
Morphological and Optical Modification of Melanosomes in Fish Integuments upon Oxidation

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Treatment
2.2. Light Microscopy
2.3. Scanning Electron Microscopy
2.4. Time-Resolved and Steady-State Fluorimetry
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinoshita, S. Structural Colors in the Realm of Nature; World Scientific: Singapore, 2008. [Google Scholar]
- Mouchet, S.R.; Vukusic, P. Structural colours in lepidopteran scales. In Advances in Insect Physiology; Elsevier: London, UK, 2018; Volume 54, pp. 1–53. [Google Scholar] [CrossRef]
- Mouchet, S.R.; Deparis, O. Natural Photonics and Bioinspiration; Artech House: Norwood, MA, USA, 2021; Available online: https://us.artechhouse.com/Natural-Photonics-and-Bioinspiration-P2221.aspx (accessed on 28 June 2023).
- Denton, E. Reflectors in Fishes. Sci. Am. 1971, 224, 64–72. [Google Scholar] [CrossRef]
- Brown, F., Jr. Chromatophores and color change. In Comparative Animal Physiology; WB Saunders: Philadelphia, PA, USA, 1973; pp. 915–950. [Google Scholar]
- Hawkes, J.W. The structure of fish skin. Cell Tissue Res. 1974, 149, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Kelsh, R.N.; Schmid, B.; Eisen, J.S. Genetic analysis of melanophore development in zebrafish embryos. Dev. Biol. 2000, 225, 277–293. [Google Scholar] [CrossRef]
- Michiels, N.; Anthes, N.; Hart, N.; Herler, J.; Meixner, A.J.; Schleifenbaum, F.; Schulte, G.; Siebeck, U.E.; Sprenger, D.; Wucherer, M.F. Red fluorescence in reef fish: A novel signalling mechanism? BMC Ecol. 2008, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Lagorio, M.G.; Cordon, G.B.; Iriel, A. Reviewing the relevance of fluorescence in biological systems. Photochem. Photobiol. Sci. 2015, 14, 1538–1559. [Google Scholar] [CrossRef]
- Marshall, J.; Johnsen, S. Fluorescence as a means of colour signal enhancement. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160335. [Google Scholar] [CrossRef]
- Bond, C. Biology of Fishes; WB Sounders Company: Philadelphia, PA, USA, 1979. [Google Scholar]
- Stepien, C.A. Color pattern and habitat differences between male, female and juvenile giant kelpfish (Blennioidei: Clinidae). Bull. Mar. Sci. 1987, 41, 45–58. [Google Scholar]
