial features. The study identified short-chain fatty acids (SCFA)—microbial fermentation products—as regulators of microglial homeostasis. Mice lacking the SCFA receptor FFAR2 showed similar microglial defects as seen in GF conditions. These findings underscore the vital role of host bacteria in regulating microglia maturation and function, with complex microbiota offering some remedial potential for microglial impairment [49].
2.2. Pattern Recognition Receptors in Microglia
Microglia express a repertoire of pattern recognition receptors (PRRs), including toll-like receptors (TLRs), nucleotide-binding oligomerization domain (NOD)-like receptors, and scavenger receptors. These receptors enable microglia to recognize pathogen-associated molecular patterns (PAMPs) on microbial surfaces [45,46].
PAMPs are molecular structures possessing specific physicochemical traits, typically found in repetitive or polymeric forms which are recognized by PRRs rather than chemically defined epitopes [46,50]. Among the extensively studied PRRs are toll-like receptors (TLRs). Other membrane-bound PRRs include the scavenger receptor family, the receptor for advanced glycation end products (RAGE), and C-type lectin receptors [51,52]. C-type lectins are specifically expressed on microglia but not on neurons or astrocytes. They play a crucial role in identifying pathogenic invasions and are specified to identify carbohydrates [52,53].
The complement system constitutes another category of extracellular PRRs wherein the membrane signaling components (complement receptors) are distinct from the detection and effector components (soluble complement factors). Furthermore, numerous intracellular PRRs have been identified, capable of recognizing PAMPs within various intracellular compartments [51,54].
Microglia express a wide array of toll-like receptors (TLRs), along with the necessary adaptor molecules for downstream TLR signaling [42]. Upon activation, microglia exhibit an upregulation of TLR expression, consistent with their role as the primary defenders against invading pathogens in the brain. Even with a simplified model using a single artificial ligand, microglia respond to TLR activation by producing various antiviral and antibacterial molecules such as TNF, IFN-β, inducible nitric oxide synthetase, a range of chemokines, and both pro- and anti-inflammatory interleukins, along with promoting cell proliferation. Specific stimulation of microglia via either TLR2 or TLR4 with lipoteichoic acid (LTA) or lipopolysaccharide (LPS), respectively, results in the release of proinflammatory cytokines [42,48,54,55].
When microglia are co-stimulated with two or more TLR ligands, each leading to a submaximal stimulus individually, a synergistic response is induced [56]. Combining different TLR2 or TLR4 ligands or mixing TLR2/TLR4, TLR4/TLR9, and TLR2/TLR9 ligands prompts microglia to release nitric oxide synergistically. This finding holds significance for effective immune responses since many microorganisms express ligands for multiple TLR receptors. For instance, while peptidoglycans (PGN) or LTA from Staphylococcus aureus can activate microglia via TLR2, the presence of TLR2 is not indispensable for recognizing intact gram-positive bacteria. Deleting TLR2 reduces proinflammatory cytokine production in Staphylococcus-aureus-infected CNS tissue [46,50].
2.3. Neuroinflammatory Responses and Microglia
Microglial activation is a key component of the neuroinflammatory response, which involves the recruitment of immune cells, release of inflammatory mediators, and alterations in the local microenvironment. Upon activation of microglia with pattern recognition receptors, several pro-inflammatory cytokines are released that influence neighboring cells, including astrocytes and neurons, further amplifying the neuroinflammatory cascade. Additionally, activated microglia contribute to the breakdown of the blood–brain barrier, facilitating immune cell infiltration into the CNS [38,57].
The acute inflammatory response serves as a crucial defense mechanism against nearby infected, diseased, or damaged cells. Yet, if this response persists or becomes chronic, it can worsen neurodegeneration. Activated microglia represent a pathological hallmark of neurodegenerative disorders such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and amyotrophic lateral sclerosis (ALS), making the modulation of these microglial responses an area of therapeutic focus. In this section, we will delve into the pro-inflammatory signaling pathways of microglia, particularly those relevant to neurodegenerative diseases [38,47].
Microglia also contribute to the resolution of neuroinflammation by transitioning to an anti-inflammatory M2 phenotype. M2 microglia release anti-inflammatory cytokines and growth factors, promoting tissue repair and dampening the pro-inflammatory response. Dysregulation of the transition from M1 to M2 states may contribute to the persistence of neuroinflammation and the development of chronic neurological disorders [44,58]. Normal microbiome plays an essential role in the transition between these states and the general health of the central nervous system [49].
Understanding the nuanced role of microglia in modulating neuroinflammation provides insights into the complex interplay between the CNS and microbial challenges. Targeting microglial functions may hold therapeutic potential for managing neuroinflammatory conditions and maintaining neural health in interactions with pathogenic or commensal microorganisms.
2.4. The Role of Microglia in the Crosstalk in the Microbiome–Brain Axis
In addition to the influence of pathogenic microorganisms on shaping the response and polarity of our microglial cells within the nervous system, recent insights suggest that our resident microbiome also plays a regulatory role in modulating the function of microglia within our central nervous system, particularly regarding the underlying mechanisms of spatial learning and memory impairment associated with microglial M1/M2 phenotypes.
Wu et al.’s study [59] uncovered that disturbances in gut microbiota trigger a shift in microglial phenotype from M2 to M1, accompanied by changes in the pro-brain-derived neurotrophic factor (proBDNF)–p75NTR–mature BDNF (mBDNF)–TrkB signaling pathway. This disruption leads to decreased hippocampal neurogenesis and levels of synaptic plasticity-related proteins (SYN, PSD95, and MAP2), ultimately impairing spatial learning and memory. Notably, certain bacterial strains such as Clostridia, Bacteroides, Lactobacillus, and Muribaculaceae play a significant role in maintaining the balance between microglial M1 and M2 phenotypes, contributing to spatial learning and memory decline following chronic METH exposure. Moreover, fecal microbial transplantation shows promise in alleviating spatial learning and memory deficits by restoring microglial M1/M2 equilibrium and subsequent proBDNF-p75NTR/mBDNF-TrkB signaling in the hippocampi of chronically METH-exposed mice [59].
In another study, Sharma et al. [60] showed that inhibiting microglial activation alone significantly reduces hypertension, accompanied by distinct changes in gut microbial communities and a notable improvement in gut health. It had been shown previously that increased microglial activation and neuroinflammation in autonomic brain regions are linked to sustained hypertension, with minocycline, an anti-inflammatory antibiotic, showing promise in mitigating these effects. To address whether the benefits of minocycline in controlling hypertension lie in its anti-inflammatory or antimicrobial action, they used chemically modified tetracycline-3 (CMT-3), known for its potent anti-inflammatory properties, and demonstrated the critical role of microglia–microbiome interactions [60].
Interestingly, through a comprehensive study, Mossad et al. [61] showed how specific aspects of microglia in aging are influenced by the gut microbiota. They conducted a study comparing microglial transcriptomes in young adult and aged mice housed under germ-free and specific pathogen-free conditions and demonstrated that the microbiota influences age-related changes in microglial gene expression. In aged mice, the absence of gut microbiota was shown to reduce oxidative stress and improve mitochondrial function in microglia. Metabolomic analyses revealed the accumulation of N6-carboxymethyllysine (CML) in microglia of the aging brain, leading to increased reactive oxygen species production and impaired mitochondrial activity. This age-dependent rise in CML levels was validated in humans. Additionally, a microbiota-dependent increase in intestinal permeability in aged mice was linked to elevated CML levels [61].
