Biologically Available Phosphorus in Biocrust-Dominated Soils of the Chihuahuan Desert

Department of Biological Sciences, University of Texas at El Paso, El Paso, TX 79968, USA
*
Author to whom correspondence should be addressed.
Soil Syst. 2018, 2(4), 56; https://doi.org/10.3390/soilsystems2040056
Submission received: 10 September 2018
/
Revised: 27 September 2018
/
Accepted: 3 October 2018
/
Published: 10 October 2018
Abstract
:In desert soils, phosphorus (P) cycling is controlled by both geochemical and biological factors and remains less studied than nitrogen and carbon. We examined these P cycling factors in the context of biological soil crusts (biocrusts), which are important drivers of nutrient cycling in drylands and have the potential to release bound labile P. We adopted the biologically-based P (BBP) method, which allows examination of biologically relevant P fractions. The BBP method incorporates four extractions: dilute calcium chloride (CaCl2), citric acid, phosphatase enzymes, and hydrochloric acid (HCl). We coupled the extractions with a 33P-labeled orthophosphate addition and incubation to assess the fate of freshly available phosphate (PO43−). Low P concentrations in the dilute CaCl2 extractions suggest that drylands lack accessible P in the soil solution, while higher amounts in the citric acid- and enzyme-extractable pools suggest that dryland microbes may acquire P through the release of organic acids and phosphatases. The addition of 33PO43− was, within 24 h, quickly adsorbed onto mineral surfaces or incorporated into hydrolysable organic compounds. Areas with biocrusts showed overall lower P concentrations across all four extractable pools. This suggests that biocrust organisms may prevent P adsorption onto mineral surfaces by incorporating P into their biomass. Overall, our results indicate that organisms may have to employ several viable strategies, including organic acid and enzyme production, to access P in dryland soils.
Keywords:
phosphorus; biocrust; dryland; Chihuahuan desert; 33P radiolabel; BBP; Jornada experimental range; New Mexico; LTER1. Introduction
Phosphorus (P) is found in soil systems in a variety of organic and inorganic forms. With no major gas-phase components, P is made available via rock weathering of minerals such as apatite (Ca10(PO4)6(OH,F,Cl)2) [1,2], dust mobilization and deposition [3], or the recycling of organic matter. In drylands, the degree of rock weathering and the level of precipitation are processes that control the dissolution and sorption of P in the soil [4]. Arid soils often have a buildup of soluble minerals such as calcium carbonate (CaCO3) which accumulate as part of caliche horizons at depth, sequestering available (labile) P [2]. Thus, the P supply to organisms in drylands depends on a mineral source from parent material or dust and the chemical means to release that P into the soil solution. Soil organic matter (SOM) concentrations are low in arid soils, which lowers the amount of P available via decomposition by phosphatases. Organic P can make up 80% of the total P in mesic systems [5], while organic P has been found to make up less than 13% of the total P in the Chihuahuan desert [6]. In drylands, P is more likely to be found in mineral or mineral-associated forms than organic forms [7]. Previous studies in the northern Chihuahuan Desert have found HCl-extractable P to be the largest P pool, while directly available P in the soil solution makes up less than 1% of the total P, with the overall P content being relatively low [8,9].
Many insights into dryland P cycling come from the use of the Hedley fractionation method, which was created to characterize the physical and biochemical sources of P [10,11]. This procedure separates P into inorganic and organic P pools that vary in their degree of availability for plants from labile to occluded [12]. The sequential fractionation method first removes inorganic and organic P that is accessible in the soil solution with a resin membrane, followed by an extraction with sodium bicarbonate on the remaining material. The sum of these two fractions represent labile P that is thought to cycle over the short term [9,12,13,14]. The resin-extractable pool characterizes the available P that is directly accessible for plant or microbial uptake in the soil solution. The remaining P fractions are accessed with sodium hydroxide (NaOH), 1 M hydrogen chloride (HCl), concentrated HCl, hydrogen peroxide (H2O2), and concentrated sulfuric acid (H2SO4) to determine moderately labile (non-occluded) to highly recalcitrant forms of P [11]. Hedley fractionation data from the Chihuahuan desert suggest that P does not remain in the soil solution easily; studies have consistently found resin-extractable P to be low overall, making up a small portion of the total P in Chihuahuan desert systems [2,6,8,9]. For arid environments, this suggests that when P is released by the enzymatic breakdown of organically bound phosphorus, for example by the activity of phosphatases or liberated from mineral phases via leaching, it must be quickly taken up by an organism or it ends up in one of the occluded pools.
While Hedley fractionation results suggest that microbial and plant uptake play a role in P availability in the Chihuahuan desert, they provide incomplete information about the mechanisms used by plants and microorganisms to acquire P, such as root interception, acid dissolution, or enzyme hydrolysis [15]. To more fully understand the differences between biological P acquisition strategies that organisms may employ, a method has been introduced by Deluca et al. [15] for assessing biologically-based P pools (BBP). This procedure uses parallel instead of sequential extractions and the four employed extractants mimic strategies used by plants or microbes to access P. The parallel procedures include a (0.1 mmol CaCl2) extraction that determines the dilute salt-extractable pool, which simulates what is available in soil pore water through diffusion or mass flow and directly available for plant and microbial uptake. This pool is similar to the resin-extractable pool used during Hedley fractionation. The citric acid (C6H8O7) and HCl extractants assess two different acid-extractable pools (Table 1). The citric acid-extractable pool assesses P that is sorbed to clay particles or P that is weakly bound to other parts of the soil matrix. This is similar to the bound P assessed in the Hedley fractionation bicarbonate and hydroxide fractions; however, citric acid is thought to be a better analogue for the organic acids released by plant roots and microbes [16]. The HCl extractant assesses P that is strongly bound to mineral surfaces, which may be less accessible to plants and microbes [15]. This stronger acid extractant likely also accesses any P available in the CaCl2 and citric acid extractions. Finally, the phosphatase pool assesses the availability of labile organic P through enzyme hydrolysis [15]. The BBP method does not measure biomass P in the way that, for example, a chloroform fumigation would. However, by including a weak acid similar to organic exudates such as citric acid, oxalic acid, or acetic acid [4] and assessing the phosphatase-available pool consisting of organically bound P, the BBP approach provides a means to more directly illuminate the bioavailable forms of P than what can be achieved with the classical Hedley-type extractions on which the BBP method is built.
