
Irradiation of ZnPPIX Complexed with Bovine β-Lactoglobulin Causes Chemical Modifications and Conformational Changes of the Protein

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Buffers and Stock Solutions
2.3. Sample Preparation
2.4. Experimental Techniques
2.4.1. UV-Vis Absorption Spectroscopy
2.4.2. Steady-State Fluorescence Spectroscopy
2.4.3. Fluorescence Lifetime
2.5. Laser Irradiation
2.6. Femtosecond Transient Absorption Spectroscopy (FTAS)
2.7. 1O2 Detection
2.8. Native Gel Electrophoresis
3. Results and Discussion
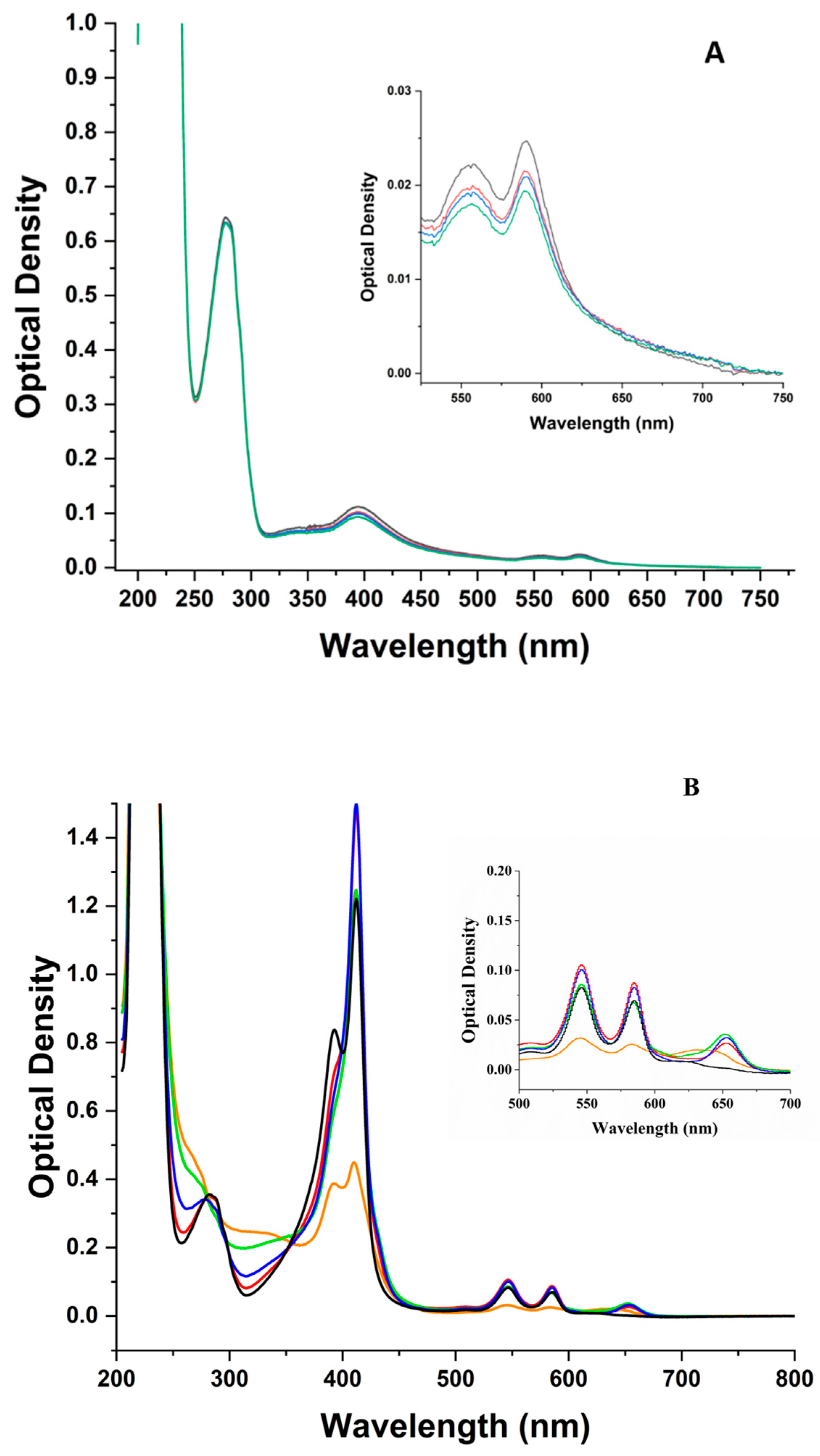
3.1. UV-Visible Absorption
3.1.1. pH 5
3.1.2. pH 9
3.2. BLG Emission
