3.1. Global Terrain Recalculation
While raw global data is available (e.g., from UN-FAO’s “
Global Terrain Slope and Aspect Data”), this is uncompiled, so an estimate of global slope is extracted from 30 arc-second resolution (ca. 1-km) summary data (Nunn & Puga 2004: appendix) [
11] with mean slope for all 234 nation and dependent states (excluding Antarctica) here calculated as 3.94% or nearly 4% (ca. 2.29°). A 4% slope is 4 cm rise per metre run with the hypotenuse just 100.08 cm, or an extra 0.08% length, which is also an extra 0.08% area. Considering each country’s area and slope separately about doubles this to an extra 0.154% land overall (as calculated in attached data file), but this is still unrealistic.
A recent 2007 calculation from USGS’s Global Slope Dataset (
pubs.usgs.gov/of/2007/1188/pdf/OF07-1188_508.pdf) of “
accurate summary statistics at 30-arc-seconds describing the underlying 3-arc-second data” fails to yield a summary. An earlier paper [
52] at five arc-minutes (10-km) had much lower global terrestrial slope of between 0–1.5°, whereas a later paper [
51] shows that such high scales widely underestimate the true situation. Since land surfaces at 1-km scales are quite unrepresentative, so published terrain data at lesser scales are presented and reviewed in the succeeding sections.
3.1.1. Macro: Terrain
Recently, Ying et al. (2014) [
51] claimed the first comprehensive estimate of the contributions of topography to the surface-area of the whole of China using Incremental Area Coefficients (IACs) as the percentage area increase of the surface area when compared with the projected area. This metric is the same as a tortuosity index. They highlighted scale-related factors and some potential environmental revisions of natural resources and ecosystem functions when area needs are taken into account. For China at 30-m resolution and a vertical error of less than 20-m, they calculated a mean surface area increase of 4.6% with the largest increment for a 50 km × 50 km cell being >45%. At 100-m resolution, the mean increase was 3.76%; at 1000-m (1-km) it was 0.5%; while at 10,000-m it was negligible (0%). Extrapolating these values linearly would give more than 4.5% increase in surface area at the 1-m scale (attached Excel chart). But, they also clearly showed (their figs. 5 and 9) that the results are exponentially dependent upon scale of observation: as resolutions approach the 1-m scale the area estimates increase markedly indicating threshold values for different classes of landscape below which the surface-area increment caused by topographic relief cannot be ignored.
Ying et al. [
51] also found that the mean slope of the DEM across China at the spatial resolution of 30-m was 10.92° (19.29% slope), at 100-m it was about 9° (15.84%), while at 1000-m it was reduced to 3.53° (6.17%), and at 10,000-m it too was negligible; extrapolating this linearly would give about 12° (21% slope) at a 1-m scale for China. This compares to Nunn & Puga [
11] data that, at the horizontal scale of 30 arc-seconds (926-m), have a mean slope of China of 5.49% (3.14°) just lower than Ying et al.’s 1000-m value and 3.8 times lower than the estimated 1-m value. It may thus be concluded that Nunn & Puga’s values are at least four times underestimations of likely 1-m scale values. Nunn & Puga’s overall Global average land area increase, based on slope at 1000-m resolution, was recalculated (Excel file attached) to be +0.154% of the flat area estimation, this multiplied four times to comply with an extrapolated 1-m scale from Ying et al.’s equivalence data, gives a value of around +0.616% overall globally. The following table summarizes these findings for China alone (
Table 3).
A real-world study of DEMs at finer scales is by Milevski & Milevska (2015: tab. 1) [
53] (5–90-m resolutions) on a patch of ground (20 × 20 km = 400 km
2) in the Skopje area of Macedonia. They found that slope accuracy increased 25 percentage points from a mean slope of 8.8° at 90-m to 11° at 5-m. This represents an increase in land area from its 400 km
2 base to 404.8 km
2 (+1.2%) and to 407.5 km
2 (+1.9%), respectively, with projection to >2% at 1-m resolution.
In the mountainous state of Himachal Pradesh in India, calculation by the local government [
54] (tab. 3) gave 3-D TSA of 86,384.77 km
2 from original 2-D TMA of 55,342.79 km
2 or an increase of approximately 56.1%. However, resolution was at only 24-m or 71-m scale. At finer increment—say 1-m or less—the TSA can be expected to yield a much higher figure. Tentative, true surface areas from a study using 90-m SRTM DEM for the rugged states of Jammu and Kashmir [
55] found 3-D and 2-D areas differed by nearly 25%: “
(296,513 km2 vs. 222,236 km2, respectively)”. Finer slope resolution will considerably increase the surface reality to the planimetric model, and refined rugosity more so. Further real-world examples at the higher scale are needed.
Although more accurate datasets are increasingly available (e.g.,
eorc.jaxa.jp/ALOS/en/aw3d30/), it is expected that, as the resolution decreases from 30-m, the total land may easily double at each iteration, possibly approaching 100% at 1-m scale, i.e., double the land surface area to the map area. In support, a study using a 10 × 10 km plot in the mountainous Pyrenees (Nogués-Bravo & Araújo 2006: fig. 1) [
56] has actual surface area of 280 km
2 or (180% greater area with ratio of 1:2.8) at 100-m scale; more than double that of 130 km
2 (30%) at the 500-m scale; while at the 1-km scale the surface area appears to be only about 110 km
2 (or just 10% larger).
In order to calculate the Soil Organic Carbon (SOC) in Chinese soils, Zhang et al. (2008) [
57] calculated 3-D terrain for three mountainous states. The results increased soil surface area from 2-D of 78.04 Mha to 3-D area value of 84.02 Mha (7.7% increase although only at coarse scale of 90-m). From this, they calculated the SOC storage to 1 m depth increased from 10.9 to 11.9 Gt (+9.2%), which is of interest to a later section of this report.
Sutton & Lopez (2003) [
58] “
ironed out” Colorado finding it ~12% larger (at scale of 90-m).
3.1.2. Meso: Tortuosity and Soil Roughness or Rugosity
The meso scale relates to an important measure of insolation defined as solar irradiance with energy measured in watt-hours per square metre (Wh/m
2) or in the Langley, which is one calorie per square centimetre (= 41,840 Jm
−2). These are both defined for horizontal area values and the latter cm
2 scale is approximately the same size as an earthworm burrow or surface cast. This is an appropriate level of observation for measuring basic ecological interactions locally and then extrapolating to a global value (as is routinely done by NASA, UN, FAO, IPCC, etc.). See also [
38].
From the foregoing, it seems that tortuosity is strongly influenced by the observational factor: the more intense the scale, the higher the tortuosity index (T value). Indeed, a study in Canada by Martin et al. (2008) [
59] shows a fourfold increase in bare earth tortuosity only when resolution was reduced to less than 10-cm starting from one metre scale. Martin (2008: fig. 5, tab. 1) [
59] show a T
B value of 16 based upon a T
A Tortuosity index of 1.2 from a TSA of 240 m
2 and TMA of 200 m
2, i.e., 20% greater surface area for bare soil at their 0.75-cm scale. However, it appears this study, as with several others, did not adequately consider slope foreshortening, which for a straight hypotenuse of about 20 m and stated angle of 18 degrees gives a baseline of 19 m or 5% lesser base length. If we unrealistically assume that the slope is smooth and constant for its width, this then gives a simple Tortuosity Index of at least 240/190 = 1.26 (+26%), which is 5% above their calculation. Note that in this study [
59] (fig. 5a), the vegetated rather than bare-soil hillslope had a tortuosity index of about 1.5 or a DSM area at least twice the bare earth DTM value.
A study from Brazil using a 3-D laser profile scanner at intervals of 1-cm (Bramorski et al. 2012: tab. 2) [
60] reported soil tortuosity under conventional and no-tillage with mean index (T) values of 89.62 and 57.4 giving an overall mean index of 73.5 or +7250%! This tortuosity index was stated to be based on that of [
61]. Communication with the author (Julieta Bramorski, email pers. comm. 11–18 July 2017) confirmed a mistake in their calculations and a new mean value of 1.33 (+33%) was arrived at. Yet, my re-working of the same data (kindly supplied by the primary author) gives a Tortuosity index (T
i) of around 4.56 that recalculated to allow for curved arc lengths rather than straight hypotenuses, gave a mean T
i of 7.16 (+616%). The constant ratio between these two means is 1.57 (+57%) and the combined mean of these two values gives a compromise of T
i = 3.6 (or +260%). The source data and Excel calculations are attached (“Julieta” section of Excel spreadsheet data file).
The mean for all four independent calculations at the mm scale is +94.0%.
3.1.3. Micro: Biodiversity, Productivity and Respiration
Of two German micro scale studies, one compares different methods of measurement but provides no usable data [
62] (fig. A1); another [
63] (tab. 2) has mean field index value of 1.23 (i.e., +23%) at 2 or 3-mm grid spacing with height accuracy better than 0.5 mm. A French study at 90 × 90-mm had a mean tortuosity index of around 2, i.e., double relief length to same projected length or +100% (Mirazai et al. 2008: fig. 6) [
64]. Also, in Europe, (Tarolli et al. 2017: tab. 1; fig. 6) [
44] summarized the various Roughness Indices and showed tortuosity doubling or quadrupling logarithmically when scale reduces from 40-mm to 4-mm scale with mean field index around 0.35 (a slight mistake in the legend is index “T
A” while text has “T
P”), this translates as an increase of 35% or 1.35 from their formulae (in their tab. 1) at this finest scale.
While defining Tortuosity-index as the ratio of total surface area to the map area i.e., T
B = TSA/TMA after Helming et al. (1992) [
63], an Austrian report (Grims et al. 2014: tab. 3) [
65] at 1-mm resolution has a field value mean of T
B = 2.63 that implies a true surface area more than two and a half times the flat horizontal footprint (i.e., +163%). [Mislabelled as “TB (%)” in Grims et al. (2014: tab. 3) [
65], the primary author confirmed by email (pers. comm. 27 July 2017) that this is in fact the dimensionless index value not percentage]. Incidentally, this paper also measured soil organic carbon (SOC) and reported a mean value of 2.0% humus (= SOM or SOC?) in the study fields.
An online accessible but possibly unpublished Canadian thesis has cultivated soil surface area up to almost double the flat area (1.9 m/m
2) with a mean value of laser roughness at the less than 1-mm scale of 1.6 (+60%) (Koiter 2008: sects. 2.3, 2.6, 2.7) [
66].
The mean value for all five mm scale results is +108.2%.
3.1.4. Sub-Micro: SOM Surface Areas and Gaseous Exchanges
At the microporous scale, soil organic matter (SOM) and its colloids are reported to have adsorbic surface area for gaseous exchange of CO
2 of between 94−174 m
2 g
−1 (de Jonge 1996: tab. 2) [
67] with a mean of 130 m
2 g
−1 (this value of 130 m
2 g
−1 is used in calculations of humic SOM bulk densities below and in an attached summary report). His paper quoted earlier studies showing SOM surface areas up to 800 m
2 g
−1, or six times greater, and this latter value approaches that of mineral zeolite or montmorillonite (also known as bentonite) clay. However, other studies only found 1 m
2 g
−1 [
68]. The SOM data are on an “
ash free basis”, i.e., just the dry, organic content of the sample is calculated even though a non-porous, inert mineral component was present in the samples. The solid phase densities average about 1.1 g cm
−3 (de Jonge 1996: tab. 2) [
67], and, regardless of whether from square or cylindrical measurements, the base area would be about 1 cm
2. The ratio of surface area (130 m
2) to flat area (1 cm
2) is thus approximately (10,000 × 130 =) 1.3 million times. As soil on a ‘flat-Earth’ occupies ~12 Gha then this would theoretically have surface area increase by 12 × (1.3 × 10
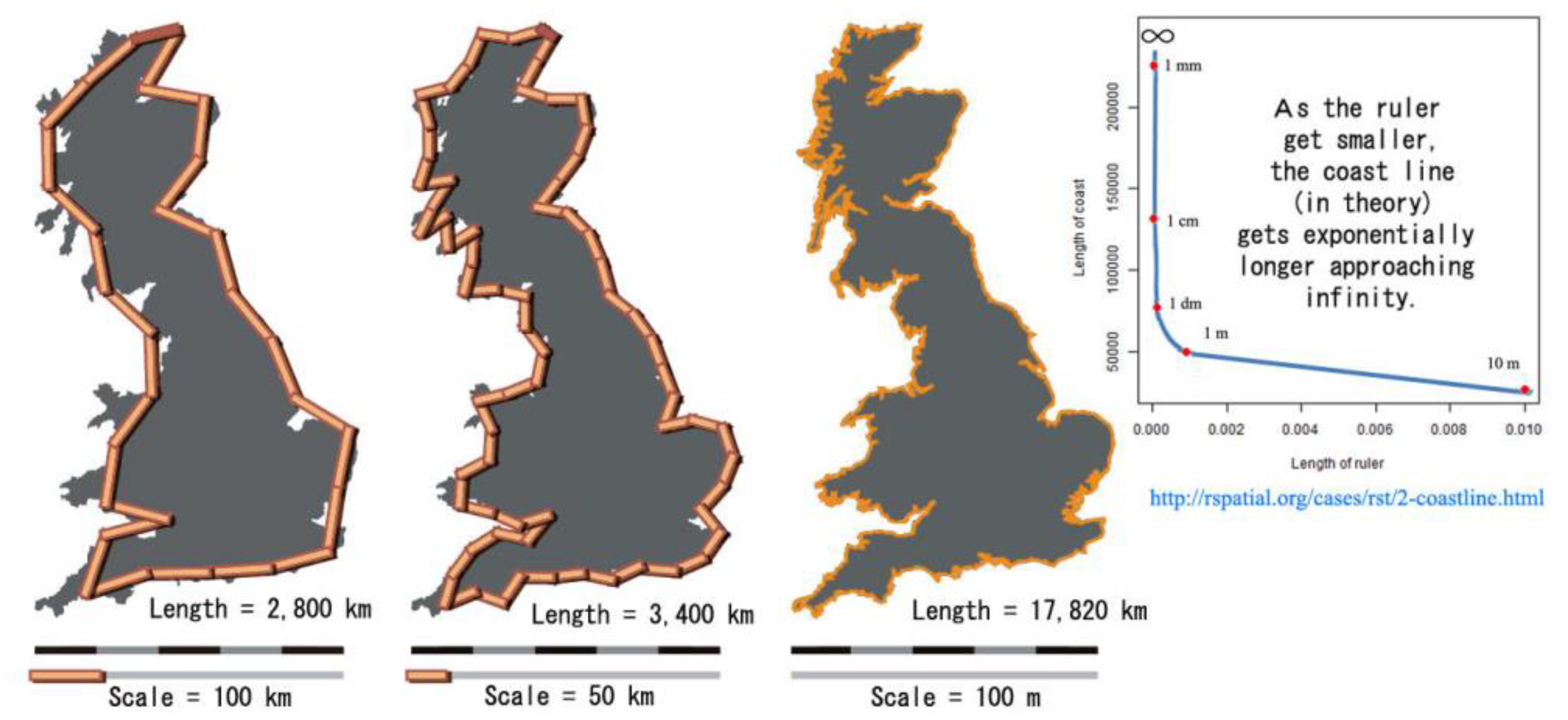
6) = 15.6 Pha. This implies that true absorbic surface area of soil exposed to the atmosphere is almost infinitely expandable—as with the coastal paradox cited in
Appendix A and as for the theoretical DTM and DSM models newly re-calculated in a section below.