3. Astrocytes: Orchestrators of Homeostasis
Astrocytes, long regarded as supportive cells, are now recognized for their active involvement in CNS–microbiome communication [18,62,63,64]. Astrocytes are intimately involved in regulating synaptic function and plasticity. By modulating neurotransmitter levels, astrocytes can impact synaptic strength and the balance between excitation and inhibition. The bidirectional communication between astrocytes and neurons allows for the fine-tuning of synaptic transmission, ensuring optimal neural network activity [2,5,15,19,63,65].
Recent studies argue that astrocytes can sense bacterial metabolites and respond by modulating neurotransmitter levels, influencing synaptic function, and contributing to neuroinflammation [7,8,20,66]. This bidirectional communication suggests a potential role for astrocytes in shaping the neural response to microbial challenges and the normal microbiome.
It is conceivable that, upon detecting bacterial metabolites, astrocytes could regulate neurotransmitter levels in the extracellular space and, consequently, regulate neural networks. For instance, it is known that astrocytes uptake the excessive accumulated glutamate from the synaptic cleft via glutamate transporters [64,67,68]. Glutamate is the main excitatory neurotransmitter in our central nervous system [2,69,70]. If bacterial metabolites indeed have an impact on the expression and function of these transporters, such modulation could potentially disrupt the clearance of neurotransmitters, leading to changes in synaptic transmission. However, it remains to be confirmed through empirical testing and experimental validation.
3.1. Sensing and Responding to Bacterial Metabolites
Astrocytes have long been known to react to activators of innate immune responses, such as microbial products, environmental toxins, and various forms of tissue damage. In healthy tissue, astrocytes express numerous receptors, including G protein-coupled receptors (GPCRs) and pattern recognition receptors (PRRs), for pathogen-associated molecular patterns (PAMPs) and damage- or danger-associated molecular patterns (DAMPs). These receptors are capable of initiating innate immune responses, particularly through toll-like receptors (TLRs) among which TLR4 is commonly associated with microglia but is also present in astrocytes [71]. Bacterial metabolites, such as short-chain fatty acids (SCFAs) or some tryptophan metabolites produced by gut microbiota, can cross the blood–brain barrier and reach astrocytes, acting as signaling molecules [7,8,20,66].
Studies have shown that astrocytes respond to SCFAs, which are bacterial metabolites produced by gut bacteria during the fermentation of dietary fiber. SCFAs can modulate astrocytic function and influence neurotransmitter release [27].
Sodium butyrate, an abnormal metabolic product of short-chain fatty acids (SCFAs), has been found to significantly increase the activation of astrocytes, reduce dopamine and 5-HT levels, worsen the decline of dopaminergic neurons, and ultimately exacerbate neuroinflammation in the brains of MPTP-treated mice. The dopaminergic neurotoxin MPTP specifically targets dopamine neurons in the substantia nigra pars compacta [72]. Moreover, in vitro studies have showed that sodium butyrate is able to exert potent anti-inflammatory effects against LPS-induced responses in neural co-cultures comprising microglial cells, astrocytes, and cerebellar granule neurons [73]. Additionally, extracts from myxobacteria who produce various bioactive metabolites, have been found to enhance cellular glutathione levels, which protects human primary astrocytic cultures from oxidative stress [74].
Dysregulation of astrocyte responses to bacterial signals has been implicated in conditions such as neurodegenerative diseases, emphasizing the potential role of astrocytes in shaping the neural response to microbial challenges. Understanding the nuanced ways in which astrocytes sense bacterial metabolites and contribute to neural responses opens new avenues for investigating the impact of the microbiome on brain health. Further research into astrocyte–microbiome interactions holds promise for uncovering therapeutic targets and strategies for managing neurological disorders influenced by dysregulated neural–microbial communication.
3.2. Contribution to the Regulation of Neuroinflammation
Astrocytes actively participate in neuroinflammatory responses. Bacterial metabolites can trigger signaling pathways within astrocytes, leading to the release of pro-inflammatory cytokines and chemokines. After identifying pathogens, astrocytes, along with microglia, promptly release various soluble immune factors, including the well-known inflammatory cytokines interleukin (IL)-1β and IL-6. These responses facilitate the recruitment and activation of leukocytes upon reaching the infection site, a process which is crucial for effectively clearing pathogens. However, excessive or prolonged responses can lead to detrimental effects. Chronic exposure to microbial challenges or dysregulation of astrocyte responses may contribute to sustained neuroinflammation, which is implicated in the pathogenesis of various neurological disorders.
In contrast, Burmeister et al. have demonstrated that IL-19 and IL-24, similar to IL-10, are expressed by astrocytes in response to a bacterial challenge, albeit with a delay, and that they exert an immunosuppressive effect on glial cells [75,76]. Another member of the IL-10 family, IL-20, is also suggested to be produced by cells within the central nervous system (CNS). Studies have shown its expression in glioblastoma cells, and staining associated with IL-20 has been observed in glia-like cells in murine models of ischemic stroke [77]. Additionally, bacterial lipopolysaccharide (LPS) has been found to induce the expression of IL-20 mRNA in mixed glial cell cultures. This effect has been shown to be dependent on myeloid differentiation factor 88 (MyD88) via induction through TLR4 signaling [78]. Notably, the expression of subunits composing type 1 and type 2 IL-20 receptors has been reported in murine astrocytes, suggesting the susceptibility of astrocytes to the immunosuppressive effects of this cytokine [29].
3.3. Astrocytic Interactions with Microorganisms at the Blood–Brain Barrier
Astrocytes are intimately associated with the Blood–Brain Barrier (BBB), with their end-feet processes ensheathing the cerebral vasculature, providing structural support, and participating in BBB integrity maintenance and immune regulation [79]. Astrocytes, as a type of glial cell contributing to BBB, play a crucial role in defending the brain against harmful microorganisms. However, further investigation is needed to explore the implications of communication between the normal microbiome and the astrocytic members of the BBB for the maintenance of brain homeostasis and overall health. Due to their significant role in maintaining the integrity of the BBB, we review their interactions with microorganisms in general.
The blood–brain barrier is a highly specialized interface comprised of endothelial cells, pericytes, and astrocytic end-feet processes. It forms a selective barrier that tightly regulates the exchange of molecules between the bloodstream and the central CNS [80]. Beyond its conventional role in maintaining CNS homeostasis, the BBB also serves as the primary line of defense against invading pathogens, including bacteria, viruses, and fungi.