There is little information on the size of weak acid and enzyme-extractable P pools in dryland soils, but knowledge about the fate of freshly available PO43− is even more scarce. In mesic systems, the incorporation of P into different soil P fractions has been investigated with the use of radioisotopes [17]. Using either 32P (half-life of 14 days) or 33P (half-life of 25 days), these radioisotope studies have shown that added labeled inorganic P may transform into soil organic P via microbial uptake or become incorporated into occluded forms of P [18,19]. For example, in an agricultural soil fertilized with P, added 33P was stored in more easily assessible pools for plant uptake, while a similar soil with low P content incorporated the 33P into less exchangeable pools [20]. In another study involving a 33P radiotracer, focused on mycorrhizal associations in forested systems, fungal associations increased P uptake from organic matter in comparison to nonmycorrhizal plants [21]. Despite these interesting findings in mesic and agricultural systems, similar studies have not been applied in dryland soils.
In this study, we investigate biologically available forms of P and the fate of freshly added PO43− using dryland soils with and without intact biological soil crusts (biocrusts). We chose to focus on biocrusts because they have been shown to be particularly important for nutrient cycling in arid systems and are a crucial component of most dryland soils [4]. Phosphorus and other nutrients enter dryland soils via dust or litter deposition and weathering of upper soil horizons, resulting in high nutrient concentration on the surface [22,23]. Thus, P bioavailability has been suggested to be more tightly connected to the amount of accessible P that enters the system through surface adsorption than to the release of P from carbonate-bound forms [24]. Organisms found in surface biocrusts such as nitrogen-fixing cyanobacteria, algae, fungi, lichens, and mosses [25] may contribute to the release of unavailable P from surface rock or dust deposits through the release of organic compounds. Cyanobacteria, algae, and fungi have been found to (a) excrete phosphatases that access P through enzyme hydrolysis; (b) secrete organic acids (e.g., citric acid or oxalic acid) that may release P from mineral surfaces; or (c) excrete hydrogen ions (H+) during respiration that induce carbonate dissolution, thereby releasing carbonate-bound P, all of which increase P availability for plants and other soil biota [4,26,27]. For example, dryland cyanobacteria have been shown to aid in the liberation and acquisition of P by mobilizing inorganic P from hydroxyapatite [28]. Once P is in biotic pools, it can be recycled via the decomposition of organically bound P driven by phosphatase enzyme production. Indeed, phosphatase activity has been shown to increase under developed biocrusts compared to areas with bare soil [29], despite phosphatase release being highly controlled by SOM [7]. In addition, biocrusts may aid in the capture of P-containing dust particles and prevent P loss or redistribution through the formation of an extracellular sheath through the excretion of exopolymers that create a sticky surface [4]. Through these processes, concentrated microbial communities may substantially influence P cycling in drylands, creating conditions that are significantly different from areas with disturbed crusts or bare soils.
The aim of this study is to investigate the influence biocrust-dominated soils have on P dynamics in dryland soils that have characteristically low levels of organic matter and low P availability. Using the BBP method coupled with a 33P orthophosphate label addition, we address the following questions: (1) How is biologically available P distributed between different pools in drylands? (2) How does added P become incorporated into the different pools? and (3) How do soils with intact surface crusts affect P cycling in comparison to soils without biocrusts (disturbed crusts)?
2. Materials and Methods
2.1. Site Information and Sample Collection
Samples were collected from areas with intact biocrusts (crusts) and areas of disturbance (no intact crusts) (Figure 1) from a cattle exclosure site within the Jornada experimental range outside Las Cruces, New Mexico (32.5876689, −106.740027) in March of 2018. The site is dominated by black grama (Bouteloua eriopoda), and light cyanobacterial biocrusts mostly consisting of Microcoleus vaginatus [30]. The sampling area within the exclosure was chosen due to its abundance of light crusts in areas with little to no vegetation nearby. The intact and disturbed crust samples were collected from 12 distinct areas (six replicates for each crust type) within the exclosure at least six feet apart in all directions and away from vegetation. The areas of disturbance chosen for sampling previously had intact crusts but have been disturbed due to recent and constant human trampling that began approximately 18 months prior to sample collection. Thus, areas of disturbance had no visible intact crusts.
2.2. Radiolabel Addition and Incubation
Soil cores were collected from the surface at 0–10 cm depth from areas with and without intact surface crusts. Intact crusts made up about 0–2 mm depth within the cores. Samples were sieved using a 2 mm mesh. Any visible plant or animal material in the samples were removed. It is important to note that sieving intact crusts may alter how P is processed in the lab versus in the field. However, the collection and sieving process likely does not kill the crust-associated organisms, which are primarily cyanobacteria. Fresh soil (5 g) was dried for three days at 60 °C for gravimetric water content (average 1.5% moisture), and 20 g fresh soil was subset into 50 mL tubes for radio-label addition and 24-h incubation. Fresh soils were labeled with 1 mL of 0.0384 kBq g−1 orthophosphate (Perkin Elmer Health Sciences, Sheldon, CT, USA) and incubated at 3 °C for 24 h The 24-h incubation period was included to provide time for potential microbial uptake and to assess the biological response of added P across the biologically available P pools analyzed by the four BBP extractants.
2.3. Extractions and P Determination
Following the 24-h incubation, 0.5 g of the radiolabeled sample was subset into 15 mL tubes for the extraction procedure (four vials per sample). Extractions were conducted in parallel by shaking the sample with 10 mL of each of the four extractants—0.01 M CaCl2, 0.01 M citrate, 0.2 enzyme unit phosphatase, and 1 M HCl (Table 1)—for 3 h (200 rev min−1), followed by a centrifugation for 30 min (3020 rev min−1) [15].
Following the four extractions, a 10 mL aliquot of supernatant was placed into a clean 15 mL tube for PO43− determination. If necessary, samples were placed in the fridge (2 days maximum) or frozen for storage prior to analysis. Samples were analyzed colorimetrically (630 nm) using a modified malachite-green method [31] on a Bio-Tek Synergy HT microplate reader (Bio-Tek Inc., Winooski, VT, USA). Samples were adjusted for the dry weight of bulk soil.
Finally, the total P content of each sample was analyzed by borate fusion wavelength dispersion X-ray fluorescence (WD-XRF) spectrometry. Samples were crushed and pulverized and formed into a homogenous glass disk through the fusion of the sample and a borate mixture. The total P procedure and analysis was conducted by SGS Mineral Services (Lakefield, ON, Canada).