3.1.5. Total Recalibration for New Land Surface Areas
Mean values from the studies reported above are tabulated in summary (
Table 4).
In summary, the table above confirms km scale readings are unrepresentative. The three 1-m scale projections give mean +2.38% area, while the mean of all 16 macro scale readings is +21.25%, this latter possibly being the most applicable to more hilly terrains. For the meso cm-scale, the mean of all four results is +94.0%, while the five micro mm-scale results give mean of +108.2%. Thus, to a basic flat land area of 15 Gha we may apply between 2.4–21.3% increase, and, to 80% of this product (equivalent to a flat 12 Gha of soil), the other two progressive increases may be overlain. Finally, the approximately 20% (ca. 3 Gha) non-soil area initially subtracted, should be added to give a new total land surface, as it is calculated in the contingency summary (
Table 5).
Antarctica and Greenland include sub-ice terrain thus 15 Gha is the base value upon which macro tortuosity indices are imposed. Then, for soil-bearing land only or about 80% of outcome area (12.3–14.6 Gha), a median land increase is of ((×3.5 + 4.2)/2) = × 3.85, which is nearly four times original 15 Gha land. Combined with an immutable flat ocean area of 36 Gha, the new land area of 53–63 Gha gives a new World area of 89–99 Gha, albeit superimposed upon this is a theoretically infinite SOM microporosity. As to which set of scales is selected, this depends upon what practical calculations (e.g., biomass, NPP, gas exchange, proper allocation of grant funds, etc.) are deemed most relevant for a particular study. A Fermi calculation (below) allows area increase to ~100 Gha.
3.2. Theoretical DTM Model and Fermi Calculations
Jenness (2008) [
47] noted slope-aspect algorithms generated indices around 1.6–2.0 (as per Hodgson 1995 [
48]) with a median 1.8. Thus, from a land surface of 15 Gha, at the metre or dm scale, this may increase to 27 Gha, and, as ~80% supports soil, its tortuosity at the cm scale may be similarly increased by 1.8 times (27 × 0.8 × 1.8 =) 38.9 Gha. It is possible to argue that the mm scale allows a further 1.8 times area to give a final total of (38.9 × 1.8 =) 69.98 or about 70 Gha. This plus 36 Gha ocean and 5.4 Gha barren land (27 × 20%) gives a theoretical new total surface area of ~111.4 Gha, which is tolerably close to the values (around 100 Gha) that were calculated above from on-the-ground field readings.
In support of higher land area, a study from Germany [
69] discusses the problems, technical issues, and recent developments whilst providing examples from model terrains seemingly at 1-m resolution at least for test square mapped landscapes with perimeters of 400 m (but 2-D area of just ca. 1000 m
2 or 31.3 m side or perimeter of 126.5 m in their fig. 4?) derived from their fig. 2 of square patch areas (after Jenness 2004 [
47]). Increases of patch areas show in their fig. 4 are from 2-D of about 1000 m
2 to 3-D of up to 10,000 m
2 or 20,000 m
2, i.e., by ten or twenty-fold (or 900–1900%). Their (fig. 4) “
Average Surface Roughness” indices go from an obvious zero in 2-D up to eight in 3-D, or by an infinite amount but implied as an eightfold area increase (+700%). This gives further support for current fourfold landscape increase (from 15 → ca. 60 Gha or +300%) as being entirely reasonable if not a wide theoretical underestimation of total land area.
Because the true surface of the land is paradoxical and it depends upon arbitrary, shifting, and overlaid scales of observation, the most pragmatic solution is perhaps to accept a compromise Fermi value pending further acuity. To transpose the scale problem it may be more practicable to arrive at a reasonable and ‘convenient’ working model of global surface area (i.e., the surface directly exposed to sunlight, atmospheric gas exchange and rainfall) as 100 Gha with 64 Gha attributed to landscapes and topsoils.
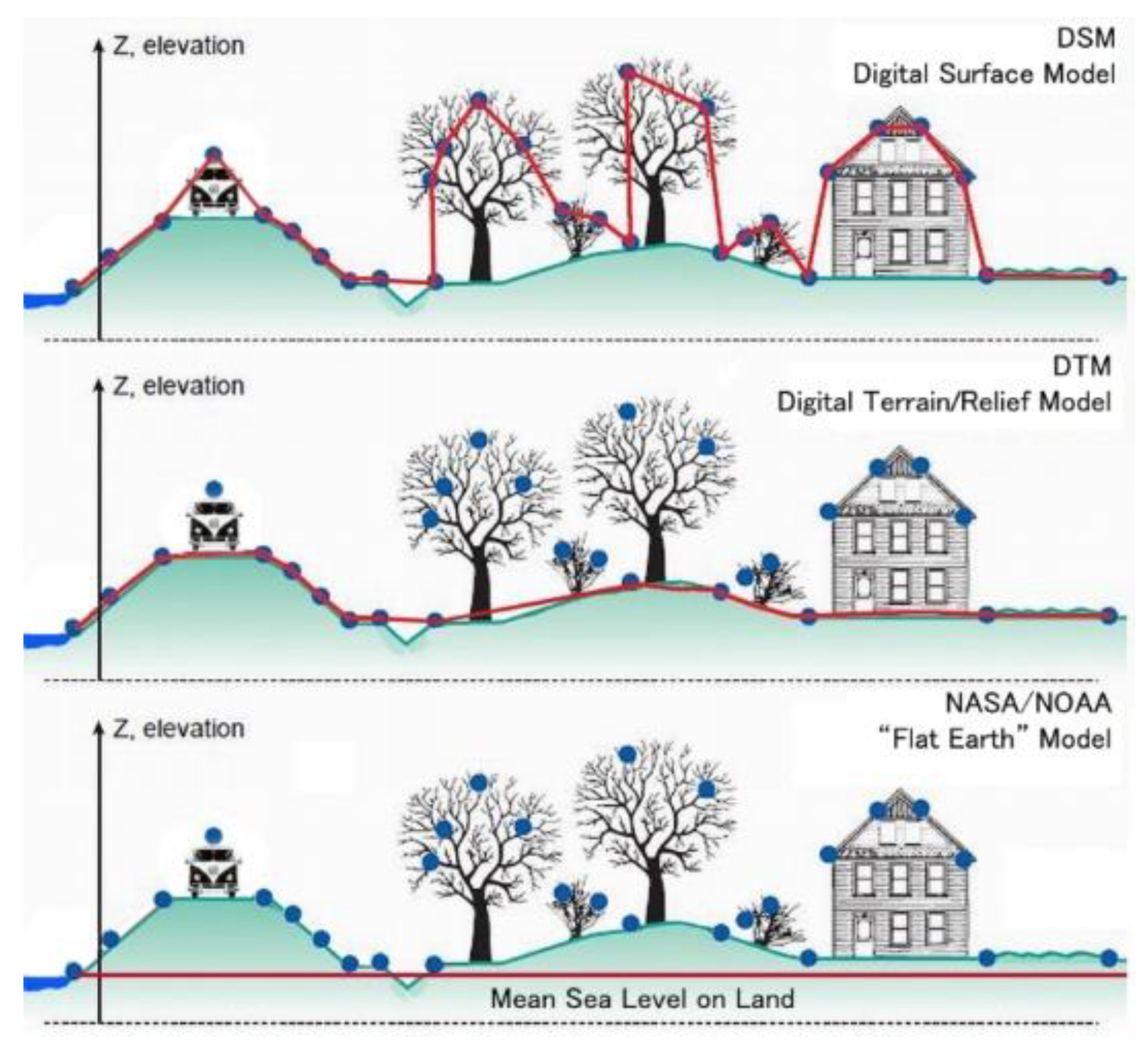
DTM for DSM Recalculations
Overlain upon the bare-earth terrain DTM is an increasing superficial DSM (cf.
Figure 6). An estimate of effective DSM is possible if we apply the Leaf-Area-Index (LAI). This dimensionless quantity characterizes effective plant cover defined as the one-sided green leaf area perpendicular to flat unit ground surface area (LAI = leaf area/flat ground area, m
2/m
2). LAI ranges from 0 (bare ground) to ~18 (dense forest canopies) and a global average (from Asner et al. 2003) [
70] is 4.5. These authors state “
LAI is a key variable for regional and global models of biosphere-atmosphere exchanges of energy, carbon dioxide, water vapour, and other materials”. It is surely just as important to have estimates of a global DTM and DSM too. Prof. Greg Asner (pers. comm. email 20 July 2017) kindly clarified: “
That estimate is the average of studies published for different vegetated ecosystems, so it does not represent the actual global land area”. Thus only soil bearing terrain is considered in the following calculations.
As about 80% of land supports soil, on the conventional flat-Earth view and in the new view, a rough estimate of prior, conventional, DSM is of 12 Gha × 4.5 = 54 plus 3 Gha ice or desert-covered land = 57 Gha. From my new topographical calculation DSM is (64 Gha × 80% =) 51 Gha soil × 4.5 LAI = 230 Gha, which is an important measure related to global photosynthesis potential (plus a lesser ocean contribution). Since LAI is for one side of the leaf, then the total for both sides of a leaf presumably gives 230 × 2 = 460 Gha DSM plus 3 Gha non-soil land, plus 36 Gha from flat oceans = 499 Gha global DSM estimate. Cities or townscapes occupy about 1–3% of land area with additional parks, gardens, verges, etc. that would add slightly to this very rough estimate of DSM. Moreover, this may be an underestimation, as, rather than a LAI of 4.5, Whitman et al. (1998: 6580) [
23] assumed a more than double LAI of 10 (but source and whether single-sided were unstated).
Microporosity will futher increase the DSM since, strictly, any internal surfaces or pore spaces are also part of the surface area if defined as the interface of solids or liquids exposed to air. Just considering plant respiration, this internal areas of stomata of leaves is possibly unquantifiable. However, leaves are a major contributor to humic soil organic matter (SOM) with its micro-porous surface area for gas exchange shown as between 1.5–120 Pha and an argument may be made that this is the truly astronomical surface area of land making the mere quadrupling of the DTM of ‘flat-Earth’ area of just 15 Gha to 64 Gha seem entirely reasonable and easily justified, as indeed is the almost ten times increase of coarse DSM from 57 Gha to 499 Gha.
Subterranea (e.g., caves, caverns, or karsts) are an additional but minor ‘surface’ area consideration, but earthworm burrows may be considerable. Burrows systems, as noted above, were found to extend for up to 888 m/m
2 in length (=8880 km ha
−1) and their void volume varied tenfold from 1.3–12.0 m
2/m
2 ground surface in the upper 1.2 m of soil during an observation period of 1.5 years (Lee 1985: pp. 196, 208) [
29,
49]. On conventional 12 Gha flat soil, this is at least 1.3 ha/ha × 12 Gha = 15.6 Gha. However, on rugose topsoil, this would be about four times greater, i.e., at least 62.4 Gha that may vary up to 624 Gha or a 0.6 Tera-hectare volume of below-ground earthworm burrow voids. It should be noted that that study mainly represented pasture soils in France, but the samples excluded both the 0–6 cm layer of soil and also burrows <2 mm in diameter (Lee 1985: 196) [
29]. Including these smaller burrows and other micro pore spaces, in the topsoil especially, would presumably increase underground volumes of sub-surface spaces substantially. Nevertheless, including sub-soil voids may double the DSM to (499 + 624 =) 1123 Gha or 1.1 Tera-hectare. The flat ocean’s surface (that exposed to Sun, air, rain) remains at 36 Gha and its bathymetry or rugosity largely an irrelevancy.
3.3. Bulk Density (BD) Backcheck
Support for the current terrain argument is from bulk density (BD) that compels revision. Tangible sub-samples are taken on the ground at fixed core sample volumes with a constant planimetric area (cm−2 or m−2 perpendicular to the centre of the Earth) and then multiplied by a biome’s area, thus mass may be adjusted to comply only by adding biome area by adding terrain/topsoil relief.
For habitable biomes supposedly totaling 12.3 (flat) Gha, (Whitman et al. 1998: tab. 2) [
23] gave mean soil bulk density as 1.3 g cm
−3 (= tm
−3) and (Lee 1985: 195) [
29] assumed a bulk density of 1.4 g cm
−3, so a reasonable mean may be 1.35 g cm
−3. Total SOC to one metre recalculated (from FAO’s Harmonized World Soil Database, HWSD, as noted in attached
Supplementary data file) gives median values for SOC of around 1.3% and their mean soil BD is ~1.4 g cm
−3 (close to 1.35 g cm
−3). Total conventional ‘flat-Earth’ topsoil mass to 1 m depth would then be [(123 × 10
12 m
3) × 1.35 tm
−3 = 166 × 10
12 t =] 166,000 Gt topsoil and 1.3% SOC = 2158 Gt C.