- Hakkinen, J.; Korhonen, H.; Oikari, A.; Karjalainen, J. Melanin concentrations in vendace (Coregonus albula) Whitefish (Coregonus lavaretus) Larvae Five Boreal lakes with different optical properties. Boreal Environ. Res. 2003, 8, 193–202. [Google Scholar]
- Stieb, S.M.; Cortesi, F.; Sueess, L.; Carleton, K.L.; Salzburger, W.; Marshall, N.J. Why UV vision and red vision are important for damselfish (Pomacentridae): Structural and expression variation in opsin genes. Mol. Ecol. 2017, 26, 1323–1342. [Google Scholar] [CrossRef]
- Blough, N.V.; Zepp, R.G. Reactive oxygen species in natural waters. In Active Oxygen in Chemistry; Springer: Dordrecht, the Netherlands, 1995; pp. 280–333. [Google Scholar]
- Manjun, Z. Determination of photochemically-generated reactive oxygen species in natural water. J. Environ. Sci. 2009, 21, 303–306. [Google Scholar]
- Kieber, D.J.; Peake, B.; Scully, N.M. Reactive oxygen species in aquatic ecosystems. In UV Effects in Aquatic Organisms and Ecosystems; Helbling, E.W., Ed.; Royal Society of Chemistry: London, UK, 2003; pp. 251–276. [Google Scholar]
- Morris, J.J.; Rose, A.L.; Lu, Z. Reactive oxygen species in the world ocean and their impacts on marine ecosystems. Redox Biol. 2022, 52, 102285. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.M. Photochemical and biochemical controls on reactive oxygen and iron speciation in the pelagic surface ocean. Mar. Chem. 2008, 109, 152–164. [Google Scholar] [CrossRef]
- Müller, R.; Desel, C.; Steinhoff, F.S.; Wiencke, C.; Bischof, K. UV-radiation and elevated temperatures induce formation of reactive oxygen species in gametophytes of cold-temperate/Arctic kelps (Laminariales, Phaeophyceae). Phycol. Res. 2012, 60, 27–36. [Google Scholar] [CrossRef]
- Celis-Plá, P.S.; Moenne, F.; Rodríguez-Rojas, F.; Pardo, D.; Lavergne, C.; Moenne, A.; Brown, M.T.; Huovinen, P.; Gómez, I.; Navarro, N.; et al. Antarctic intertidal macroalgae under predicted increased temperatures mediated by global climate change: Would they cope? Sci. Total Environ. 2020, 740, 140379. [Google Scholar] [CrossRef]
- Allen, G.R.; Emery, A.R. A review of the pomacentrid fishes of the genus Stegastes from the Indo-Pacific, with descriptions of two new species. In Indo-Pacific Fishes; Bishop Museum: Honolulu, HI, USA, 1985; Volume 3, pp. 1–31. [Google Scholar]
- Sköld, H.N.; Aspengren, S.; Cheney, K.L.; Wallin, M. Fish Chromatophores—From Molecular Motors to Animal Behavior. Int. Rev. Cell Mol. Biol. 2016, 321, 171–219. [Google Scholar] [CrossRef]