2.4. 33P Label Recovery
Following the extractions, an additional 3 mL aliquot of supernatant from each sample with respective extractant was placed in a scintillation vial with 15 mL of scintillation cocktail (Ultima-Gold Scintillation Cocktail, Perkin Elmer) and counted for radioactivity using a 2900 TriCarb Liquid Scintillation Counter at the Biomolecule Analysis Core Facility (BACF) in the UTEP Biological Sciences Department. Samples were counted for 2 min; the counting was repeated three times. The average counts per minute (CPM) and disintegrations per minute (DPM) were determined by the scintillation counter and converted to kBq and used to determine the sample and introduced radioactivity. The label recovery of each sample was determined using the following equation:
where r and R represent the amount of radioactivity or sample activity (kBq kg−1) recovered in each individual and the total applied (introduced) radioactivity in each sample prior to incubation, respectively [32]. This approach in determining 33P recovery may overestimate the amount of P recovered in each individual sample as we did not take into account soil P prior to label addition [33]. Subsampling after the label addition for radioactivity analysis may also result in an overestimation of the 33P recovery. Thus, it is important to note that samples may exceed 100% recovery.
33P Recovery (%) = (r/R) × 100
2.5. Statistics
The BBP technique assesses the relative effectiveness of different biological mechanisms for accessing P. Thus, aside from the raw pool sizes, which we report as mean ± SE, the most important quantitative outcomes of the assay are the proportional magnitudes of the P pool sizes (e.g., the citric acid/phosphatase ratio). To examine the relative sizes of phosphate pools from the BBP fractions and label recovery within each sample across crust type, mean pool sizes from the 12 samples (6 intact; 6 disturbed) were divided by one another (e.g., citric acid mean/CaCl2 mean) to obtain the ratios among pools. These are important because they indicate the relative effectiveness of different P acquisition strategies. For example, if organic acids are a more effective strategy than phosphatases, the citric acid/phosphatase ratio will be high. Similarly, if the HCl/citric acid ratio is high, it would indicate that P is strongly occluded and not able to be released by weak organic acids alone. We have summarized the interpretation of the ratios in Table 2. Uncertainty for these ratios was estimated by generating 95% bootstrap confidence intervals [34]. Bootstrap confidence intervals were calculated using the bias corrected and accelerated (BCa) method [35]. All statistical analyses were conducted using the R Studio Version 1.0.136 [36].
3. Results
3.1. Biologically Available P Pools Across Crust Type
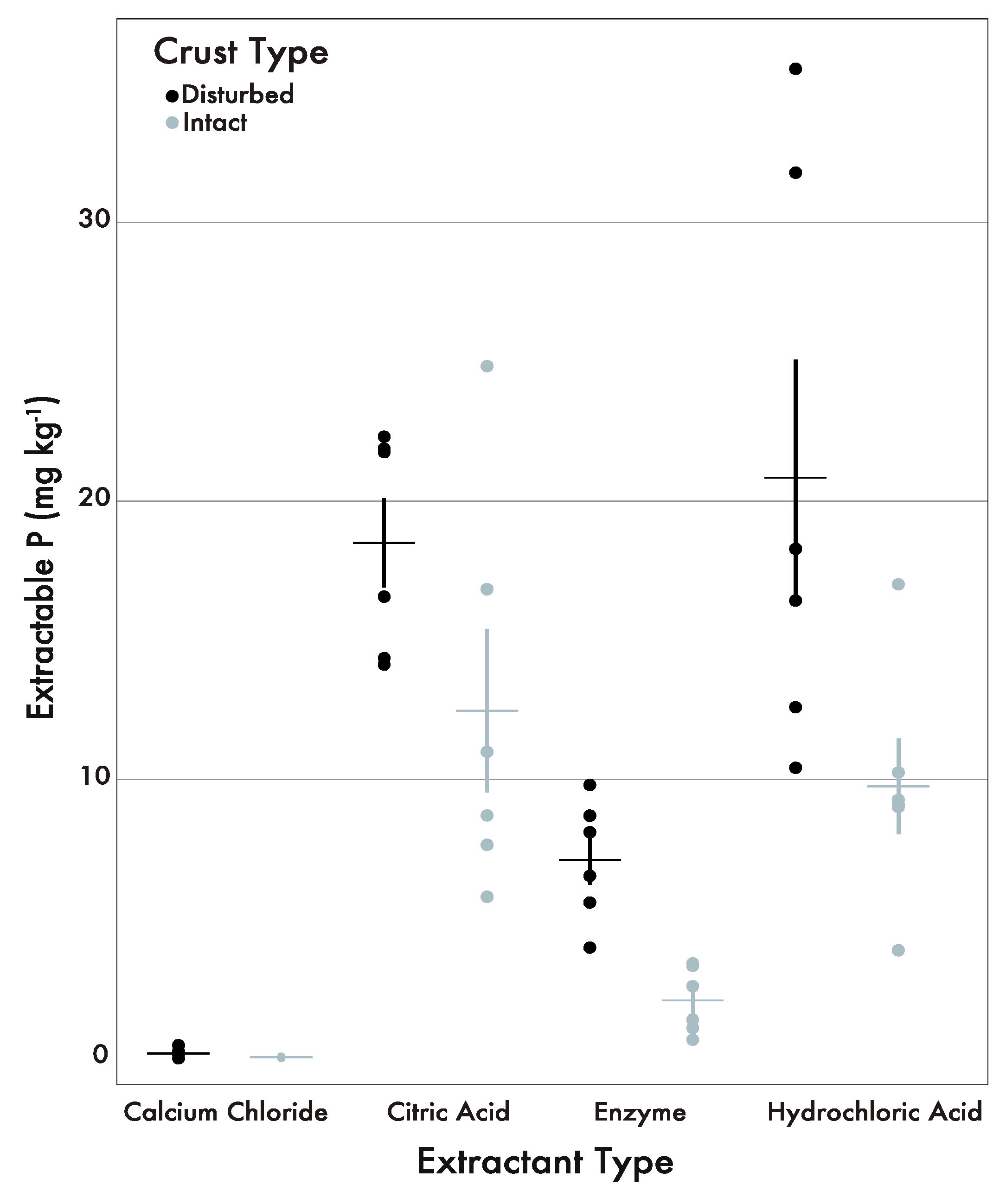
The phosphatase-available pool was lower on average (intact: 2.07 mg kg−1; disturbed: 7.12 mg kg−1) compared to the citric acid-extractable (intact: 12.47 mg kg−1; disturbed: 18.5 mg kg−1) and HCl-extractable (intact: 9.76 mg kg−1; disturbed: 20.84 mg kg−1) pools in intact and disturbed areas (Figure 2). The CaCl2 pool contained very little P in both crust types (intact: 0.03 mg kg−1; disturbed: 0.17 mg kg−1) (Figure 2). Phosphorus concentration in the citric acid, enzyme, and HCl pools were highest in areas of disturbance compared to areas with intact crusts.
The total P content for each of the disturbed and intact crust samples was below the detection limit of 100 mg kg−1 for the borate-fusion WD-XRF technique. Taken together with the results from the P extracts, this indicates that there is no large unaccounted P pool, if any, in the analyzed samples.