As mentioned earlier, astrocytes express an array of pattern recognition receptors (PRRs), including toll-like receptors (TLRs) as well as nucleotide-binding oligomerization domain-like receptors (NLRs) and retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs), enabling them to recognize pathogen-associated molecular patterns (PAMPs) derived from invading microorganisms [46,81]. Upon detection of microbial components, astrocytes initiate immune signaling cascades, leading to the activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and the production of pro-inflammatory cytokines, such as interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF-α). Additionally, astrocytes release antimicrobial peptides and reactive oxygen species (ROS), contributing to the elimination of microbial pathogens and the containment of CNS infections [82]. Furthermore, astrocyte-derived factors modulate the expression of tight junction proteins in brain endothelial cells, reinforcing the barrier function of the BBB and limiting microbial penetration into the CNS parenchyma [83]. The principal structural components of the blood–brain barrier are the tight junctions found among the endothelial cells of brain capillaries. It is not yet clear if astrocytes within the BBB use the same molecular mechanisms to communicate with metabolites of the microbiome despite their important role as the first line of defense in the brain.
3.4. The Role of Astrocytes in the Crosstalk in the Microbiome-Brain Axis
The gut–brain axis serves as a conduit for communication between the gut microbiota and the CNS. Astrocytes, positioned at the interface between blood vessels and neurons, can integrate signals from the periphery and influence local neural circuits. Through this crosstalk, astrocytes may play a role in translating gut-derived signals into neural responses, influencing behavior and cognitive function.
In addition to our previous understanding of interactions and the harmful effects of microbial toxins, enzymes, and products, emerging research is uncovering the regulatory role of microbiome products in the overall brain health and function. For instance, Rothhammer et al. [20,66] have demonstrated that dietary tryptophan is metabolized by gut microbiota into AHR agonists, which play a role in limiting CNS inflammation by affecting astrocytes. Ampicillin treatment during the recovery phase has been shown to lead to increased EAE scores, while supplementation with tryptophan metabolites such as indole, indoxyl-3-sulfate, indole-3-propionic acid, and indole-3-aldehyde, or the bacterial enzyme tryptophanase, has been shown to reduce CNS inflammation in antibiotic-treated mice. The EAE score refers to the clinical scoring system used in experimental autoimmune encephalomyelitis (EAE), which is an animal model of multiple sclerosis (MS). Individuals with multiple sclerosis (MS) have been shown to exhibit decreased circulating levels of AHR agonists. These findings suggest that interferon-Is produced within the CNS, along with metabolites derived from dietary tryptophan by the gut flora, cooperate to activate AHR signaling in astrocytes, thereby suppressing CNS inflammation [20,66].
Another study has illustrated how the metabolism of amino acids such as tyrosine and phenylalanine by gut microbes, resulting in p-cresol and subsequent conversion to p-cresol glucuronide (pCG) by host enzymes, can profoundly influence components of the blood–brain barrier, including the astrocytic members of BBB [84]. This study examined pCG impact on the cerebral vasculature and brain function both in vitro and in vivo, utilizing male C57Bl/6 J mice to gauge BBB permeability and brain-wide genetic expression changes following pCG treatment. Additionally, the study investigated its effects on a human cerebromicrovascular endothelial cell line, hCMEC/D3, evaluating permeability, electrical resistance, and barrier protein expression. Mice treated with pCG exhibited decreased BBB permeability and significant shifts in brain gene expression. Surprisingly, pCG alone had minimal effects on hCMEC/D3 cells until co-administered with bacterial lipopolysaccharide, which then prevented endotoxin-induced permeability. Further analysis suggested that pCG acts as an antagonist to the primary lipopolysaccharide receptor, TLR4. This highlights pCG’s biological activity at the BBB, countering the effects of circulating lipopolysaccharide. These findings challenge the perception of glucuronide conjugates as mere metabolic byproducts and underscore the intricate communication pathways between gut microbes and the host, particularly in the gut–brain axis [84].
4. Oligodendrocytes: Guardians of Axial Integrity in the CNS Microbial Communication
Oligodendrocytes, primarily known for their crucial role in myelinating axons to facilitate efficient nerve signal transmission, have recently emerged as intriguing players in the intricate dialogue between the CNS and microbial entities [85,86]. While the field is still evolving, studies suggest that oligodendrocytes may participate in CNS–microbiome communication through mechanisms that extend beyond their classical functions.
4.1. Oligodendrocyte Sensing of Bacterial Signals
Oligodendrocytes express a variety of receptors, including toll-like receptors (TLRs) and other pattern recognition receptors (PRRs). These receptors enable oligodendrocytes to detect bacterial signals, such as pathogen-associated molecular patterns (PAMPs), present on the surface of microbes [46]. Upon recognition of bacterial signals, oligodendrocytes may initiate intracellular signaling cascades that could influence their function and interactions with neighboring cells.
4.2. Oligodendrocytes in the Crosstalk in the Microbiome–Brain Axis
Myelin, the fatty substance produced by oligodendrocytes, plays a critical role in insulating axons and facilitating rapid electrical signal transmission. Studies suggest that bacterial signals may modulate oligodendrocyte function, impacting myelin maintenance and integrity. Dysregulation in myelin maintenance due to aberrant responses to microbial signals could potentially contribute to demyelinating disorders or exacerbate existing conditions.
The prefrontal cortex (PFC), located in the frontal lobe, regulates behavior and social interaction. This part of the brain undergoes extended myelination, which is crucial for its role [87]. Studies have shown that socially withdrawn mice exhibit impaired myelination in the PFC, accompanied by reduced expression of major myelin genes, a phenomenon which has been attributed to changes in their gut microbial metabolome [88,89].
Specifically, socially withdrawn mice have been shown to exhibit shifts in their gut microbiome, leading to increased levels of p-cresol, a product of microbial breakdown of dietary tyrosine. P-cresol, known to inhibit oligodendrocyte differentiation in vitro, has been shown to be implicated in the observed myelination deficits [88]. However, p-cresol is not the sole microbial metabolite influencing myelination and behavior. Other studies have explored the role of butyrate, a SCFA produced by gut microbiota through dietary fiber fermentation, in regulating microglial activity [73]. Butyrate, which shares similarities with valproic acid—a mood stabilizing drug known to inhibit oligodendrocyte differentiation—is suggested to directly influence oligodendrocyte progenitor cell (OPC) differentiation and myelination due to its histone deacetylase inhibitory activity [85].
During early life, rapid myelination is crucial, and it has been shown that neonatal gut dysbiosis can disrupt this process by affecting immune responses and neuronal differentiation. Keogh et al. [31] demonstrated that neonatal antibiotic-induced dysbiosis disrupts host–microbe interactions, impairing brain myelination and altering the gut–brain axis. They found that, when neonatal C57BL/6 mice were given antibiotics or water daily from postnatal day 7 until weaning to induce dysbiosis, dysbiosis persisted in adulthood, affecting intestinal physiology and bacterial metabolites. They reported that this led to cognitive deficits and anxiety-like behavior. Gene expression related to myelination and the fact that oligodendrocyte development increased in the prefrontal cortex (PFC) of antibiotic-treated mice were confirmed. Administering the short-chain fatty acid butyrate post-antibiotic treatment restored intestinal physiology, behavior, and myelination, indicating the crucial role of gut microbiota [31].