Allowing for organic soils having lower BD than mineral soils, highly organic, peaty Histosol humic-SOM BD is 0.1 g cm
−3 (Köchy et al. 2015: 354) [
71] as an ideal for SOM organic matter with 50% C (from Pribyl 2010) [
72]. Prior best estimate of total SOC to 1 m depth (e.g., by IPCC 2013,
www.4p1000.org, etc.) was 1500 Gt giving total × 2 SOM of 3000 Gt on planimetric 12 Gha land or 120,000 m
3 to 1 m depth. Thus, a BD was of (3000/120,000 GtGm
−3 =) 0.025 g cm
−3, which is below the required SOM BD of 0.1 g cm
−3 and thus needs × 4 mass. The only plausible way to increase mass is by increasing real biome area to allow for terrain/topsoil. When the soil surface is doubled for terrain and again for topsoil micro-relief then mass of soil increases. Since BD measurements typically use a core cylinder of fixed volume, thus the actual undulating surface area is immaterial. For demonstrative purposes of real BD, if we assume quadruple SOM 3000 Gt → 12,000 Gt whilst maintaining 12 Gha planimetric area (or rather its volumetric equivalent to 1 m depth), the resulting bulk density of 0.1 tm
−3 exactly matches the required mean of 0.1 tm
−3 (
Q.E.D.).
Is it reasonable to increase land area values fourfold? Given a BD mean of 1.35 g cm−3 (or tm−3) and allowing for a fourfold increase in soil occupied land area (i.e., 12 Gha × 4 = 48 Gha), then total soil mass to 1 m would be (480,000 Gm−3 × 1.35 t) = 648,000 Gt globally. If SOC is 1.3%, then the total SOC to 1 m is 8424 Gt (that tolerably agrees with a 8580 Gt value calculated below from empirical sources).
Similarly, a planimetric soil area of 12 Gha to 3 m depth (= 360,000 Gm3) requires a new SOM of 36,000 Gt to give the required 0.1 g cm−3. If 3 m SOC doubles from 8580 → 17,160 × 2 = 34,320 Gt SOM giving BD of (34,320/360,000 =) 0.095 tm−3 or tolerably 0.1 g cm−3 (Q.E.D.). However, both mean bulk density and SOC % are perhaps less reliable at depths greater than 1 m.
Another calculation, possibly artifactual, is with prior SOC >1 m depth (Köchy et al. 2015) [
71] of 3000 Gt × 2 for 6000 Gt SOM on planimetric 12 Gha if to a sample depth of, say, 3 m = 360,000 Gm
3 giving real SOM bulk density of just 0.016 tm
−3 or out by a factor of six for average BD of peaty SOM of around 0.1 tm
−3. This discrepancy may be resolved with reference to terrain/relief by about ×6 from flat 12 Gha to about 72 Gha that, plus 3 Gha hot or ice deserts and 36 ocean, gives total area of 111 Gha. Seeming slightly excessive this may be ultimately reasonable and is, coincidentally, nearly the same value of 111.4 Gha as arrived at earlier with theoretical DTM models.
Reasoned indications thus point to Earth’s real surface area in the realm of 111 Gha with 75 Gha bare-earth (68%) and just 36 Gha sea (32%) or about two-thirds of the World being land-based.
Standard BD reference of planimetric 12 Gha to the centre of the Earth, overlain by terrain/soil relief, etc. by using multiplication factors are summarized, assuming the global mean BD 1.35 gm
−3 and SOC 1.3%, as compared to current conventional SOC values (of 1500 Gt to 1 m or 3000 Gt to 3 m depth), showing their multiplication shortfalls (
Table 6).
This table shows IPCC’s current conventional 1 m SOC estimates (ca. 1500 Gt) is out by a factor of 1.4, and other possible terrain scenarios by between 2.8–8.4 times. Terrain × factors are for coarse landforms, and also for superficial cm2 + mm2 relief details that, at both these finer scales, are mainly composed of superficial SOM-humus/earthworm casts.
For reference (from Wikipedia), amorphous carbon densities are 1.8–2.1 g cm−3 differing from dry soil bulk density that varies in its minerals, biotic, as well as its air space voids (porosity).
Although clearly revealing conventional underestimations of SOC/SOM, these variable result from BD calculations probably relate to difficulties in obtaining global BD means and their complexity with soil depth. The upper 1 m results are likely most reliable. Full calculations and justification for bulk density assay may be scrutinized in the attached
Supplementary Files.
3.4. Soil, C and a “Missing Sink” Discrepancy
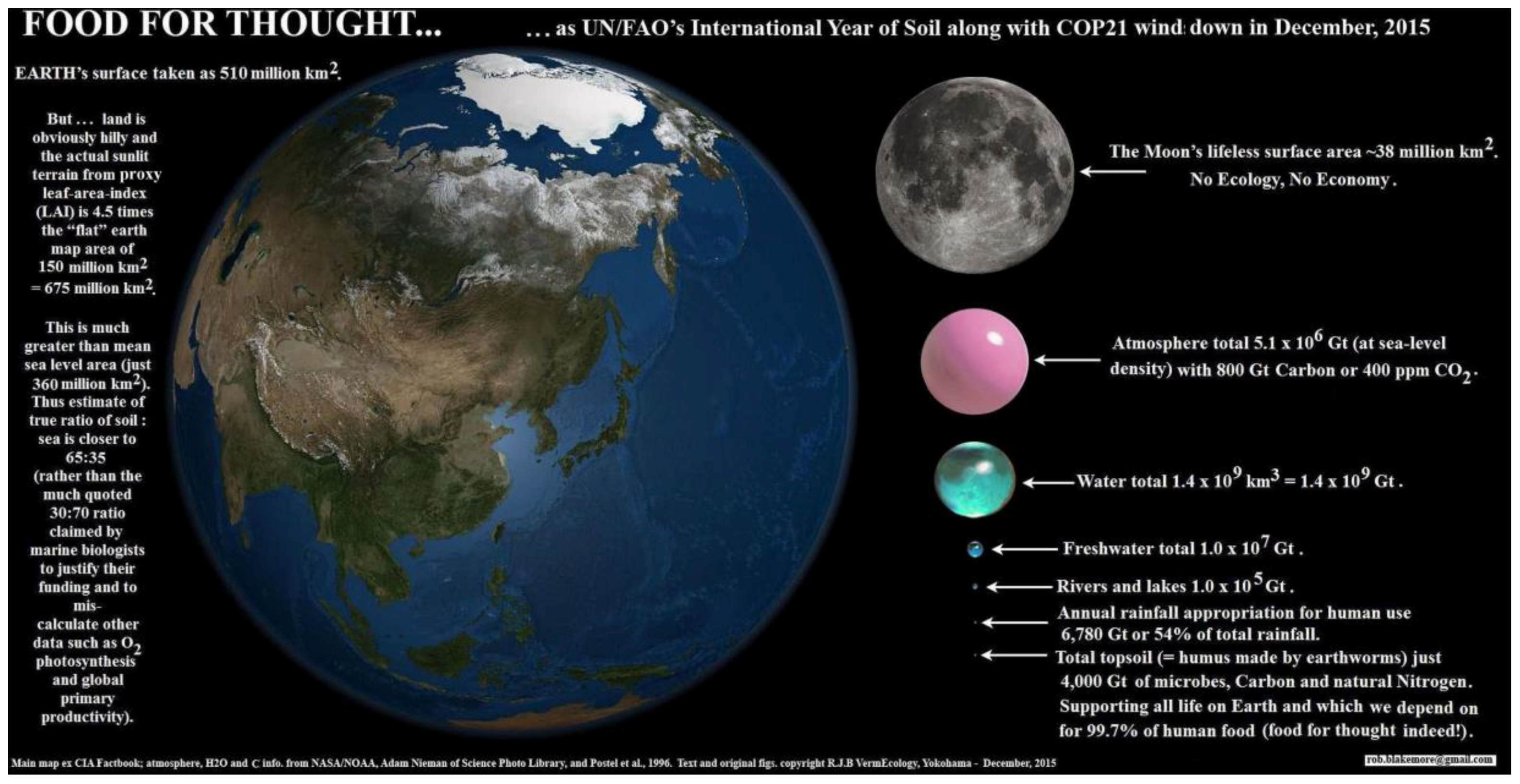
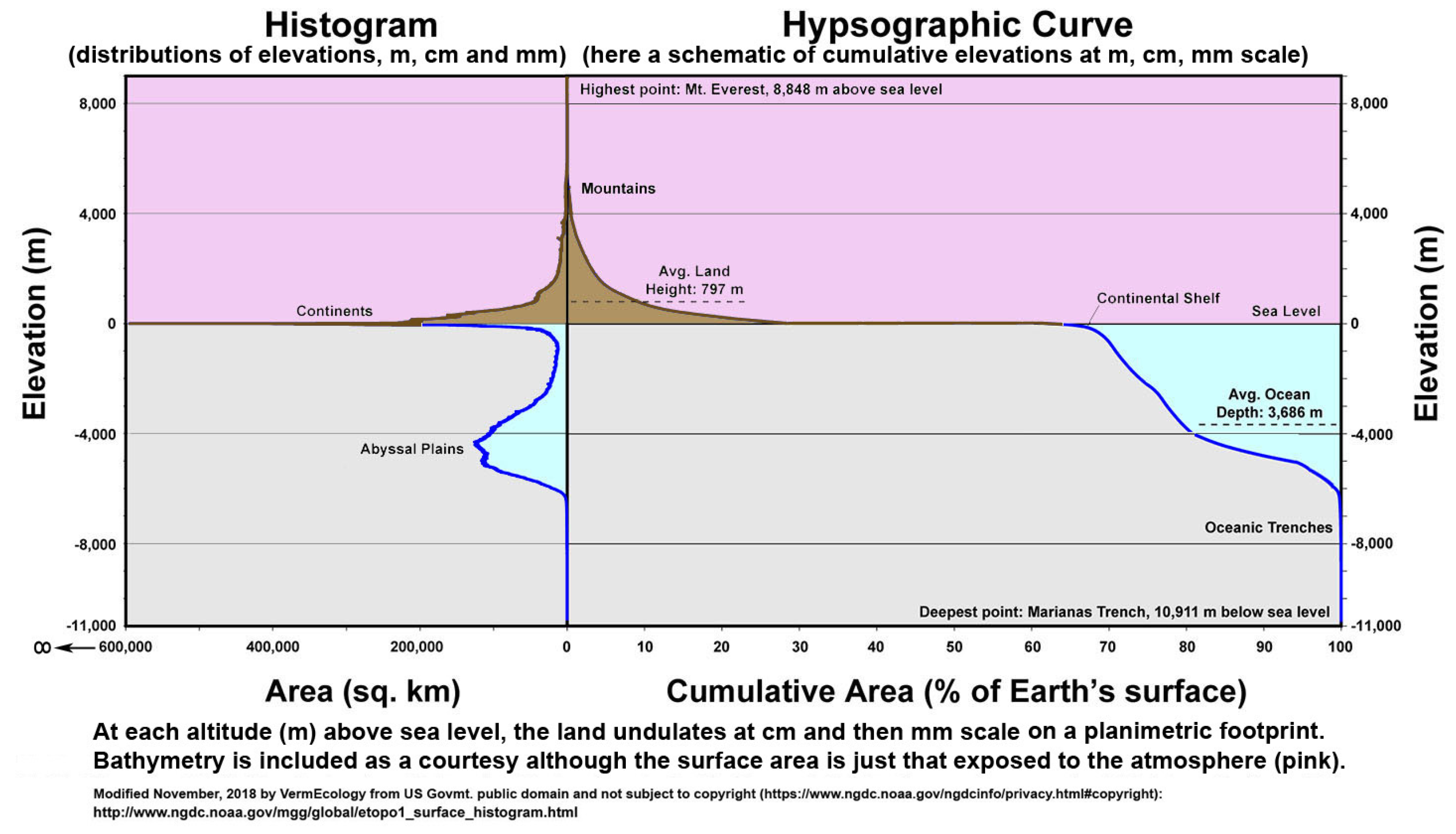
Primary sources of global carbon budgets as used by IPCC (e.g., by authors such as Batjes, Haughton, Jackson & Jobbágy, and Prof. Rattan Lal) invariably give a land area total of about 15 Gha on a globe of around 51 Gha; however, this is for an idealized flat surface whereas it is self-evident that land is hilly (cf.
Figure 1). With topological consideration, all land areas may be slightly increased at one kilometre scale (by ~1–5%). As already noted above, on study showed topsoil surface area from 2-D to 3-D increased by 7.7% at a coarse scale of 90-m and its SOC storage to 1 m depth was upped by +9.2% [
57]. Also, as calculated above, 1-m scale projections give mean land increases of +2.38–21.25% (median about 10%), and soil carbon may certainly be increased, likely doubled or quadrupled, at finer resolutions. Factors are: sub-surface SOC, roots, and soil biota. Justification is that these are measured at the mm to cm scale and then applied at the m to km scale.
Lal (2008: fig. 1) [
73] cites a “
missing sink” of 2.6 Gt/yr C, as discussed in a
Supplementary file.
3.4.1. Total Soil Carbon (SOC/SOM) for Recalculation of Global Carbon Budget
Relating to global warming and Greenhouse Gasses (GHGs), carbon is by far the major issue with the problem, and the solution, to be found mainly in the ground (
Table 7,
Figure 10).