- Salis, P.; Lorin, T.; Laudet, V.; Frédérich, B. Magic Traits in Magic Fish: Understanding Color Pattern Evolution Using Reef Fish. Trends Genet. 2019, 35, 265–278. [Google Scholar] [CrossRef]
- Smith, R.; Garrett, B.; Naqvi, K.; Fülöp, A.; Godfrey, S.; Marsh, J.; Chechik, V. Mechanistic insights into the bleaching of melanin by alkaline hydrogen peroxide. Free Radic. Biol. Med. 2017, 108, 110–117. [Google Scholar] [CrossRef]
- Goda, M.; Fujii, R. Coloration and Chromatophores of the Domino Damsel, Dascyllus trimaculatus. Zool. Sci. 2001, 18, 165–174. [Google Scholar] [CrossRef]
- Futter, C.E.; Ramalho, J.S.; Jaissle, G.B.; Seeliger, M.W.; Seabra, M.C. The Role of Rab27a in the Regulation of Melanosome Distribution within Retinal Pigment Epithelial Cells. Mol. Biol. Cell 2004, 15, 2264–2275. [Google Scholar] [CrossRef]
- Franzen, A.J.; Cunha, M.M.; Miranda, K.; Hentschel, J.; Plattner, H.; da Silva, M.B.; Salgado, C.G.; de Souza, W.; Rozental, S. Ultrastructural characterization of melanosomes of the human pathogenic fungus Fonsecaea pedrosoi. J. Struct. Biol. 2008, 162, 75–84. [Google Scholar] [CrossRef]
- Amiri, M.; Shaheen, H.M. Chromatophores and color revelation in the blue variant of the Siamese fighting fish (Betta splendens). Micron 2012, 43, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Naaz, I. Comparative light and electron microscopic studies of dorsal skin melanophores of Indian toad, Bufo melanostictus. J. Microsc. Ultrastruct. 2014, 2, 230–235. [Google Scholar] [CrossRef]
- Mouchet, S.R.; Van Hooijdonk, E.; Welch, V.L.; Louette, P.; Colomer, J.F.; Su, B.L.; Deparis, O. Liquid-induced colour change in a beetle: The concept of a photonic cell. Sci. Rep. 2016, 6, 19322. [Google Scholar] [CrossRef]
- Dhungel, B.; Ohno, Y.; Matayoshi, R.; Iwasaki, M.; Taira, W.; Adhikari, K.; Gurung, R.; Otaki, J.M. Distal-less induces elemental color patterns in Junonia butterfly wings. Zool. Lett. 2016, 2, 4. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Introduction to fluorescence. In Principles of Fluorescence Spectroscopy; Springer: New York, NY, USA, 1999; pp. 1–23. [Google Scholar]
- Lindgren, J.; Moyer, A.; Schweitzer, M.H.; Sjövall, P.; Uvdal, P.; Nilsson, D.E.; Heimdal, J.; Engdahl, A.; Gren, J.A.; Schultz, B.P.; et al. Interpreting melanin-based coloration through deep time: A critical review. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150614. [Google Scholar] [CrossRef] [PubMed]