Table 3 shows the results of the paired bootstrap statistical analysis for the phosphate concentration by individual sample, crust types and extractant. The citric acid pool was 78% {95% CI = 48, 139} as large as the HCl pool in the intact areas and 113% {79, 144} in the disturbed areas, putting them at a similar magnitude. The citric acid and HCl extractants had the most similar ratios across all comparisons. When compared with the enzyme extractant, the citric acid and HCl ratios revealed comparable ratios between the two crust types (intact: 6.02, 4.71; disturbed: 2.60, 2.93 respectively). Extractant comparisons with CaCl2 were several orders of magnitudes smaller and had a larger degree of variability from comparisons made between the citric acid, enzyme, and HCl extractants (Table 3). The CaCl2 extractant was distinct from the other three extractants and had the least effect on P concentration across individual samples.
3.2. 33P Label Recovery
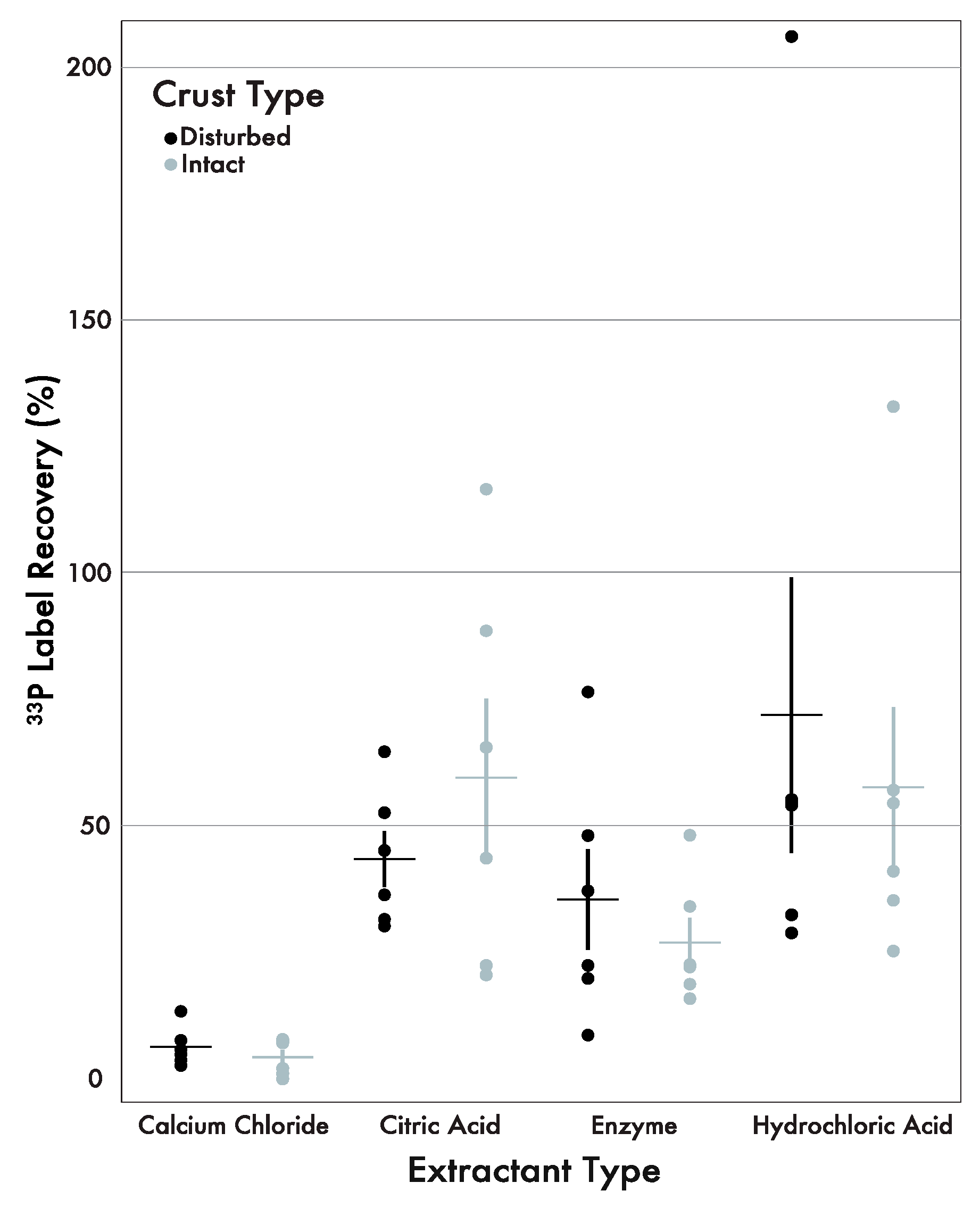
On average, little 33P label was recovered in the CaCl2 extractable pool (5–7%), while significant amounts of label recovery were observed in the citric acid, HCl, and enzyme pools across both crust types (Figure 3; Table 4). The citric acid and HCl pools, in both intact and disturbed areas, showed more P accumulation and label recovery compared to the other two extractants with about 45–70% recovery of the added P. The citric acid pool was the only pool to show increased recovery (15%) in areas with intact crusts compared to areas of disturbance. The enzyme pool showed similar amounts of recovery in both crust types (intact: 27.6%; disturbed: 36.1%).
Table 4 shows the results of the paired bootstrap statistical analysis for the 33P label recovery by individual sample, soil type and extractant. The analysis revealed a similar trend as observed for the P concentration. With a value of 96% {49, 157} the citric acid pool was as large as the HCl pool. These extractants were the most similar across individual samples and crust types compared to the other extractants. When compared with the enzyme extractant, the citric acid and HCl extractants had similar ratios with little variability (intact: 2.18, 2.11; disturbed; 1.22, 2.01 respectively). Unlike the P concentration, the extractant comparisons with CaCl2 revealed overall lower ratios when compared with the citric acid, enzyme, and HCl extractants, along with less variability (Table 4). The CaCl2 extractant had the least effect on 33P label recovery across individual samples.
4. Discussion
4.1. Biologically Available P in Drylands
Consistent with the lower SOM levels in dryland soils, our results show overall low concentrations of extractable P accessed with phosphatase (mean 4.6 mg kg−1) and the dilute CaCl2 extractant (mean 0.10 mg kg−1), compared to concentrations from the citric acid and HCl extractants (mean 15.4 mg kg−1). A BBP study conducted by Deluca et al. [15] in a series of mesic vegetation types across Northern Wales observed, in contrast, larger P concentrations in the soil solution (CaCl2-extractable) and enzyme-extractable P pools, between 2 to 30 mg kg−1 and ~500 mg kg−1 respectively. This contrast suggests that our dryland system maintains relatively low levels of available P that are directly accessible for microbial or plant root uptake via diffusion or transpiration-driven mass flow. We conclude that very little P is directly available in the soil solution, and that most P is surface-bound or incorporated into organic matter in these soils.