Global SOM-humus stock data are not readily available but they may be calculated from global soil organic carbon (SOC) given as 1500 Gt by IPCC 2013,
www.4p1000.org 2015,
http://www.fao.org/3/a-i6937e.pdf 2017, see [
74] (fig 1), 2300 [
41], 2397 [
75], or as 2956.5 that is quoted as ~3000 Gt [
71]. Value differences are largely due to depth of topsoil sampling [
8,
74], the first is 0–1 m, the second is 0–3 m, and the third and fourth most recent values include soil greater than 1 m [
71] (by Köchy et al. 2015 who possibly have mean 4.0 m for peats or to depth of soil for other types?). Then, taking their higher value of 3000 Gt and applying the revised van Bemmelen factor of SOM = 2 × SOC (Pribyl 2010) [
72], the total SOM is 6000 Gt on a dry-weight or an “
ash free basis”. However, all values are for ‘flat-Earth’ calculations of just ~12 Gha soil area having a SOM bulk density (BD) of 6000/120,000 = 0.05 tm
−3, and if this is doubled for terrain and coarse relief, then the total topsoil mass is presumably increased too. That is, for SOC from 3000 → 6000 Gt and for SOM humus 6000 → 12,000 Gt with a new SOM bulk density, keeping same area due to fixed core sample volumes, as 12,000/120,000 = 0.1 tm
−3 the significance of which is already noted in the BD section above. These new increased values, however, may themselves be underestimations.
In addition to terrain considerations, [
74] (p. 11) noted that: “
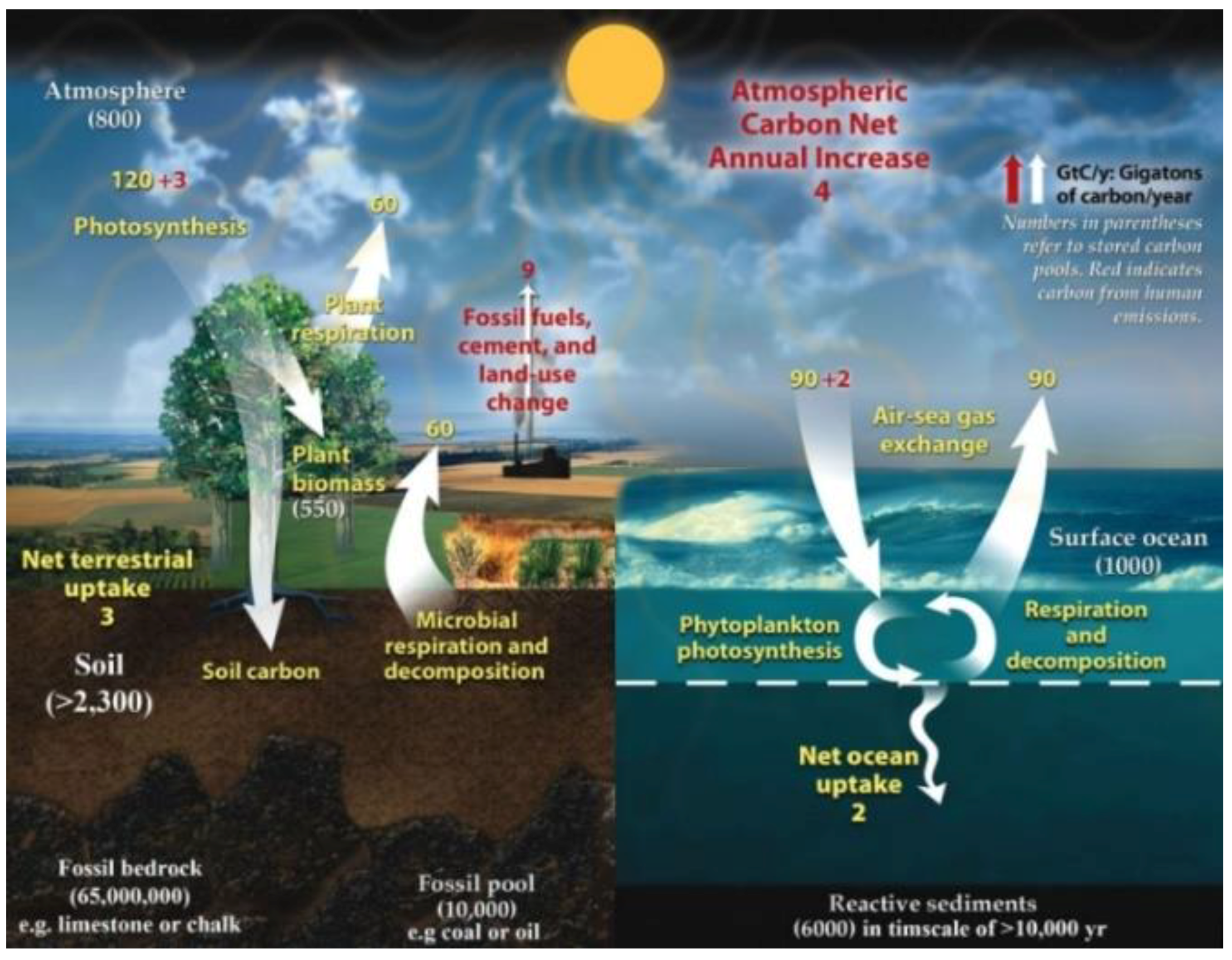
Soil carbon values require allowance for intractable glomalin adding a further 5–27% to almost all SOC tallies (Comis, 2002). Plus data from deep soils may increase budgets: e.g., Harper & Tibbett (2013) found C up to five times greater in Australian soils at depth >1 m and down to 35 m in some cases. The Walkley-Black method itself underestimates total C by about 20% with a correction factor of ca. 1.3 often required [this W-B correction is from Pribyl, 2010], whereas latest techniques using mid-infrared (MIR) spectroscopy give more accurate readings. These three factors combined would surely increase soil SOC totals”.Thus, assuming that soil depth factors are already included with terrain area, 6000 Gt SOC × 1.3 W-B correction = 7800 Gt plus, say, median value 10% for glomalin = 8580 Gt total soil carbon. Worldwide, the reactive organic carbon stored in soils (herein from 3000 → 8580 Gt) greatly exceeds the most generous amounts that are attributed in above-ground phytomass (700 Gt), plus atmosphere (800 Gt) and surface oceans (1000 Gt), which equal just 2500 Gt in total when combined (cf.
Figure 10).
Global topsoil humic SOM is then also raised from 8580 SOC × 2 to approximately 17,160 Gt (but, as calculated above, to greater than 1 m depth this may be doubled again to ~34,320 Gt).
Turnover time for fast pool carbon is estimated at 23 yrs [
75] cf. 10–15 yrs according to (IPCC 2007) [
76]. These then would also be duration for processing of humic SOM by detritivore earthworms, as indeed Darwin (1881) [
31] extrapolated from his minute observations: “
All the fertile areas of this planet have at least once passed through the bodies of earthworms”. From this, it was reasoned [
74] that all atmospheric carbon is theoretically processed via leaf-litter through the intestines of earthworms in ~12-year cycles. That is, unless populations are severely depleted [
15].
3.4.2. Root Stocks, Vesicular-Arbuscular Mycorrhiza (VAM) Hyphae, Litter, Crusts, and Earthworms
Relating to above-ground vegetation are the often ignored underground root-area-indices (RAIs) with fine roots a prominent sink for carbon, often much greater than that of vegetation above ground. Extending many metres below ground, interlinking with kilometers of symbiotic VAM fungal hyphae, roots are routinely excluded from soil samples by manual removal and sieving.
Estimated total root biomass was 292 Gt containing 146 Gt carbon and representing 33% of total annual net primary productivity (Jackson et al. 1997: tabs. 2–3) [
37]; however, this seemingly was updated by Mokany et al. (2005: 95) [
77] to 241 Gt C for roots. UNEP (2002: 10) [
26] estimate that probably over 80% of plant production enters the soil system either through plant roots or as leaf-litter. It was also shown that perhaps 50% of below-ground allocation is released as extra-root carbon exudates [
78], some being ‘traded’ with microbes for Nitrogen fixation or other growth factors. Additionally, estimates are of at least 15 Gt C for soil mycorrhizal VAM hyphae [
79].
Some vegetation surveys, but certainly not all, make allowance for below ground biota and for living or dormant biomass and dead necromass. Also, generally excluded from calculations of SOC (and SOM) mass is leaf-litter—an important part of the soil profile transitioning to humus—that contributes considerably to the global carbon budget with a “
pedologic pool” of 40–80 Gt giving a median stock value of 60 Gt (Lal 2008: fig. 1) [
73,
80]. Moreover, autotrophic biofilm or biocrust (e.g., bryophytic liverworts, hornworts, and mosses plus microfungi/yeasts, photosynthetic green algae, lichens, and Cyanobacteria or Cyanophyta) also coat and inhabit the convoluted superficial and interstitial surface rocks, topsoil, and sand. These ‘cryptogamic covers’ of biocrust total 5 Gt C [
81].
Complete soil carbon thus strictly includes root mass (241 Gt), leaf-litter (60 Gt), plus VAM (15 Gt), biocrusts (5 Gt), and earthworms (2–4 Gt from [
82]) to total 323 Gt that may all be reasonably doubled to allow for terrain to (323 × 2 =) ~650 Gt carbon. If this is then added to the 8580 SOC × 2 = 17,160 SOM calculation from above, it gives a new total SOM to depth of about 17,810 Gt (as per Abstract, but, as also estimated above, this may likely be doubled again to ~34,320 Gt).
3.4.3. Microbial Biotic Carbon: Living, Dormant or Dead (Including Fossils and Geology)
Regarding microbial biotic carbon (most of which is included in the SOC data), a much-cited study (Whitman et al. 1998: tabs. 2, 5) [
23] of prokaryotes [viz. Monera (simple bacteria) and Archaea] estimated their total cellular carbon biomass as up to 450 Pg (= 450 Gt) that these authors stated to equal the carbon storage in land plants. Their allocation of prokaryotic mass was approximately 50:50 ocean to land (actually 48–241 Gt carbon in soil versus 305.2 in sea). But, their land estimates (tab. 2), although up to 8 m depth, is for ‘flat-Earth’ biome areas, which they say totals 12.3 Gha excluding ice, multiplied by numbers of microbe cells sampled from each biome; whereas, for ocean in (their tabs. 1 and 3) are unit volume of sea (cells/mL) thus immutable, or cells/cm
3 in sediments at depths (most in 0.1–10 m depth). It is likely that terrain/relief will more than double the land count and thus the total biomass by at least one third. Taking their upper 241 Gt value × 2 for land terrain and ×2 for topsoil relief = 964 Gt (plus 305.2 Gt in sea = 1269.2 Gt total biotic carbon). However, a more recent ocean re-assessment [
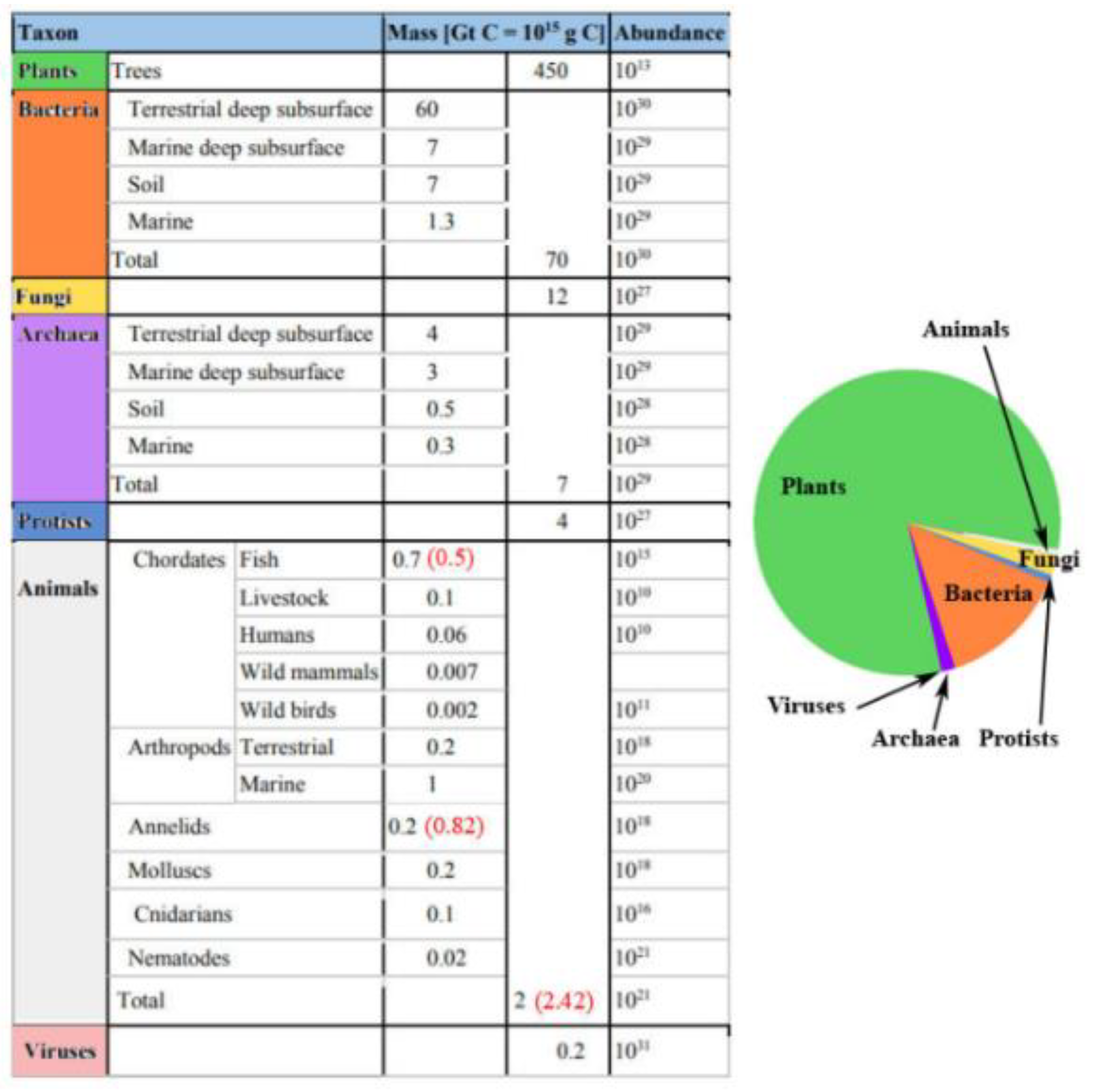
6] reduced microbial biomass on the seafloor due to paucity in actual deep ocean cores from their original 303 billion tonnes of C to just 4.1 billion tonnes representing just 0.6% of Earth’s total living biomass and reducing the total global biotic carbon to about [964 + (2.2 + 4.1) =] 970.3 Gt with most (i.e., 964 Gt) in soil. Thus, land’s C allocation (99.35%) is yet again greatly enhanced proportionately to that of the ocean (0.65%) (cf.