- Donatti, L.; Fanta, E. Morphology of the retina in the freshwater fish Metynnis Roosevelti Eiggenmann (Characidae, Serrasalminae) and the effects of monochromatic red light. Rev. Bras. Zool. 1999, 16, 151–173. [Google Scholar] [CrossRef]
- Kawauchi, H. Functions of Melanin-Concentrating Hormone in Fish. J. Exp. Zool. 2006, 305A, 751–760. [Google Scholar] [CrossRef]
- Wolnicka-Glubis, A.; Pecio, A.; Podkowa, D.; Kolodziejczyk, L.M.; Plonka, P.M. Pheomelanin in the skin of Hymenochirus Boettgeri (Amphibia: Anura: Pipidae). Exp. Dermatol. 2012, 21, 535–561. [Google Scholar] [CrossRef]
- Colleary, C.; Dolocan, A.; Gardner, J.; Singh, S.; Wuttke, M.; Rabenstein, R.; Habersetzer, J.; Schaal, S.; Feseha, M.; Clemens, M.; et al. Chemical, experimental, and morphological evidence for diagenetically altered melanin in exceptionally preserved fossils. Proc. Natl. Acad. Sci. USA 2015, 112, 12592–12597. [Google Scholar] [CrossRef]
- Kottler, V.A.; Künstner, A.; Schartl, M. Pheomelanin in fish? Pigment Cell Melanoma Res. 2015, 28, 355–356. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, X.; Wang, X.; Li, J.; Liu, G.; Kuang, Y.; Xu, J.; Zheng, X.; Ren, L.; Wang, G.; et al. Genome sequence and genetic diversity of the common carp, Cyprinus Carpio. Nat. Genet. 2014, 46, 1212–1219. [Google Scholar] [CrossRef]
- Riley, P. Materia Melanica: Further Dark Thoughts. Pigment Cell Res. 1992, 5, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Bonser, R.H.C. Melanin and the Abrasion Resistance of Feathers. Condor 1995, 97, 590–591. [Google Scholar]
- Liu, Y.; Hong, L.; Wakamatsu, K.; Ito, S.; Adhyaru, B.; Cheng, C.Y.; Bowers, C.R.; Simon, J.D. Comparison of Structural and Chemical Properties of Black and Red Human Hair Melanosomes. Photochem. Photobiol. 2005, 81, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Moses, D.N.; Harreld, J.H.; Stucky, G.D.; Waite, J.H. Melanin and Glycera Jaws. J. Biol. Chem. 2006, 281, 34826–34832. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, R.W.; Schreiber, E.A.; Peele, A.M.; Burtt, E.H. Pattern of Damage to Albino Great Frigatebird Flight Feathers Supports Hypothesis of Abrasion by Airborne Particles. Condor 2006, 108, 736–741. [Google Scholar] [CrossRef]