Our BBP results show that the highest concentrations of P were in the two acid-extractable pools. We found similar P concentrations using the citric acid and HCl extractants (mean 15.49 mg kg−1 and mean 15.28 mg kg−1, respectively). This similarity suggests that both acid extractants are effective at accessing adsorbed or mineral forms of P [15,37]. The small discrepancy between the detection limit of 100 mg kg−1 for the total P analysis via borate fusion WD-XRF and the HCl extractable pool indicates that there is no large P pool that is missed by the BBP extractions. The HCl extractable pool is likely close to the total P content in these soils. Deluca et al. [15] also observed a strong correlation between the citric acid- and HCl-extractable pools (r = 0.856), which is comparable to the proportional magnitudes we observed in our samples. We expected the HCl-extractable pool to include P extractable via a weaker acid as it has been reported that the HCl extraction is likely to capture extractable P via the citric acid, enzyme, and CaCl2 extractants [36]. However, we observed similar sizes for pools associated with the two acid extractants. This similarity suggests that the release of citric acid or a similar low molecular weight organic acid such as acetate or oxalate by plants or microbes may be an adequate strategy to acquire P compared to the release of a stronger acid.
Considering the low P yields for the enzyme extractions and the level of P limitation in this arid system, phosphatase production, in contrast to acids, may be a less suitable P acquisition strategy in drylands in comparison to more mesic ecosystems. Deluca et al. [15] observed P concentrations that were about double in the phosphatase-extractable pool compared to the citric acid-extractable pool (mean ~200 mg kg−1), while we observed the opposite relationship. The citric acid-extractable pool was roughly three times higher than the phosphatase pool in our analysis. However, despite low P concentrations, there is still the formation of at least some phosphatase-available organic P (i.e., organic P); thus, we cannot rule out the role of enzyme-based P acquisition in our dryland system. Although it has been shown that phosphatase activity will increase in response to P-deficient soils [38], the comparably small size of the phosphatase pool suggests enzyme hydrolysis may not be as an effective strategy in drylands. Thus, based on our results, we hypothesize dryland soil biota may be more likely to release a weak or low molecular weight acid, such as citric acid, while in a mesic system, phosphatase production may be the preferred strategy.
4.2. Short-Term Fate of PO43− Added to Biocrust-Dominated Soils
The radioisotope recovery results reveal that P added to the soil solution is quickly adsorbed to mineral surfaces or subject to microbial uptake as opposed to remaining free in the soil solution. The increased radiolabel presence in the acid-extractable pools compared to the other extractable pools indicates that the P introduced into this system is quickly adsorbed, which agrees with other studies that examined recovery after 24 h [39]. Thus, the release of organic acids, such as citric acid, or a stronger acid may increase the solubility of inorganic P that is added to the system via dust deposition.
The incorporation of radiolabeled P into the enzyme-extractable pool was unexpected. The labile organic P pool in the Chihuahuan desert has been found to be low in comparison to the labile inorganic pool [2], which agrees with the observation that drylands tend to have lower levels of SOM in comparison with other systems. Since phosphatase activity is highly correlated with SOM [7] and higher levels of microbial biomass, it is surprising that 25–40% of the added 33P was found in the enzyme-extractable pool. This result suggests that the PO43− was converted to a hydrolysable organic form during the 24-h period. Such a conversion is possible since microorganisms have been shown to rapidly synthesize organic P following an inorganic P addition [40,41]. Also, because arid soils are subject to P limitations due to the high CaCO3 content and lack of precipitation, organisms may be adapted to quickly take up and incorporate the P into biotic pools. In a similar radiolabel study in calcareous soils, it was observed that soils with low microbial biomass and low available P had higher label recovery in the microbial pool following a 24-h incubation [39].
The short-term fate of added PO43− provides insight on the path of P coming into the system via dust deposition, as dust is one of the main sources of nutrient input in drylands, and biocrusts that occupy surface soils are hypothesized to aid in the capture of P-containing dust particles [4]. Dryland organisms may quickly assimilate added inorganic P into their biomass. Crust organisms, including cyanobacteria, have the ability to mobilize inorganic PO43− [28] from mineral surfaces, and aquatic cyanobacteria have been found to remove orthophosphate from water sources, even at low phosphate levels [42]. Others have observed that P limitation may be a strong control in soils dominated by cyanobacteria [43,44]. Our results support the notion that drylands may be largely controlled by P limitation due to the observed recovery in the areas with intact crusts. Thus, there is a potential for tight cycling of P via phosphatase or organic acid acquisition by crust associated organisms. In addition, a previous study using an inorganic P addition and the BBP method to identify available P pools in a South Australian arid soil found low P concentration in the CaCl2-extractable and enzyme-extractable pools in soils grown with plants, suggesting that the P was taken up by the plants or microbes, as microbial biomass increased with plant growth [37]. This is comparable to what we observed in the soils with intact crusts. Active microbial communities such as biocrusts may illicit a similar response in taking up added P. Hoang and Marschner [37] also observed an increase in the CaCl2-extractable and enzyme-extractable pool in bare soils compared to soils growing with plants, suggesting that P released from the acid extractants may remain available in solution in the absence of plants [37]. However, the amount of variability between areas with and without intact surface crusts in our study does not provide strong evidence to differentiate between organismal strategies. The relative recovery across all samples regardless of crust type represents the possibility of a variety of soil microorganisms contributing to the release and uptake of P following organic acid or enzyme release.
4.3. Bioavailable P in Intact Crusts versus Areas of Disturbance
Across all four biologically available P pools (CaCl2, citric acid, enzyme and HCl-extractable P), P concentrations were higher in areas of disturbance and lower in areas with intact crusts. A possibility is that in areas with intact surface crusts, cyanobacteria and other crust organism may be rapidly taking up the P by incorporating P into their biomass or aiding in the retention of P in biotic pools, preventing a buildup on mineral surfaces in comparison to with areas of disturbance. The disturbed areas are loose soils with no intact surface crusts, and the observed high concentrations in these areas may be due to the lack of microbial activity. Similarly, Hoang and Marschner [37] found extractable-P across all pools to decrease in the presence of plants compared to bare loose soils. We cannot guarantee that the disturbed areas are devoid of free-living crust organisms and certainly contain other soil biota that may be contributing to microbial uptake or retention of P; however, on average, we found that areas with intact crusts have 30–80% less extractable P compared to areas of disturbance. Biocrusts in drylands have been shown to produce compounds such as oxalic acid, citric acid, and malic acid to release P [4], and cyanobacteria and microbes in this system are likely to produce a weak organic acid to access available inorganic forms of P. Biocrusts have been shown to induce bioweathering [45] through the release of weak acidic compounds. The low concentrations in the citric acid-extractable pool in areas with intact crusts may indicate a microbial strategy to induce bioweathering through the release of a weak organic acid. As a byproduct, P is released from the mineral soil and possibly lost via hydrologic transport to roots or leaching. Bioweathering initiated by crust organisms and other soil biota may lead to an increase in P bioavailability in the system. Our data are consistent with a system in which microbes in the Chihuahuan desert are contributing to the acquisition and release of P through the production of organic acids and increasing the bioavailable P pool.