Table 2,
Figure 3).
The UNEP (2002: tab. 2.1) [
26] “
World Atlas of Biodiversity”, despite claiming global coverage, is mainly concerned with marine/ocean/water and barely mentioned soils, nevertheless, had total carbon content of Earth as ~100,000,000 Gt C, allocated as in the following, modified, and corrected, tables (
Table 8 and
Table 9).
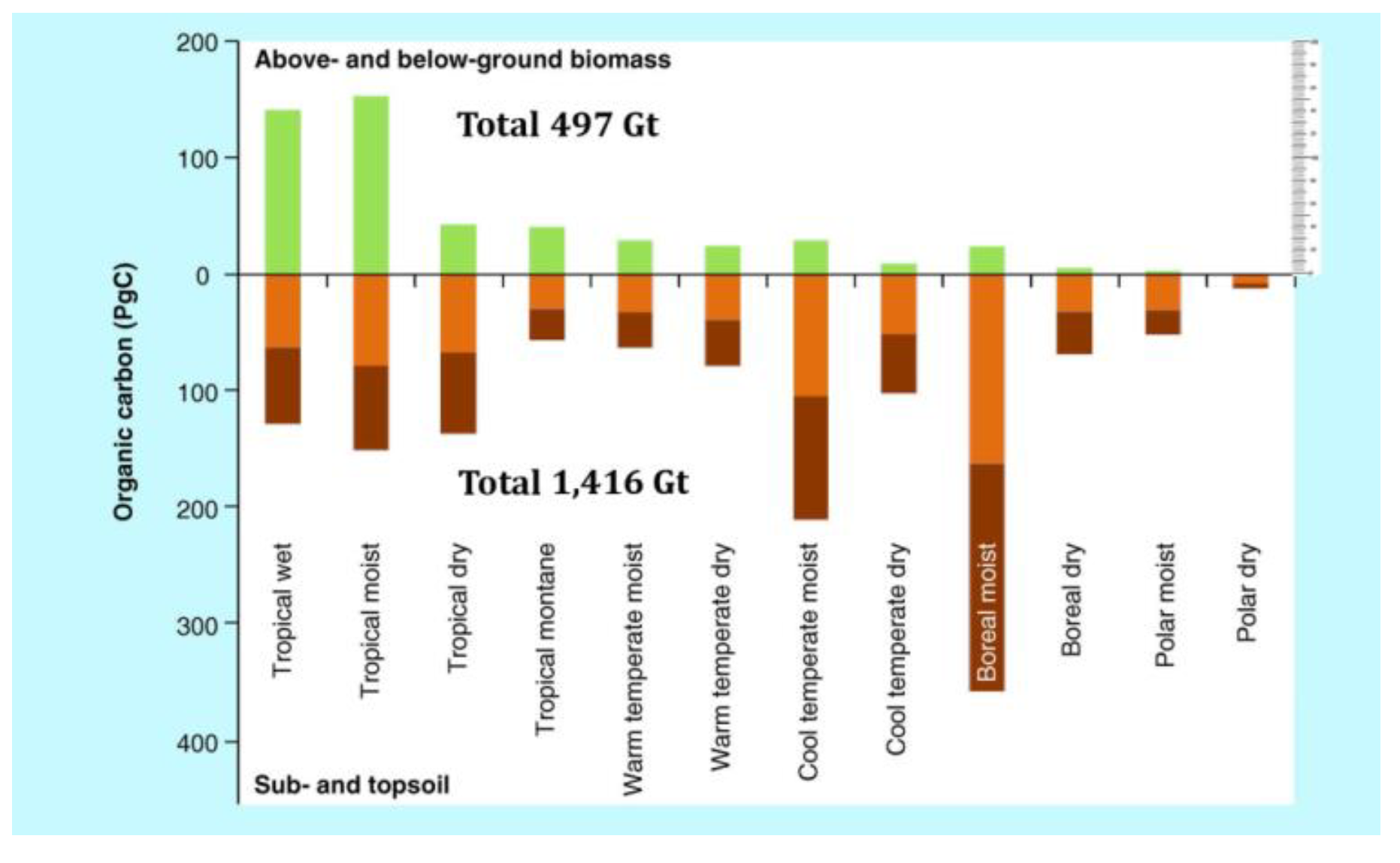
3.4.4. Above and Below-Ground Biodiversity and Biomass Carbon Rechecked (plus Ocean C)
It is remarkable that almost always overlooked or undervalued in biodiversity assessments are the communities and networks of below-ground soil biota that represent both the Earth’s highest diversity and its greatest biomass (even without consideration of terrain effects) (
Figure 11 and
Figure 12).
Most calculations of terrestrial fauna and flora (microbes, plants, fungi and animals) based upon ‘flat-Earth’ biomes or habitats require revision and likely doubling or quadrupling, and this affects relative ocean proportions. Although the total animal biomass appears to be insignificant in comparison to land plants [
26,
81] just considering megadrile earthworms, recent calculations [
82] of 1.3 quadrillion worms with fresh weight ‘vermi-mass’ of 4–8 Gt, may be doubled for terrain relief to 2.6 quadrillion and a massive 8–16 Gt (with carbon content up to 4 Gt). If correct, earthworms would be truly significant (as Darwin 1881 surmised), even though they are apparently annihilated under conventional, chemical agriculture [
15,
29]. When compared to a recent best estimate of global fish “
wet weight” of just 1–2 Gt [
85] (with carbon at most 0.5 Gt), this casts glib comments about worms being good fishing bait in a whole new light.
Life on Earth may be elevated as summarized in carbon calculations above. However, as noted, an ignored sub-surface biomass in the rhizosphere of VAM fungi and roots substantially increase the land proportion [
37]. For roots Mokany et al. (2005: 95) [
77] said: “
Our results yield an estimated global root stock of 241 Pg C, a similar value to that proposed by Robinson (2004), but about 50% higher than the 160 Pg C estimated by Saugier et al. (2001). This dramatic increase in estimated global root carbon stock corresponds to a 12% increase in estimated total carbon stock of the worlds vegetation (from 652 to 733 Pg)”. Searching their sources, the value 652 Pg is likely above-ground vegetation (from “
Saugier et al.
2001”) of 492 Pg, plus Robinson’s (2004) [
78] estimate of 160 Pg root (492 + 160 = 652). The 733 is seemingly from the same above-ground value plus their own estimate of 241 Pg root carbon (492 + 241 = 733 Pg = Gt).
Thus, a total of above- and below-ground land vegetation are reasonably accepted as 733 Gt C, which, along with bacteria from Whitman et al. (1998:
Table 5) [
23] and as re-assessed by [
6] of 241 Gt vs. 6.3 Gt in soil vs. sea, respectively, gives biomass carbon on land of (733 + 241 =) 974 Gt C. Mycorrhizal VAM-fungal hyphae and biocrusts add 15 and 5 Gt (974 + 20 = 994), plus 2–4 Gt earthworms [
82] and 7 Gt for other organisms [
81] = ~1000 Gt total. This terrestrial carbon may be doubled for terrain (and possibly doubled again for soil relief, especially for microbes) to give between 2000–4000 Gt land C, plus an ocean contribution of just 14.8 to total at least 2014.8 Gt of living, respiring, biotic carbon (
Table 10).
As carbon is universally about 50% dry weight, a new value is at least (2000 × 2) = 4000 Gt dry biomass on land plus (14.8 × 2) = 29.6 Gt in sea. Since water content is taken as 50% [~30% in wood (
www.wood-database.com/wood-articles/wood-and-moisture/) and 40–70% in bacteria [
86,
87] with median value ~50%], then this value is doubled again to at least 8000 Gt wet weight on land plus (14.8 × 4 =) ~60 Gt in sea to give new total for Earth’s living, respiring, fresh mass of ~8060 Gt, or roughly ~8 Tera-tonnes (Tt) of biomass.
These data compare to [
88] Vaclav Smil’s (2011) total dry biomass of Life on Earth he estimated as just 1600 Gt (here more than doubled to at least 4029.6 Gt maybe 8060 Gt). As a cross-check, the total biosphere carbon is estimated at between one to four Trillion tons [
89]; thus, my current estimate of around 2000 Gt C (2 Tt) is about mid-range but is closer to the best case scenario of 4 Tt.
Total terrestrial carbon of at least 2000 Gt in land organisms mostly intermixes with the 8580 Gt or so SOC in SOM or humus as active carbon stored and recycled on land, as compared to just 900–1000 Gt reactive carbon in the oceans (Lal 2008: fig. 1) [
41,
73]. Observable today, as in the geological past, is how biologically active (vermi-)compost—part of SOM-humus—rapidly recycles organic remains, hence one reason why topsoil leaves few soft tissue fossils or manures when compared to water submersion, anaerobic inundation, or mud that all stifle decomposition and give rise to both fossils and bio-sedimentary rocks that are formed from macro- or meso-biota and microbial remains.
3.5. Biodiversity of Species
Concomitant with the increasing detail of terrain is a realization that the biological scale of life on Earth is also increasingly refined to reduce the major living components from the scale of giant trees and massive mammals, to that of invertebrates, and finally to the microbial components that, on most recent revisions, have the largest biomass, biodiversity, and contributions to biotic energy cycles (also as a biopharmaceutical resource). As Ying et al. (2014) [
51] succinctly state: “
The increase in surface area with spatial resolution should mean more living space and a more diversified environment for smaller sized organisms, which comprise the majority of species (and thus contribute more to biodiversity). This trend also leads to underestimation of the role of environmental processes occurring at finer scale”.Terrain increase has most significance to smaller, superficial microbes and soil Arthropoda (mainly insects), but it has less relevance for colonial soil societies, such as ants or termites with colonies that are concentrated in localized nests or communal mounds rather than individuals being widely and deeply dispersed as indeed are earthworms. While the present recalibration makes only a moderate difference to habitable land at the metre scale—that is, for large animals like humans and their livestock or for large plants—it makes a greater change to habitat space for organisms in the realms of the cm scale (for example larger insects and earthworms) and a massive difference for the majority of animals and plant life that are measured in mm or less down to the micrometre (µm) microbe scale. Greater land surface gives higher abundance, biomass, and biodiversity, especially for the hordes of autotrophic, heterotrophic, symbiotic, and parasitic microbes, including fungi, which already dominate the Earth and exist mainly in the living soil provisioning our vital services, essential resources, and providing our new medicines too (
Figure 13).
Below are conventional global biodiversity and biomass calculations (
Figure 14 and
Figure 15).