Not only has the effect of the microbiome in neural myelination been documented, but also the role of local myelination on the microbial composition of the gut has been reported. Bostancıklıoğlu and colleagues explored the correlation between myelin biology and gut microbiome profile [32]. Using 16S rRNA metagenomic sequencing, molecular analysis of myelin biology-associated proteins and behavioral tests, they found that hippocampal myelin damage led to significant gut dysbiosis (p < 0.05), altered memory performance (p < 0.05), and affected emotional responses. Treatment with clemastine (promoting myelin repair) improved gut dysbiosis and behavioral deviations. This research provides animal-based evidence of the direct interaction between glial biology in the hippocampus and gut microbiome profile, suggesting a framework for generating new hypotheses connecting different systems to the gut–brain axis [32,90].
These findings underscore the significance of the microbiome–gut–behavior axis in modulating myelination and social behaviors, highlighting microbial metabolites as critical mediators in gut-microbiota–oligodendrocyte communication.
4.3. Crosstalk among Oligodendrocytes, Microglia, and Microorganisms
Oligodendrocytes are in close proximity to microglia, the resident immune cells of the CNS. Growing evidence demonstrates the role that microglia play in the regulation of myelination by oligodendrocytes [40,91]. Microbial signals can effectively activate microglial cells, and this activation may influence oligodendrocyte function. The crosstalk between oligodendrocytes and microglia could involve immune modulation, where the release of cytokines and other signaling molecules by microglia influences the responses of oligodendrocytes to microbial signals.
4.4. Microbe–Oligodendrocyte Interactions in Neurodegenerative Diseases
The potential impact of oligodendrocyte–microbe crosstalk on neurodegenerative diseases is an area that warrants further exploration. Neurodegenerative disorders, such as multiple sclerosis (MS), involve demyelination and axonal damage [92,93].
Understanding how oligodendrocytes respond to microbial signals may offer insights into the pathogenesis of these diseases and open new avenues for therapeutic interventions aimed at preserving myelin integrity.
Oligodendrocytes not only contribute to myelination, but also play a role in supporting axonal health and integrity. It is conceivable that microbial signals may influence the trophic support provided by oligodendrocytes to axons. Investigating the impact of microbial cues on oligodendrocyte-mediated support for axonal health may unveil novel mechanisms relevant to neuroprotection and disease progression.
5. Neurotransmitter Modulation in Glia–Microbe Interactions
Microorganisms, particularly those residing in the gut, can produce neurotransmitter-like molecules or precursors that impact neurotransmitter levels in the CNS. Bacteria have demonstrated the ability to generate and/or metabolize various mammalian neurotransmitters such as dopamine, norepinephrine, serotonin, and gamma-aminobutyric acid (GABA) [11]. Neuroglial cells, including astrocytes, are equipped with receptors for these molecules, allowing them to potentially sense and respond to microbial signals [94,95,96]. The modulation of neurotransmitter levels by microorganisms may influence synaptic transmission, neuronal excitability, and, ultimately, cognitive and behavioral outcomes which, basically, are regulated by neuroglial cells. While there is growing interest in understanding the interaction of the neurotransmitters that microorganisms produce with the human nervous system, the pathways and molecular mechanisms of these interactions remain to be elucidated. Much of our understanding of microbial neuroactive compound production and their impact on brain function stems from animal studies. However, there is a need to broaden our knowledge and understanding of the interaction between neuroglia and microorganisms in human health and disease.
For example, a neurotransmitter has been found in the roundworm Caenorhabditis elegans, called tyramine, which is produced by gut-dwelling Providencia bacteria [97]. It influences the worm’s sensory choices independently of the worm’s own tyramine production. The bacteria-produced tyramine is likely transformed into octopamine by a host enzyme called tyramine β-hydroxylase. Octopamine then interacts with the OCTR-1 octopamine receptor on specific sensory neurons, affecting the worm’s response to unpleasant smells. The researchers pinpointed the genes responsible for tyramine production in Providencia and found that these genes play a crucial role in influencing the worm’s behavior. Moreover, they observed that C. elegans preferentially selects Providencia bacteria when given a choice of food sources, and this preference relies on the presence of bacterially produced tyramine and the worm’s own octopamine signaling [97]. Various glial cells in C. elegans play specific roles in modulating neuron-based activity and behavior [98]. Cephalic sheath glia are essential for the development, maintenance, and activity of central synapses. Meanwhile, amphid glia are implicated in regulating the sensitivity of sensory synapses. These glial cell types originate from the ectoderm. Additionally, mesoderm-derived glial-like cells in the nerve ring (GLRs) are involved in the circuitry responsible for initiating motor movement in the anterior region of the worm [98].
6. Implications for Neurological Disorders
Understanding the glial-mediated communication between the central nervous system (CNS) and microorganisms, specifically members of our microbiome, has far-reaching implications for the general health and progression of various neurological disorders. Dysregulation of these intricate pathways may contribute to the development and exacerbation of conditions spanning neurodegenerative diseases, mood disorders, and neuroinflammatory conditions [6,8,12,27,31,99,100,101,102]. Here, we delve into specific examples that underscore the role of glial cells and potential microbial influences in these disorders. Glial cells, particularly microglia and astrocytes, play crucial roles in the progression of neurodegenerative diseases such as Alzheimer’s, Parkinson’s, and Huntington’s diseases [43,103,104,105].
Alzheimer’s disease (AD) is a progressive neurodegenerative disease that is characterized, pathologically, by the development of amyloid beta (Aβ) plaques and neurofibrillary tau tangles in the brain [104,106,107]. Research indicates that altering the gut microbiome with antibiotics (abx) reduces amyloid beta (Aβ) plaques and diminishes the proinflammatory microglial response in male APPPS1-21 mice. Chandra et al. [103] revealed that the gut microbiome influences how astrocytes respond to Aβ amyloidosis through mechanisms involving both microglia and other pathways. Their study demonstrated that treating male APPPS1-21 mice with broad-spectrum antibiotics after birth, leading to gut microbiome disruption, decreases the presence of GFAP+ reactive astrocytes and PAAs. This suggests that the gut microbiome plays a role in regulating the induction and recruitment of astrocytes reactive to Aβ plaques [103].
Parkinson’s disease (PD) stands as the second most prevalent neurodegenerative condition globally, marked by the degeneration of dopaminergic neurons in the substantia nigra pars compacta [108,109]. Increasing evidence suggests that oxidative and nitrosative stress in astrocytes contributes significantly to PD development, a phenomenon which can intensify in response to microbial signals. Given their pivotal function in generating antioxidants and neutralizing reactive oxygen and nitrogen species (ROS/RNS), astrocytic oxidative/nitrosative stress emerges as a pivotal factor in the onset of PD [28].
Pioneering studies have revealed the potential role of gut microbiome dysbiosis as a disease biomarker and modifier in PD patients. Since Scheperjans et al.’s initial report on gut microbiota alterations in PD in 2015, subsequent research, including studies by Aho et al., Li et al., Lin et al., and Petrov et al., has further supported this link [105,106,107,108]. However, how this microbiome dysbiosis draws its regulatory impact on astrocytic oxidative/nitrosative stress remains to be elucidated.
Microglial activation, often triggered by microbial components or dysbiosis-related inflammation, has been implicated in the chronic neuroinflammation characteristic of these conditions [110]. Additionally, astrocytes, responding to microbial signals, may contribute to the disruption of synaptic homeostasis and to the accumulation of misfolded proteins seen in these diseases. Exploring the specific neuroglial pathways influenced by microbial factors may unveil novel targets for therapeutic interventions in neurodegenerative disorders.