3.2.1. pH 5
3.2.2. pH 9
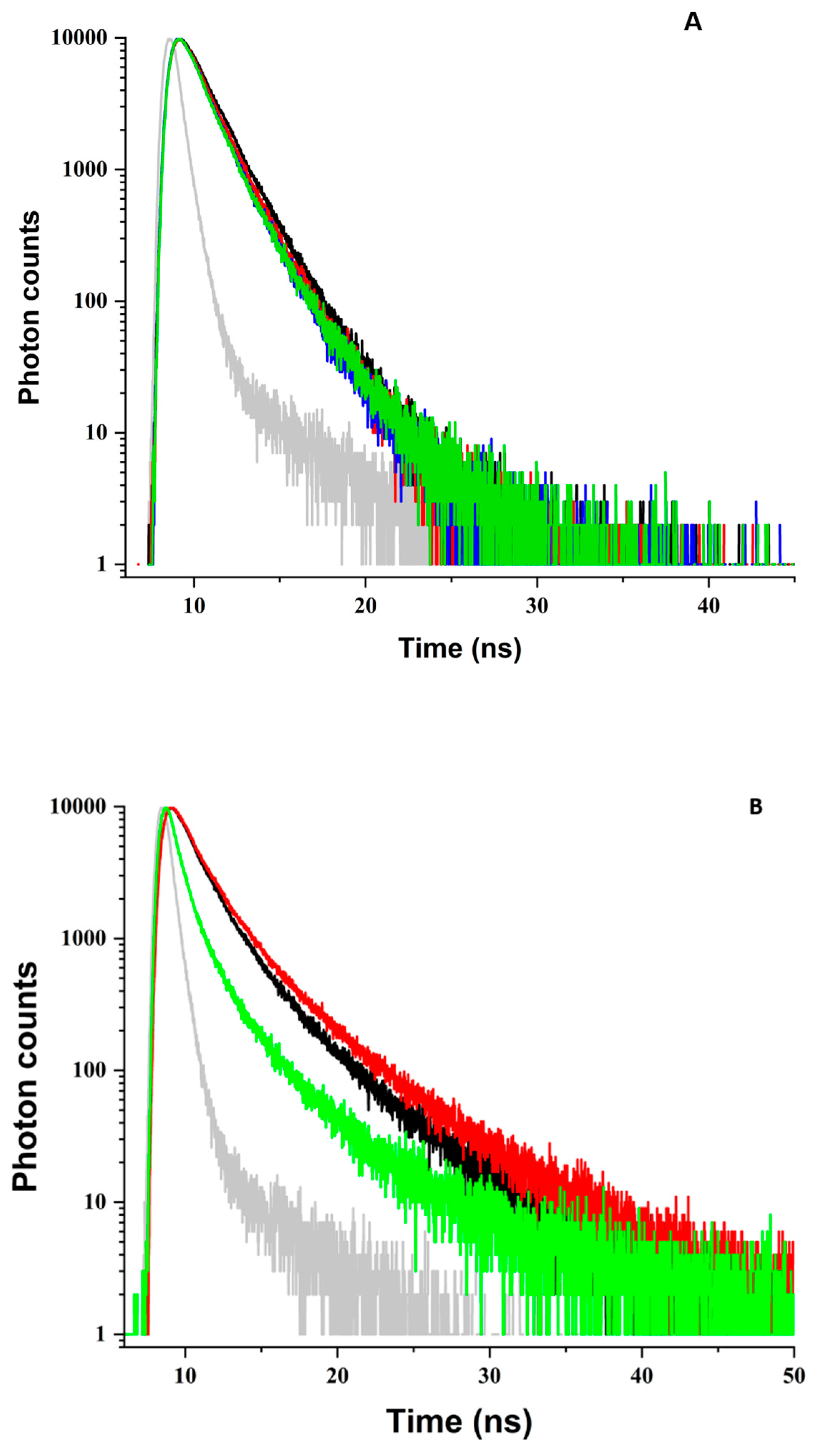
3.3. BLG Fluorescence Decay
3.3.1. pH 5
3.3.2. pH 9
3.3.3. Partial Summary