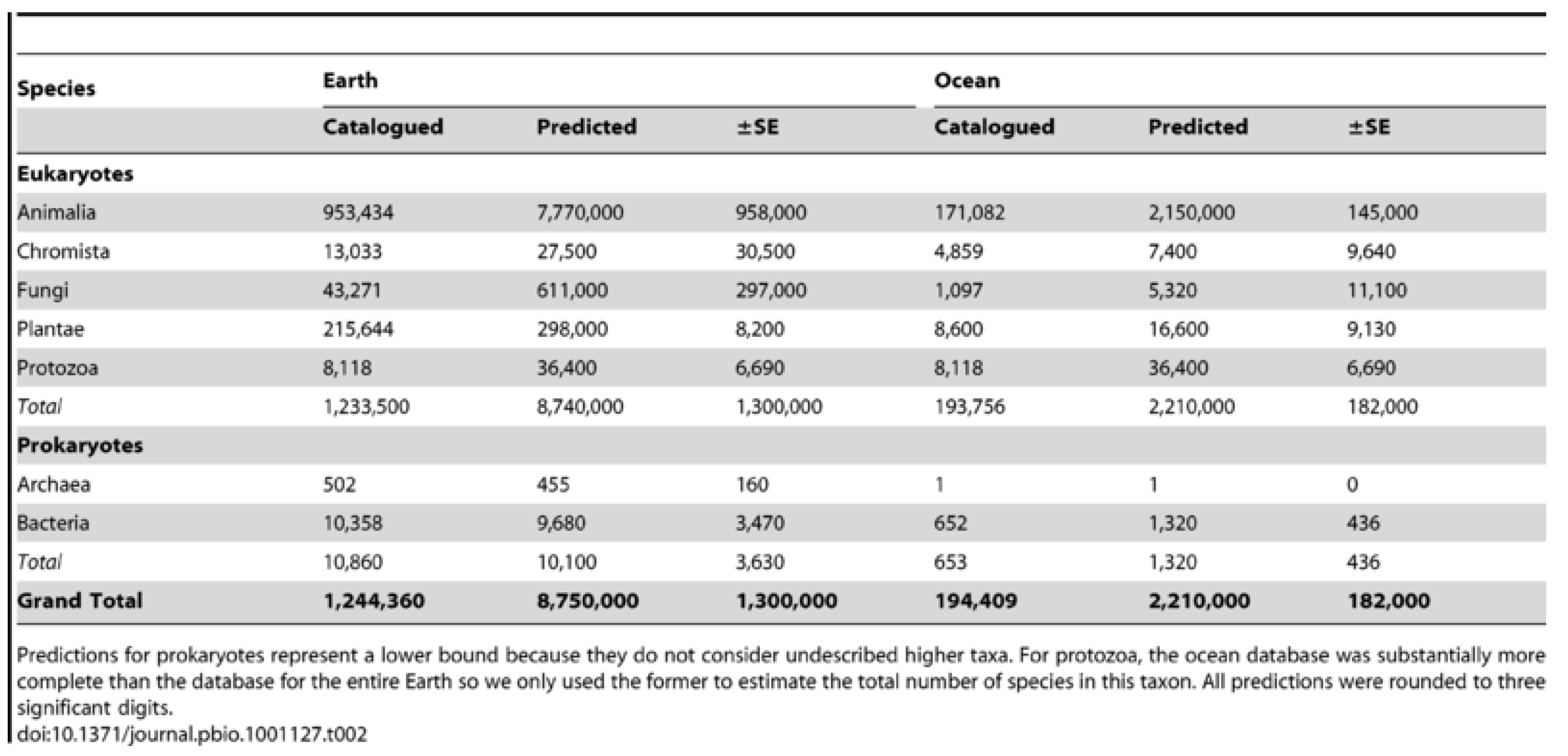
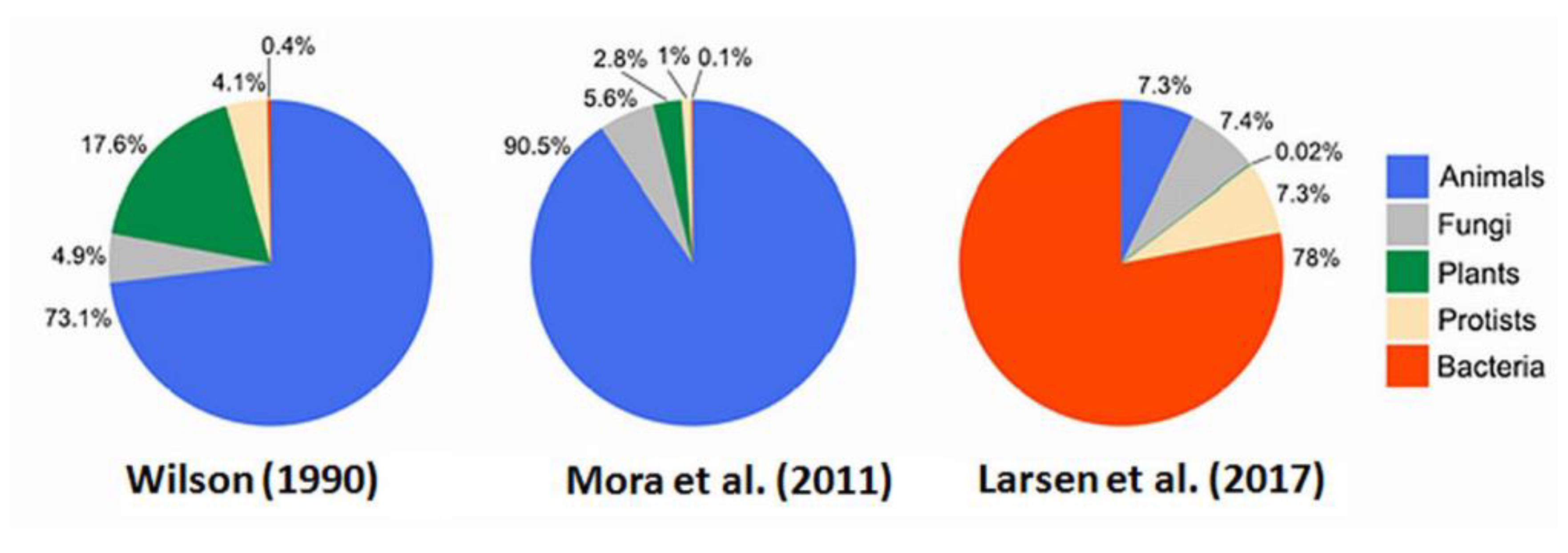
While about two million species have been formally described, global biodiversity recently revised to consider the unique symbionts and parasites of animals produced a new “
pie of life” of up to two billion species
in toto [
92] (cf.
Figure 15), a thousand times increase. Some other estimates using scaling laws to predict species go as high as a trillion taxa when all virus and microbes are tallied, e.g., Locey & Lennon (2016) [
93]. Any or all of these estimates if based upon the ‘flat-Earth’ land model require up-scaling for terrain, relief, etc., as is proposed herein.
3.6. Topsoil Resource
Returning to the initial questions about the Earth’s organic, microbe-rich topsoil. It is vitally important to determine and to conserve this limited resource or, as Darwin (1881: 39) [
31] has it in his swansong book on earthworms: “
The vegetable mould [=topsoil humus] which covers, as with a mantle, the surface of the land”. Soils occupy ~81% of land that is not (yet) extreme desert, rock, sand, ice, or waterlogged (19%) (Jackson et al. 1997: tab. 2) [
37], and its topsoil fragility is as visualized (
Figure 16).
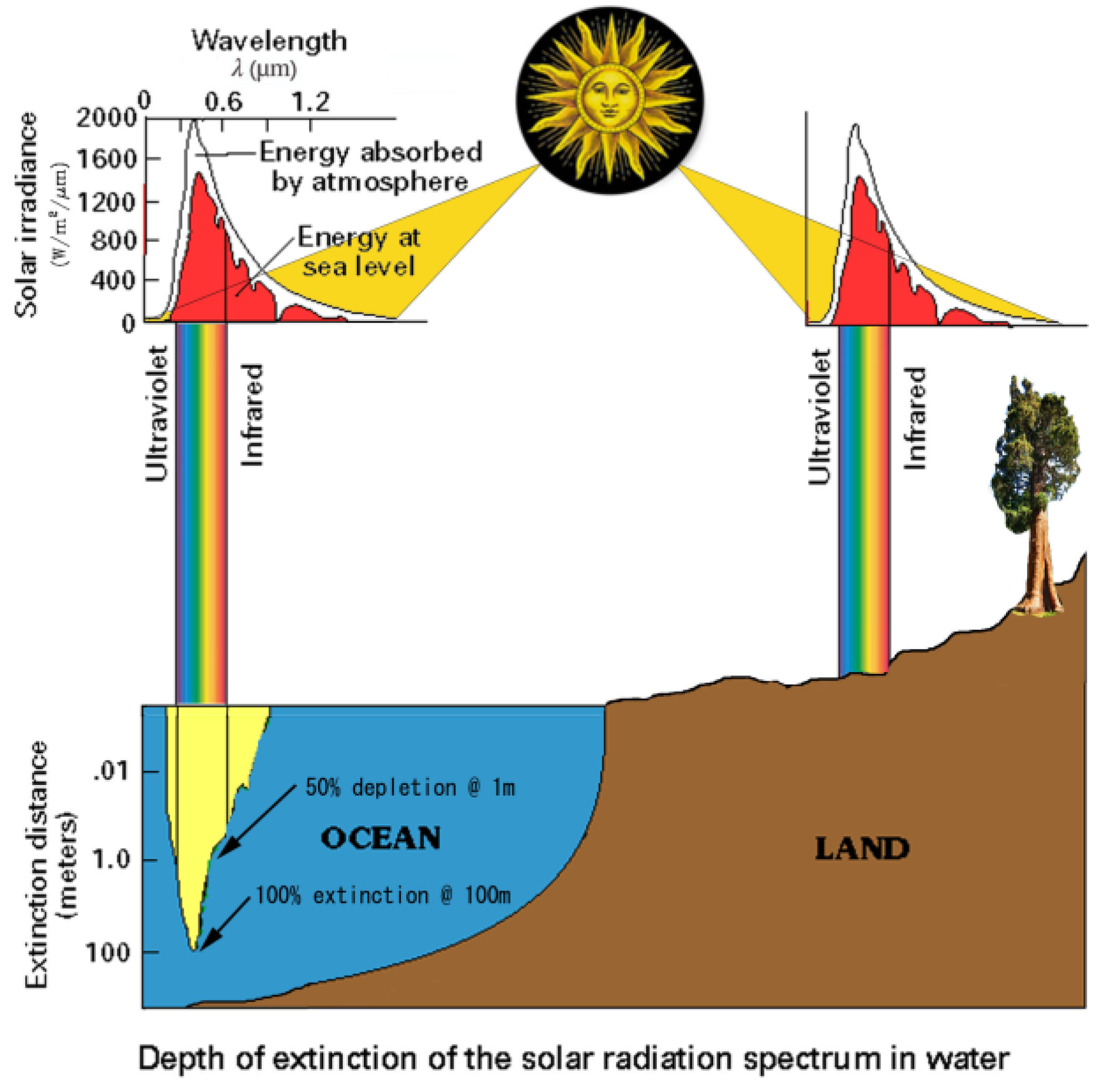
The surface of the Earth is primarily composed of an interface between three essential components, which, in order of volume and levity (antonym of density), are: air, water, and soil that together support abundances of biodiversity in the reverse order. The superficial topsoil that covers all habitable surfaces of the land as a moist, living, breathing skin that manifestly has the highest density and least volume of the three, but overwhelmingly supports the greatest productivity and biomass. The oceans are relatively depauperate, despite moderate volume. The atmosphere has the largest volume with the lowest (negligible) productivity and biomass, much of it transitory: e.g., seeds, insects, spiders, and other aeronauts (volant animals), including cavernicolous bats, microbes, and occasional flying-fish/squid. As well as biota, there is material exchange between these elements in the soil’s moisture and aeration, the silt and (low levels of) dissolved gasses in water, and the humidity and dust in the air. The Sun’s incident visible spectrum energy (for photosynthesis) is depleted by about 25% in the atmosphere, the remainder rapidly reduced by 50% at −1 m and completely extinguished at −100 m depth in salty seawater, whilst on land it is variously absorbed or reflected by plants. Sunlight barely penetrates the superficial soil and litter layers, which is why land plants strive to compete by elevation and extension with the giant
Sequoia reaching up to 100 m skywards, while its roots and symbiotic VAM fungi may extend equally deep earthwards (
Figure 17).
Clinging to land, autotrophic biofilm, or biocrust contributions to productivity at smaller scales are mostly unquantified. Values [
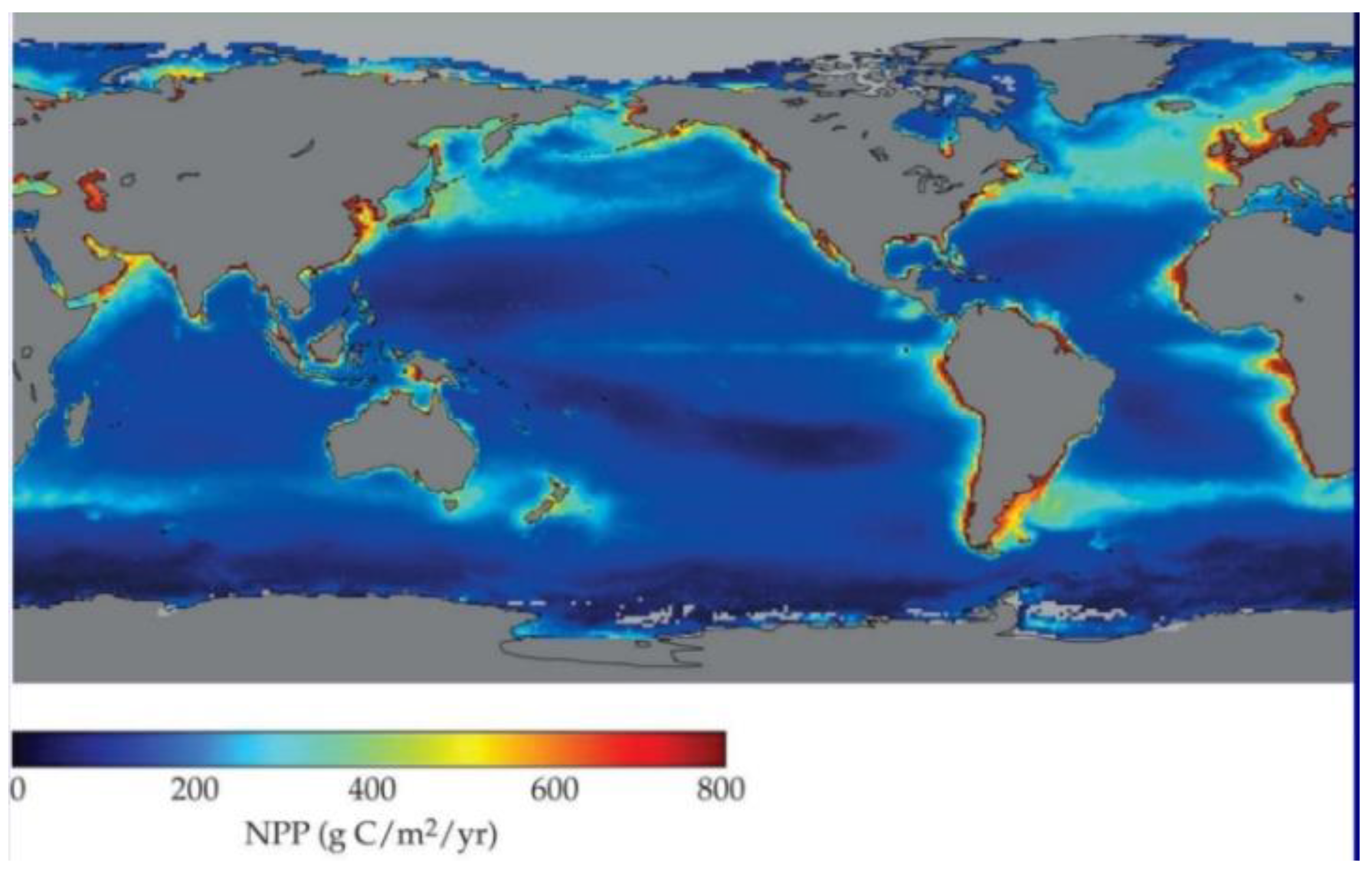
81] of 5 Gt C for ‘cryptogamic covers’, here upped to 10–20 Gt, are higher than the total biomass of mangroves (4 Gt C), seagrasses (0.1 Gt C), and at least double upper estimate of global standing stock of all marine microalgae taxa (0.0075–2.55 Gt C) (cf.
Figure 3 of NPP).