- Pannkuk, E.L.; Siefferman, L.M.; Butts, J.A. Colour phases of the eastern screech owl: A comparison of biomechanical variables of body contour feathers. Funct. Ecol. 2010, 24, 347–353. [Google Scholar] [CrossRef]
- Galván, I.; Solano, F. Bird Integumentary Melanins: Biosynthesis, Forms, Function and Evolution. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Wang, Y.; Li, T.; Wang, X.; Ma, P.; Bai, H.; Dong, W.; Xie, Y.; Chen, M. Superior Performance of Polyurethane Based on Natural Melanin Nanoparticles. Biomacromolecules 2016, 17, 3782–3789. [Google Scholar] [CrossRef]
- Huang, Z.; Zeng, H.; Hamzavi, I.; Alajlan, A.; Tan, E.; McLean, D.I.; Lui, H. Cutaneous melanin exhibiting fluorescence emission under near-infrared light excitation. J. Biomed. Opt. 2006, 11, 034010. [Google Scholar] [CrossRef]
- Fernandes, B.; Matamá, T.; Guimarães, D.; Gomes, A.; Cavaco-Paulo, A. Fluorescent quantification of melanin. Pigment Cell Melanoma Res. 2016, 29, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, M.; Goujon, D.; Conde, R.; Forrer, M.; van den Bergh, H.; Wagnières, G. Absolute autofluorescence spectra of human healthy, metaplastic, and early cancerous bronchial tissue in vivo. Appl. Opt. 2001, 40, 3784–3791. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Wang, W.; Yang, Y.; Alfano, R.R. Native fluorescence spectra of human cancerous and normal breast tissues analyzed with non-negative constraint methods. Appl. Opt. 2013, 52, 1293–1301. [Google Scholar] [CrossRef]
- Sachs, H.W. Über die autogenen Pigmente, besonders das Lipofuscin und seine Abrenzung von Melanin. Beitr. Pathol. Anat. Allg. Pathol. 1943, 108, 268–314. [Google Scholar]
- Kayatz, P.; Thumann, G.; Luther, T.T.; Jordan, J.F.; Bartz–Schmidt, K.U.; Esser, P.J.; Schraermeyer, U. Oxidation Causes Melanin Fluorescence. Investig. Ophthalmol. Vis. Sci. 2001, 42, 241–246. [Google Scholar]
- Sarathy, S.R.; Mohseni, M. The impact of UV/H2O2 advanced oxidation on molecular size distribution of chromophoric natural organic matter. Environ. Sci. Technol. 2007, 41, 8315–8320. [Google Scholar] [CrossRef]
- Iriel, A.; Lagorio, M.G. Is the flower fluorescence relevant in biocommunication? Naturwissenschaften 2010, 97, 915–924. [Google Scholar] [CrossRef]
- Iriel, A.; Lagorio, M.G. Implications of reflectance and fluorescence of Rhododendr. Indicum Flowers Biosignaling. Photochem. Photobiol. Sci. 2010, 9, 342–348. [Google Scholar] [CrossRef]