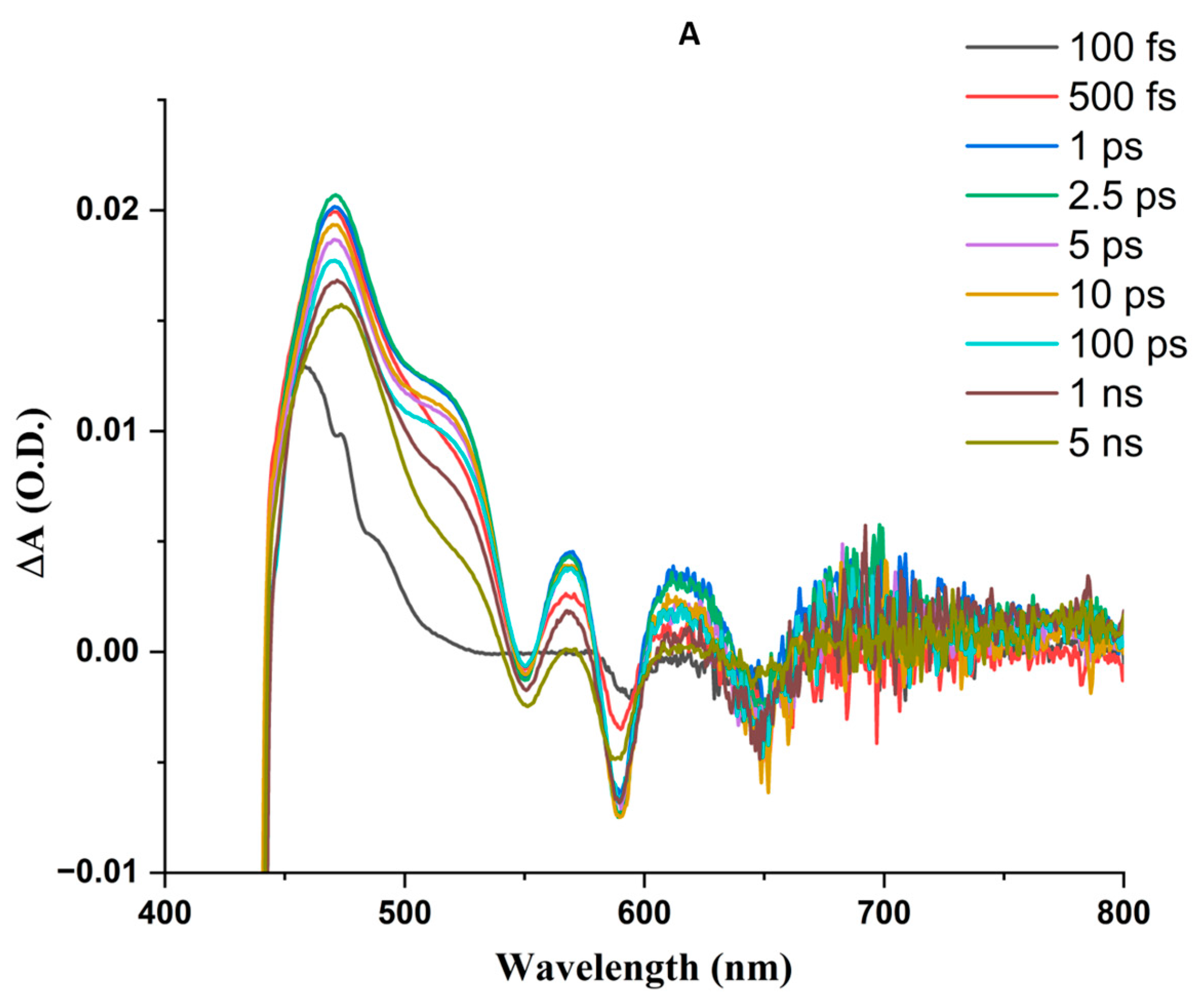
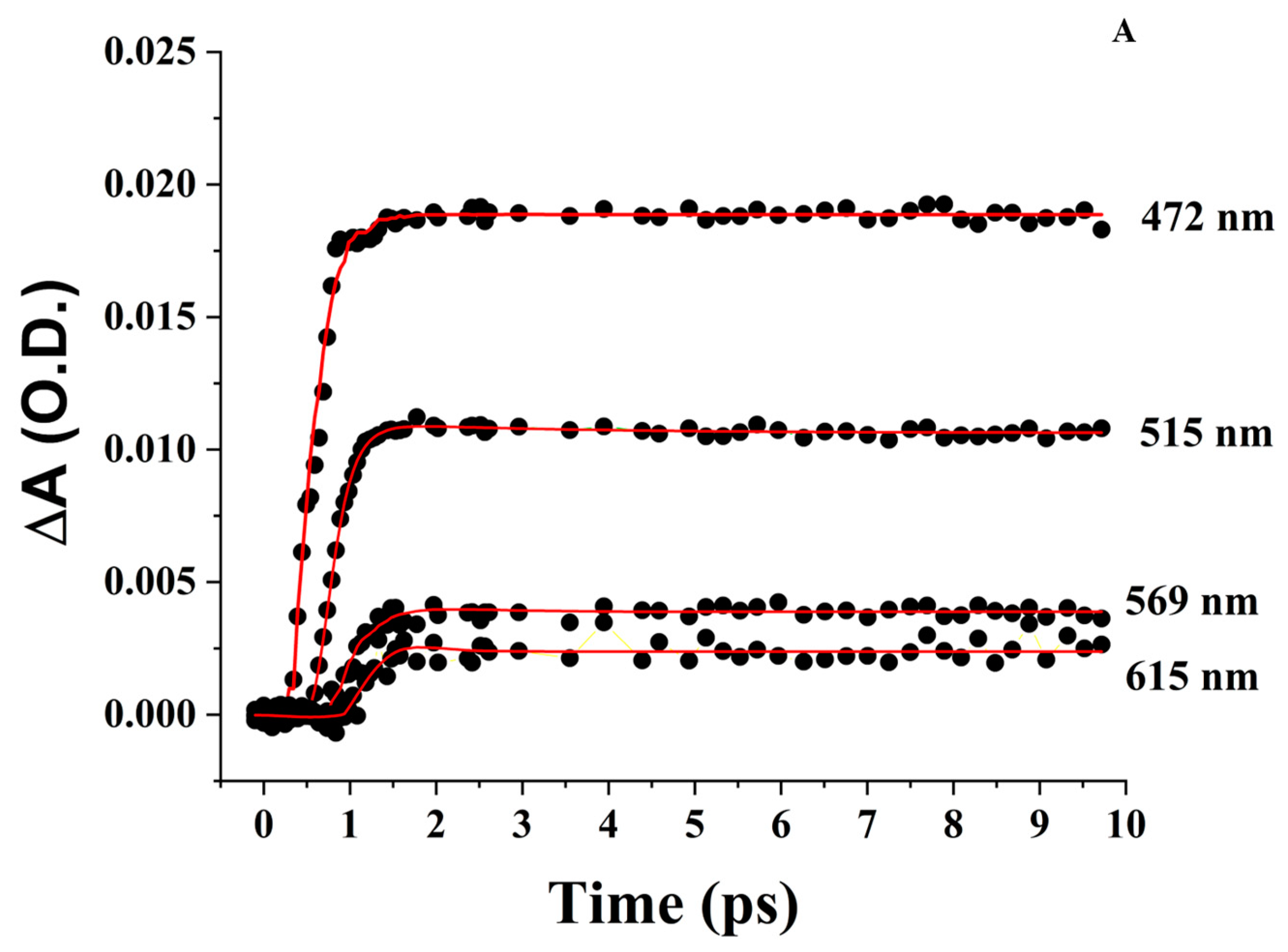
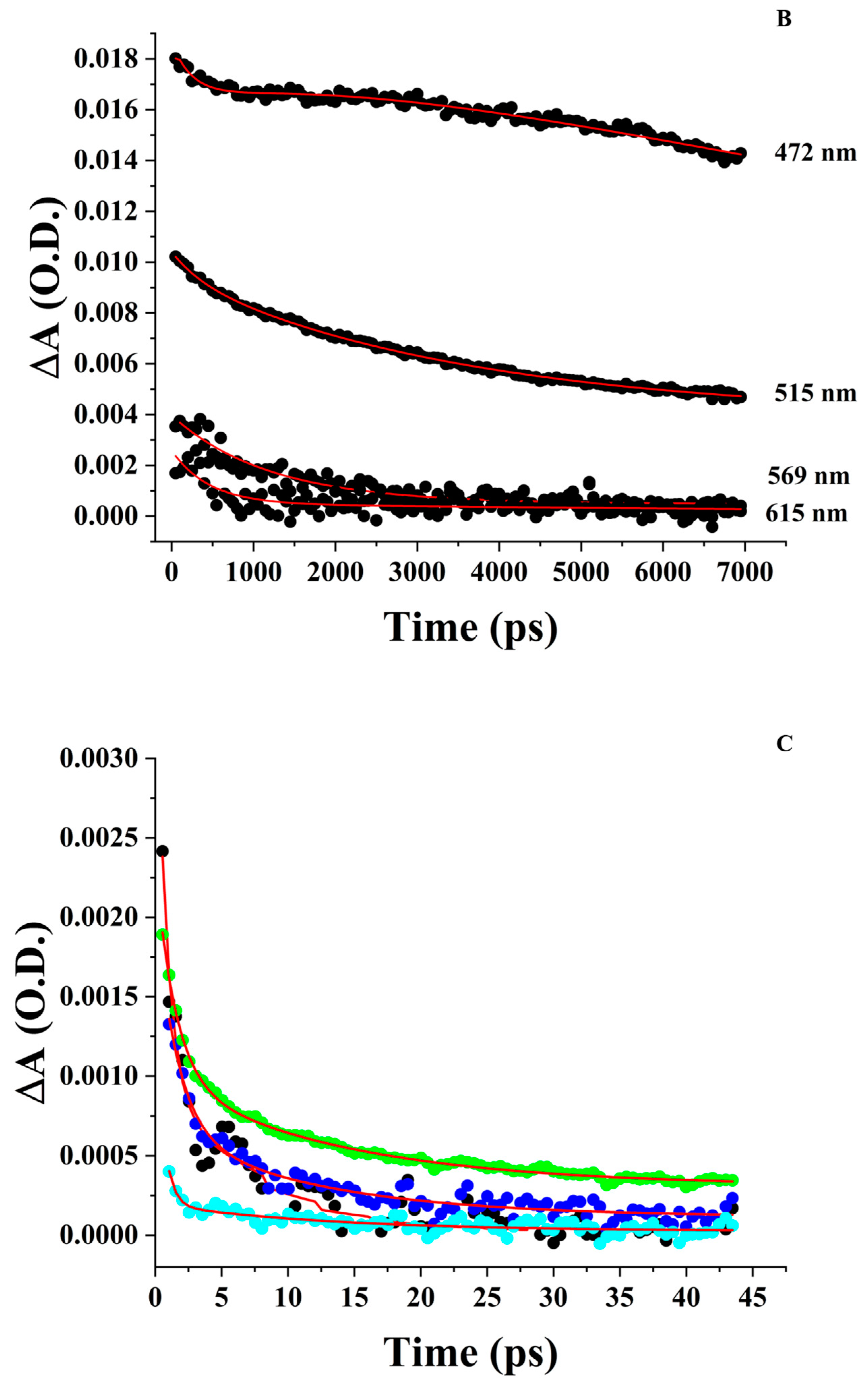
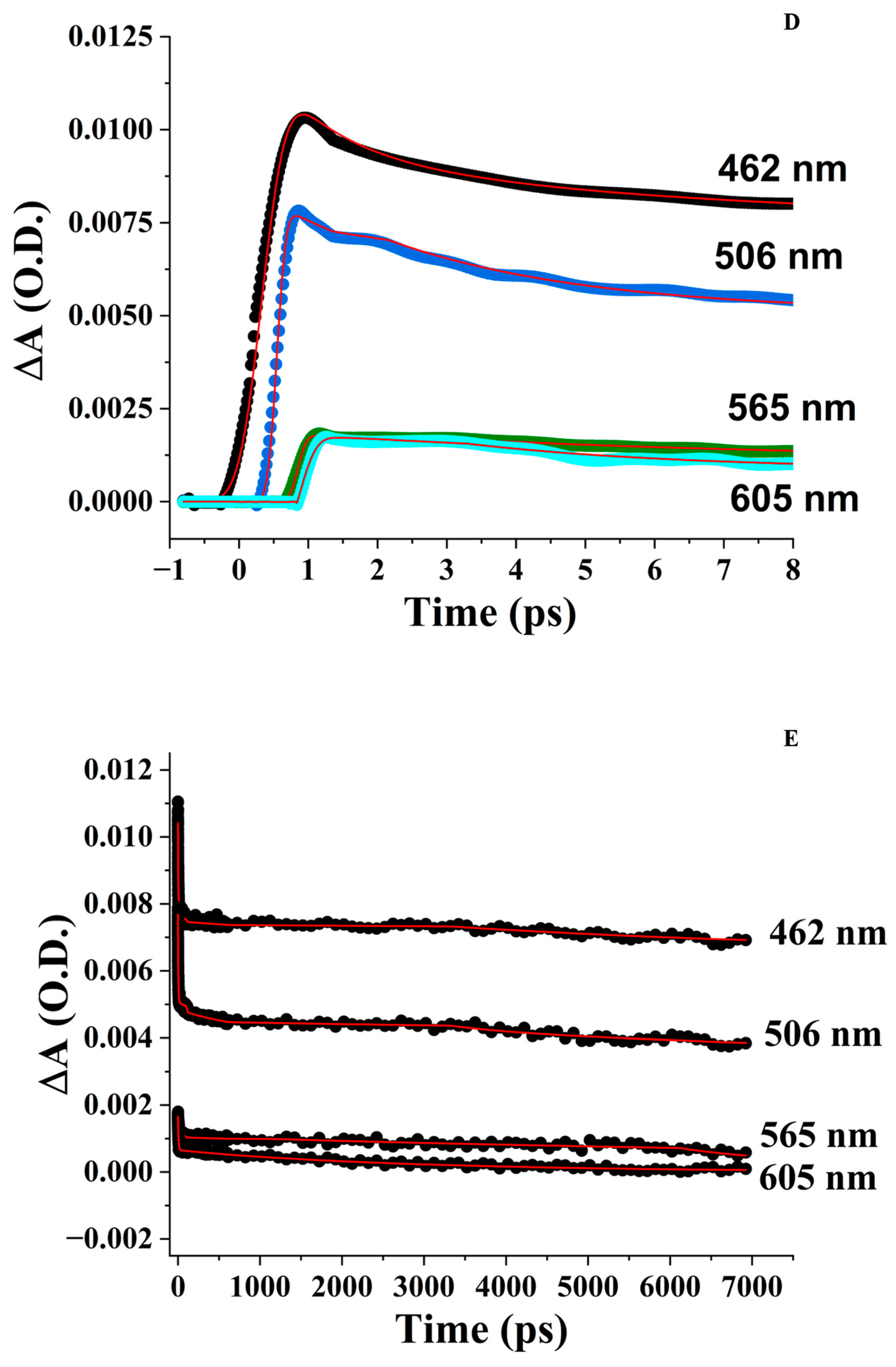
