
Glow on Sharks: State of the Art on Bioluminescence Research
by
 , , and
, , and
Laurent Duchatelet
1,* ,
,
Julien M. Claes
1,
Jérôme Delroisse
2 ,
,
Patrick Flammang
2 and
Jérôme Mallefet
1,*
1
Marine Biology Laboratory, Earth and Life Institute, University of Louvain—UCLouvain, 1348 Louvain-la-Neuve, Belgium
2
Biology of Marine Organisms and Biomimetics Unit, Research Institute for Biosciences, University of Mons—UMONS, 7000 Mons, Belgium
*
Authors to whom correspondence should be addressed.
Oceans 2021, 2(4), 822-842; https://doi.org/10.3390/oceans2040047
Submission received: 1 September 2021
/
Revised: 25 November 2021
/
Accepted: 3 December 2021
/
Published: 17 December 2021
(This article belongs to the Special Issue Current Advances and Challenges in Ocean Science—Feature Papers for the Founding of Oceans)
Abstract
:This review presents a synthesis of shark bioluminescence knowledge. Up to date, bioluminescent sharks are found only in Squaliformes, and specifically in Etmopteridae, Dalatiidae and Somniosidae families. The state-of-the-art knowledge about the evolution, ecological functions, histological structure, the associated squamation and physiological control of the photogenic organs of these elusive deep-sea sharks is presented. Special focus is given to their unique and singular hormonal luminescence control mechanism. In this context, the implication of the photophore-associated extraocular photoreception—which complements the visual adaptations of bioluminescent sharks to perceive residual downwelling light and luminescence in dim light environment—in the hormonally based luminescence control is depicted in detail. Similarities and differences between shark families are highlighted and support the hypothesis of an evolutionary unique ancestral appearance of luminescence in elasmobranchs. Finally, potential areas for future research on shark luminescence are presented.
Keywords:
shark; luminescence; Etmopteridae; Dalatiidae; Somniosidae; photophore; hormonal control; counter-illumination1. Introduction
Bioluminescence is the ability of living organisms to produce visible light [1]. Mention of this phenomenon dates back to antiquity with the description of “cold light” by Aristotle in his book “De Anima”. Many centuries later, Charles Darwin, on board the Beagle, witnessed and described light in water as “milky sea” in his logbook. The first studies demonstrating mechanisms underlying bioluminescence appeared in 1667 with Robert Boyle, who depicted the oxygen requirement for luminescence production.
Bioluminescence is the result of a spontaneous exergonic chemical reaction involving the oxidation of a luciferin catalyzed by a luciferase [2], which produces a transitory excited state that finally relaxes by emitting a photon with oxyluciferin as final product [3,4,5]. Luminous systems involve either luciferase and luciferin as separate components or a complex molecule called “photoprotein” comprising a preoxidized luciferin and a luciferasic activity [5]. Luciferins are often common to different taxa while luciferases are thought to be typically species-specific [6] but exceptions to this rule have been recently highlighted [7,8,9]. Similar luciferases, first described in species from same phyla (within a clade), are now found between phylogenetically distant species supporting the existence of multiphyletic distribution of these luciferases [7,8,9].
Rare in terms of species number, luminescence nevertheless arose in over 700 genera from 13 phyla covering all kingdoms, except plants and archaea, e.g., [6,10,11]. This phylogenetic diversity results from numerous independent evolutions of light-producing capability—in fact, it is estimated that bioluminescence arose independently more than 90 times during evolution [1,3,9,10,12,13], which suggests that luminescence is of paramount ecological importance, but also that the acquisition of light emission capability is a relatively easy and quick process [1].
Eighty percent of luminous taxa inhabit marine environments, from coastal shallow waters to abyssal depths, and are mainly found in bacteria, protists, ctenophores, cnidarians, annelids, mollusks, chaetognaths, crustaceans, echinoderms, tunicates, and fishes [1,6,14,15]. Within the water column, the mesopelagic zone (i.e., 200–1000 m depth) has been estimated to host a high proportion of luminous organisms, e.g., 70% of mesopelagic bony fishes appear to be luminous [16]. Comparatively, terrestrial and freshwater luminous animals are only represented by earthworms, snails and limpets, fireflies, beetles, and some insect larvae, e.g., [17,18,19,20,21,22,23].
Although observations of shark luminescence have been reported for almost two centuries [24], the first research projects dedicated to shark luminescence were only initiated in 2005 and focused on the study of the ecological functions and physiological control of the photophores of a single species, the velvet belly lanternshark, Etmopterus spinax [25]. Since then, shark luminescence research has flourished, with detailed phylogenetical, ecological, histological, and physiological studies now available for numerous species, e.g., [26,27]. By synthesizing the findings of those studies, this article aims to provide not only a holistic view of current shark luminescence knowledge but also perspectives for future research.
2. Luminous Shark Diversity
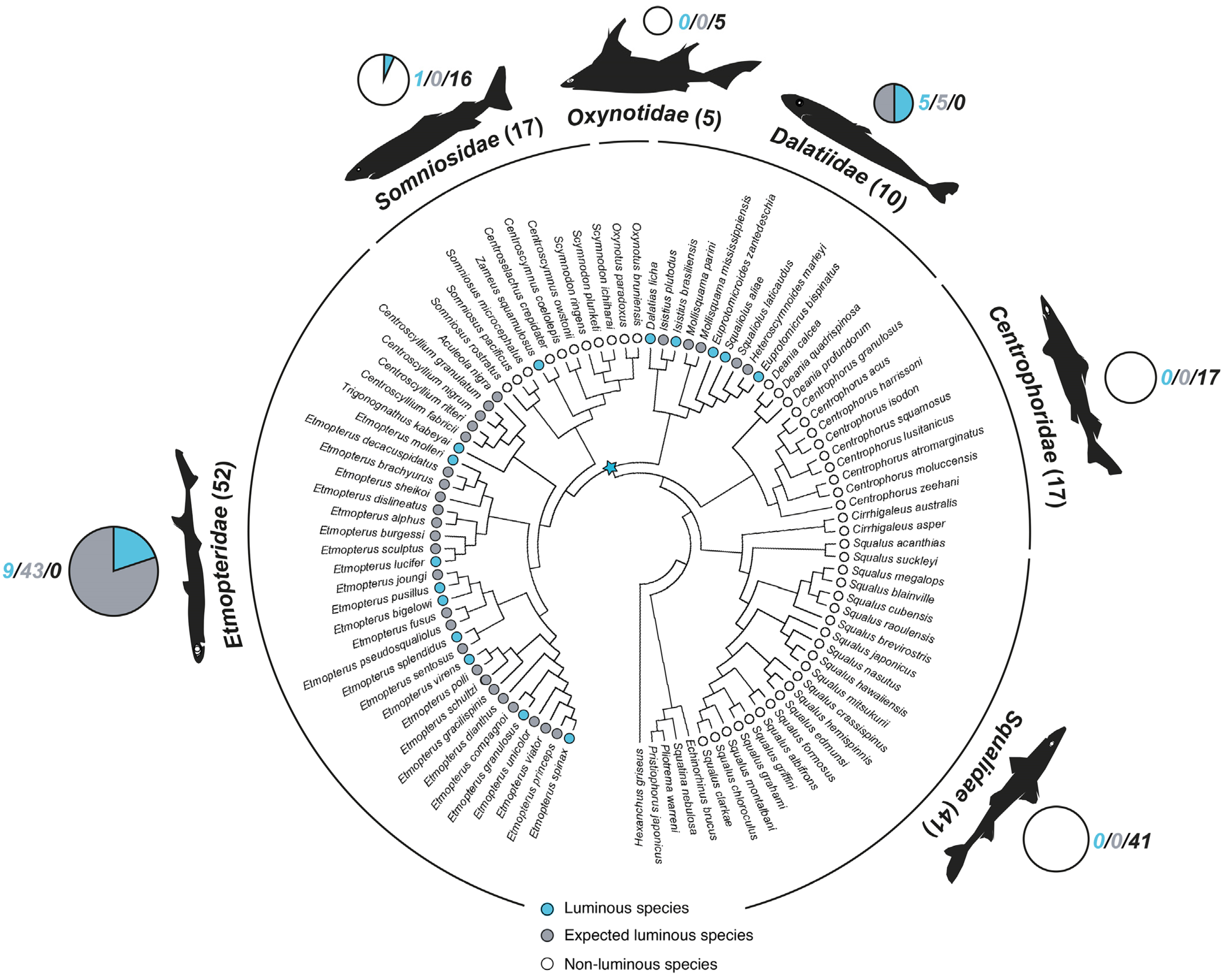
Among cartilaginous fishes, only sharks have evolved the ability to emit light. Indeed, no report of bioluminescence exists for ratfishes (Chimaeriformes) apart from the mention of a luminous fluid on the rabbit fish, Chimaera monstrosa, in 1810 interpreted as coming from luminous bacteria due to the degradation of the harvested organism [28]. In addition, the anecdotal observation of putative photogenic organs in the deep-sea dark blind ray, Benthobatis moresbyi [29], has not been confirmed by more recent and detailed morphological studies [30,31]. Bioluminescence in sharks appears currently restricted to Squaliformes, where only three families (Dalatiidae, Somniosidae, and Etmopteridae) contain luminescent representatives (Figure 1). Indeed, although bioluminescence has once been suggested for the specific supralabial white band of the megamouth shark, Megachasma pelagios, recent work invalidated this assumption [32]. Although the luminescence reported for the species that belong to Etmopteridae and Dalatidae was initially thought to have been acquired independently [33,34], the recent discovery of luminescence from a somniosid, the velvet dogfish, Zameus squamulosus [35,36], now strongly suggests that the acquisition of luminescence capability in sharks represents a single evolutionary event, which occurred during the deep-sea radiation of Squaliformes at the end of the Cretaceous [35]. Although fossil studies estimate the Etmopteridae radiation around 90 million years ago [37], molecular data presents a separation of Etmopteridae from other Squaliformes during the Upper Cretaceous (i.e., 65–90 million years ago) [35,38]. The Dalatiidae family radiated later, during the Paleocene after the Cretaceous/Paleocene mass extinction, 65–105 million years ago, when they substituted extinct marine reptiles and fishes in the epipelagic fauna before reaching the deep sea [34,35,39].
Interestingly, the photogenic structures appear ubiquitous in Etmopteridae (four genera: Trigonognathus, Aculeola, Centroscyllium and Etmopterus; 52 species) and Dalatiidae (seven genera: Dalatias, Isistius, Mollisquama, Euprotomicroides, Squaliolus, Euprotomicrus and Heteroscymnoides; 10 species). Nevertheless, Z. squamulosus is the only somniosid shark known to possess photophores (Figure 1; Table S1). In parallel, luminescence has been observed in live in 15 species only, however, covering most clades, i.e., the blurred smooth lanternshark, Etmopterus bigelowi [40]; the southern lanternshark, Etmopterus granulosus [41]; the blackbelly lanternshark, Etmopterus lucifer [41]; the slendertail lanternshark, Etmopterus molleri [42,43]; the smooth lanternshark, Etmopterus pusillus [44]; E. spinax [45,46]; the splendid lanternshark, Etmopterus splendidus [47]; the green lanternshark, Etmopterus virens [40]; the smalleye pygmy shark, Squaliolus aliae [43,48]; the kitefin shark, Dalatias licha [41]; the taillight shark, Euprotomicroides zantedeschia [49]; the pygmy shark, Euprotomicrus bispinatus [50]; the cookiecutter shark, Isistius brasiliensis [51,52]; Z. squamulosus [36]; the viper dogfish, Trigonognathus kabeyai (Mallefet, unpublished data); Figure 1 and Figure 2; Table S1. In addition, expected luminous species are encountered in Etmopteridae and Dalatiidae, based on the presence of photophore structures and/or flank marks in the holotype description [33,35,36,38,40,41,42,43,44,45,46,47,48,49,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90]; Table S1. This led to a total number of 62 luminous sharks, i.e., ~11% of currently described species (550 in total [91]), while in comparison, luminescence is thought to have appeared in only ~5% of bony fishes [12].
Figure 1.
Shark luminescence distribution within Squaliformes families based on published phylogenies [35,38,53,92,93,94,95,96]. Circles inside the tree represent the luminous (blue), expected luminous (gray), and non-luminous (white) status of each represented species. Statuses are based on in vivo pictures, physiological studies (luminous), the presence of photophores or flank marks (expected luminous), and none of these criteria (non-luminous) (see Table S1). Circles outside the tree, scaled to the total number of species (number in brackets) within a given family [54,91], indicate the proportion of luminous (blue), expected luminous (gray), as well as non-luminous (white) species. For each family, total number of luminous/expected luminous/non-luminous species are given next to the outside circle. Blue star indicates the expected origin of luminescence in sharks.
Figure 1.