- Taboada, C.; Brunetti, A.E.; Pedron, F.N.; Carnevale Neto, F.; Estrin, D.A.; Bari, S.E.; Chemes, L.B.; Peporine Lopes, N.; Lagorio, M.G.; Faivovich, J. Naturally occurring fluorescence in frogs. Proc. Natl. Acad. Sci. USA 2017, 114, 3672–3677. [Google Scholar] [CrossRef]
- Ladouce, M.; Barakat, T.; Su, B.L.; Deparis, O.; Mouchet, S.R. Scattering of ultraviolet light by avian eggshells. Faraday Discuss. 2020, 223, 63–80. [Google Scholar] [CrossRef]
- Mouchet, S.R.; Verstraete, C.; Bokic, B.; Mara, D.; Dellieu, L.; Orr, A.G.; Deparis, O.; Van Deun, R.; Verbiest, T.; Vukusic, P.; et al. Revealing natural fluorescence in transparent insect wings by linear and nonlinear optical techniques. J. Lumin. 2023, 254, 119490. [Google Scholar] [CrossRef]
- Hulburt, E.O. The Penetration of Ultraviolet Light into Pure Water and Sea Water. J. Opt. Soc. Am. 1928, 17, 15–22. [Google Scholar] [CrossRef]
- Moan, J. Visible light and UV radiation. In Radiation at Home, Outdoors and in the Workplace; Brune, A., Hellborg, R., Persson, B., Pääkkönen, R., Eds.; Scandinavian Science Publisher: Oslo, Norway, 2001; pp. 69–85. [Google Scholar]


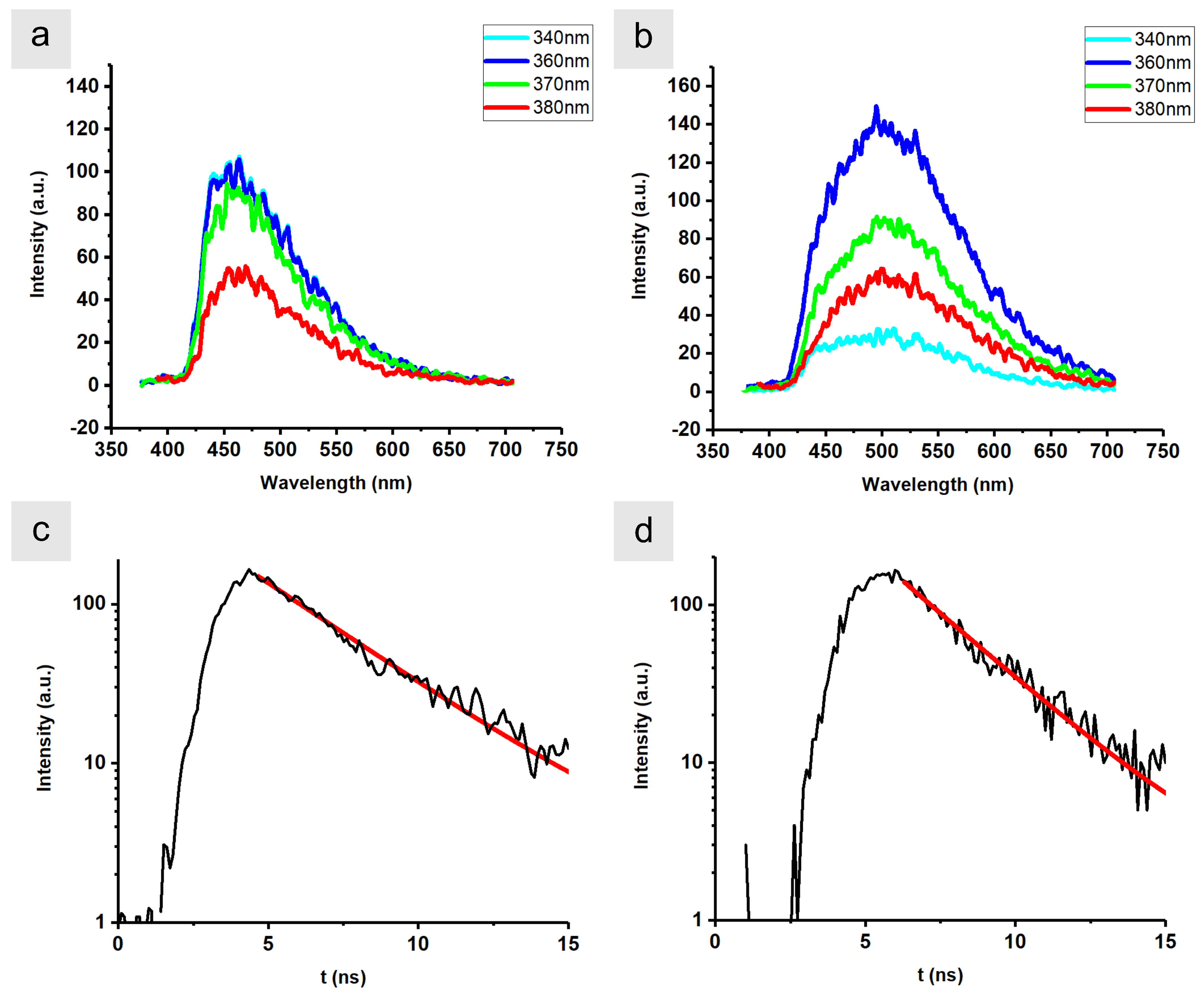
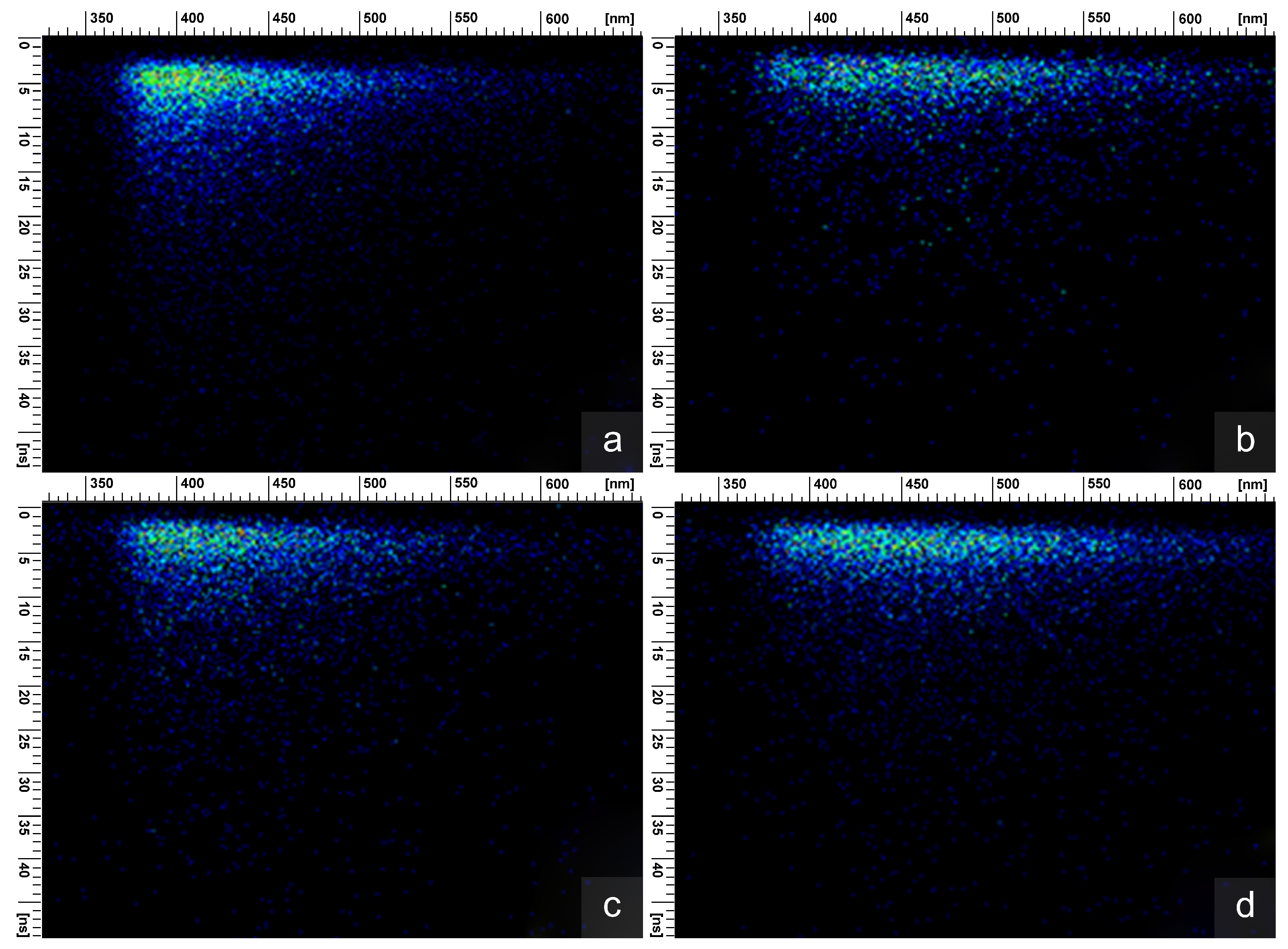

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavelength (nm) | ||||
|---|---|---|---|---|
| 340 | 360 | 370 | 380 | |
| (nm) | 474 | 476 | 474 | 478 |
| (nm) | 523 | 524 | 528 | 531 |
| (ns) | 2.70 | 2.25 | 2.69 | 2.61 |
| (ns) | 1.66 | 1.75 | 1.92 | 2.02 |
| (nm) | 93 | 94 | 86 | 101 |
| (nm) | 122 | 130 | 126 | 126 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouchet, S.R.; Cortesi, F.; Bokic, B.; Lazovic, V.; Vukusic, P.; Marshall, N.J.; Kolaric, B. Morphological and Optical Modification of Melanosomes in Fish Integuments upon Oxidation. Optics 2023, 4, 563-572. https://doi.org/10.3390/opt4040041
Mouchet SR, Cortesi F, Bokic B, Lazovic V, Vukusic P, Marshall NJ, Kolaric B. Morphological and Optical Modification of Melanosomes in Fish Integuments upon Oxidation. Optics. 2023; 4(4):563-572. https://doi.org/10.3390/opt4040041
Chicago/Turabian StyleMouchet, Sébastien R., Fabio Cortesi, Bojana Bokic, Vladimir Lazovic, Pete Vukusic, N. Justin Marshall, and Branko Kolaric. 2023. "Morphological and Optical Modification of Melanosomes in Fish Integuments upon Oxidation" Optics 4, no. 4: 563-572. https://doi.org/10.3390/opt4040041