3.4. Femtosecond Transient Absorption Spectroscopy (FTAS)
3.4.1. ZnPPIX in DMSO
3.4.2. Aqueous ZnPPIX
3.4.3. ZnPPIX:BLG at pH 9
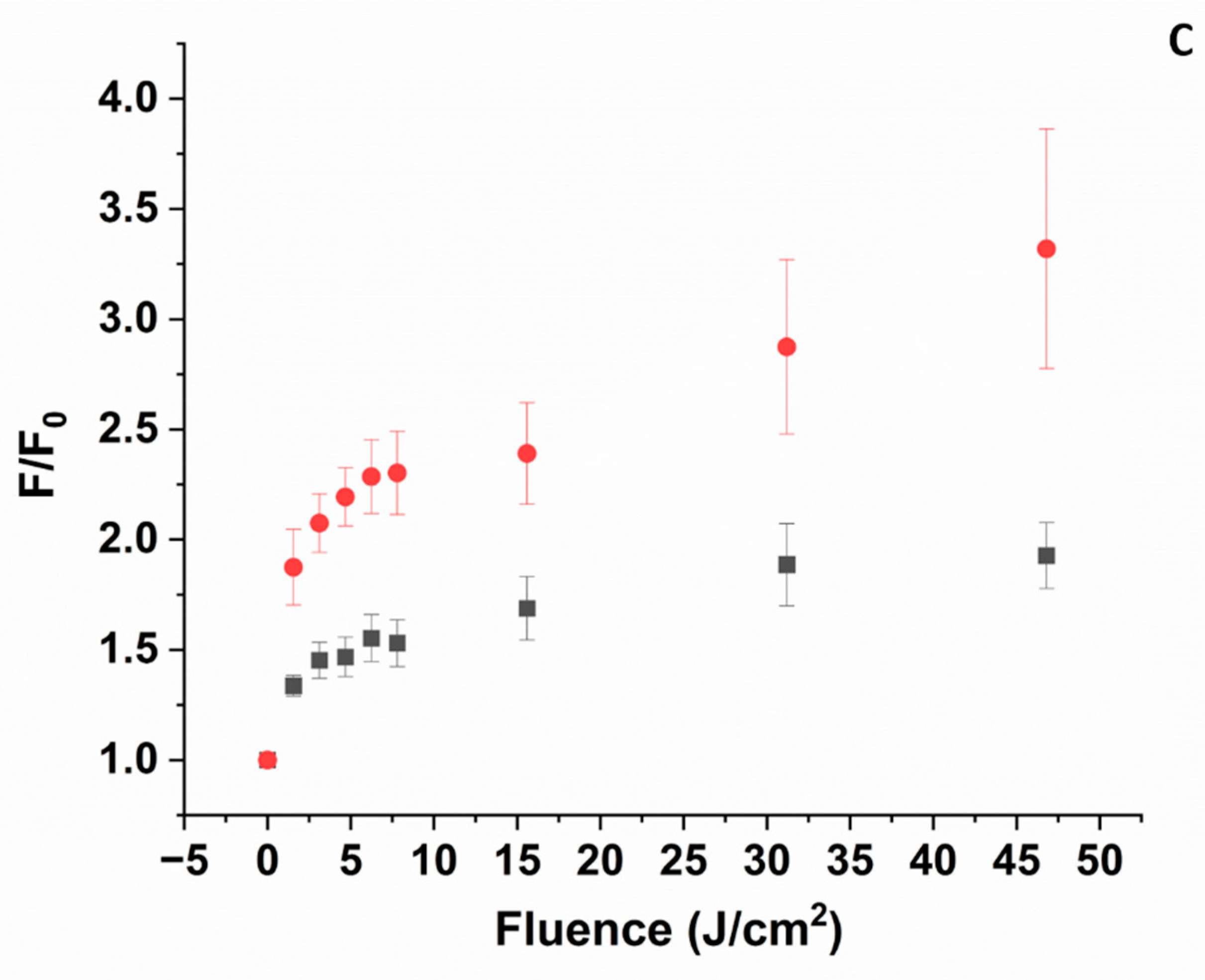