Shark luminescence distribution within Squaliformes families based on published phylogenies [35,38,53,92,93,94,95,96]. Circles inside the tree represent the luminous (blue), expected luminous (gray), and non-luminous (white) status of each represented species. Statuses are based on in vivo pictures, physiological studies (luminous), the presence of photophores or flank marks (expected luminous), and none of these criteria (non-luminous) (see Table S1). Circles outside the tree, scaled to the total number of species (number in brackets) within a given family [54,91], indicate the proportion of luminous (blue), expected luminous (gray), as well as non-luminous (white) species. For each family, total number of luminous/expected luminous/non-luminous species are given next to the outside circle. Blue star indicates the expected origin of luminescence in sharks.

Figure 2.
Shark luminescent patterns. Squaliform cladogram with taxonomic grouping where photophores/flank markings (gray shade) or live luminescence (blue shade) have been observed [35,36,40,41,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,97,98] and spontaneous luminescence of representative shark species lateral view for D. licha, S. aliae, E. bispinatus, excreted bioluminescent fluid for E. zantedeschia (blue bubbles on the cladogram), and ventral view for the others]. D, Dalatiidae; E, Etmopteridae; FM, flank markings; S, Somniosidae. Indicative scale bars, 5 cm. Shark drawings are modified from [34,74]. Photographs courtesy by D. Perrine (E. bispinatus), T. Raczynski (E. zantedeschia [49]) and J. Mallefet (other species; [36,41,65], Mallefet, unpublished).
Figure 2.
Shark luminescent patterns. Squaliform cladogram with taxonomic grouping where photophores/flank markings (gray shade) or live luminescence (blue shade) have been observed [35,36,40,41,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,97,98] and spontaneous luminescence of representative shark species lateral view for D. licha, S. aliae, E. bispinatus, excreted bioluminescent fluid for E. zantedeschia (blue bubbles on the cladogram), and ventral view for the others]. D, Dalatiidae; E, Etmopteridae; FM, flank markings; S, Somniosidae. Indicative scale bars, 5 cm. Shark drawings are modified from [34,74]. Photographs courtesy by D. Perrine (E. bispinatus), T. Raczynski (E. zantedeschia [49]) and J. Mallefet (other species; [36,41,65], Mallefet, unpublished).

3. Ecology of Shark Luminescence
Deciphering the ecological functions of luminescence from elusive animals such as deep-sea sharks is extremely challenging. Indeed, field observations are scarce and unbiased lab experiments proved to be difficult to perform. As a matter of fact, most functions formulated over years regarding the function of shark luminescence remain undemonstrated, mainly due to the difficulties of observing and collecting these rare and elusive organisms to conduct ethological studies on the function of their luminescence. Fortunately, however, detailed analyses of photophore distribution (luminescent “patterns”) as well as physical characteristics (intensity, wavelength and angular distribution) and kinetics of luminescence coupled to physical models for pelagic vision and molecular phylogenetic analyses, now allow us to draw a clearer picture of the adaptive benefits of luminescence in sharks.
In this context, counterillumination, i.e., a camouflage technique used by midwater organisms cloaking their silhouette from upward-looking organisms using a ventral glow mimicking downwelling sunlight, e.g., [99,100,101,102,103], is probably the primary function of shark luminescence, for both defensive and predatory purposes [41,65]. Indeed, shark photophores are predominantly situated on the ventral surface area (Figure 2 and Figure 3) and produce a light whose color (wavelength) which is similar to that found in coastal (blue–green) and oceanic (blue) environment (Figure 3b; Table 1). Moreover, the intensity and angular distribution of light emission have been correlated to capture depth residual light parameters in three species: E. spinax [103], E. splendidus [65], and S. aliae [65].
Figure 3.
Shark luminescence functions. (a) Composite pictures of E. molleri and E. spinax luminescence patterns. Spontaneous dorsal, lateral and ventral luminescence from E. molleri, with key photophores areas highlighted. Circular inserts represent spine-associated photophores (SAPs) of the dorsal fins of E. spinax [104]; these photophores are absent in E. molleri (which instead has spine base-associated photophores; [105]). (b) Angular distribution of dalatiid (S. aliae; mid body) and etmopterid (E. spinax; mid body and lateral flank marks) luminescence as well as downwelling sunlight; the perfect match observed for E. spinax is achieved thanks to a centripetal change in photophore orientation modified from [65,103]. (c) Simplified bioluminescent shark phylogeny providing an overview of demonstrated, experimentally supported and putative functions of shark luminescence [27,41,74,75,104,105,106,107,108], Claes and Duchatelet, unpublished observation. AP, associated photophores; FP, frontal photophores; LZ, luminous zone; OP, ocular photophores; SAP1, spine-associated photophores of 1st dorsal fin; SAP2, spine-associated photophores of 2nd dorsal fin; SBAP, spine base-associated photophores. Scale bar, 5 cm. Photographs by J. Mallefet.
Figure 3.
Shark luminescence functions. (a) Composite pictures of E. molleri and E. spinax luminescence patterns. Spontaneous dorsal, lateral and ventral luminescence from E. molleri, with key photophores areas highlighted. Circular inserts represent spine-associated photophores (SAPs) of the dorsal fins of E. spinax [104]; these photophores are absent in E. molleri (which instead has spine base-associated photophores; [105]). (b) Angular distribution of dalatiid (S. aliae; mid body) and etmopterid (E. spinax; mid body and lateral flank marks) luminescence as well as downwelling sunlight; the perfect match observed for E. spinax is achieved thanks to a centripetal change in photophore orientation modified from [65,103]. (c) Simplified bioluminescent shark phylogeny providing an overview of demonstrated, experimentally supported and putative functions of shark luminescence [27,41,74,75,104,105,106,107,108], Claes and Duchatelet, unpublished observation. AP, associated photophores; FP, frontal photophores; LZ, luminous zone; OP, ocular photophores; SAP1, spine-associated photophores of 1st dorsal fin; SAP2, spine-associated photophores of 2nd dorsal fin; SBAP, spine base-associated photophores. Scale bar, 5 cm. Photographs by J. Mallefet.


Table 1.
Shark luminescence color.
| Species | Photogenic Structure | Luminescence Color (Wavelength Peak) | References |
|---|---|---|---|
| Isistius brasiliensis | Photophores | Dark blue (455 nm) | [16] |
| Squaliolus aliae | Photophores | Dark blue (457 nm) | [65] |
| Euprotomicroides zantedeschia | Pelvic pouch (fluid) | Dark blue | [49] |
| Dalatias licha | Photophores | Blue | [41] |
| Euprotomicrus bispinatus | Photophores | Blue | This study |
| Etmopterus splendidus | Photophores | Blue (476 nm) | [65] |
| Etmopterus molleri | Photophores | Blue (477 nm) | [65] |
| Etmopterus bigelowi | Photophores | Blue | [40] |
| Etmopterus granulosus | Photophores | Blue | [41] |
| Etmopterus lucifer | Photophores | Blue | [41] |
| Trigonognathus kabeyai | Photophores | Blue | This study |
| Etmopterus spinax | Photophores | Blue-green (488 nm) | [65] |
| Zameus squamulosus | Photophores | Blue-green | [36] |
| Etmopterus virens | Photophores | Green | [40] |
| Etmopterus pusillus | Photophores | “Whitish” | [44] |
Interestingly, “water box” experiments suggested that E. spinax, contrary to other counter-illuminating animals, was not able to adapt rapidly the intensity of its luminescence by more than one order of magnitude in response to changing light regime [103]. Hence an “isolume follower” hypothesis was formulated, stating that sharks move up and down in the water column to remain cryptic [103]. This has been further supported by a large comparative study showing that daytime capture depth of luminescent sharks could be predicted from their ventral photophore cover (e.g., the percentage of the ventral surface area covered with photophores), which varies from 2% in the bareskin dogfish Centroscyllium kamoharai to 56% in the viper dogfish T. kabeyai [65]. In addition, recent studies show that some etmopterid species have a higher swimming speed and muscular enzymatic activities than their non-luminous counterparts living in the same deep environment [109,110]. These findings could be correlated with a greater physiological demand to perform vertical migration to remain camouflaged. The second most widespread and well-supported function of shark luminescence is intraspecific communication. Although Dalatiidae and Somniosidae present “simple” luminescent patterns where photophores follow a dorsoventral density gradient [33,34,36,41,52] (Figure 2 and Figure 3), etmopterid sharks display complex luminescent photophore aggregations on the ventral area, but also on the flanks, the fins, the tail, around the eyes, the spiracles, the gills, and the epidermal tissue surrounding dorsal spines [34,104,105,106,111] (Figure 2 and Figure 3). Since these patterns are species-specific and do not show sexual dimorphism (except photophores associated with primary sexual characters, e.g., male claspers, as explained below [112]), they are often used as taxonomic determination tools [34,54,63,68]. Interestingly the shape of lateral luminous areas (flank markings) appears to be clade-specific [38,65] (Figure 2) and their presence correlates with an increased speciation rate [75] and a moderate predation risk [65]. This, and the ability for those sharks to discern the flank marking shape (and other specific luminous zones) at a biologically meaningful distance (1–3 m, or three to ten body lengths, as indicated by visual modeling [111]) strongly support the idea that etmopterid flank markings represent an exaptation of counter-illuminating photophores to facilitate deep-sea communication [65]. Luminescence could also be used as a mating aid, which allows males to identify females from a distance (since photophores cover male claspers of all species [34]; Figure 3a) and visualize their cloaca and pectoral fins (brighter in etmopterid [34]; Figure 2 and Figure 3a) in the darkness of the deep sea (sharks display internal fertilization during which the male stabilizes itself by biting the female’s pectoral fin [97]). Sexual dimorphism in the luminescence time course of pharmacologically stimulated pelvic photophores from E. spinax further supports this hypothesis [107].
The last luminescence function for which experimental support is available is the aposematism, a mechanism by which an animal advertises potential predators that it is not worth attacking or eating [113,114]. Etmopterid sharks (contrary to dalatiid and somniosid species) have large sharp defensive spines associated with their dorsal fins [97]. In some species, photophores either placed on the edge of the dorsal fins (E. spinax; [104]) or around the base of the spines (E. molleri; [105]), allow these spines to be seen in the dark from a distance by potential predators, and hence potentially work as an aposematic signal as strongly suggested by experimental data [104,105] (Figure 3a–c).
In addition to these relatively well-established functions, researchers have formulated more speculative functional hypotheses regarding the luminescence of some species (Figure 3c). For instance, the luminescent liquid ejected by the pelvic pouch of E. zantedeschia probably works as a defensive “smokescreen” mechanism [49]. Given the morphological similarity of the pectoral glands, an identical function has been suggested for the pocket sharks, Mollisquama spp. [55,74]. The ocular and frontal photophores of E. spinax [103] and the ventral photophore of D. licha [41] could be used as a vision aid (Figure 3c). In addition, the tail of D. licha, S. aliae, Z. squamulosus, T. kabeyai, and Etmopterus species, which is more mobile and brighter than the rest of the ventral surface area, could work as a distracting lure, and hence be analogous to the caudal photophores of myctophid and tubeshoulder fishes [115,116,117]. Conversely, the idea that the dark, photophore-free collar of I. brasiliensis acts as a lure to attract bigger pelagic fishes or marine mammals on which it feeds [51], now seems at the very least dubious given that numerous common preys of this shark species are either filter feeders or top predators for which such a mechanism is useless [65], and that the closely related species, the largetooth cookiecutter shark, I. plutodus, which has a similar diet [97], lacks such a collar [70].
Etmopterid and dalatiid sharks display a set of visual features not found in non-bioluminescent sharks, which strongly supports the idea that their visual system coevolved with their ability to produce light. Aphakic gaps in S. aliae [111] and E. bispinatus Claes, personal observation] and translucent upper eyelid found in etmopterid sharks (genus Trigonognathus and Etmopterus; [111]) probably play a role in counterilluminating, e.g., facilitating perception of downwelling light intensity to ensure the perfect match, similarly to what was recently shown for deep-sea bony fishes [118]. On the other hand, the spectral absorbance (485–488 nm) of etmopterid rod photoreceptor as well as the retinal distribution of ganglion cells of those shark (which appear species-specific) appear finely tuned to detect their own bioluminescence, especially from flank markings [111].
4. Photogenic Structures and Specialized Squamation of Bioluminescent Sharks
Sharks display two types of photogenic structures: photophores (Figure 4), with internal luminescence present in all bioluminescent shark species, and secretory glands ejecting a bioluminescent fluid in the environment (external or secretory luminescence; Figure 4) present in E. zantedeschia (pelvic pouch) and in Mollisquama spp. (pectoral pockets).
Shark photophores are small, reaching a maximum diameter of ~50, ~100, and ~200 µm for Somniosidae, Dalatiidae, and Etmopteridae, respectively [34,36,41,48,52,65,76]. Comparatively, the photophores of bony fishes can reach up to 1 cm [119]. Given their high photophore density (sometimes over ten thousand per square centimeter; [65]), sharks are probably the luminous organisms with the highest number of photophores (a 52 cm TL male E. spinax has been estimated to bear about 440,000 photophores; [34]). The general structure of photophores is similar across families (Figure 4a,c,d).