As we observed in the acid extractions, the lower concentration in the intact surface crusts following the enzyme extraction may represent an acquisition strategy used by biocrust organisms and soil biota to access P. Biocrust organisms, specifically cyanobacteria, have the potential to increase phosphatase activity in soils by the release of extracellular phosphatases [4], and our data suggest that the release of phosphatases may be an additional mechanistic approach to acquire P. In a study focused on biocrust organisms and associated soils, Baumann et al. [45] found that biocrust organisms are considerable factors influencing concentrations of accessible P in temperate soils, and biocrust-associated organisms may excrete phosphatases to release P from Fe or Al. This is a likely mechanism used by dryland cyanobacteria and biocrust organisms in the Chihuahuan desert due to the Fe and Al mineralogy, and high carbonate composition. Thus, increased phosphatase production and excretion is a likely strategy used by biocrust organisms and other dryland soil biota to access inorganic P.
It is important to note that we assessed soils with and without biocrusts and not isolated biocrusts from the soil surface. Thus, one limitation of this study is the collection of soils from 0-10 cm depth. Biocrusts usually occupy the first 2 mm of the soil, while some associated organisms have been found in the top 1 cm [4]. The additional soil likely introduced other soil microbes with similar mechanistic strategies, and the acquisition mechanisms observed throughout our study may not be solely representative of strategies used by biocrust organisms. However, the differences we observed between the two areas of collection indicate a difference in microbial presence and potential strategic approaches in accessing P in drylands.
5. Conclusions
The results of the BBP assay revealed P stocks in these Chihuahuan desert soils that were mostly bound to mineral surfaces, showing the importance of geochemical controls on P cycling. However, our data also indicated that biological processes may be key to P cycling, especially in areas with intact biocrusts. Our results show that microbes may be accessing P through the release of organic acids and, to a lesser extent, phosphatase enzymes. In particular, the size of the citric acid-extractable pool suggests that organic acid exudation is a viable strategy to acquire P in this dryland ecosystem as it was nearly as effective as the release of a stronger acid. Phosphatase activity may also be a viable strategy to acquire P as we observed substantial P concentrations in the enzyme-extractable pool. The rapid incorporation of radiolabel into the enzyme-extractable pool further suggests that the organic P pool is still an important part of the P cycle in these soils despite low organic matter levels. The high radiolabel recovery observed in the acid and enzyme pools also suggests that newly added P can be quickly adsorbed or become subject to microbial uptake and does not remain in the soil solution for direct root uptake. The P concentrations and increased recovery in the citric acid- and enzyme-extractable pools suggest that there are two mechanistic approaches used by biocrust organisms and other soil microbes to prevent the sequestration of P into the mineral soil, which may increase the uptake potential and P available for plants and other soil biota in drylands.
Author Contributions
Conceptualization, G.M.C., A.D.-N., J.R.M. and B.B.; investigation, G.M.C.; resources, A.D.-N. and J.R.M.; writing and original draft preparation, G.M.C.; writing, review & editing, A.D.-N., J.R.M. and B.B.
Funding
This research was funded by National Science Foundation grant number 1557162 as well as startup funds from the University of Texas at El Paso to both A. Darrouzet-Nardi and J. McLaren.
Acknowledgments
I would like to thank Renato Aguilera for providing laboratory space, Armando Valera for his advice with radioactivity, the Biomolecule Analysis Core Facility at the University of Texas at El Paso for equipment access, and Tim Ferdelman (MPI Bremen) and Jan Jansa (AVCR) for their email correspondences.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, X.; Post, W.M. Phosphorus transformations as a function of pedogenesis: A synthesis of soil phosphorus data using Hedley fractionation method. Biogeosciences 2011, 8, 2907–2916. [Google Scholar] [CrossRef] [Green Version]
- Lajtha, K.; Schlesinger, H. The biogeochemistry of phosphorus cycling and phosphorus availability along a desert soil chronosequence. Ecology 1988, 69, 24–39. [Google Scholar] [CrossRef]
- Porder, S.; Ramachandran, S. The phosphorus concentration of common rocks-a potential driver of ecosystem P status. Plant Soil 2013, 367, 41–55. [Google Scholar] [CrossRef]
- Belnap, J. Biological phosphorus cycling in dryland regions. Phosphorus Action 2011, 26, 371–406. [Google Scholar]
- Dalal, R.C. Soil organic phosphorus. Adv. Agron. 1977, 29, 83–117. [Google Scholar]
- Cross, A.F.; Schlesinger, W.H. Biological and chemical controls on phosphorus fraction in semiarid soils. Biogeochemistry 2001, 52, 155–172. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, E.T.; Schlesinger, W.H. A comparison of fractionation methods for forms of phosphorus in soils. Biogeochemistry 1999, 47, 25–38. [Google Scholar] [CrossRef]
- Cross, A.F.; Schlesinger, W.H. A literature review and evaluation of the Hedley fractionation: Applications to the biogeochemical cycle of soil P in natural ecosystems. Geoderma 1995, 64, 197–214. [Google Scholar] [CrossRef]
- Hedley, M.J.; Stewart, J.W. Method to measure microbial phosphate in soils. Soil Biol. Biochem. 1982, 14, 377–385. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of available P by sequential extraction. Soil Sampl. Methods Anal. 1993, 7, 5–229. [Google Scholar]