3.7. NPP
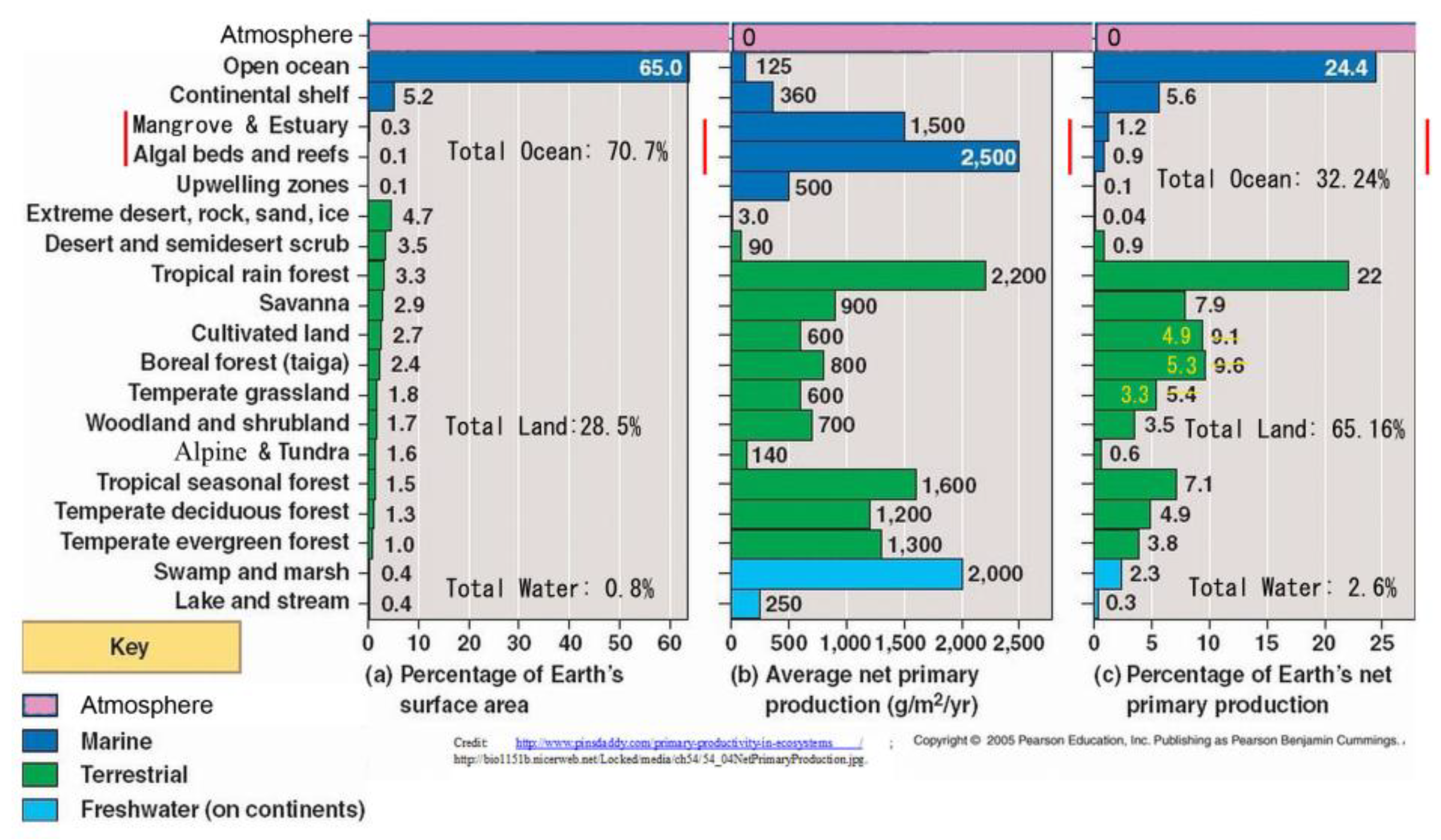
Marine productivity is minor and mainly coastal (e.g., in rockpools), with most open ocean a desolate ‘wet desert’ (
Figure 18).
Topsoil naturally relates to net primary productivity (NPP) with land’s contribution, until now put at somewhere around 45–68% (cf.
Table 2,
Figure 3), yet with correct terrain/topsoil relief factors this would be increased possibly by two or four (or maybe more) to total over 218 Gt C on land. This represents a minimum productivity ratio of soil : sea as 4 : 1 or 80% vs. 20% (
Table 11).
This table shows that NPP per annum has apparently been doubled from 48 Gt to 99 Gt, then up to 170 Gt. Each time with more refinement for the land contribution. The current study continues this trajectory to yield total values of >270 Gt/yr (81% from 30 Gha land), albeit such conclusion requires practical, on-the-ground confirmation. Consideration of finer soil detail and of biocrusts may allow a higher productivity total of 488 Gt C/yr (89% from 60 Gha land).
Pertinent to this are calculations of land productivity per unit area from ecological quadrats that may need to be revised upwards, by ~1–5%, to account for terrain slope/relief (
Appendix A). This too applies to earthworm surveys, conventionally tied to a flat 1 m
2 metric; these too may require a 1–5% increase, but this is minor consideration to their doubling for more refined land surfaces.
Getting to the crux of the Net Primary Productivity (NPP), carbon sequestration, and climate change issue, a recent report stated that: “
At a certainty level of 75%, soil C mass will not change if CO2-induced increase of NPP is limited by nutrients” [
71]. The present paper increases soil C mass by increasing soil area/volume, whereas, to the conventional, but problematical, agrichemical advocates this certainty statement would imply that even more synthetic Nitrogen and other chemicals need to be added to soils (cf.
Figure 2). To agroecology aficionados, the same statement implies a need to recycle all organic wastes back to the soil to “
close the circle”, preferably with more rapid and enhanced nutrient benefits of earthworm vermin-composting, in order to fulfill what Sir Albert Howard (1945) [
95] called the ‘Law of Return’.
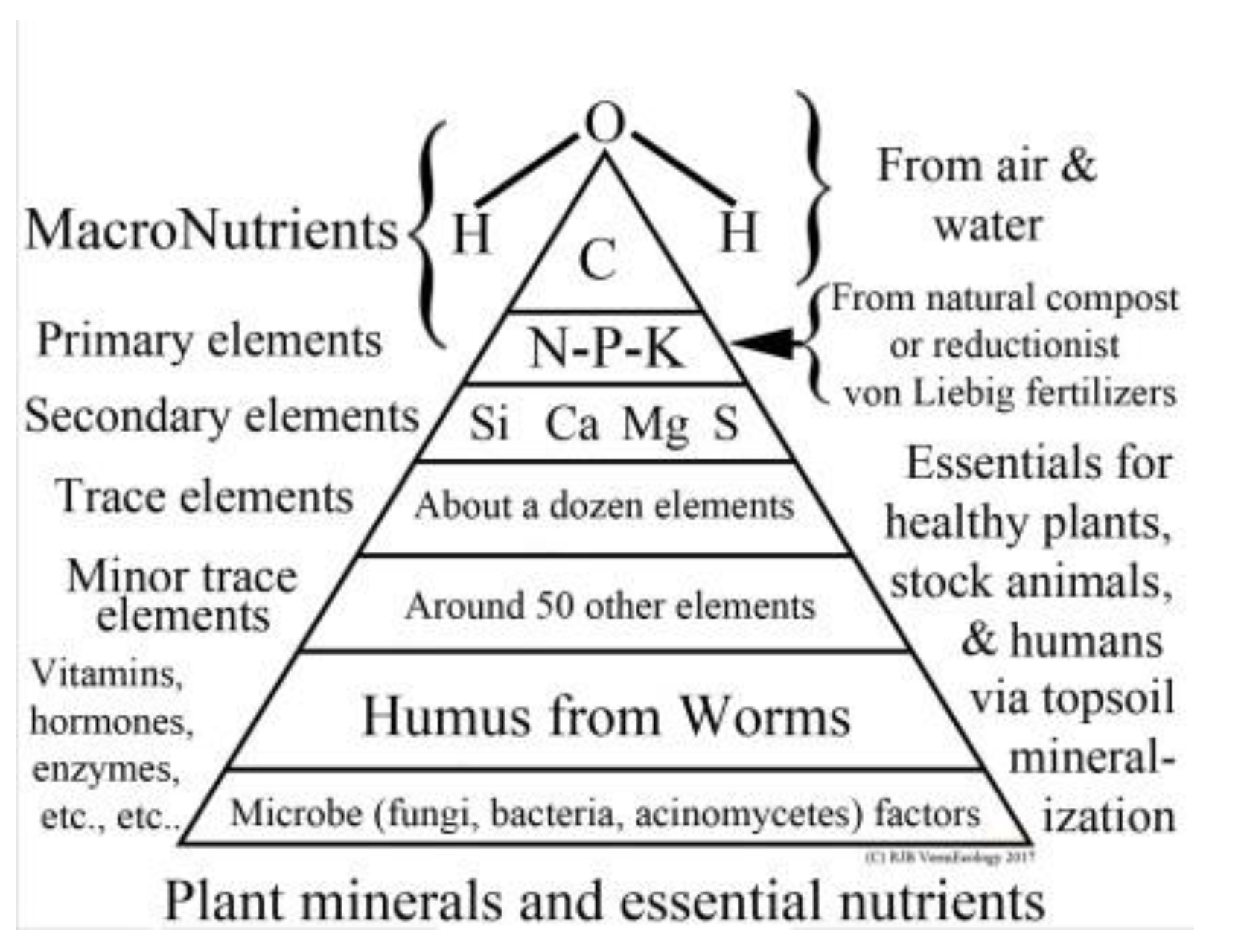
Spontaneous generation has long been debunked, and, similarly, it is not possible for any higher organism to exist without tangible resources as alluded to above:
viz. sunlight, water, gasses, nutrients, symbionts, and habitat. Conventionally, soil nutrients are only considered in terms of simplistic von Liebig agrichemicals N-P-K, whereas the proper plant requirements are complex and mainly carbon based, as shown in Permaculture’s nutrient-pyramid charted below (
Figure 19).
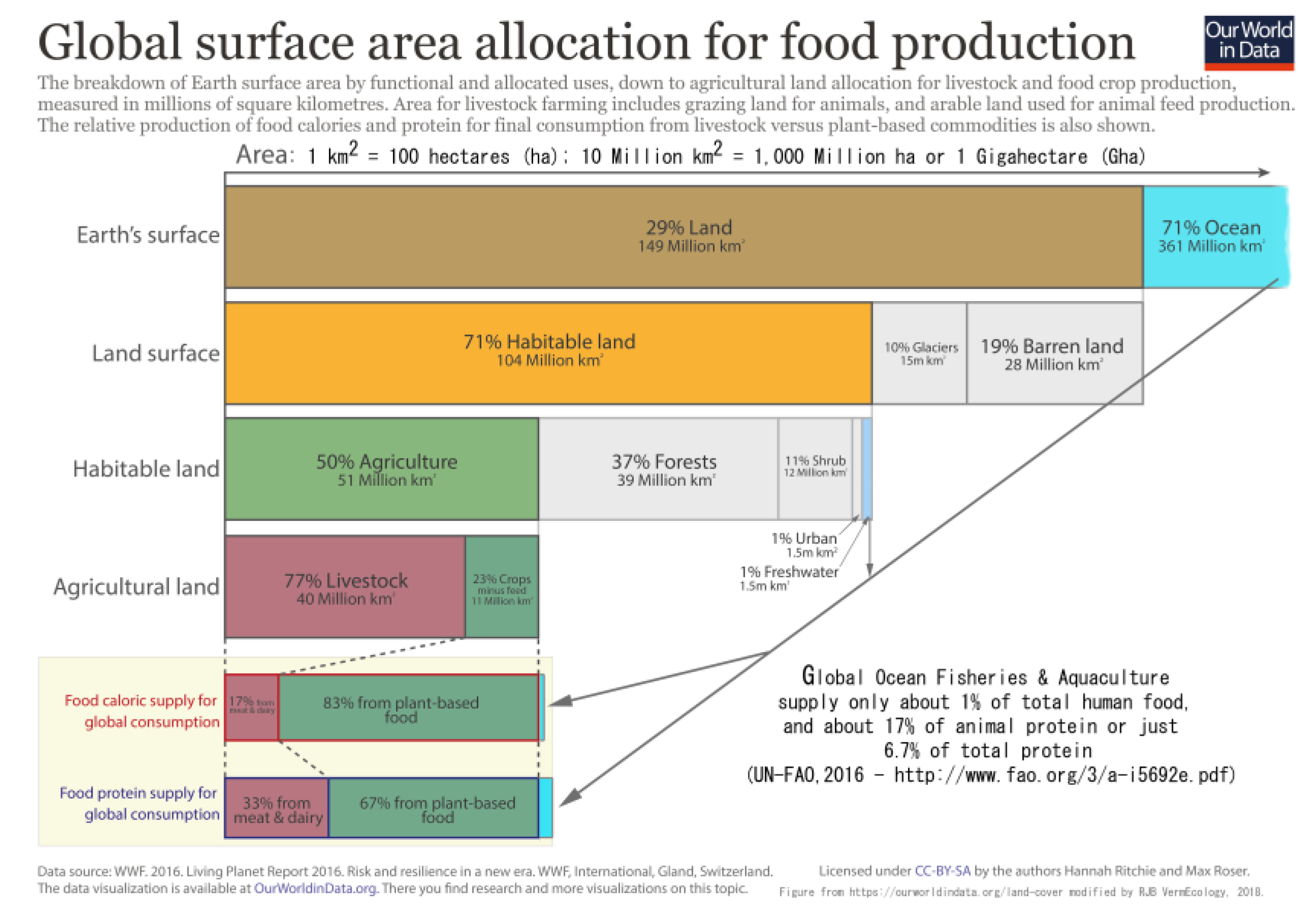
The context of this recycling is that approximately 50% of global soils are managed, often deleteriously, on chemical farms, in burnt or resown pastures and regrowth forests [
84] (cf.
Figure 4). The greatest task facing humanity today is to restore topsoils to their full potential with proper management also to repair or reclaim arid and semi-desert lands using Permaculture methods (Mollison, 1988) [
96].