Figure 4.
Shark photogenic organs. (a) Photophores (modified from [32,34]); (b) secretory photogenic structures: light-producing columnar epithelium from pouch of the taillight shark (modified from [34]; left) and putative light-producing cubic tissue from pocket shark’s pectoral pockets (modified from [74]; right); (c) histological section of I. brasiliensis photophore; (d) histological section of E. lucifer photophore; (e) ultrastructure of E. spinax photophore; (f) ultrastructure of E. spinax photocyte; (g) autofluorescence from E. lucifer photophore; (h) autofluorescence from photocyte vesicle from E. lucifer; (i) autofluorescence from pocket shark’s pocket epithelium. A, apical cells; B, basal cells; Ba, basal lamina; Bs, blood sinus; Ca, cartilage rod from pectoral fin. C1, cellular type 1; C2, cellular type 2; E, epidermis; G, granular area; I, inclusion; ILS, iris-like structure; L, lens cell; N, nucleus; P, photocyte; Ps, secretory photocyte; R, reticulated layer (reflector); S, pigmented sheath; V, photocyte vesicles; Va, vesicular area. Indicative scale bars in (a–e,g,i), 50 μm; in (f,h), 10 μm.
Figure 4.
Shark photogenic organs. (a) Photophores (modified from [32,34]); (b) secretory photogenic structures: light-producing columnar epithelium from pouch of the taillight shark (modified from [34]; left) and putative light-producing cubic tissue from pocket shark’s pectoral pockets (modified from [74]; right); (c) histological section of I. brasiliensis photophore; (d) histological section of E. lucifer photophore; (e) ultrastructure of E. spinax photophore; (f) ultrastructure of E. spinax photocyte; (g) autofluorescence from E. lucifer photophore; (h) autofluorescence from photocyte vesicle from E. lucifer; (i) autofluorescence from pocket shark’s pocket epithelium. A, apical cells; B, basal cells; Ba, basal lamina; Bs, blood sinus; Ca, cartilage rod from pectoral fin. C1, cellular type 1; C2, cellular type 2; E, epidermis; G, granular area; I, inclusion; ILS, iris-like structure; L, lens cell; N, nucleus; P, photocyte; Ps, secretory photocyte; R, reticulated layer (reflector); S, pigmented sheath; V, photocyte vesicles; Va, vesicular area. Indicative scale bars in (a–e,g,i), 50 μm; in (f,h), 10 μm.
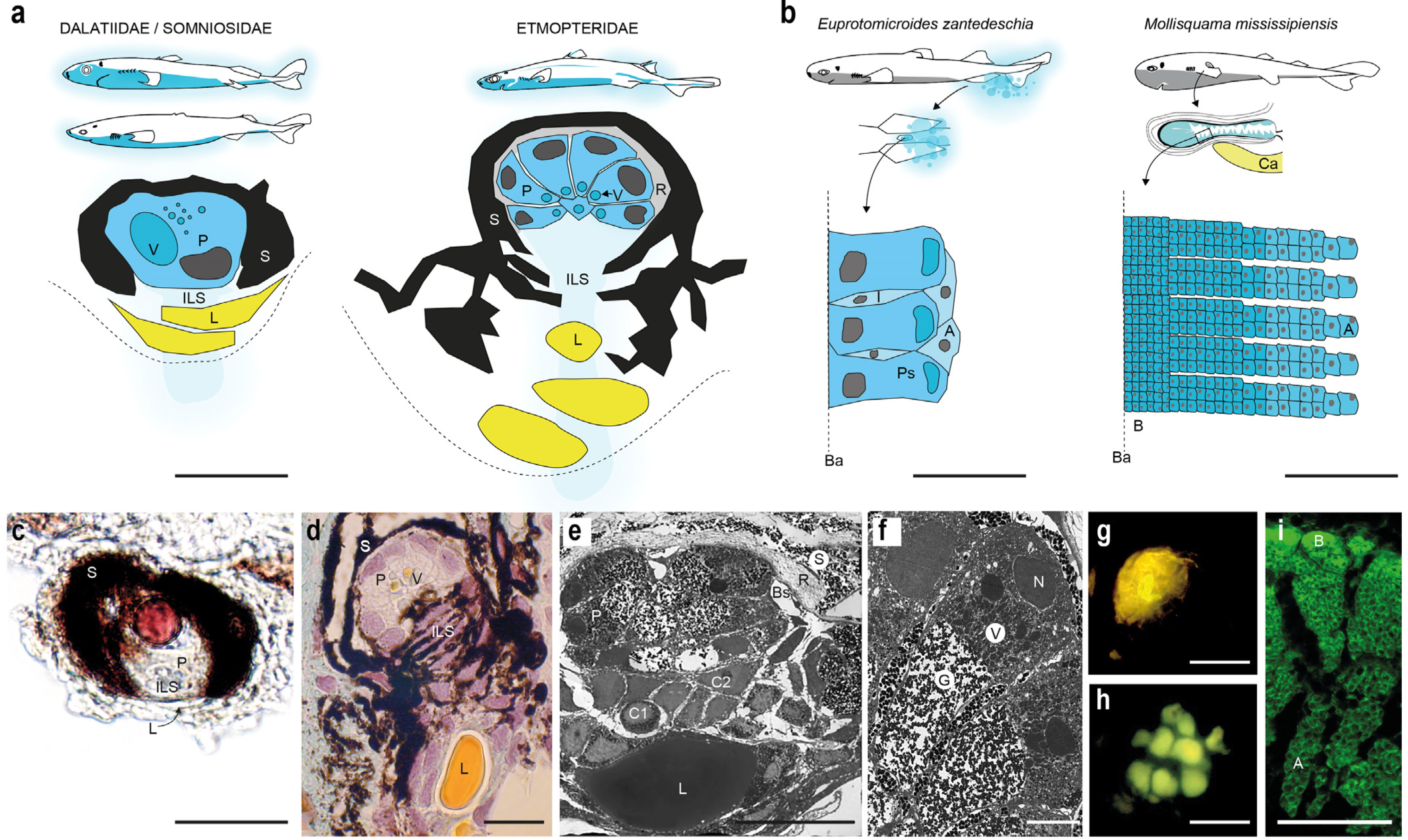
Etmopteridae photophores are composed of 6–14 emitting cells (photocytes) embedded in a cup-shaped pigmented cell sheath, covered by a reflective layer containing guanine crystals and capped by one or several lens cells [16,41,44,76,120]. A multilayer cell zone, called iris-like structure (ILS), is present between the lens cells and the photocytes and is used as a photophore shutter [43,76,77,120,121,122]. Renwart et al., 2014, described three different cell types constituting the ILS of E. spinax photophores: (i) the cellular type I, which contains fibrous material and was assumed to stabilize the lens cells; (ii) the cellular type II, with nucleus only visible via electron microscopy and whose function is unclear, and (iii) the pigmented cells, affiliated to melanophores, presenting pseudopodia-like cellular projections [76] (Figure 4). Photocytes of E. spinax were ultrastructurally depicted as containing three distinct areas: the nucleus, the vesicular and the granular areas (Figure 4e,f). The granular area is assumed to be the location of the bioluminescence chemical reaction and associated microsources were named “glowons” [76,120]. Green autofluorescence is observed within photocytes (especially from their granular area) under blue/UV light exposure (Figure 4g,h), which is assumed to be due to the fluorescence properties of the bioluminescent substrate (as it is the case, e.g., for luciferin; [45,47,76,77]). Shark photophores appear to lack intra-organ innervation [46,76], with terminal epidermal nerves reaching only the surrounding of the photophore as demonstrated through acetylated-tubulin labelling [123].
Comparatively, Dalatiidae and Somniosidae harbor less complex and smaller light organs containing a single photocyte, a pigmented sheath and a group of small lens cells [33,34,36,41,52] (Figure 4a). Pigmented cells, attributed to ILS cells, surmounting the photocytes also act as a photophore shutter [41,77]. Photophore development has been studied in E. spinax and S. aliae, revealing a similar four-phase morphogenesis pattern: (i) apparition of pigmented cells; (ii) formation of the pigmented sheath; (iii) apparition of the protophotocyte (i.e., photocytes not able to produce light); (iv) maturation of the photocyte during which the photocyte acquires its luminescence competence (revealed by the presence of fluorescent vesicles in photocytes; [45,77], Duchatelet, personal observation).
The secretory epithelium from the pelvic pouch of E. zantedeschia displays a pseudostratified columnar epithelium with three distinct cell types: columnar cells with a large apical inclusion (which are probably secretory cells), flattened cells present between columnar cells and extending from the basal lamina towards the free surface of the epithelium, and superficial cells forming a thin cover above the apical part of columnar cells [61]. The epithelium of the putatively bioluminescent fluid-producing pectoral pockets of Mollisquama mississippiensis shows a surprisingly different structure, since it appears to be stratified, made of over 50 layers of cuboidal cells displaying green autofluorescence and probably involved in the production of a holocrine secretion [74] (Figure 4b,i).
Given that shark photophores are embedded in the shark epidermis, they compete for space with dermal denticles (placoid scales). As a consequence, bioluminescent sharks evolved specific squamation patterns, i.e., pavement-like, cross-, bristle-, hook-, and simple/alveolar leaf-shaped patterns [32,34,35,36,41,124,125]; Figure 5, which allows the accommodation of photophores between (pavement-like, cross-, bristle-, hook-shaped types) or below (leaf-shaped types) the placoid scales. Histology and light transmission analyses of leaf-shaped type denticles from Z. squamulosus recently revealed specific honeycomb structures allowing the transmission of at least 50% of the light produced [36]. On the specific rostral area, the kitefin shark, D. licha, also presents highly translucent leaf-shaped placoid scales but without honeycomb structure [41].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5. Control of Bioluminescence from Shark Photophores
5.1. Hormonal Control
Luminous sharks occupy a peculiar place among bioluminescent organisms regarding the control of their luminescence. Indeed, their photogenic organs are primarily controlled by hormones rather than by nerves, the general condition for intrinsic photogenic organs [42,43,47,48,108]. Although the majority of bony fishes have a nervous control of the light emitted (e.g., via adrenaline and noradrenaline, [126,127,128,129]), such classical neurotransmitters have been demonstrated to be inefficient to regulate the shark light emission [16,108]. Intraperitoneal injection of drugs such as adrenalin or acetylcholine in I. brasiliensis failed to trigger light emission [16]. Claes and Mallefet (2009) also attempted, without success, to induce light response in the etmopterid E. spinax after the application of neural agents (e.g., adrenalin, noradrenalin, serotonin, carbachol, and the classical depolarizing agent KCl) on isolated ventral photogenic skin patches, a technique which became the standard for pharmacological studies of organism luminescence control [108]. The absence of photophore innervations, and the inefficiency of classical neurotransmitters to trigger light emission, led to assumptions of a hormonal control of luminescence in sharks [28,108].
Using E. spinax as a model species, and hypothesizing that the shark light emission involves pigment motion within the ILS-melanophores, Claes and Mallefet (2009) have highlighted the implication of melatonin (MT), prolactin (PRL) and alpha melanocyte-stimulating hormone (α-MSH) in the light emission process [108] (Figure 6a). These three hormones are known to be involved in the physiological skin color modulation in elasmobranchs (i.e., sharks and rays): PRL, and α-MSH stimulate melanophore pigment dispersion and, thus, induce skin darkening, and MT regulate melanophore pigment aggregation, leading to a paler skin [130,131]. A few years later, the hormonal control was described in other species such as E. splendidus [47], E. molleri [42], E. lucifer [41] and E. granulosus [41], as well as in two Dalatiidae species, S. aliae [48] and D. licha [41]. Recently, the adrenocorticotropic hormone (ACTH), a melanocortin hormone also involved in vertebrate skin pigment motion within melanophores, was found to inhibit light emission induced by MT in both Etmopteridae and Dalatiidae [41,43] (Figure 6a). Conversely, the melanin-concentrating hormone (MCH), another hormone responsible of the pigment granule aggregation in bony fishes [132,133,134,135] had no effect on shark luminescence [136]. Although the pharmacological control of Z. squamulosus photophores has not been investigated yet, these organs can be assumed to be under hormonal control as well, given their structural similarity with dalatiid photophores [35,36]. Future work on this subject will allow us to confirm this hypothesis.