- Johnson, A.H.; Frizano, J.; Vann, D.R. Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure. Oecologia 2003, 135, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Fox, T.R.; Miller, B.W.; Rubilar, R.; Stape, J.L.; Albaugh, T.J. Phosphorus Nutrition of Forest Plantations: The Role of Inorganic and Organic Phosphorus. Phosphorus Action 2011, 26, 317–338. [Google Scholar]
- Bowman, R.A.; Cole, C.V. An exploratory method for fractionation of organic phosphorus from grassland soils. Soil Sci. 1978, 2, 94–101. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Glanville, H.C.; Harris, M.; Emmett, B.A.; Pingree, M.R.A.; de Sosa, L.L.; Jones, D.L. A novel biologically-based approach to evaluating soil phosphorus availability across complex landscapes. Soil Biol. Biochem. 2015, 88, 110–119. [Google Scholar] [CrossRef]
- Darch, T.; Blackwell, M.S.A.; Chadwick, D.; Haygarth, P.M.; Hawkins, J.M.B.; Turner, B.L. Assessment of bioavailable organic phosphorus in tropical forest soils by organic acid extraction and phosphatase hydrolysis. Geoderma 2016, 284, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Fardeau, J.C. Dynamics of phosphate in soils. An isotopic outlook. Fertil. Res. 1995, 45, 91–100. [Google Scholar] [CrossRef]
- Di, H.J.; Condron, L.M.; Frossard, E. Isotope techniques to study phosphorus cycling in agricultural and forest soils: A review. Biol. Fertil. Soils 1997, 24, 1–12. [Google Scholar] [CrossRef]
- Barrow, N.J. A mechanistic model for describing the sorption and desorption of phosphate by soil. J. Soil Sci. 1983, 34, 733–750. [Google Scholar] [CrossRef]
- Buehler, S.; Oberson, A.R.; Rao, I.M.; Friesen, D.K.; Frossard, E. Sequential phosphorus extraction of a 33P-labeled Oxisol under contrasting agricultural systems. Soil Sci. Soc. Am. J. 2002, 66, 868–877. [Google Scholar] [CrossRef]
- Joner, E.; Jakobsen, I. Uptake of 32P from labelled organic matter by mycorrhizal and non-mycorrhizal subterranean clover (Tri-folium subterraneum L.). Plant Soil 1985, 172, 221–227. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Thomas, A.D.; Dougill, A.J. Spatial and temporal distribution of cyanobacterial soil crusts in the Kalahari: Implications for soil surface properties. Geomorphology 2007, 85, 17–29. [Google Scholar] [CrossRef]
- Murmann, R.P.; Peech, M. Relative significance of labile and crystalline phosphates in soil. Soil Sci. 1969, 107, 249–255. [Google Scholar] [CrossRef]
- Belnap, J.; Lange, O.L. Structure and functioning of biological soil crusts: A synthesis. Biol. Soil Crusts Struct. Funct. Manag. 2001, 150, 471–479. [Google Scholar]
- Reed, S.C.; Townsend, A.R.; Taylor, P.G.; Cleveland, C.C. Phosphorus cycling in tropical forests growing on highly weathered soils. Phosphorus Action 2011, 26, 339–369. [Google Scholar]
- Jones, D.L.; Oburger, E. Solubilization of phosphorus by soil microorganisms. Phosphorus 2011, 26, 169–198. [Google Scholar]
- Whitton, B.A. Soils and rice-fields. Ecology Cyanobacteria 2000, 257–259. [Google Scholar]
- Bolton, H.J.; Smith, J.L.; Link, S.O. Soil microbial biomass and activity of a disturbed and undisturbed shrub-steppe ecosystem. Soil Biol. Biochem. 1993, 25, 545–552. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Loza, V.; Marusenko, Y.; Mateo, P.; Potrafka, R.M. Temperature drives the continental-scale distribution of key microbes in topsoil communities. Science 2013, 28, 1574. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Crutchfield, J.; Vandiviere, M. Rapid sensitive, microscale determination of phosphate in water and soil. J. Environ. Qual. 2001, 30, 2206–2209. [Google Scholar] [CrossRef] [PubMed]
- Bünemann, E.K.; Steinebrunner, F.; Smithson, P.C.; Frossard, E.; Oberson, A. Phosphorus dynamics in a highly weathered soil as revealed by isotopic labeling techniques. Soil Sci. Soc. Am. J. 2004, 68, 1645–1655. [Google Scholar] [CrossRef]
- Carpenter, J.; Bithell, J. Bootstrap confidence intervals: When, which, what? A practical guide for medical statisticians. Stat. Med. 2000, 19, 1141–1164. [Google Scholar] [CrossRef]
- Chien, S.H.; Sikora, F.J.; Gilkes, R.J.; McLaughlin, M.J. Comparing of the difference and balance methods to calculate percent recovery of fertilizer phosphorus applied to soils: A critical discussion. Nutrient Cycl. Agroecosyst. 2012, 92, 1–8. [Google Scholar] [CrossRef]
- Efron, B. Better bootstrap confidence intervals. J. Am. Stat. Assoc. 1987, 82, 171–185. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2018. Available online: http://www.R-project.org (accessed on 21 May 2018).
- Hoang, K.T.K.; Marschner, P. Plant and microbial-induced changes in P pools in soil amended with straw and inorganic P. J. Soil Sci. Plant Nutr. 2017, 17, 1088–1101. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.D.; Voroney, R.P.; Lynch, D.H.; Oberson, A.; Frossard, E.; Bünemann, E.K. Microbially-mediated P fluxes in calcareous soils as a function of water-extractable phosphate. Soil Biol. Biochem. 2017, 106, 51–60. [Google Scholar] [CrossRef]
- Tate, K.R. The biological transformation of P in soil. Biol. Process. Soil Fertil. 1984, 11, 245–256. [Google Scholar]
- Stewart, J.W.; Tiessen, H. Dynamics of soil organic phosphorus. Biogeochemistry 1987, 4, 41–60. [Google Scholar] [CrossRef]
- Gaffney, A.M.; Markov, S.A.; Gunasekaran, M. Utilization of cyanobacteria in photobioreactors for orthophosphate removal from water. Appl. Biochem. Biotechnol. 2001, 91, 185–193. [Google Scholar] [CrossRef]
- Knelman, J.E.; Schmidt, S.K.; Lynch, R.C.; Darcy, J.L.; Castle, S.C.; Cleveland, C.C.; Nemergut, D.R. Nutrient addition dramatically accelerates microbial community succession. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Darcy, J.L.; Schmidt, S.K.; Knelman, J.E.; Cleveland, C.C.; Castle, S.C.; Nemergut, D.R. Phosphorus, not nitrogen, limits plants and microbial primary producers following glacial retreat. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Glaser, K.; Mutz, J.E.; Karsten, U.; MacLennan, A.; Hu, Y.; Leinweber, P. Biological soil crusts of temperate forests: Their role in P cycling. Soil Biol. Biochem. 2017, 109, 156–166. [Google Scholar] [CrossRef]
Figure 1.
Areas of intact surface crusts (left) versus areas of disturbance (right).

Figure 2.