3.8. Oceans and Space: Diversions and Distractions to the Problems on Earth
Copley (2017) [
97] reveals that the entire ocean floor has now been surveyed to a maximum resolution of around 5 km and that: “
NASA’s Magellan spacecraft mapped 98% of the surface of Venus to a resolution of around 100 m. The entire Martian surface has also been mapped at that resolution and just over 60% of the Red Planet has now been mapped at around 20 m resolution. Meanwhile, selenographers have mapped all of the lunar surface at around 100 m resolution and now even at seven metre resolution”. For Earth, global data is available from the 2000 Shuttle Radar Topography Mission (SRTM) and ASTER Global Digital Elevation Model (
https://asterweb.jpl.nasa.gov/) with a one arc-second, or about 30-m sampling and some datasets have trees and other non-terrain features removed. However, where is the compiled data for the earth beneath our feet?
For bathymetry, a surface of 36.066 Gha has a seabed at 2–20-km resolution of 36.138 Gha (Costello et al., 2010: tab. 1) [
98]. These authors claim this is important as it somehow relates to ocean fisheries that absolutely supply just <0.5% of human food (the other 0.5% mainly from freshwater aquaculture) [
99] (cf.
Figure 4). Nevertheless, only the surface of the ocean is oxygenated and exposed to sunlight, thus bathymetry is a completely irrelevant diversion, as are other planets’ topographies, for calculations of primary productivity and biota here on Earth upon which oceanographers and astronomers entirely depend for their survival, as does everyone else. Moreover, marine scientists are unequivocal that the ocean surface does not include the seafloor as they universally quote its surface area as 36 Gha, i.e., the flat interface between the water, the air, and the coastline abutment, even allowing them an (ever increasing) high water mark.
Mars misventures and the latest
$10+ billion space telescope (
https://jwst.nasa.gov/about.html) aiming yet again to seek “
life on planets like Earth” seems much lower priorities as compared to the rapidly declining life
on planet Earth of which we yet know but a fraction. The same amount of funding could seed urgently needed Soil Ecology Institutes on each Continent. Similarly, submarine surveys of deep-sea hydrothermal vents costing
$ millions to find just a few new species, which will still be there tomorrow, while essential soil species are being lost to erosion daily. Basic equipment for soil survey is a spade. How justifiable is it to dabble in space or deep oceans when we do not yet know how many earthworm species exist on the eponymous Earth, barely nothing of their ecology or conservation status, and even less of their symbiotic/parasitic co-evolutionaries? When the latest report (IPCC 2018) [
100] gives us just 12 years to act in order to prevent catastrophic change, studies of deep space or the abyss seem irrational, inessential, and unjustifiable funding choices that misdirect talent and resources from critical issues emanating from and solvable only in and on our homeland turf.
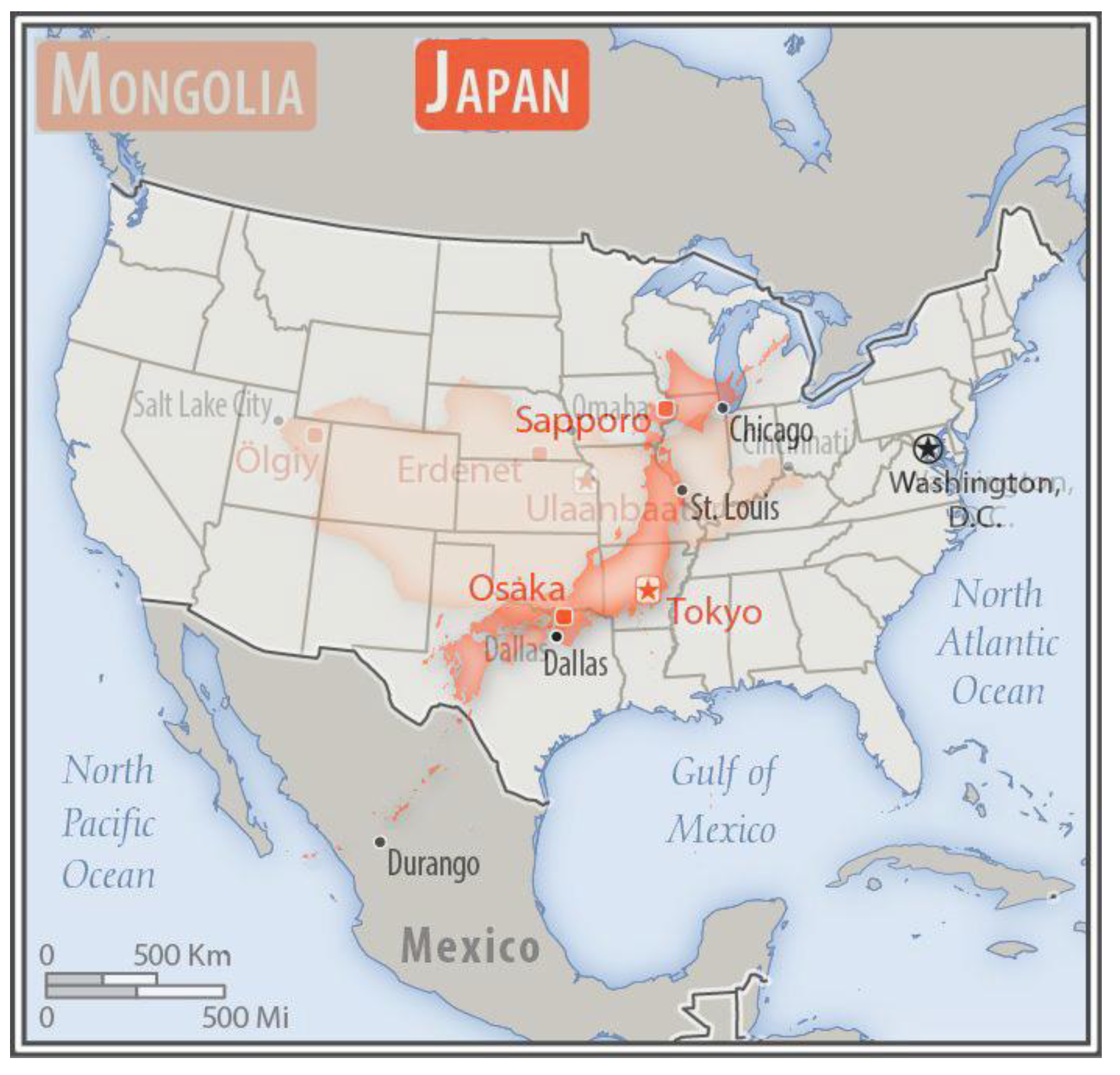
3.9. Worked Example for Samos Island and the Land of the State of Japan
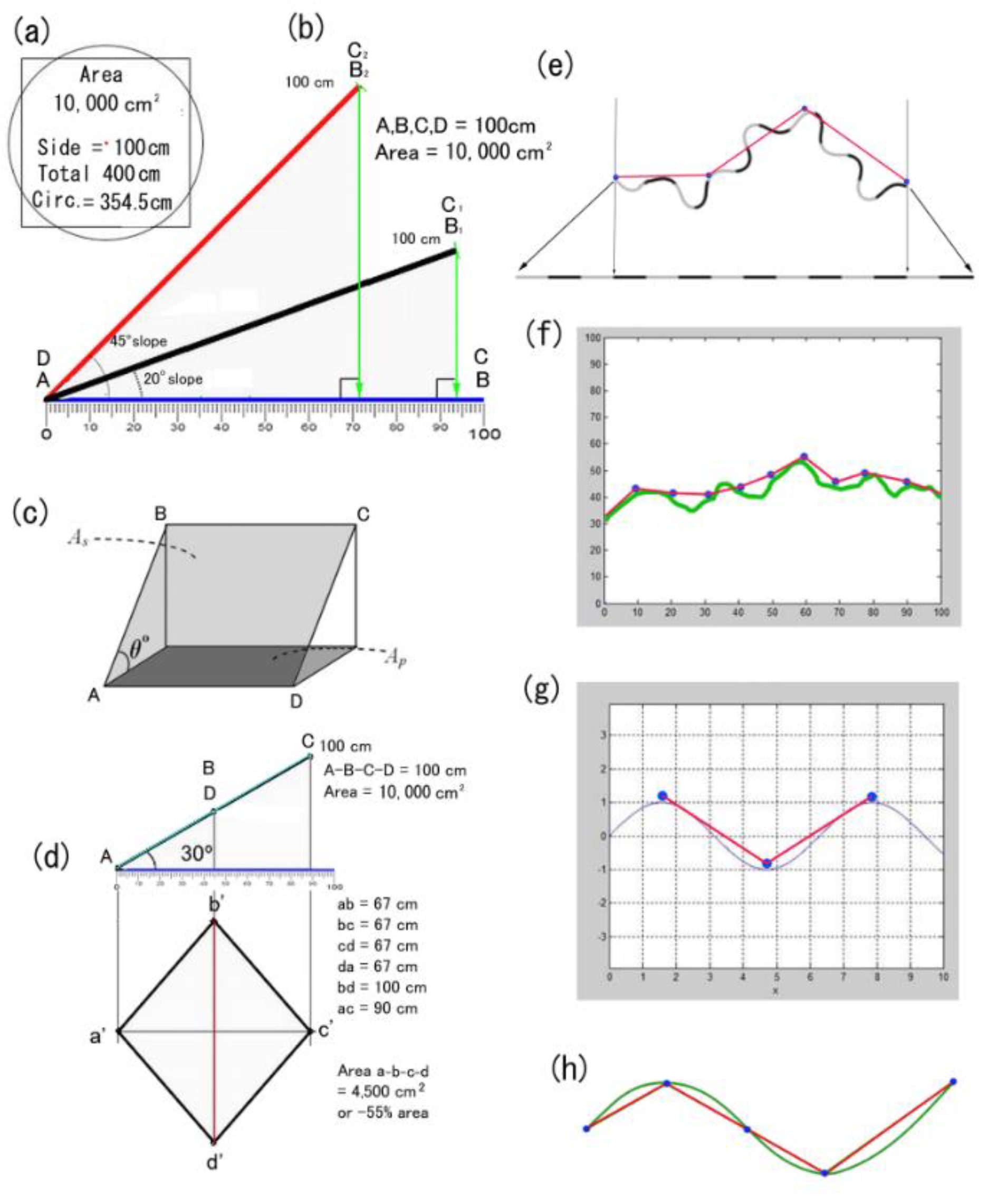
Aristarchus of Samos is credited with the first concept of a spherical Earth revolving around the Sun, an idea later supported by Aristotle on empirical grounds. Appropriately fitting is an attempt to define the topography of Aristarchus’s and Pythagoras’s island of Samos with its central volcanic peak, Vigla, at 1434 m. Its planimetric area of 477.4 km2, which, if circular, would give the island a radius of 12.33 km. Thus a crude approximation using Pythagorean hypotenuse as 12.41 km (= new radius) gives a new surface area 483.8 km² that is only about 1.3% larger at the km scale. However, allowing topographic undulations at one metre, or less, to increase area by 50% totals 716.1 km2 that may itself be doubled for fractal tortuosity at cm scale to about 1432.2 km2 or a 200% increase over original. If hypotenuse/radius is increased 50% to allow for undulating curvatures (i.e., to 18.5), then area is 1075 km2, which, if doubled for relief to 2150 km2, is substantially (350%) larger.
For Japan, [
11] its land area is 36,450,000 ha (0.0365 Gha excluding lakes, e.g., Biwako) and average slope of 6.275% (3.59°). If the flat area was considered a circle with base diameter 6812 units, its hypotenuse of 6850 differs by ~0.6% or about 41 units giving a proper diameter of 6853 and a new area of 36,885,132 ha. This extra 435,132 ha (4351 km
2), which is the least possible, is only a modest 1.2% extra, but an increase in surface area is likely closer to 400% with finer resolutions, as found in the current study. From the worked examples above, its hilly m
2 terrain allows 21.25% extra land (at least) and, because soil occupies most of her land, then by 94% cm
2 tortuosity and then again by mm
2 108.2% micro-relief. This gives Japan a practical land of (0.0365 × 1.2125 = 0.044 × 1.94 = 0.085 × 2.082 =) ~0.17 Gha, or × 4.7, which is larger than Mongolia’s flat surface area that is in the realm of 0.15 Gha before its own required readjustments (
Figure 20).
3.10. Flaws in Un-Flattening the Earth?
Possible flaws in this land surface argument are that the estimation of quadrupled land area may be excessive, or it may be an underestimation depending upon what scale is chosen. The question is why nobody knows this basic data about Earth? Certainly, the present IPCC or NASA/NOAA values are wrong. Other criticisms may be that Landsat and other satellites, if set to measure perpendicular/planimetric values, make terrain less relevant. Because land productivity calculation is more difficult when compared to ocean or atmosphere budgets, IPCC [
101] estimates soil carbon contributions based upon emissions minus atmospheric and oceanic uptake. The residual difference is reasonably ascribed to the land that appears a quite valid method and the ‘missing sink’ discrepancy easily attributed to underestimation of the sub-soil components. Carbon sink calculations when ascribed to biomes may also be artificial due to boundary differences affecting relative % (which may be independent of topography). For example, FAO [
102] have grasslands covering 40.5% of land comprised of woody savannah/savannah (13.8%), open/closed shrub (12.7%), non-woodly grassland (8.5%), and tundra (5.7%); whereas, other sources separate these biomes. Calculations relating to carbon stored and released (either eroded or respired) from agriculture, forestry, and other land-use changes, primary productivity and biodiversity studies, however, certainly do need to employ topography details down to cm or mm scale for true tallies.
Regarding soil biomass, as carbon values are drawn from loss-on-ignition (LOI) or Walkley- Black, they may include much of the microbiota (although certainly not the larger megadrile earthworms nor sieved roots/hyphae), whereas microbial measurements often take smaller samples and either extract DNA or use plate cultures to estimate biomass and diversity. Thus, the intermesh of chemical and biotic factors may unintentionally overlap to overstate total carbon in SOM humus.
Conversely, when soil carbon or microbes, or any other organisms, are ascribed to a ‘flat-Earth’ biome then the calculations are invariably and undeniably wide underestimations of both soil depth and of probable land surface area that they occupy both in reality or potentially.




























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}