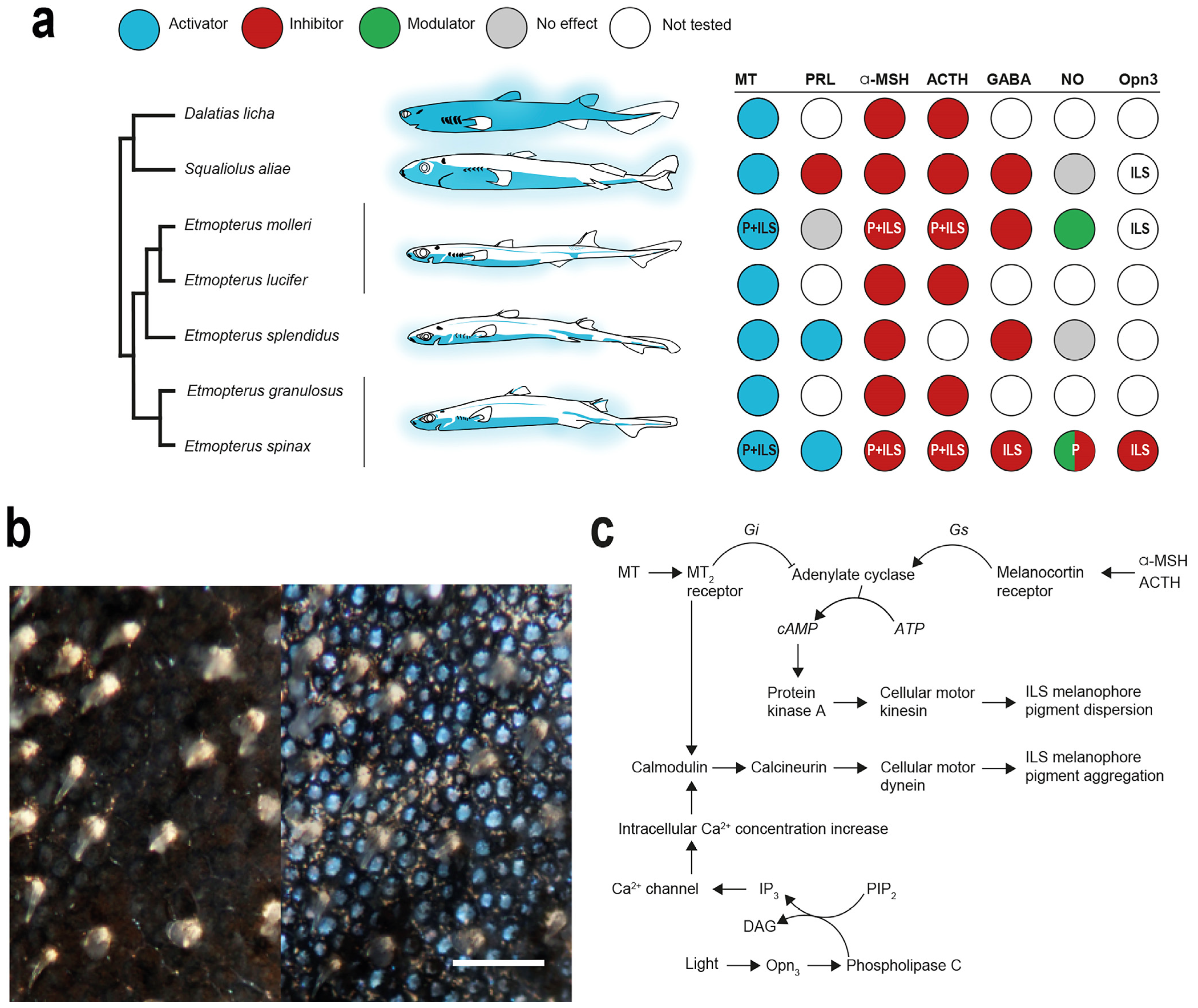
Figure 6.
Shark photophore control. (a) Overview of shark luminescence control agents mapped against experimentally investigated species over the last 15 years [25,27,41,42,43,47,48,77,107,108,121,122,137,138,139]. Letters in circles indicate the identified targets of the agent (ILS, iris-like structure; P, photocytes); (b) photophore-containing skin patch before hormonal stimulation (left) and at luminescence peak (right) from E. spinax [121]; (c) primary hormonal control pathway of shark photophore regulating ILS shutter function (modified from [122]).
Figure 6.
Shark photophore control. (a) Overview of shark luminescence control agents mapped against experimentally investigated species over the last 15 years [25,27,41,42,43,47,48,77,107,108,121,122,137,138,139]. Letters in circles indicate the identified targets of the agent (ILS, iris-like structure; P, photocytes); (b) photophore-containing skin patch before hormonal stimulation (left) and at luminescence peak (right) from E. spinax [121]; (c) primary hormonal control pathway of shark photophore regulating ILS shutter function (modified from [122]).

Overall, the photophore responses to hormone application appear similar across investigated species, except for PRL (Figure 6a). In Etmopteridae, MT and PRL, both at a concentration of 10−6 mol L−1, trigger light emission, while α-MSH at 10−6 mol L−1 and ACTH at 10−5 mol L−1 inhibit it [42,43,47,108] (Figure 6a). Both stimulating hormones present different light-emission curves when applied on photogenic ventral skin patches. Although MT triggers a slow increasing and long-lasting luminescence for up to one hour depending on the species, PRL application induces a fast–high response, decreasing rapidly after a few minutes. Nevertheless, it has been shown that both hormones act on the ILS cells pigment movement to open the “iris” and allow the outward light emission [43,121] (Figure 6b). Conversely, α-MSH and ACTH inhibit the light emission with a decrease in the amount of light produced after both MT or PRL applications, and also act on the ILS cells pigment movement by occulting the photocyte, and hence preventing the light from being emitted outside [41,43,108,121,122]. In Dalatiidae, the hormonal control is similar, except for PRL, which was demonstrated to inhibit the light emission process in S. aliae [41,48]. The physiological effects of each hormone for each studied shark species are summarized in Figure 6a.
In silico transcriptome analyses and immune localization allowed us to confirm the presence and location of MT and α-MSH/ACTH receptors within the photophore from two Etmopteridae species, E. spinax and E. molleri [137] (Figure 6a). These G protein-coupled receptors (GPCR), known to be coupled with specific G proteins, are mainly localized within the cell membrane of photocytes and ILS cells [137]. Researchers also highlighted that PRL receptor mRNA sequences are absent from E. spinax ventral skin reference transcriptome [137], in agreement with previous works revealing the secondary loss of PRL receptor within the elasmobranch lineages during the growth hormone/prolactin protein family evolution [140]. Therefore, how PRL could cellularly have been perceived by the photophore to trigger luminescence remains enigmatic and assumptions have been made on the involvement of the closely related growth hormone and its specific receptor in the light emission control.
An integrative model of the hormonal control of shark photophore luminescence, highlighting the role of MT, α-MSH and ACTH on both the photocyte and the ILS, has been recently proposed [122] (Figure 6c). MT, through its perception by MT receptor, triggers the release of Gi proteins, inhibiting the adenylate cyclase activity [43,122]. Adenylate cyclase is known to directly act on cellular cAMP concentration [141,142,143]. Confirming results from Claes and Mallefet, 2009 [108], cAMP concentration assay after MT-induced light emission revealed a drastic decrease in cAMP levels [43,122]. In fish melanophore, a decrease in the cell cAMP concentration triggers the aggregation of melanin pigment toward the nucleus periphery [142,144,145]. Moreover, MT application was demonstrated to activate, through a calmodulin/calcineurin pathway, the cellular motor dynein, which carries pigmented granules towards melanophore-like nucleus periphery [122]. This MT pathway regulates the “opening” of the ILS cells and allows the emitted light to reach the outside of the photophore [122] (Figure 6). The ultrastructure analyses of the ILS structure before and after the MT-induced luminescence confirmed the pigment motion within the ILS-associated melanophores [120]. On the other hand, analysis of the α-MSH/ACTH pathway revealed how these hormones “close” the photophore and, hence, inhibit light emission. Through melanocortin hormone (α-MSH/ACTH) perception, its receptors release a Gs protein, activating the adenylate cyclase, and an increase in the intra-ILS cells cAMP concentrations [43,122]. The final step of this inhibiting pathway involves the cellular motor kinesin, which carries pigment granules on cytoskeleton towards the ILS melanophore pseudopodial-like projection, occulting the light produced [122] (Figure 6c). These two ILS-regulating pathways seem to be conserved across Etmopteridae and Dalatiidae [41,43,47,122] (Figure 6a).
In addition to the hormonal control, pharmacological studies also revealed that γ-aminobutyric acid (GABA) and nitric oxide (NO) can modulate the light emission in certain species [42,48,123,139] (Figure 6a). In the model species, E. spinax, GABA inhibits and NO modulates the MT/PRL-induced luminescence [123,139]. Conversely, NO application on S. aliae photogenic skin patches, after MT/PRL light emission triggering, had no effects [48] (Figure 6a).
Further research is needed to determine the intracellular interplay between the neuromodulators (NO and GABA) and the hormones as well as to better understand the intracellular events occurring in the photocytes upon hormonal stimulation.
5.2. Extraocular Photoreception and Pigment Motion Regulation
Extraocular photoreception, the ability to detect and respond to light clues outside of the “eyes”, has been suggested to be involved in the bioluminescence control in various invertebrate taxa. In the bobtail squid, Euprymna scolopes, extraocular photoreceptors and photocytes are colocalized within photophores. Expression of eye-like developing genes in the light “pocket” of E. scolopes underlines such a coupling between photoemission and photoreception mechanisms [146,147]. In the comb jelly, Memniopsis leidyi, and the burrowing ophiuroid, Amphiura filiformis, both photoreceptor and photocyte molecular markers (i.e., opsin and photoprotein/luciferase, respectively) are coexpressed within the same cells. These observations support the existence of a functional link between light perception and bioluminescence control for these species [8,148,149,150]. By analyzing the opsin and phototransduction genes expression within photophores, light organ photosensitivity was also suggested for a counterilluminating deep-sea shrimp, Janicella spinicauda, where it was assumed to play a role in the light emission regulation to ensure a match with the residual downwelling light [151].
As indicated by recent analyses of ventral skin and eye transcriptomes of E. spinax, photoreceptors and phototransduction actors were expressed in both tissues [152]. This species displays only two ocular opsins, one rhodopsin and one putatively associated peropsin, highlighting its monochromatic vision [152]. A specific extraocular photopigment, the encephalopsin (Opn3), was also detected in the skin transcriptome of E. spinax [152]. The colocation of this extraocular opsin (Es-Opn3) with the ventral skin photophores provides fuel for the putative functional coupling between light emission and light perception in luminous organisms [152]. More precisely, membranes of the ILS cells were shown to be the main expression site of the Es-Opn3, while no expression was found at the photocyte level [152]. Besides, the expression of this opsin was demonstrated to be concomitant with the in utero development of the photophores in E. spinax and S. aliae embryos, appearing with the transformation of protophotocytes to photocytes (i.e., when photocytes can produce luminescence; [77], Duchatelet, unpublished data), which supports the idea that the Es-Opn3 is used to detect embryonic luminescence [77]. The evaluation of the absorbance spectrum of this luminous shark extraocular opsin has added new evidence of the link between the two photobiological processes, since a clear overlap exists between the light emission spectrum of E. spinax and the Es-Opn3 photopigment light absorbance [122].
Going further in the phototransduction cascade, Duchatelet et al., 2020, demonstrated an impact of blue light exposure on the intracellular concentration of inositol triphosphate (IP3) from photogenic ventral skin [122]. This IP3 concentration modulation confirmed opsin activity and highlighted the first step of the phototransduction cascade occurring at the photophore level. Pharmacological analyses unveiled the next steps of the phototransduction pathway, with the implication of calcium, the Ca2+-dependent calmodulin, and the cytoplasmic motor dynein [122], clearly demonstrating the interconnection between these pathway steps and the pathway leading to the pigment granule aggregation in melanophores. Therefore, Duchatelet et al., 2020, proposed a model of light emission control in E. spinax based on the photoperception of its own luminescence that regulates ILS melanophore pigment aggregation and, thus, the aperture of the photophore, which regulates the light output [122] (Figure 6c). Interestingly, while hormones operate at both the photocyte, triggering light emission through an unknown biochemical reaction (i.e., unknown luciferin/luciferase system or photoprotein) and ILS (to regulate the amount of light transmitted as a camera diaphragm) levels, Es-Opn3 appears to be involved only in the pigment motion regulation of the ILS. This dual control is assumed to occur at least in the luminous species belonging to Dalatiidae and Etmopteridae (Duchatelet, unpublished data). The close link between photoemission and photoreception at the light-organ level, putatively regulating the amount of light emitted, seems to have emerged independently in phylogenetically distant luminous organisms: ctenophores, cephalopods, crustaceans, echinoderms, and luminous sharks [8,122,146,147,148,149,150,151,152]. It is a fascinating example of convergent evolution.