7. Implications for Health and Disease
Perturbations in the microbial ecosystem, commonly observed in dysbiosis or microbial imbalances, have been shown to impact neuroglial function and contribute to neurological disorders. Understanding the ecological aspects of glial–microbial interactions provides a holistic perspective on the maintenance of neural health. Therapeutic interventions aimed at restoring microbial balance could potentially influence neuroglial responses, offering novel strategies for managing neurological disorders influenced by dysregulated neural–microbial communication.
Incorporating the evolutionary and ecological dimensions of microorganisms’ impacts on the human nervous system, including the vital role of microglia, astrocytes, and oligodendrocytes, enriches our understanding of the intricate relationship between the microbial world and the evolution of the human brain. This broader perspective opens avenues for interdisciplinary research at the intersection of microbiology, neuroscience, and evolutionary biology, fostering insights into the complex dynamics shaping brain health and disease.
8. Future Directions
The field of glia–-microbiome crosstalk is rapidly evolving, and future research should delve into the intricacies of the modulatory effects exerted by microorganisms on the CNS through neuroglial cells. Elucidating the molecular mechanisms, identifying specific microbial components involved, and understanding the functional consequences of this bidirectional communication will be crucial for developing targeted interventions for neurological disorders. Elucidating the mechanisms and consequences of this crosstalk holds promise for developing novel therapeutic strategies for neurological disorders and advancing our understanding of the dynamic interplay between the nervous system and the microbiome.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Carson, M.J.; Doose, J.M.; Melchior, B.; Schmid, C.D.; Ploix, C.C. CNS Immune Privilege: Hiding in Plain Sight. Immunol. Rev. 2006, 213, 48–65. [Google Scholar] [CrossRef]
- Parpura, V.; Basarsky, T.A.; Liu, F.; Jeftinija, K.K.; Jeftinija, S.; Haydon, P.G. Glutamate-Mediated Astrocyte-Neuron Signalling. Lett. Nat. 1994, 369, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.S.; Xu, Q.; Arcuino, G.; Kang, J.; Nedergaard, M. Astrocyte-Mediated Activation of Neuronal Kainate Receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 3172–3177. [Google Scholar] [CrossRef] [PubMed]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.A.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins Are Astrocyte-Secreted Proteins That Promote CNS Synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Kucukdereli, H.; Allen, N.J.; Lee, A.T.; Feng, A.; Ozlu, M.I.; Conatser, L.M.; Chakraborty, C.; Workman, G.; Weaver, M.; Sage, E.H.; et al. Control of Excitatory CNS Synaptogenesis by Astrocyte-Secreted Proteins Hevin and SPARC. Proc. Natl. Acad. Sci. USA 2011, 108, E440–E449. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Bose, C.; Mande, S.S. Tryptophan Metabolism by Gut Microbiome and Gut-Brain-Axis: An in Silico Analysis. Front. Neurosci. 2019, 13, 1365. [Google Scholar] [CrossRef] [PubMed]
- Sanmarco, L.M.; Wheeler, M.A.; Gutiérrez-Vázquez, C.; Polonio, C.M.; Linnerbauer, M.; Pinho-Ribeiro, F.A.; Li, Z.; Giovannoni, F.; Batterman, K.V.; Scalisi, G.; et al. Gut-Licensed IFNγ+ NK Cells Drive LAMP1+TRAIL+Anti-Inflammatory Astrocytes. Nature 2021, 590, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Gudkov, S.V.; Burmistrov, D.E.; Kondakova, E.V.; Sarimov, R.M.; Yarkov, R.S.; Franceschi, C.; Vedunova, M.V. An Emerging Role of Astrocytes in Aging/Neuroinflammation and Gut-Brain Axis with Consequences on Sleep and Sleep Disorders. Ageing Res. Rev. 2023, 83, 101775. [Google Scholar] [CrossRef] [PubMed]
- Barroso, A.; Mahler, J.V.; Fonseca-Castro, P.H.; Quintana, F.J. The Aryl Hydrocarbon Receptor and the Gut–Brain Axis. Cell Mol. Immunol. 2021, 18, 259–268. [Google Scholar] [CrossRef]
- Zhou, L.; Foster, J.A. Psychobiotics and the Gut–Brain Axis: In the Pursuit of Happiness. Neuropsychiatr. Dis. Treat. 2015, 11, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter Modulation by the Gut Microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. How Gut Microbes Could Drive Brain Disorders. Nature 2021, 590, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Cryan, J.F.; Paul Ross, R.; Fitzgerald, G.F.; Dinan, T.G.; Stanton, C. Bacterial Neuroactive Compounds Produced by Psychobiotics. Adv. Exp. Med. Biol. 2014, 817, 221–239. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria–Gut–Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Barres, B.A. Glia—More than Just Brain Glue. Nature 2009, 457, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Barres, B.A. Signaling between Glia and Neurons: Focus on Synaptic Plasticity. Curr. Opin. Neurobiol. 2005, 15, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Ransom, B.; Goldman, S.A. New Roles for Astrocytes: Redefining the Functional Architecture of the Brain. Trends Neurosci. 2003, 26, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.-S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Rothhammer, V.; Kenison, J.E.; Li, Z.; Tjon, E.; Takenaka, M.C.; Chao, C.C.; Alves De Lima, K.; Borucki, D.M.; Kaye, J.; Quintana, F.J. Aryl Hydrocarbon Receptor Activation in Astrocytes by Laquinimod Ameliorates Autoimmune Inflammation in the CNS. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e946. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Lim, S.; Hoeffel, G.; Low, D.; Huber, T. Origin and Differentiation of Microglia. Front. Cell Neurosci. 2013, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Brioschi, S.; Zhou, Y.; Colonna, M. Brain Macrophages in Development, Homeostasis and Disease. J. Immunol. 2020, 204, 294. [Google Scholar] [CrossRef] [PubMed]