Extractable P across all four extractant types and crust types (black = disturbed, gray = intact). For both crust types, the citric acid and HCl extractions yielded higher P contents than the CaCl2 and phosphatase extractants. The horizontal lines indicate the mean and the vertical lines indicate the standard error.
Figure 2.
Extractable P across all four extractant types and crust types (black = disturbed, gray = intact). For both crust types, the citric acid and HCl extractions yielded higher P contents than the CaCl2 and phosphatase extractants. The horizontal lines indicate the mean and the vertical lines indicate the standard error.

Figure 3.
33P recovery across all four extractant types and crust type (black = disturbed, gray = intact). The citric acid, phosphatase, and HCl pools showed the highest label recovery in comparison to the CaCl2 extractant. The horizontal lines indicate the mean and the vertical lines indicate the standard error.
Figure 3.
33P recovery across all four extractant types and crust type (black = disturbed, gray = intact). The citric acid, phosphatase, and HCl pools showed the highest label recovery in comparison to the CaCl2 extractant. The horizontal lines indicate the mean and the vertical lines indicate the standard error.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Extractants used to access different forms of phosphorus that provide evidence for different microbial or plant acquisition strategies [15].
Table 1.
Extractants used to access different forms of phosphorus that provide evidence for different microbial or plant acquisition strategies [15].
| Extractant Type | Form of P Accessed | Biotic System Mimicked by Extraction Method |
|---|---|---|
| 0.01 M CaCl2 | Weakly adsorbed inorganic P | P accessed by root interception & diffusion |
| 0.01 M citrate | Active inorganic P sorbed to clay particles or weakly bound in inorganic precipitates | Organic acid release by plants and microorganisms |
| 0.2 enzyme unit (wheat germ phosphatase) | Organic P readily attached by acid phosphatase and phytase enzymes | Enzyme release by plants and microorganisms to access labile organic P |
| 1 M HCl | Soluble, active and moderately stable inorganic P adsorbed to mineral surfaces or present in inorganic precipitates. | Proton release by plants and microorganisms to access adsorbed and precipitated P. |
Table 2.
Explanation of the relative effectiveness ratios of different biological mechanisms assessed by the biologically-based P pools (BBP) method. Categories of high, similar and low refer to the magnitude of the ratios.
Table 2.
Explanation of the relative effectiveness ratios of different biological mechanisms assessed by the biologically-based P pools (BBP) method. Categories of high, similar and low refer to the magnitude of the ratios.
| Ratio | High (>1) | Similar (≈1) | Low (<1) |
|---|---|---|---|
| Citric acid/CaCl2 Enzyme/CaCl2 HCl/CaCl2 | Acids or enzymes may be effective P acquisition strategies beyond taking up soil pore water. | Most P is readily available in soil pore water | Unlikely to observe since CaCl2 pool should be a subset of other pools |
| Citric acid/Enzyme | Weak acids may have more potential than phosphatases for P acquisition. | Weak acids have similar potential to phosphatases for P acquisition. | Weak acids may have less potential than phosphatases for P acquisition |
| HCl/Citric acid | Weak acids are not sufficient to release P from the soil matrix; inorganic P strongly occluded. | Weak acids are sufficient to release P from the soil matrix. | Unlikely to observe since citric acid pool is likely a subset of the HCl (a stronger acid) pool |
| HCl/Enzyme | Most P in the soil is inorganic. | There are substantial organic and inorganic P pools | Most P in the soil is organic |
Table 3.
Results of the paired bootstrap analysis across individual samples and crust type for P concentration following four extractants.
Table 3.
Results of the paired bootstrap analysis across individual samples and crust type for P concentration following four extractants.
| Crust Type | Comparison | Effectiveness Ratio | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|
| Intact | Citric acid/CaCl2 | 374 | 211 | 966 |
| Enzyme/CaCl2 | 62.2 | 32.4 | 245 | |
| HCl/CaCl2 | 293 | 124 | 667 | |
| Citric acid/enzyme | 6.02 | 4.17 | 7.63 | |
| HCl/citric acid | 0.782 | 0.483 | 1.39 | |
| HCl/enzyme | 4.71 | 3.21 | 7.69 | |
| Disturbed | Citric acid/CaCl2 | 107 | 65.6 | 324 |
| Enzyme/CaCl2 | 41.3 | 21.8 | 136 | |
| HCl/CaCl2 | 121 | 51.1 | 422 | |
| Citric acid/enzyme | 2.60 | 2.17 | 3.13 | |
| HCl/citric acid | 1.13 | 0.794 | 1.44 | |
| HCl/enzyme | 2.93 | 2.31 | 3.77 |
Table 4.
Results of the paired bootstrap analysis across individual samples for 33P label recovery among four extractants and crust types.
Table 4.
Results of the paired bootstrap analysis across individual samples for 33P label recovery among four extractants and crust types.
| Crust Type | Comparison | Effectiveness Ratio | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|
| Intact | Citric acid/CaCl2 | 12.1 | 7.54 | 31 |
| Enzyme/CaCl2 | 5.56 | 3.29 | 11.7 | |
| HCl/CaCl2 | 11.7 | 6.69 | 19.7 | |
| Citric acid/enzyme | 2.18 | 1.48 | 2.61 | |
| HCl/citric acid | 0.969 | 0.491 | 1.57 | |
| HCl/enzyme | 2.11 | 1.07 | 3.12 | |
| Disturbed | Citric acid/CaCl2 | 6.29 | 4.43 | 11.5 |
| Enzyme/CaCl2 | 5.15 | 2.68 | 9.84 | |
| HCl/CaCl2 | 10.4 | 5.13 | 29.8 | |
| Citric acid/enzyme | 1.22 | 0.76 | 1.73 | |
| HCl/citric acid | 1.65 | 1.06 | 2.81 | |
| HCl/enzyme | 2.01 | 1.05 | 3.63 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Crain, G.M.; McLaren, J.R.; Brunner, B.; Darrouzet-Nardi, A. Biologically Available Phosphorus in Biocrust-Dominated Soils of the Chihuahuan Desert. Soil Syst. 2018, 2, 56. https://doi.org/10.3390/soilsystems2040056
AMA Style
Crain GM, McLaren JR, Brunner B, Darrouzet-Nardi A. Biologically Available Phosphorus in Biocrust-Dominated Soils of the Chihuahuan Desert. Soil Systems. 2018; 2(4):56. https://doi.org/10.3390/soilsystems2040056
Chicago/Turabian StyleCrain, Grace M., Jennie R. McLaren, Benjamin Brunner, and Anthony Darrouzet-Nardi. 2018. "Biologically Available Phosphorus in Biocrust-Dominated Soils of the Chihuahuan Desert" Soil Systems 2, no. 4: 56. https://doi.org/10.3390/soilsystems2040056