6. Biochemistry of Shark Luminescence
One of the remaining questions is what type of bioluminescent system is involved in the emission of light in sharks. Two categories of light systems are known to date: (i) luciferase–luciferin systems, and (ii) photoprotein systems. In the former, a substrate, the luciferin, is oxidized by an enzyme, the luciferase, in the presence of oxygen and often other cofactors [5]. In the latter, an enzyme complex as well as a preoxidized luciferin form together a complex protein, the so-called photoprotein, requiring only the contribution of a cofactor (often an ion) to produce light [5,153]. Various luciferin types have been molecularly described in the marine environments to date, such as the coelenterazine (the most phylogenetically widespread luciferin type in the oceanic environment) and its derivatives (dehydrocoelenterazine and enol-sulfate coelenterazine), the dinoflagellate luciferin, or the Cypridina luciferin (also called vargulin) [1,13,154]. Associated with these luciferins, different types of luciferases (nine different types) and photoprotein (three different types) have been molecularly depicted [1,5,9,13]. These substrates and enzymes, although initially considered to be species-specific [28], can sometimes be shared by phylogenetically very distant species [5,6,7,8,9]. This has led scientists to hypothesize that certain species can acquire the necessary components for the luminous reaction through their diet [1,5,155,156,157,158,159]. Recently, the golden sweeper fish, Parapriacanthus ransonnetti, has been shown to acquire not only its luciferin, but also its luciferase by feeding on luminous ostracods [160]. It is noted that the only species described as being able to synthesize de novo their luciferins are the midwater shrimp Systellaspis debilis [161], the ostracod Metridia pacifica [162], and the ctenophores Bolinopsis infundibulum and M. leidyi [163].
Attempts were made to identify the bioluminescent compound responsible for the light emission in E. spinax by analyzing the cross-reactivity of known luciferins with extract of shark skin assumed to contain the catalyst (i.e., luciferase). In parallel, the putative diet acquisition of a luciferin through the food chain was hypothesized. Only coelenterazine, the most commonly found luciferin in marine taxa [5,164,165,166,167], was found in the digestive tract of E. spinax, but none of the tested luciferins (coelenterazine, dinoflagellate/euphausiid luciferin, and Cypridina luciferin) has reacted with the shark photogenic skin extract [168]. Similar negative results have been obtained for the cross-reactivity with the respective luciferase: coelenterazine-dependent luciferase (Renilla luciferase), cypridinid luciferase, or euphausid luciferase [168]. Analysis of the transcriptomic sequence available for the photogenic skin failed to pinpoint any putative homologs of known luciferase/photoprotein sequences [52,152], leading to assume the involvement of an unknown light-emitting system in luminous sharks, including either an unknown light-emitting system (photoprotein) or a specific active or storage form of a known luciferin using a shark-specific luciferase. Research combining data from transcriptomics, proteomics, and bioinformatics modeling are ongoing, and could allow us to decipher this enigmatic bioluminescent system.
7. Conclusions and Perspectives
Bioluminescent sharks have fascinated humans for almost two centuries. Yet, dedicated research on these enigmatic deep-sea inhabitants involving spectrophotometry, luminometry, pharmacology, light/electron microscopy, biochemistry, molecular analyses, and transcriptomics started only 15 years ago. Results from those studies (over 50 publications in total) have been synthesized in the current review. Overall, findings suggest that luminescence acquisition has been a unique though pivotal evolutionary event for squaliform sharks, which greatly facilitated their radiation in deep-sea habitats and strongly shaped their visual system. From a primary function of camouflage (coopted from the hormonally controlled crypsis mechanism of shallow water elasmobranchs) in Dalatiidae, Somniosidae and basal Etmopteridae, shark bioluminescent patterns progressively became an intra- and interspecific communication tool in derived etmopterid sharks (e.g., Etmopterus species), an exaptation that considerably increased their speciation rate and has probably been facilitated by the increase in size and complexity of etmopterid photophores (which allows better orientation of light output, e.g., tangential to the body surface). The secretory luminescence observed in E. zantedeschia (and putatively present in pocket sharks, Mollisquama sp.), which might have evolved from the invagination and modification of epidermal photophores, represents an example of the high selective pressure occurring in the darkness of the deep sea.
It clearly appears that the future of shark bioluminescence research will also be driven by new molecular data and techniques. The Next-Generation Sequencing methods recently allowed the emergence of transcriptomic studies in non-model organisms such as some selected luminous shark species (i.e., E. spinax, I. brasiliensis [52,152]). These studies paved the way for future transcriptomic, proteomic, and genomic studies on luminous sharks. Among an infinite number of fascinating questions, these studies could focus on the identification of the light-emitting molecular toolkit (luciferase, photoprotein, etc.) in luminous sharks.
Up to now, genomic resources for cartilaginous fish are still very limited (i.e., genomes available for the great white shark, Carcharodon carcharias, the whale shark, Rhincodon typus, the elephant shark, Callorhinchus milii, and the little skate, Leucoraja erinacea) and absent for luminous sharks. Genome size estimation in different Etmopterid species revealed that these species have among the largest genomes of all investigated Chondrichthyes (e.g., genome size of E. spinax might reach 16 Gbp, against 4.63 Gbp in C. cacharias, and 3 Gbp in R. typus) [169,170,171,172,173]. These impressive genome sizes possibly reveal lineage-specific genome expansion and large-scale alteration events such as gene/genome duplications, transposon insertions or events such as polyploidy [174], and confirm the challenging future of genomic studies on these species.
Further research on these sharks is planned to understand their ecology. In vivo ethological studies could lead to a better understanding of their behavior, notably the “isolumes follower” hypothesis by passive acoustic tagging of these sharks. Another approach could be to follow “in situ” reaction to stimuli mimicking the light emission of conspecifics.
Although recent research allowed to draw a clearer picture on the evolution, ecology and physiology of shark luminescence, gaps in our knowledge of these fascinating animals still exist. In particular, the interplay between hormonal control pathway and neuromodulation as well as the chemistry of shark luminescence remain to be determined. Work is underway to clarify these shadow areas.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/oceans2040047/s1, Table S1: Overview table of the luminous species within the luminous Squaliformes (Etmopteridae, Dalatiidae, Somniosidae) according to the literature.
Author Contributions
Conceptualization, L.D., J.M.C. and J.M.; software, L.D.; investigation, L.D., J.M.C., J.D. and J.M.; writing—original draft preparation, L.D., J.M.C. and J.D.; writing—review and editing, L.D., J.M.C., J.D., P.F. and J.M.; supervision, J.M.; project administration, L.D.; funding acquisition, P.F. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fonds National de la Recherche Scientifique (F.R.S.-FNRS, Belgium), grant number T.0169.20 awarded to the University of Louvain—UCLouvain Marine Biology laboratory and the University of Mons—UMONS Biology of Marine Organisms and Biomimetics laboratory. L.D., J.M.C., J.D., P.F. and J.M. are, respectively a postdoctoral researcher, scientific collaborator of the UCLouvain laboratory, postdoctoral researcher F.R.S.-FNRS, research director F.R.S.-FNRS, and research associate F.R.S.-FNRS.
Acknowledgments
This review is the contribution BRC # 381 of the Biodiversity Research Center (UCLouvain) from the Earth and Life Institute Biodiversity (ELIB) and the “Centre Interuniversitaire de Biologie Marine” (CIBIM).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Haddock, S.H.D.; Moline, M.A.; Case, J.F. Bioluminescence in the sea. Annu. Rev. Mar. Sci. 2010, 2, 443–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastings, J.W.; Morin, J.G. Bioluminescence. In Comparative Animal Physiology. Neural and Integrative Animal Physiology; Prosser, C.L., Ed.; John Wiley and Sons: Chichester, NY, USA, 1991; Chapter 3; pp. 131–170. [Google Scholar]
- Hastings, J.W. Biological diversity, chemical mechanisms, and the evolutionary origins of bioluminescent systems. J. Mol. Evol. 1983, 19, 309–321. [Google Scholar] [CrossRef]
- Wilson, T.; Hastings, J.W. Bioluminescence. Ann. Rev. Cell. Dev. Biol. 1998, 14, 197–230. [Google Scholar] [CrossRef]
- Shimomura, O. Bioluminescence: Chemical Principles and Methods; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2012. [Google Scholar]
- Widder, E.A. Bioluminescence in the ocean: Origins of biological, chemical, and ecological diversity. Science 2010, 328, 704–708. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, G.; Metcalf, P.; Paterson, N.G.; Sharpe, M.L. Mass spectrometry analysis and transcriptome sequencing reveal glowing squid crystal proteins are in the same superfamily as firefly luciferase. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delroisse, J.; Ullrich-Lüter, E.; Blaue, S.; Ortega-Martinez, O.; Eeckhaut, I.; Flammang, P.; Mallefet, J. A puzzling homology: A brittle star using a putative cnidarian-type luciferase for bioluminescence. Open Biol. 2017, 7, 160300. [Google Scholar] [CrossRef] [Green Version]
- Delroisse, J.; Duchatelet, L.; Flammang, P.; Mallefet, J. Leaving the dark side? Insights into evolution of luciferases. Front. Mar. Sci. 2021, 8, 673620. [Google Scholar] [CrossRef]
- Hastings, J.W. Bioluminescence. In Cell Physiology Source Book; Sperelakis, N., Ed.; Academic Press: Cambridge, MA, USA, 1995; pp. 665–681. [Google Scholar] [CrossRef]
- Oba, Y.; Schultz, D.T. Eco-Evo bioluminescence on land and in the sea. In Bioluminescence: Fundamentals and Applications in Biotechnology—Volume 1. Advances in Biochemical Engineering/Biotechnology; Thouand, G., Marks, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 144, pp. 3–36. [Google Scholar] [CrossRef]
- Davis, M.P.; Sparks, J.S.; Smith, W.L. Repeated and widespread evolution of bioluminescence in marine fishes. PLoS ONE 2016, 11, e0155154. [Google Scholar] [CrossRef]
- Lau, E.S.; Oakley, T.H. Multi-level convergence of complex traits and the evolution of bioluminescence. Biol. Rev. 2020, 96, 673–691. [Google Scholar] [CrossRef]
- Morin, J.G. Coastal bioluminescence: Patterns and functions. Bull. Mar. Sci. 1983, 33, 787–817. [Google Scholar]
- Martini, S.; Kuhnz, L.; Mallefet, J.; Haddock, S.H.D. Distribution and quantification of bioluminescence as an ecological trait in the deep sea benthos. Sci. Rep. 2019, 9, 14654. [Google Scholar] [CrossRef] [Green Version]
- Herring, P.J.; Morin, J.G. Bioluminescence in fishes. In Bioluminescence in Action; Herring, P.J., Ed.; New York Academic Press: New York, NY, USA, 1978; pp. 273–329. [Google Scholar]
- Haneda, Y. Further studies on a luminous land snail, Quantula striata, in Malaya. Sci. Rep. Yokosuka City Mus. 1963, 8, 1–9. [Google Scholar]
- Rudie, N.G.; Mulkerrin, M.G.; Wampler, J.E. Earthworm bioluminescence: Characterization of high specific activity Diplocardia longa luciferase and the reaction it catalyzes. Biochemistry 1981, 20, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.V. The chemical mechanism and evolutionary development of beetle bioluminescence. Photochem. Photobiol. 1995, 62, 662–673. [Google Scholar] [CrossRef]
- Broadley, R.A.; Stringer, I.A.N. Prey attraction by larvae of the New Zealand glowworm, Arachnocampa luminosa (Diptera: Mycetophilidae). Invertebr. Biol. 2001, 120, 170–177. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Moore, S. Hitherto unreported aspects of the ecology and anatomy of a unique gastropod: The bioluminescent freshwater pulmonated Latia neritoides. In Bioluminescence in Focus—A Collection of Illuminating Essays; Meyer-Rochow, V.B., Ed.; Research Signpost: Kerala, India, 2009; pp. 85–104. [Google Scholar]
- Oba, Y.; Branham, M.A.; Fukatsu, T. The terrestrial bioluminescent animals of Japan. Zool. Sci. 2011, 28, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.J.; Branham, M.A.; Whiting, M.F.; Bybee, S.M. Total evidence phylogeny and the evolution of adult bioluminescence in fireflies (Coleoptera: Lampyridae). Mol. Phylogenetics Evol. 2017, 107, 564–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, F.D. Narrative of a Whaling Voyage Round the Globe, from the Year 1833 to 1836; Рипoл Классик: Moscow, Russia, 1840; Volume 2. [Google Scholar]
- Claes, J.M. Function and Control of Luminescence from Lantern Shark (Etmopterus spinax) Photophores. Ph.D. Thesis, Université catholique de Louvain—UCLouvain, Ottignies-Louvain-la-Neuve, Belgium, 4 May 2010. [Google Scholar]
- Renwart, M. Ultrastructure and Biochemistry of the Light-Emitting System of Lantern Shark (Etmopterus spinax) Photophores. Ph.D. Thesis, Université catholique de Louvain—UCLouvain, Ottignies-Louvain-la-Neuve, Belgium, 18 December 2014. [Google Scholar]