- Tizabi, Y.; Getachew, B.; Tsytsarev, V.; Csoka, A.B.; Copeland, R.L.; Heinbockel, T.; Tizabi, Y.; Getachew, B.; Tsytsarev, V.; Csoka, A.B.; et al. Central Nicotinic and Muscarinic Receptors in Health and Disease. In Acetylcholine—Recent Advances and New Perspectives; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Del Bigio, M.R. Ependymal Cells: Biology and Pathology. Acta Neuropathol. 2010, 119, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Götz, M. Radial Glia: Multi-Purpose Cells for Vertebrate Brain Development. Trends Neurosci. 2002, 25, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Weissman, T.A.; Riquelme, P.A.; Ivic, L.; Flint, A.C.; Kriegstein, A.R. Calcium Waves Propagate through Radial Glial Cells and Modulate Proliferation in the Developing Neocortex. Neuron 2004, 43, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-F.; Wei, D.-N.; Tang, Y. Gut Microbiota Regulate Astrocytic Functions in the Brain: Possible Therapeutic Consequences. Curr. Neuropharmacol. 2021, 19, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Rizor, A.; Pajarillo, E.; Johnson, J.; Aschner, M.; Lee, E. Astrocytic Oxidative/Nitrosative Stress Contributes to Parkinson’s Disease Pathogenesis: The Dual Role of Reactive Astrocytes. Antioxidants 2019, 8, 265. [Google Scholar] [CrossRef]
- Burmeister, A.R.; Johnson, M.B.; Marriott, I. Murine Astrocytes Are Responsive to the Pro-Inflammatory Effects of IL-20. Neurosci. Lett. 2019, 708, 134334. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Keogh, C.E.; Kim, D.H.J.; Pusceddu, M.M.; Knotts, T.A.; Rabasa, G.; Sladek, J.A.; Hsieh, M.T.; Honeycutt, M.; Brust-Mascher, I.; Barboza, M.; et al. Myelin as a Regulator of Development of the Microbiota-Gut-Brain Axis. Brain Behav. Immun. 2021, 91, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Bostancıklıoğlu, M.; Kaplan, D.S.; Temiz, E.; Yiğit, E. Local Myelin Damage in the Hippocampus Fluctuates Gut Microbiome Profile and Memory. J. Psychiatr. Res. 2023, 158, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Bradl, M.; Lassmann, H. Oligodendrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Wang, X.; Wu, H.; Wu, Q.; Zhang, J. Microglia and Neuroinflammation: Crucial Pathological Mechanisms in Traumatic Brain Injury-Induced Neurodegeneration. Front. Aging Neurosci. 2022, 14, 825086. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, V.L.; Nikonenko, I.R.; Skibo, G.G.; McKanna, J.A. Distribution of Microglia and Astrocytes in Different Regions of the Normal Adult Rat Brain. Neurophysiology 1997, 29, 343–351. [Google Scholar] [CrossRef]
- Yang, T.T.; Lin, C.; Hsu, C.T.; Wang, T.F.; Ke, F.Y.; Kuo, Y.M. Differential Distribution and Activation of Microglia in the Brain of Male C57BL/6J Mice. Brain Struct. Funct. 2013, 218, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Satoh, J.I. Gene Expression Profiles of M1 and M2 Microglia Characterized by Comparative Analysis of Public Datasets. Clin. Exp. Neuroimmunol. 2018, 9, 124–138. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, J.A.; Kavanagh, E.; Engskog-Vlachos, P.; Engskog, M.K.R.; Herrera, A.J.; Espinosa-Oliva, A.M.; Joseph, B.; Hajji, N.; Venero, J.L.; Burguillos, M.A. Cells Microglia: Agents of the CNS Pro-Inflammatory Response. Cells 2020, 9, 1717. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 Polarization and Metabolic States. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.N.; Douglas Fields, R. Regulation of Myelination by Microglia. Sci. Adv. 2021, 7, 1131. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, J.; Zhang, Y.; Huang, Y.; Chen, D.; Shi, Z.; Smith, A.D.; Li, W.; Gao, Y. Central Nervous System Diseases Related to Pathological Microglial Phagocytosis. CNS Neurosci. Ther. 2021, 27, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Fiebich, B.L.; Ribeiro, C.; Batista, A.; Saliba, S.W.; Yousif, N.M.; Pinheiro De Oliveira, A.C.; Burguillos, M.A.; Airavaara, M.; Carlos, A.; De Oliveira, P. Role of Microglia TLRs in Neurodegeneration. Front. Cell. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef]
- Palpagama, T.H.; Waldvogel, H.J.; Faull, R.L.M.; Kwakowsky, A. The Role of Microglia and Astrocytes in Huntington’s Disease. Front. Mol. Neurosci. 2019, 12, 473145. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, L.A.; Di Gioia, M.; Amarante-Mendes, G.P.; Br, G.; Weinlich, R.; Bortoluci, K.R.; Adjemian, S.; Branco, L.M.; Zanetti, L.C. Pattern Recognition Receptors and the Host Cell Death Molecular Machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Pablo, J.; Vaccari, R.; Dalton Dietrich, W.; Popovich, P.G.; Keane, R.W. Pattern Recognition Receptors and Central Nervous System Repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2015, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, J.; Duan, L.; Xiong, H.; Jiang, Y.; Liang, H. Microglia Activation Mediated by Toll-like Receptor-4 Impairs Brain White Matter Tracts in Rats. J. Biomed. Res. 2018, 32, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; De Angelis, A.L.H.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Falsig, J.; Van Beek, J.; Hermann, C.; Leist, M. Review Molecular Basis for Detection of Invading Pathogens in the Brain. J. Neurosci. Res. 2007, 86, 1434–1447. [Google Scholar] [CrossRef]
- Hori, O.; Brett, J.; Slattery, T.; Cao, R.; Zhang, J.; Chen, J.X.; Nagashima, M.; Lundh, E.R.; Vijay, S.; Nitecki, D.; et al. The Receptor for Advanced Glycation End Products (RAGE) Is a Cellular Binding Site for Amphoterin. J. Biol. Chem. 1995, 270, 25752–25761. [Google Scholar] [CrossRef]
- Figdor, C.G.; van Kooyk, Y.; Adema Gosse, J. C-Type Lectin Receptors on Dendritic Cells and Langerhans Cells. Nat. Rev. Immunol. 2002, 2, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Choi, H.; Prabagar, M.G.V.; Choi, W.S.; Kim, S.J.; Cheong, C.; Park, C.G.; Chin, C.Y.; Kang, Y.S. The C-Type Lectin CD209b Is Expressed on Microglia and It Mediates the Uptake of Capsular Polysaccharides of Streptococcus Pneumoniae. Neurosci. Lett. 2009, 450, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, J.; Chiu, I.; Wang, Y.; Sloane, J.A.; Lü, J.; Kosaras, B.; Sidman, R.L.; Volpe, J.J.; Vartanian, T. Toll-like Receptor 8 Functions as a Negative Regulator of Neurite Outgrowth and Inducer of Neuronal Apoptosis. J. Cell Biol. 2006, 175, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, I.E.; Lewen, A.; Galow, L.V.; Cesetti, T.; Scheffel, J.; Regen, T.; Hanisch, U.K.; Kann, O. TLR4-Activated Microglia Require IFN-γ to Induce Severe Neuronal Dysfunction and Death in Situ. Proc. Natl. Acad. Sci. USA 2016, 113, 212–217. [Google Scholar] [CrossRef]
- Ebert, S.; Gerber, J.; Bader, S.; Mühlhauser, F.; Brechtel, K.; Mitchell, T.J.; Nau, R. Dose-Dependent Activation of Microglial Cells by Toll-like Receptor Agonists Alone and in Combination. J. Neuroimmunol. 2005, 159, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.M.; Rodríguez, J.; Giambartolomei, G.H. Microglia at the Crossroads of Pathogen-Induced Neuroinflammation. ASN Neuro 2022, 14, 17590914221104566. [Google Scholar] [CrossRef] [PubMed]