- Duchatelet, L. Extraocular Photoreception and Bioluminescence of Representatives of Two Luminous Shark Families, Etmopteridae and Dalatiidae. Ph.D. Thesis, Université Catholique de Louvain—UCLouvain, Ottignies-Louvain-la-Neuve, Belgium, 3 October 2019. [Google Scholar]
- Harvey, E.N. Bioluminescence. Chapter XVI Pisces; Academic Press Inc.: New York, NY, USA, 1952; pp. 494–553. [Google Scholar]
- Alcock, A. A Naturalist in Indian Seas: Or, Four Years with the Royal Indian Marine Survey Ship “Investigator”; Dutton: New York, NY, USA, 1902. [Google Scholar]
- de Carvalho, M.R. A synopsis of the deep-sea genus Benthobatis Alcock, with a redescription of the type species Benthobatis moresbyi Alcock, 1898 (Chondrichthyes, Torpediniformes, Narcinidae). In Proceedings of the 5th Indo-Pacific Fish Conference, Nouméa, New Caledonia, 3–8 November 1997; Séret, B., Sire, J.-Y., Eds.; Societe Francaise d’ Ichtyologie: Paris, France, 1999; pp. 231–255. [Google Scholar]
- de Carvalho, M.R.; Compagno, L.J.V.; Ebert, D.A. Benthobatis yangi, a new species of blind electric ray from Taiwan (Chondrichthyes: Torpediniformes: Narcinidae). Bull. Mar. Sci. 2003, 72, 923–939. [Google Scholar]
- Duchatelet, L.; Moris, V.; Tomita, T.; Mahillon, J.; Sato, K.; Behets, C.; Mallefet, J. The megamouth shark, Megachasma pelagios, is not a luminous species. PLoS ONE 2020, 15, e0242196. [Google Scholar] [CrossRef]
- Hubbs, C.L.; Iwai, T.; Matsubara, K. External and internal characters, horizontal and vertical distribution, luminescence, and food of the dwarf pelagic shark, Euprotomicrus bispinatus. Bull. Scripps Inst. Oceanogr. 1967, 10, 1–64. [Google Scholar]
- Claes, J.M.; Mallefet, J. Bioluminescence of sharks: First synthesis. In Bioluminescence in Focus—A Collection of Illuminating Essays; Meyer-Rochow, V.B., Ed.; Research Signpost: Kerala, India, 2009; pp. 51–65. [Google Scholar]
- Straube, N.; Li, C.; Claes, J.M.; Corrigan, S.; Naylor, G.J.P. Molecular phylogeny of Squaliformes and first occurrence of bioluminescence in sharks. BMC Evol. Biol. 2015, 15, 162. [Google Scholar] [CrossRef] [Green Version]
- Duchatelet, L.; Marion, R.; Mallefet, J. A third luminous shark family: Confirmation of luminescence ability for Zameus squamulosus (Squaliformes; Somniosidae). Photochem. Photobiol. 2021, 97, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Adnet, S.; Capetta, H. A paleontological and phylogenetical analysis of Squaliform sharks (Chondrichthyes: Squaliformes) based on dental characters. Lethaia 2001, 34, 234–248. [Google Scholar] [CrossRef]
- Straube, N.; Iglésias, S.P.; Sellos, D.Y.; Kriwet, J.; Schliewen, U.K. Molecular phylogeny and node time estimation of bioluminescent lantern sharks (Elasmobranchii: Etmopteridae). Mol. Phylogenet. Evol. 2010, 56, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V.; Dando, M.; Fowler, S. Sharks of the World; Harper Collins: London, UK, 2004. [Google Scholar]
- Castro, J.I. The Sharks of North America; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Mallefet, J.; Stevens, D.W.; Duchatelet, L. Bioluminescence of the largest luminous vertebrate, the kitefin shark, Dalatias licha: First insights and comparative aspects. Front. Mar. Sci. 2021, 8, 633582. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Comparative control of luminescence in sharks: New insights from the slendertail lanternshark (Etmopterus molleri). J. Exp. Mar. Biol. Ecol. 2015, 467, 87–94. [Google Scholar] [CrossRef]
- Duchatelet, L.; Delroisse, J.; Pinte, N.; Sato, K.; Ho, H.-C.; Mallefet, J. Adrenocorticotropic hormone and cyclic adenosine monophosphate are involved in the control of shark bioluminescence. Photochem. Photobiol. 2020, 96, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H. Some observations on the luminous organs of fishes. J. Coll. Sci. Imp. Univ. Tokyo 1911, 27, 1–25. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Early development of bioluminescence suggests camouflage by counter-illumination in the velvet belly lantern shark Etmopterus spinax (Squaloidea: Etmopteridae). J. Fish Biol. 2008, 73, 1337–1350. [Google Scholar] [CrossRef]
- Johann, L.; der Organe, I.V. Über eigentümliche epitheliale Gebilde (Leuchtorgane) bei Spinax niger Aus dem zoologischen Institut der Universität Rostock. Von. Z. Wiss. Zool. 1899, 66, 136. [Google Scholar]
- Claes, J.M.; Sato, K.; Mallefet, J. Morphology and control of photogenic structures in a rare dwarf pelagic lantern shark (Etmopterus splendidus). J. Exp. Mar. Biol. Ecol. 2011, 406, 1–5. [Google Scholar] [CrossRef]
- Claes, J.M.; Ho, H.-C.; Mallefet, J. Control of luminescence from pygmy shark (Squaliolus aliae) photophores. J. Exp. Biol. 2012, 215, 1691–1699. [Google Scholar] [CrossRef] [Green Version]
- Stehmann, M.F.W.; Van Oijen, M.; Kamminga, P. Re-description of the rare taillight shark Euprotomicroides zantedeschia (Squaliformes, Dalatiidae), based on third and fourth record from off Chile. Cybium 2016, 40, 187–197. [Google Scholar]
- Dickens, D.A.G.; Marshall, N.J. Observations on Euprotomicrus. Mar. Obs. 1956, 26, 73–74. [Google Scholar]
- Widder, E.A. A predatory use of counterillumination by the squaloid shark, Isistius brasiliensis. Environ. Biol. Fishes 1998, 53, 267–273. [Google Scholar] [CrossRef]
- Delroisse, J.; Duchatelet, L.; Flammang, P.; Mallefet, J. Photophore distribution and enzymatic diversity within the photogenic integument of the cookie cutter shark Isistius brasiliensis (Chondrichthyes: Dalatiidae). Front. Mar. Sci. 2021, 8, 627045. [Google Scholar] [CrossRef]
- Ebert, D.A.; Straube, N.; Leslie, R.W.; Weigmann, S. Etmopterus alphus n. sp.: A new lanternshark (Squaliformes: Etmopteridae) from the south-western Indian Ocean. Afr. J. Mar. Sci. 2016, 38, 329–340. [Google Scholar] [CrossRef]
- Ebert, D.A.; Leslie, R.W.; Weigmann, S. Etmopterus brosei sp. nov.: A new lanternshark (Squaliformes: Etmopteridae) from the southeastern Atlantic and southwestern Indian oceans, with a revised key to the Etmopterus lucifer clade. Mar. Biodiv. 2021, 51, 53. [Google Scholar] [CrossRef]
- Grace, M.A.; Doosey, M.H.; Denton, J.S.S.; Naylor, G.J.P.; Bart, H.L.; Maisey, J.G. A new Western North Atlantic Ocean kitefin shark (Squaliformes: Dalatiidae) from the Gulf of Mexico. Zootaxa 2019, 4619, 109–120. [Google Scholar] [CrossRef]
- Burckhardt, R. On the luminous organs of selachian fishes. Ann. Mag. Nat. Hist. 1900, 7, 558–568. [Google Scholar] [CrossRef]
- Bigelow, H.B.; Schoeder, W.C.; Springer, S. New and little known sharks from the Atlantic and from the Gulf of Mexico. Bull. Mus. Comp. Zool. Harv. Coll. 1953, 109, 213–276. [Google Scholar]
- Abe, T. Description of a new squaloid shark, Centroscyllium kamoharai, from Japan. Jpn. J. Ichthyol. 1966, 13, 190–198. [Google Scholar] [CrossRef]
- Chan, W.L. New sharks from the South China Sea. J. Zool. 1966, 148, 218–237. [Google Scholar] [CrossRef]
- Dolganov, V.N. A new shark from the family Squalidae caught on the Naska submarine ridge. Zool. Z. 1984, 63, 1589–1591. [Google Scholar]
- Munk, O.; Jørgensen, J.M. Putatively luminous tissue in the abdominal pouch of a male dalatiine shark, Euprotomicroides zantedeschia Hulley & Penrith, 1966. Acta Zool. 1988, 69, 247–251. [Google Scholar] [CrossRef]
- Shirai, S.; Nakaya, K. A new squalid species of the genus Centroscyllium from the Emperor seamont chain. Jpn. J. Ichtyol. 1990, 36, 391–398. [Google Scholar] [CrossRef]
- Last, P.R.; Burgess, G.H.; Séret, B. Description of six new species of lantern-sharks of the genus Etmopterus (Squaloidea: Etmopteridae) from the Australasian region. Cybium 2002, 26, 203–223. [Google Scholar]
- Knuckey, J.D.S.; Ebert, D.A.; Burgess, G.H. Etmopterus joungi n. sp., a new species of lanternshark (Squaliformes: Etmopteridae) from Taiwan. Aqua Int. J. Ichthyol. 2010, 17, 61–72. [Google Scholar]
- Claes, J.M.; Nilsson, D.-E.; Straube, N.; Collin, S.P.; Mallefet, J. Iso-luminance counterillumination drove bioluminescent shark radiation. Sci. Rep. 2014, 4, 4328. [Google Scholar] [CrossRef] [Green Version]
- Grace, M.A.; Doosey, M.H.; Bart, H.L.; Naylor, G.J.P. First record of Mollisquama sp. (Chondrichthyes: Squaliformes: Dalatiidae) from the Gulf of Mexico, with a morphological comparison to the holotype description of Mollisquama parini Dolganov. Zootaxa 2015, 3948, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Vásquez, V.E.; Ebert, D.A.; Long, D.J. Etmopterus benchleyi n. sp., a new lanternshark (Squaliformes: Etmopteridae) from the central eastern Pacific Ocean. J. Ocean. Sci. Found 2015, 17, 43–55. [Google Scholar]
- Ebert, D.A.; Papastamatiou, Y.P.; Kajiura, S.M.; Wetherbee, B.M. Etmopterus lailae sp. nov., a new lanternshark (Squaliformes: Etmopteridae) from the Northwestern Hawaiian Islands. Zootaxa 2017, 4237, 371–382. [Google Scholar] [CrossRef] [Green Version]
- White, W.T.; Ebert, D.A.; Mana, R.R.; Corrigan, S. Etmopterus samadiae n. sp., a new lanternshark (Squaliformes: Etmopteridae) from Papua New Guinea. Zootaxa 2017, 4244, 339–354. [Google Scholar] [CrossRef]
- de Figueiredo Petean, F.; de Carvalho, M.R. Comparative morphology and systematics of the cookiecutter sharks, genus Isistius Gill (1864) (Chondrichthyes: Squaliformes: Dalatiidae). PLoS ONE 2018, 13, e0201913. [Google Scholar] [CrossRef] [PubMed]
- Dolganov, V.N.; Balanov, A.A. Etmopterus parini sp. n. (Squaliformes: Etmopteridae), a new shark species from the northwestern Pacific Ocean. Biol. Morya 2018, 44, 427–430. [Google Scholar]
- Ebert, D.A.; Van Hees, K.E. Etmopterus marshae sp. Nov, a new lanternshark (Squaliformes: Etmopteridae) from the Philippine Islands, with a revised key to the Etmopterus lucifer clade. Zootaxa 2018, 4508, 197–210. [Google Scholar] [CrossRef]
- Dolganov, V.N. On the little-known sharks Etmopterus villosus (Etmopteridae) and Scymnodalatias sherwoodi (Somniosidae) from the Pacific Ocean. J. Ichthyol. 2019, 59, 275–279. [Google Scholar] [CrossRef]
- Claes, J.M.; Delroisse, J.; Grace, M.A.; Doosey, M.H.; Duchatelet, L.; Mallefet, J. Histological evidence for secretory bioluminescence from pectoral pockets of the American pocket shark (Mollisquama mississippiensis). Sci. Rep. 2020, 10, 18762. [Google Scholar] [CrossRef]
- Claes, J.M.; Nilsson, D.-E.; Mallefet, J.; Straube, N. The presence of lateral photophores correlates with increased speciation in deep-sea bioluminescent sharks. R. Soc. Open Sci. 2015, 2, 150219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renwart, M.; Delroisse, J.; Claes, J.M.; Mallefet, J. Ultrastructural organization of lantern shark (Etmopterus spinax Linnaeus, 1758) photophores. Zoomorphology 2014, 133, 405–416. [Google Scholar] [CrossRef]
- Duchatelet, L.; Claes, J.M.; Mallefet, J. Embryonic expression of encephalopsin supports bioluminescence perception in lanternshark photophores. Mar. Biol. 2019, 166, 21. [Google Scholar] [CrossRef]
- Duchatelet, L.; Delroisse, J.; Flammang, P.; Mahillon, J.; Mallefet, J. Etmopterus spinax, the velvet belly lanternshark, does not use bacterial luminescence. Acta Histochem. 2019, 121, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Schofield, P.J.; Burgess, G.H. Etmopterus robinsi (Elasmobranchii, Etmopteridae), a new species of deepwater lantern shark from the Caribbean Sea and Western North Atlantic, with a redescription of Etmopterus hillianus. Bull. Mar. Sci. 1997, 60, 1060–1073. [Google Scholar]
- Straube, N.; Duhamel, G.; Gasco, N.; Kriwet, J.; Schliewen, U.K. Description of a new deep-sea lantern shark Etmopterus viator sp. nov. (Squaliformes: Etmopteridae) from the Southern Hemisphere. In The Kerguelen Plateau, Marine Ecosystem and Fisheries; Société Française d’Ichtyologie: Paris, France, 2011; pp. 135–148. [Google Scholar]
- Fricke, R.; Koch, I. A New Species of the Lantern Shark Genus Etmopterus from Southern Africa (Elasmobranchii: Squalidae)/With 4 Figures and 1 Table; Stuttgarter Beiträge zur Naturkunde: Stuttgart, Germany, 1990. [Google Scholar]
- Iwai, T. Luminous organs of the deep-sea squaloid shark, Centroscyllium ritteri Jordan and Fowler. Pac. Sci. 1960, 14, 51–54. [Google Scholar]
- Schaaf-DaSilva, J.A.; Ebert, D.A. Etmopterus burgessi sp. nov., a new species of lanternshark (Squaliformes: Etmopteridae) from Taiwan. Zootaxa 2006, 1373, 53–64. [Google Scholar] [CrossRef]
- Springer, S.; Burgess, G.H. Two new dwarf dogsharks (Etmopterus, Squalidae), found off the Caribbean coast of Colombia. Copeia 1985, 1985, 584–591. [Google Scholar] [CrossRef]
- Shirai, S.; Tachikawa, H. Taxonomic resolution of the Etmopterus pusillus species group (Elasmobranchii, Etmopteridae), with description of E. bigelowi, n. sp. Copeia 1993, 1993, 483–495. [Google Scholar] [CrossRef]
- Dolganov, V.N. Description of new species of sharks of the family Squalidae (Squaliformes) from the north-western part of the Pacific Ocean with remarks of validity of Etmopterus. Zool. Zhurnal 1986, 65, 149–153. [Google Scholar]
- Gubanov, E.P.; Kondyurin, V.V.; Myagkov, N.A. Sharks of the World Ocean: Identification Handbook. [=Akuly Mirovogo Okeana: Sparvochnik-Opredelitel’]; Agropromizdat: Moscow, Russia, 1986; pp. 1–272. [Google Scholar]
- Mochizuki, K.; Ohe, F. Trigonognathus kabeyai, a new genus and species of the Squalid sharks from Japan. Jpn. J. Ichthyol. 1990, 36, 385–390. [Google Scholar] [CrossRef]
- Seigel, J.A. Revision of the Dalatiid shark genus Squaliolus: Anatomy, systematics, ecology. Copeia 1978, 1978, 602–614. [Google Scholar] [CrossRef]
- Garrick, J.A.F.; Springer, S. Isistius plutodus, a new Squaloid shark from the Gulf of Mexico. Copeia 1964, 1964, 678–682. [Google Scholar] [CrossRef]
- Pollerspöck, J.; Straube, N. Bibliography Database of Living/Fossil Sharks, Rays and Chimaeras (Chondrichtyes: Elasmobranchii, Holocephali)—List of Valid Extant Species; List of Described Extant Species; Statistic, World Wide Web Electronic Publication, Version 10/2021; ISSN: 2195-6499. Available online: www.shark-references.com (accessed on 23 November 2021).
- Denton, J.S.S.; Maisey, J.G.; Grace, M.; Pradel, A.; Doosey, M.H.; Bart, H.L., Jr.; Naylor, G.J.P. Cranial morphology in Mollisquama sp. (Squaliformes; Dalatiidae) and patterns of cranial evolution in dalatiid sharks. J. Anat. 2018, 233, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Daley, R.; Appleyard, S.A.; Koopman, M. Genetic catch verification to support recovery plans for deepsea gulper sharks (genus Centrophorus, family Centrophoridae)- an Australian example using the 16S gene. Mar. Freshw. Res. 2012, 63, 708–714. [Google Scholar] [CrossRef]
- Stefanni, S.; Catarino, D.; Ribeiro, P.A.; Freitas, M.; Menezes, G.M.; Neat, F.; Stankovic, D. Molecular systematics of the long-snouted deep water dogfish (Centrophoridae, Deania) with implications for identification, taxonomy, and conservation. Front. Mar. Sci. 2021, 7, 588192. [Google Scholar] [CrossRef]
- Ziadi-Künzli, F.; Soliman, T.; Imai, H.; Sakurai, M.; Maeda, K.; Tachihara, K. Re-evaluation of deep-sea dogfishes (genus Squalus) in Japan using phylogenetic inference. Deep Sea Res. I 2020, 160, 103261. [Google Scholar] [CrossRef]
- White, W.T.; Vaz, D.F.B.; Ho, H.-C.; Ebert, D.A.; de Carvalho, M.R.; Corrigan, S.; Rochel, E.; de Carvalho, M.; Tanaka, S.; Naylor, G.J.P. Redescription of Scymnodon ichiharai Yano and Tanaka 1984 (Squaliformes: Somniosidae) from the western North Pacific, with comments on the definition of somniosid genera. Ichthyol. Res. 2015, 62, 213–229. [Google Scholar] [CrossRef]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World, a Complete Guide; Princeton University Press: Princeton, NJ, USA, 2013; pp. 1–624. [Google Scholar]
- Ebert, D.A.; Compagno, L.J.V.; De Vries, M.J. A new lanternshark (Squaliformes: Etmopteridae: Etmopterus) from Southern Africa. Copeia 2011, 2011, 379–384. [Google Scholar] [CrossRef]
- Clarke, W.D. Function of bioluminescence in mesopelagic organisms. Nature 1963, 198, 1244–1246. [Google Scholar] [CrossRef]
- Denton, E.J.; Gilpin-Brown, J.B.; Wright, P.J. The angular distribution of the light produced by some mesopelagic fish in relation to their camouflage. Proc. R. Soc. B 1972, 182, 145–158. [Google Scholar] [CrossRef]
- Young, R.E.; Kampa, E.M.; Maynard, S.D.; Mencher, F.M.; Roper, C.F.E. Counterillumination and the upper depth limits of midwater animals. Deep Sea Res. Part I Oceanogr. Res. Pap. 1980, 27, 671–691. [Google Scholar] [CrossRef]
- McFall-Ngai, M.; Morin, J.G. Camouflage by disruptive illumination in Leiognathids, a family of shallow-water, bioluminescent fishes. J. Exp. Biol. 1991, 156, 119–137. [Google Scholar] [CrossRef]
- Claes, J.M.; Aksnes, D.L.; Mallefet, J. Phantom hunter of the fjords: Camouflage by counterillumination in a shark (Etmopterus spinax). J. Exp. Mar. Biol. Ecol. 2010, 388, 28–32. [Google Scholar] [CrossRef]
- Claes, J.M.; Dean, M.N.; Nilsson, D.-E.; Hart, N.S.; Mallefet, J. A deepwater fish with ‘lightsabers’—Dorsal spine-associated luminescence in a counterilluminating lanternshark. Sci. Rep. 2013, 3, 1308. [Google Scholar] [CrossRef] [PubMed]
- Duchatelet, L.; Pinte, N.; Tomita, T.; Sato, K.; Mallefet, J. Etmopteridae bioluminescence: Dorsal pattern specificity and aposematic use. Zool. Lett. 2019, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Claes, J.M.; Mallefet, J. Ecological Functions of shark luminescence. Luminescence 2014, 29, 13. [Google Scholar]
- Claes, J.M.; Mallefet, J. Functional physiology of lantern shark (Etmopterus spinax) luminescent pattern: Differential hormonal regulation of luminous zones. J. Exp. Biol. 2010, 213, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Claes, J.M.; Mallefet, J. Hormonal control of luminescence from lantern shark (Etmopterus spinax) photophores. J. Exp. Biol. 2009, 212, 3684–3692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinte, N.; Parisot, P.; Martin, U.; Zintzen, V.; De Vleeschouwer, C.; Roberts, C.D.; Mallefet, J. Ecological features and swimming capabilities of deep-sea sharks from New Zealand. Deep-Sea Res. I 2020, 156, 103187. [Google Scholar] [CrossRef]
- Pinte, N.; Coubris, C.; Jones, E.; Mallefet, J. Red and white muscle proportions and enzyme activities in mesopelagic sharks. Comp. Biochem. Physiol. B 2021, 256, 110649. [Google Scholar] [CrossRef]
- Claes, J.M.; Partridge, J.C.; Hart, N.S.; Garza-Gisholt, E.; Ho, H.-C.; Mallefet, J.; Collin, S.P. Photon hunting in the twilight zone: Visual features of mesopelagic bioluminescent sharks. PLoS ONE 2014, 9, e104213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claes, J.M.; Mallefet, J. Ontogeny of photophore pattern in the velvet belly lanternshark, Etmopterus spinax. Zoology 2009, 112, 433–441. [Google Scholar] [CrossRef]
- Santos, J.C.; Coloma, L.A.; Cannatella, D.C. Multiple, recurring origins of aposematism and diet specialization in poison frogs. Proc. Natl. Acad. Sci. USA 2003, 100, 12792–12797. [Google Scholar] [CrossRef] [Green Version]
- Marek, P.; Papaj, D.; Yeager, J.; Molina, S.; Moore, W. Bioluminescent aposematism in millipedes. Curr. Biol. 2011, 21, R680–R681. [Google Scholar] [CrossRef] [Green Version]
- Bolin, R.L. The function of the luminous organs of deep-sea fishes. In Proceedings of the 9th Pacific Science Congress, Bangkok, Thailand, 18 November–9 December 1957; Volume 10, pp. 37–39. [Google Scholar]
- Matsui, T.; Rosenblatt, R.H. Review of the deep-sea fish family Platytroctidae (Pisces: Salmoniformes). Bull. Scripps Inst. Oceanogr. Univ. Calif. 1987, 26, 1–159. [Google Scholar]
- Poulsen, J.Y. New observations and ontogenetic transformation of photogenic tissues in the tubeshoulder Sagamichthys schnakenbecki (Platytroctidae, Alepocephaliformes). J. Fish Biol. 2019, 94, 62–76. [Google Scholar] [CrossRef]
- Davis, A.L.; Sutton, T.T.; Kier, W.M.; Johnsen, S. Evidence that eye-facing photophores serve as a reference for counterillumination in an order of deep-sea fishes. Proc. R. Soc. B 2020, 287, 20192918. [Google Scholar] [CrossRef]
- Herring, P.J.; Cope, C. Red bioluminescence in fishes: On the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. Mar. Biol. 2005, 148, 383–394. [Google Scholar] [CrossRef]
- Renwart, M.; Delroisse, J.; Flammang, P.; Claes, J.M.; Mallefet, J. Cytological changes during luminescence production in lanternshark (Etmopterus spinax Linnaeus, 1758) photophores. Zoomorphology 2015, 134, 107–116. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. The lantern shark’s light switch: Turning shallow water crypsis into midwater camouflage. Biol. Lett. 2010, 6, 685–687. [Google Scholar] [CrossRef]
- Duchatelet, L.; Sugihara, T.; Delroisse, J.; Koyanagi, M.; Rezsohazy, R.; Terakita, A.; Mallefet, J. From extraocular photoreception to pigment movement regulation: A new control mechanism of the lanternshark luminescence. Sci. Rep. 2020, 10, 10195. [Google Scholar] [CrossRef]
- Claes, J.M.; Krönström, J.; Holmgren, S.; Mallefet, J. Nitric oxide in the control of luminescence from lantern shark (Etmopterus spinax) photophores. J. Exp. Biol. 2010, 213, 3005–3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reif, W.E. Functions of scales and photophores in mesopelagic luminescent sharks. Acta Zool. 1985, 66, 111–118. [Google Scholar] [CrossRef]
- Ferrón, H.G.; Paredes-Aliaga, M.V.; Martínez-Pérez, C.; Botella, H. Bioluminescent-like squamation in the galeomorph shark Apristurus ampliceps (Chondrichthyes: Elasmobranchii). Contrib. Zool. 2018, 87, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Christophe, B.; Baguet, F. The adrenergic control of the photocytes luminescence of the Porichthys photophores. Comp. Bioch. Physiol. C Comp. Pharmacol. 1985, 81, 359–365. [Google Scholar] [CrossRef]
- Mallefet, J.; Anctil, M. Immunohistochemical detection of biogenic amines in the photophores of the midshipman fish Porichthys notatus. Can. J. Zool. 1992, 70, 1968–1975. [Google Scholar] [CrossRef]
- Zaccone, G.; Abelli, L.; Salpietro, L.; Zaccone, D.; Macrì, B.; Marino, F. Nervous control of photophores in luminescent fishes. Acta Hictochem. 2011, 113, 387–394. [Google Scholar] [CrossRef]
- Mallefet, J.; Duchatelet, L.; Hermans, C.; Baguet, F. Luminescence control of Stomiidae photophores. Acta Histochem. 2019, 121, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Visconti, M.A.; Ramanzini, G.C.; Camargo, C.R.; Castrucci, A.M.L. Elasmobranch color change: A short review and novel data on hormone regulation. J. Exp. Zool. 1999, 284, 485–491. [Google Scholar] [CrossRef]
- Gelsleichter, J.; Evans, A.N. Hormonal regulation of elasmobranch physiology. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; Chapter 11; pp. 314–338. [Google Scholar]
- Nagai, M.; Oshima, N.; Fujii, R. A comparative study of melanin concentrating hormone (MCH) action on teleost melanophores. Biol. Bull. 1986, 171, 360–370. [Google Scholar] [CrossRef]
- Oshima, N.; Kasukawa, H.; Fujii, R.; Wilkes, B.C.; Hruby, V.J.; Hadley, M.E. Action of melanin-concentrating hormone (MCH) on teleost chromatophores. Gen. Comp. Endocrinol. 1986, 64, 381–388. [Google Scholar] [CrossRef]
- Mizusawa, K.; Kobayashi, Y.; Sunuma, T.; Asahida, T.; Saito, Y.; Takahashi, A. Inhibiting roles of melanin-concentrating hormone for skin pigment dispersion in barfin flounder, Verasper moseri. Gen. Comp. Endocrinol. 2011, 171, 75–81. [Google Scholar] [CrossRef]