- Machado-Pereira, M.; Santos, T.; Ferreira, L.; Bernardino, L.; Ferreira, R. Anti-Inflammatory Strategy for M2 Microglial Polarization Using Retinoic Acid-Loaded Nanoparticles. Mediat. Inflamm. 2017, 2017, 6742427. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Dong, Z.; Jiang, X.; Qu, L.; Zhou, W.; Sun, X.; Hou, J.; Xu, H.; Cheng, M. Gut Microbiota Taxon-Dependent Transformation of Microglial M1/M2 Phenotypes Underlying Mechanisms of Spatial Learning and Memory Impairment after Chronic Methamphetamine Exposure. Microbiol. Spectr. 2023, 11, e0030223. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Yang, T.; Oliveira, A.C.; Lobaton, G.O.; Aquino, V.; Kim, S.; Richards, E.M.; Pepine, C.J.; Sumners, C.; Raizada, M.K. Microglial Cells Impact Gut Microbiota and Gut Pathology in Angiotensin II-Induced Hypertension. Circ. Res. 2019, 124, 727–736. [Google Scholar] [CrossRef]
- Mossad, O.; Batut, B.; Yilmaz, B.; Dokalis, N.; Mezö, C.; Nent, E.; Nabavi, L.S.; Mayer, M.; Maron, F.J.M.; Buescher, J.M.; et al. Gut Microbiota Drives Age-Related Oxidative Stress and Mitochondrial Damage in Microglia via the Metabolite N 6-Carboxymethyllysine. Nat. Neurosci. 2022, 25, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Barres, B.A. A Smarter Mouse with Human Astrocytes. BioEssays 2013, 35, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaee, M.; Menard, F. Glutamate Signaling and Filopodiagenesis of Astrocytoma Cells in Brain Cancers: Survey and Questions. Cells 2022, 11, 2657. [Google Scholar] [CrossRef]
- Bernardinelli, Y.; Muller, D.; Nikonenko, I. Astrocyte-Synapse Structural Plasticity. Neural Plast. 2014, 2014, 232105. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; Mascanfroni, I.D.; Bunse, L.; Takenaka, M.C.; Kenison, J.E.; Mayo, L.; Chao, C.C.; Patel, B.; Yan, R.; Blain, M.; et al. Type i Interferons and Microbial Metabolites of Tryptophan Modulate Astrocyte Activity and Central Nervous System Inflammation via the Aryl Hydrocarbon Receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaee, M. Glutamate Induced Morphological Response in Astrocytoma Cells. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2021. [Google Scholar] [CrossRef]
- Tabatabaee, M.S.; Menard, F. L-Type Voltage-Gated Calcium Channel Modulators Inhibit Glutamate-Induced Morphology Changes in U118-MG Astrocytoma Cells. Cell Mol. Neurobiol. 2020, 40, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Willard, S.S.; Koochekpour, S. Glutamate, Glutamate Receptors, and Downstream Signaling Pathways. Int. J. Biol. Sci. 2013, 9, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Malarkey, E.B.; Parpura, V. Mechanisms of Glutamate Release from Astrocytes. Neurochem. Int. 2008, 52, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Astrocyte Reactivity: Subtypes, States, and Functions in CNS Innate Immunity. Trends Immunol. 2020, 41, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.M.; Sun, M.F.; Jia, X.B.; Li, Y.; Zhang, B.P.; Zhao, L.P.; Shi, Y.; Zhou, Z.L.; Zhu, Y.L.; Cui, C.; et al. Sodium Butyrate Exacerbates Parkinson’s Disease by Aggravating Neuroinflammation and Colonic Inflammation in MPTP-Induced Mice Model. Neurochem. Res. 2020, 45, 2128–2142. [Google Scholar] [CrossRef] [PubMed]
- Huuskonen, J.; Suuronen, T.; Nuutinen, T.; Kyrylenko, S.; Salminen, A. Regulation of Microglial Inflammatory Response by Sodium Butyrate and Short-Chain Fatty Acids. Br. J. Pharmacol. 2004, 141, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Dehhaghi, M.; Tan, V.; Heng, B.; Mohammadipanah, F.; Guillemin, G.J. Protective Effects of Myxobacterial Extracts on Hydrogen Peroxide-Induced Toxicity on Human Primary Astrocytes. Neuroscience 2019, 399, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cooley, I.D.; Chauhan, V.S.; Donneyz, M.A.; Marriott, I. Astrocytes Produce IL-19 in Response to Bacterial Challenge and Are Sensitive to the Immunosuppressive Effects of This IL-10 Family Member. Glia 2014, 62, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.R.; Johnson, M.B.; Yaemmongkol, J.J.; Marriott, I. Murine Astrocytes Produce IL-24 and Are Susceptible to the Immunosuppressive Effects of This Cytokine. J. Neuroinflamm. 2019, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-Y.; Chang, M.-S. IL-20 Is Regulated by Hypoxia-Inducible Factor and up-Regulated after Experimental Ischemic Stroke. J. Immunol. 2009, 182, 5003–5012. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, T.; Wada, S.; Suzuki, S.; Okuma, Y.; Akira, S.; Matsuda, T.; Nomura, Y. Bacterial Endotoxin Induces IL-20 Expression in the Glial Cells. Mol. Brain Res. 2004, 130, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Heithoff, B.P.; George, K.K.; Phares, A.N.; Zuidhoek, I.A.; Munoz-Ballester, C.; Robel, S. Astrocytes Are Necessary for Blood–Brain Barrier Maintenance in the Adult Mouse Brain. Glia 2021, 69, 436–472. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte–Endothelial Interactions at the Blood–Brain Barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Sterka, D.; Rati, D.M.; Marriott, I. Functional Expression of NOD2, a Novel Pattern Recognition Receptor for Bacterial Motifs, in Primary Murine Astrocytes. Glia 2006, 53, 322–330. [Google Scholar] [CrossRef]
- Wang, J.; Li, G.; Wang, Z.; Zhang, X.; Yao, L.; Wang, F.; Liu, S.; Yin, J.; Ling, E.A.; Wang, L.; et al. High Glucose-Induced Expression of Inflammatory Cytokines and Reactive Oxygen Species in Cultured Astrocytes. Neuroscience 2012, 202, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Wolburg, H.; Neuhaus, J.; Kniesel, U.; Krauß, B.; Schmid, E.M.; Öcalan, M.; Farrell, C.; Risau, W. Modulation of Tight Junction Structure in Blood-Brain Barrier Endothelial Cells Effects of Tissue Culture, Second Messengers and Cocultured Astrocytes. J. Cell Sci. 1994, 107, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Stachulski, A.V.; Knausenberger, T.B.A.; Shah, S.N.; Hoyles, L.; McArthur, S. A Host–Gut Microbial Amino Acid Co-Metabolite, p-Cresol Glucuronide, Promotes Blood–Brain Barrier Integrity in Vivo. Tissue Barriers 2023, 11, 2073175. [Google Scholar] [CrossRef] [PubMed]
- Ntranos, A.; Casaccia, P. The Microbiome–Gut–Behavior Axis: Crosstalk Between the Gut Microbiome and Oligodendrocytes Modulates Behavioral Responses. Neurotherapeutics 2018, 15, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef] [PubMed]
- Franklin, T.B.; Silva, B.A.; Perova, Z.; Marrone, L.; Masferrer, M.E.; Zhan, Y.; Kaplan, A.; Greetham, L.; Verrechia, V.; Halman, A.; et al. Prefrontal Cortical Control of a Brainstem Social Behavior Circuit. Nat. Neurosci. 2017, 20, 260. [Google Scholar] [CrossRef] [PubMed]
- Hoban, A.E.; Stilling, R.M.; Ryan, F.J.; Shanahan, F.; Dinan, T.G.; Claesson, M.J.; Clarke, G.; Cryan, J.F. Regulation of Prefrontal Cortex Myelination by the Microbiota. Transl. Psychiatry 2016, 6, e774. [Google Scholar] [CrossRef]
- Gacias, M.; Gaspari, S.; Santos, P.M.G.; Tamburini, S.; Andrade, M.; Zhang, F.; Shen, N.; Tolstikov, V.; Kiebish, M.A.; Dupree, J.L.; et al. Microbiota-Driven Transcriptional Changes in Prefrontal Cortex Override Genetic Differences in Social Behavior. Elife 2016, 5, e13442. [Google Scholar] [CrossRef] [PubMed]
- Myatich, A.; Haque, A.; Sole, C.; Banik, N.L. Clemastine in Remyelination and Protection of Neurons and Skeletal Muscle after Spinal Cord Injury. Neural Regen. Res. 2023, 18, 940. [Google Scholar] [CrossRef] [PubMed]
- Kalafatakis, I.; Karagogeos, D. Oligodendrocytes and Microglia: Key Players in Myelin Development, Damage and Repair. Biomolecules 2021, 11, 1058. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Waubant, E. The Multiple Sclerosis Microbiome? Ann. Transl. Med. 2017, 5, 12015. [Google Scholar] [CrossRef] [PubMed]
- Branton, W.G.; Lu, J.Q.; Surette, M.G.; Holt, R.A.; Lind, J.; Laman, J.D.; Power, C. Brain Microbiota Disruption within Inflammatory Demyelinating Lesions in Multiple Sclerosis. Sci. Rep. 2016, 6, 37344. [Google Scholar] [CrossRef] [PubMed]
- Glaum, S.R.; Holzwarth, J.A.; Miller, R.J. Glutamate Receptors Activate Ca2+ Mobilization and Ca2+ Influx into Astrocytes. Proc. Natl. Acad. Sci. USA 1990, 87, 3454–3458. [Google Scholar] [CrossRef]
- Backus, K.H.; Kettenmann, H.; Schachner, M. Pharmacological Characterization of the Glutamate Receptor in Cultured Astrocytes. J. Neurosci. Res. 1989, 22, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.R.; Koji Takahashi, D.; Thomson, K.E.; Wilcox, K.S. The Expression of Kainate Receptor Subunits in Hippocampal Astrocytes After Experimentally Induced Status Epilepticus. J. Neuropathol. Exp. Neurol. 2013, 72, 919–932. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.P.; Fox, B.W.; Chao, P.H.; Schroeder, F.C.; Sengupta, P. A Neurotransmitter Produced by Gut Bacteria Modulates Host Sensory Behaviour. Nature 2020, 583, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Stout, R.F.; Verkhratsky, A.; Parpura, V. Caenorhabditis Elegans Glia Modulate Neuronal Activity and Behavior. Front. Cell Neurosci. 2014, 8, 81214. [Google Scholar] [CrossRef]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Dooling, S.W.; Costa-Mattioli, M. Gut Bacteria Seize Control of the Brain to Prevent Epilepsy. Cell Host Microbe 2018, 24, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Montiel-Castro, A.J.; Gonzalez-Cervantes, R.M.; Bravo-Ruiseco, G.; Pacheco-Lopez, G. The Microbiota-Gut-Brain Axis: Neurobehavioral Correlates, Health and Sociality. Front. Integr. Neurosci. 2013, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, L.; Aceto, S.; Agnisola, C.; De Paolo, S.; Dipineto, L.; Stilling, R.M.; Dinan, T.G.; Cryan, J.F.; Menna, L.F.; Fioretti, A. Probiotic Modulation of the Microbiota-Gut-Brain Axis and Behaviour in Zebrafish. Sci. Rep. 2016, 6, 70. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Di Meco, A.; Dodiya, H.B.; Popovic, J.; Cuddy, L.K.; Weigle, I.Q.; Zhang, X.; Sadleir, K.; Sisodia, S.S.; Vassar, R. The Gut Microbiome Regulates Astrocyte Reaction to Aβ Amyloidosis through Microglial Dependent and Independent Mechanisms. Mol. Neurodegener. 2023, 18, 45. [Google Scholar] [CrossRef]
- Bulgart, H.R.; Neczypor, E.W.; Wold, L.E.; Mackos, A.R. Microbial Involvement in Alzheimer Disease Development and Progression. Mol. Neurodegener. 2020, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Cannon, T.; Gruenheid, S. Microbes and Parkinson’s Disease: From Associations to Mechanisms. Trends Microbiol. 2022, 30, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Hickman, R.A.; Faustin, A.; Wisniewski, T. Alzheimer Disease and Its Growing Epidemic: Risk Factors, Biomarkers, and the Urgent Need for Therapeutics. Neurol. Clin. 2016, 34, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Sheng, M.; Sabatini, B.L.; Südhof, T.C. Synapses and Alzheimer’s Disease. Cold Spring Harb. Perspect. Biol. 2012, 4, a005777. [Google Scholar] [CrossRef] [PubMed]
- Stefanis, L. α-Synuclein in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009399. [Google Scholar] [CrossRef]
- Nunes-Costa, D.; Magalhães, J.D.; G-Fernandes, M.; Cardoso, S.M.; Empadinhas, N. Microbial BMAA and the Pathway for Parkinson’s Disease Neurodegeneration. Front. Aging Neurosci. 2020, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Erin, R.; Reyes, N.; Gao, L.; Asatryan, L. Neuroimmunology and Neuroinflammation Microbiome Meets Microglia in Neuroinflammation and Neurological Disorders. Neurosciences 2020, 7, 215–233. [Google Scholar] [CrossRef]
Figure 1.
Overview of Neuroglia. Microglia serve as the immune cells of the brain, engaging in interactions with neurons and neighboring blood vessels. Oligodendrocytes form myelin sheaths around axons, enhancing neuronal transmission speed. Astrocytes extend processes to interface with blood vessels and encase synapses.
Figure 1.
Overview of Neuroglia. Microglia serve as the immune cells of the brain, engaging in interactions with neurons and neighboring blood vessels. Oligodendrocytes form myelin sheaths around axons, enhancing neuronal transmission speed. Astrocytes extend processes to interface with blood vessels and encase synapses.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tabatabaee, M. Microbiome-Glia Crosstalk: Bridging the Communication Divide in the Central Nervous System. Neuroglia 2024, 5, 89-104. https://doi.org/10.3390/neuroglia5020007
AMA Style
Tabatabaee M. Microbiome-Glia Crosstalk: Bridging the Communication Divide in the Central Nervous System. Neuroglia. 2024; 5(2):89-104. https://doi.org/10.3390/neuroglia5020007
Chicago/Turabian StyleTabatabaee, Mitra. 2024. "Microbiome-Glia Crosstalk: Bridging the Communication Divide in the Central Nervous System" Neuroglia 5, no. 2: 89-104. https://doi.org/10.3390/neuroglia5020007