- Mizusawa, K.; Amiya, N.; Yamaguchi, Y.; Takabe, S.; Amano, M.; Breves, J.P.; Fox, B.K.; Grau, E.G.; Hyodo, S.; Takahashi, A. Identification of mRNAs coding for mammalian-type melanin-concentrating hormone and its receptors in the scalloped hammerhead shark Sphyrna lewini. Gen. Comp. Endocrinol. 2012, 179, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Duchatelet, L.; Delroisse, J.; Mallefet, J. Melanin-concentrating hormone is not involved in luminescence emission in the velvet belly lanternshark, Etmopterus spinax. Mar. Biol. 2019, 166, 140. [Google Scholar] [CrossRef]
- Duchatelet, L.; Delroisse, J.; Mallefet, J. Bioluminescence in lanternsharks: Insight from hormone receptor localization. Gen. Comp. Endocrinol. 2020, 294, 113488. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Control of luminescence from lantern shark (Etmopterus spinax) photophores. Commun. Integr. Biol. 2011, 4, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Claes, J.M.; Krönström, J.; Holmgren, S.; Mallefet, J. GABA inhibition of luminescence from lantern shark (Etmopterus spinax) photophores. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 231–236. [Google Scholar] [CrossRef]
- Ocampo Daza, D.; Larhammar, D. Evolution of the receptors for growth hormone, prolactin, erythropoietin and thrombopoietin in relation to the vertebrate tetraploidizations. Gen. Comp. Endocrinol. 2018, 257, 143–160. [Google Scholar] [CrossRef]
- Vanacek, J. Cellular mechanisms of melatonin action. Physiol. Rev. 1998, 78, 687–721. [Google Scholar] [CrossRef]
- Busca, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Sugden, D.; Davidson, K.; Hough, K.A.; The, M.-T. Melatonin, melatonin receptors and melanophores: A moving story. Pigment Cell Res. 2004, 17, 454–460. [Google Scholar] [CrossRef]
- Visconti, M.A.; Castrucci, A.M.L. Melanotropin receptors in the cartilaginous fish, Potamotrygon reticulatus and in the lungfish, Lepidosiren paradoxa. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1993, 106, 523–528. [Google Scholar] [CrossRef]
- Nery, L.E.M.; Castrucci, A.M.L. Pigment cell signalling for physiological color change. Comp. Biochem. Physiol. A Mol. Interg. Physiol. 1997, 118, 1135–1144. [Google Scholar] [CrossRef]
- Tong, D.; Rozas, N.S.; Oakley, T.H.; Mitchell, J.; Colley, N.J.; McFall-Ngai. Evidence for light perception in a bioluminescent organ. Proc. Natl. Acad. Sci. USA 2009, 106, 9836–9841. [Google Scholar] [CrossRef] [Green Version]
- McFall-Ngai, M.; Heath-Heckman, E.A.C.; Gillette, A.A.; Peyer, S.M.; Harvie, E.A. The secret languages of coevolved symbioses: Insights from the Euprymna scolopes-Vibrio fischeri symbiosis. Semin. Immunol. 2012, 24, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, C.E.; Pang, K.; Powers, M.L.; Reitzel, A.M.; Ryan, J.F.; Simmons, D.; Tada, T.; Park, M.; Gupta, J.; Brooks, S.Y.; et al. Genomic organization, evolution, and expression of photoprotein and opsin genes in Mnemiopsis leidyi: A new view of ctenophore photocytes. BMC Biol. 2012, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delroisse, J.; Ullrich-Luter, E.; Ortega-Martinez, O.; Dupont, S.; Arnone, M.I.; Mallefet, J.; Flammang, P. High opsin diversity in a non-visual infaunal brittle star. BMC Genom. 2014, 15, 1035. [Google Scholar] [CrossRef] [Green Version]
- Delroisse, J.; Ullrich-Lüter, E.; Blaue, S.; Ortega-Martinez, O.; Eeckhaut, I.; Flammang, P.; Mallefet, J. Fine structure of the luminous spines and luciferase detection in the brittle star Amphiura filiformis. Zool. Anz. 2017, 269, 1–12. [Google Scholar] [CrossRef]
- Bracken-Grissom, H.D.; DeLeo, D.M.; Porter, M.L.; Iwanicki, T.; Sickles, J.; Frank, T.M. Light organ photosensitivity in deep-sea shrimp may suggest a novel role in counterillumination. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Delroisse, J.; Duchatelet, L.; Flammang, P.; Mallefet, J. De novo transcriptome analyses provide insights into opsin-based photoreception in the lanternshark Etmopterus spinax. PLoS ONE 2018, 13, e0209767. [Google Scholar] [CrossRef] [PubMed]
- McCapra, F. Chemical mechanisms in bioluminescence. Acc. Chem. Res. 1976, 9, 201–208. [Google Scholar] [CrossRef]
- Kaskova, Z.M.; Tsarkova, A.S.; Yampolsky, I.V. 1001 lights: Luciferins, luciferases, their mechanisms of action and applications in chemical analysis, biology and medicine. Chem. Soc. Rev. 2016, 45, 6048–6077. [Google Scholar] [CrossRef]
- Warner, J.A.; Case, J.F. The zoogeography and dietary induction of bioluminescence in the midshipman fish, Porichthys notatus. Biol. Bull. 1980, 159, 231–246. [Google Scholar] [CrossRef]
- Frank, T.M.; Widder, E.A.; Latz, M.I.; Case, J.F. Dietary maintenance of bioluminescence in a deep-sea mysid. J. Exp. Biol. 1984, 109, 385–389. [Google Scholar] [CrossRef]
- Thompson, E.M.; Nafpaktitis, B.G.; Tsuji, F.I. Dietary uptake and blood transport of Vargula (crustacean) luciferin in the bioluminescent fish, Porichthys notatus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1988, 89, 203–209. [Google Scholar] [CrossRef]
- Haddock, S.H.D.; Rivers, T.J.; Robison, B.H. Can coelenterates make coelenterazine? Dietary requirement for luciferin in cnidarian bioluminescence. Proc. Natl. Acad. Sci. USA 2001, 98, 11151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallefet, J.; Duchatelet, L.; Coubris, C. Bioluminescence induction in the ophiuroid Amphiura filiformis (Echinodermata). J. Exp. Biol. 2020, 223, jeb218719. [Google Scholar] [CrossRef] [PubMed]
- Bessho-Uehara, M.; Yamamoto, N.; Shigenobu, S.; Mori, H.; Kuwata, K.; Oba, Y. Kleptoprotein bioluminescence: Parapriacanthus fish obtain luciferase from ostracod prey. Sci. Adv. 2020, 6, eaax4942. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.M.; Herring, P.J.; Campbell, A.K. Evidence for de novo biosynthesis of coelenterazine in the bioluminescent midwater shrimp, Systellaspis debilis. J. Mar. Biol. Assoc. UK 1995, 75, 165–171. [Google Scholar] [CrossRef]
- Oba, Y.; Kato, S.-I.; Ojika, M.; Inouye, S. Biosynthesis of coelenterazine in the deep-sea copepod, Metridia pacifica. Biochem. Biophys. Res. Commun. 2009, 390, 684–688. [Google Scholar] [CrossRef]
- Bessho-Uehara, M.; Huang, W.; Patry, W.L.; Browne, W.E.; Weng, J.-K.; Haddock, S.H.D. Evidence for de novo biosynthesis of the luminous substrate coelenterazine in ctenophores. iScience 2020, 23, 101859. [Google Scholar] [CrossRef]
- Shimomura, O. Presence of coelenterazine in non-bioluminescent marine organisms. Comp. Biochem. Physiol. 1987, 86B, 361–363. [Google Scholar] [CrossRef]
- Mallefet, J.; Shimomura, O. Presence of coelenterazine in mesopelagic fishes from the Strait of Messina. Mar. Biol. 1995, 124, 381–385. [Google Scholar] [CrossRef]
- Thomson, C.M.; Herring, P.J.; Campbell, A.K. The widespread occurrence and tissue distribution of the imidazolopyrazine luciferins. J. Biol. Chem. 1997, 2, 87–91. [Google Scholar] [CrossRef]
- Duchatelet, L.; Hermans, C.; Duhamel, G.; Cherel, Y.; Guinet, C.; Mallefet, J. Coelenterazine detection in five myctophid species from the Kerguelen Plateau. In The Kerguelen Plateau: Marine Ecosystem and Fisheries. Proceedings of the Second Symposium; Welsford, D., Dell, J., Duhamel, G., Eds.; Australian Antartic Division: Kingston, TAS, Australia; pp. 31–41.
- Renwart, M.; Mallefet, J. First study of the chemistry of the luminous system in a deep-sea shark, Etmopterus spinax Linnaeus, 1758 (Chondrichthyes: Etmopteridae). J. Exp. Mar. Biol. Ecol. 2013, 448, 214–219. [Google Scholar] [CrossRef]
- Stingo, V.; Du Buit, M.-H.; Odierna, G. Genome size of some selachian fishes. Boll. Zool. 1980, 47, 129–137. [Google Scholar] [CrossRef]
- Ojima, Y.; Yamamoto, K. Cellular DNA contents of fishes determined by flow cytometry. La Kromosomo II 1990, 57, 1871–1888. [Google Scholar]
- Hardie, D.C.; Hebert, P.D.N. The nucleotypic effects of cellular DNA content in cartilaginous and ray-finned fishes. Genome 2003, 46, 683–706. [Google Scholar] [CrossRef] [Green Version]
- Marra, N.J.; Stanhope, M.J.; Jue, N.K.; Wang, M.; Sun, Q.; Bitar, P.P.; Richards, V.P.; Komissarov, A.; Rayko, M.; Kliver, S.; et al. White shark genome reveals ancient elasmobranch adaptations associated with wound healing and the maintenance of genome stability. Proc. Natl. Acad. Sci. USA 2019, 116, 4446–4455. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Park, S.G.; Luria, V.; Jeon, S.; Kim, H.-M.; Jeon, Y.; Bhak, Y.; Jun, J.H.; Kim, S.W.; Hong, W.H.; et al. The whale shark genome reveals how genomic and physiological properties scale with body size. Proc. Natl. Acad. Sci. USA 2020, 117, 20662–20671. [Google Scholar] [CrossRef]
- Dufresne, F.; Jeffery, N. A guided tour of large genome size in animals: What we know and where we are heading. Chromosome Res. 2011, 19, 925–938. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Duchatelet, L.; Claes, J.M.; Delroisse, J.; Flammang, P.; Mallefet, J. Glow on Sharks: State of the Art on Bioluminescence Research. Oceans 2021, 2, 822-842. https://doi.org/10.3390/oceans2040047
AMA Style
Duchatelet L, Claes JM, Delroisse J, Flammang P, Mallefet J. Glow on Sharks: State of the Art on Bioluminescence Research. Oceans. 2021; 2(4):822-842. https://doi.org/10.3390/oceans2040047
Chicago/Turabian StyleDuchatelet, Laurent, Julien M. Claes, Jérôme Delroisse, Patrick Flammang, and Jérôme Mallefet. 2021. "Glow on Sharks: State of the Art on Bioluminescence Research" Oceans 2, no. 4: 822-842. https://doi.org/10.3390/oceans2040047