3.5. 1O2 Formation
3.5.1. pH 5
3.5.2. pH 9
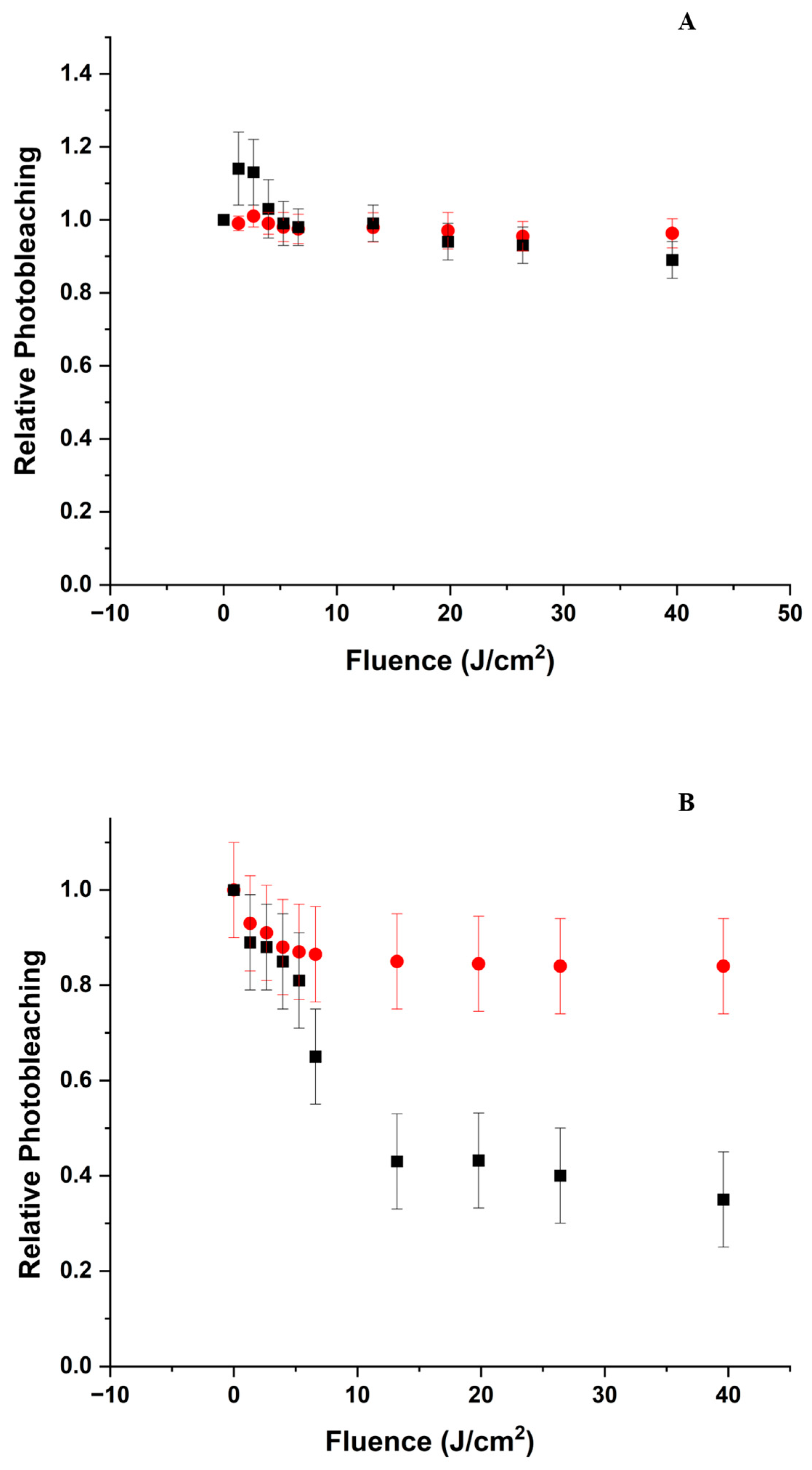
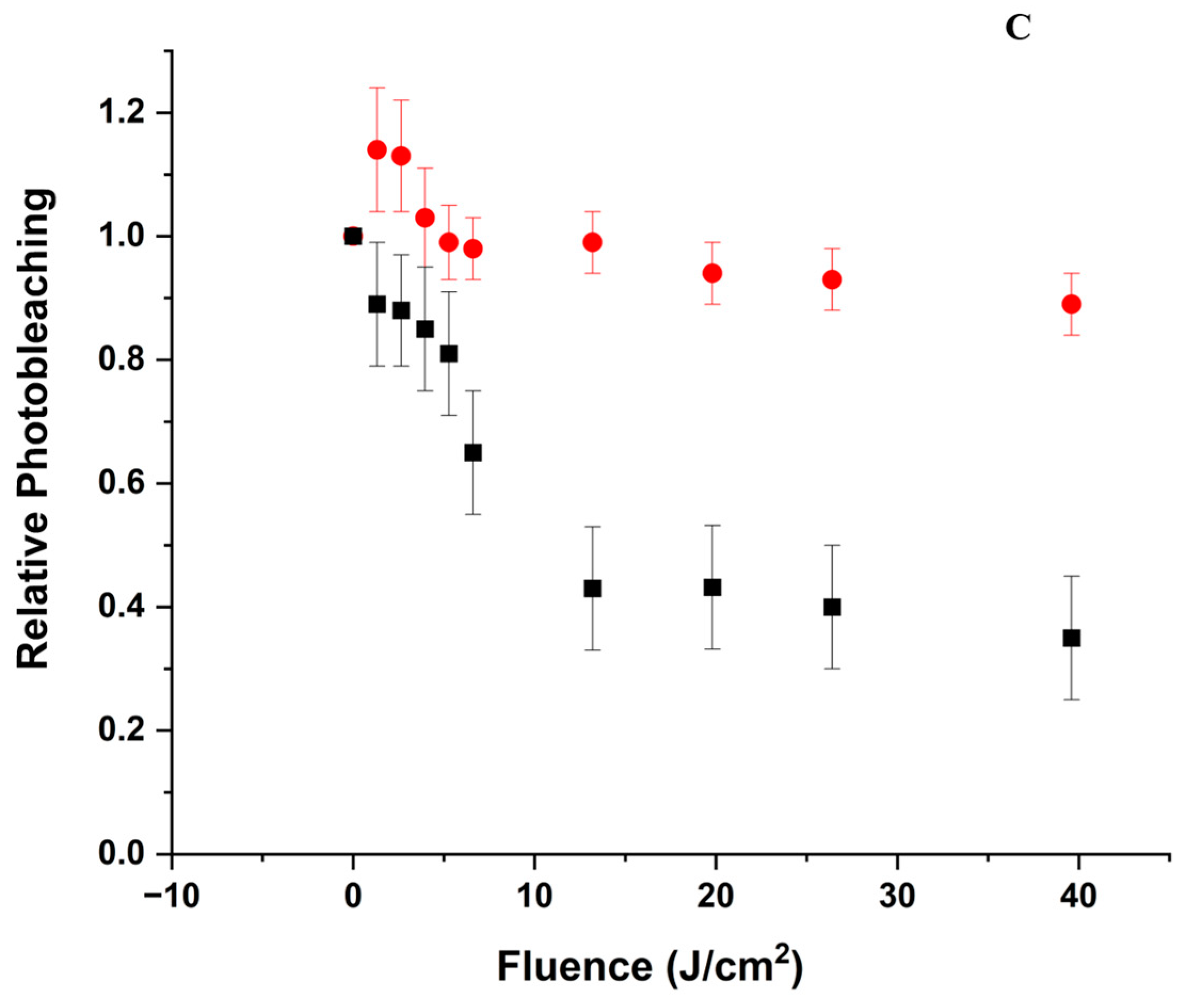
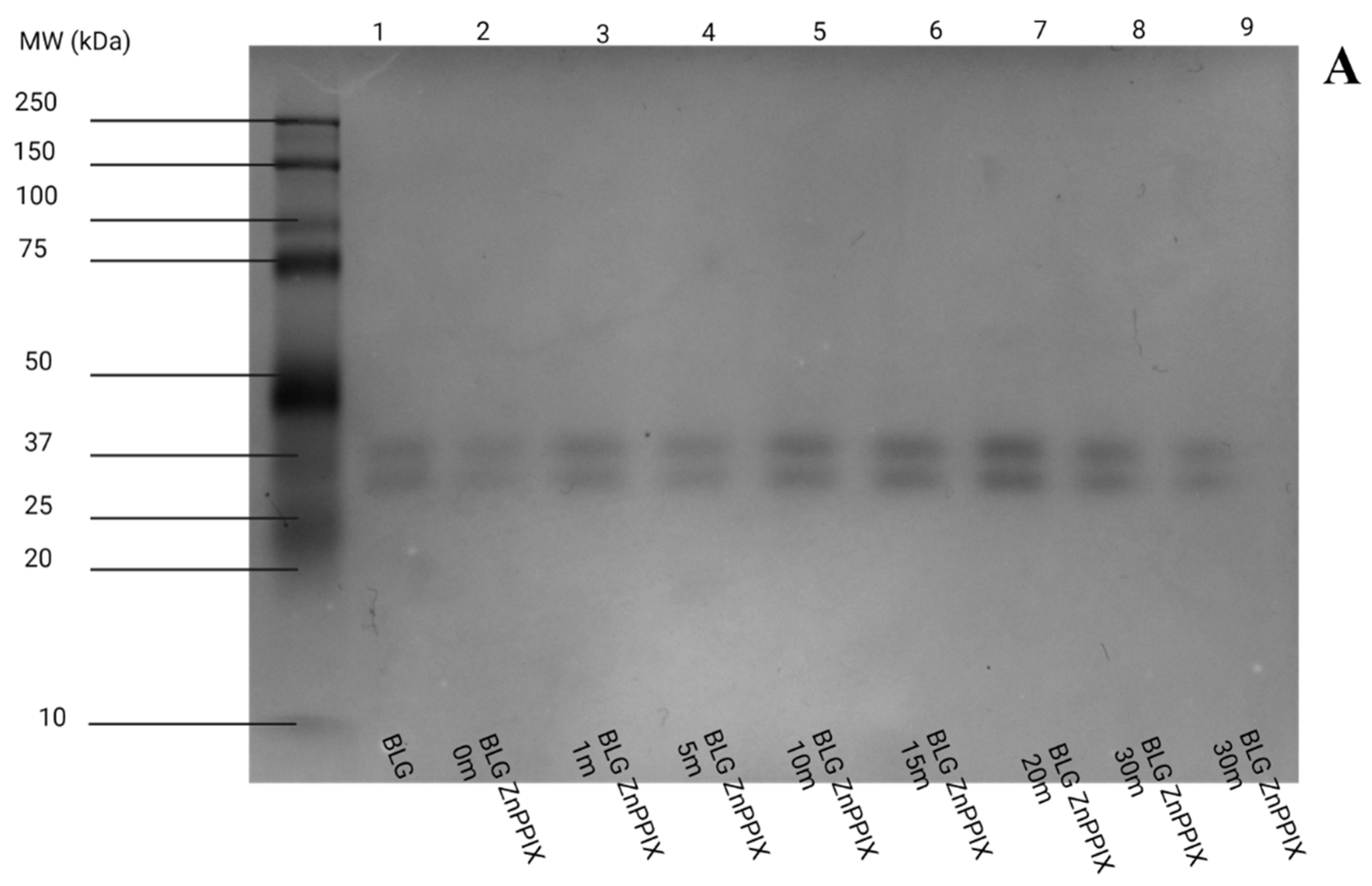
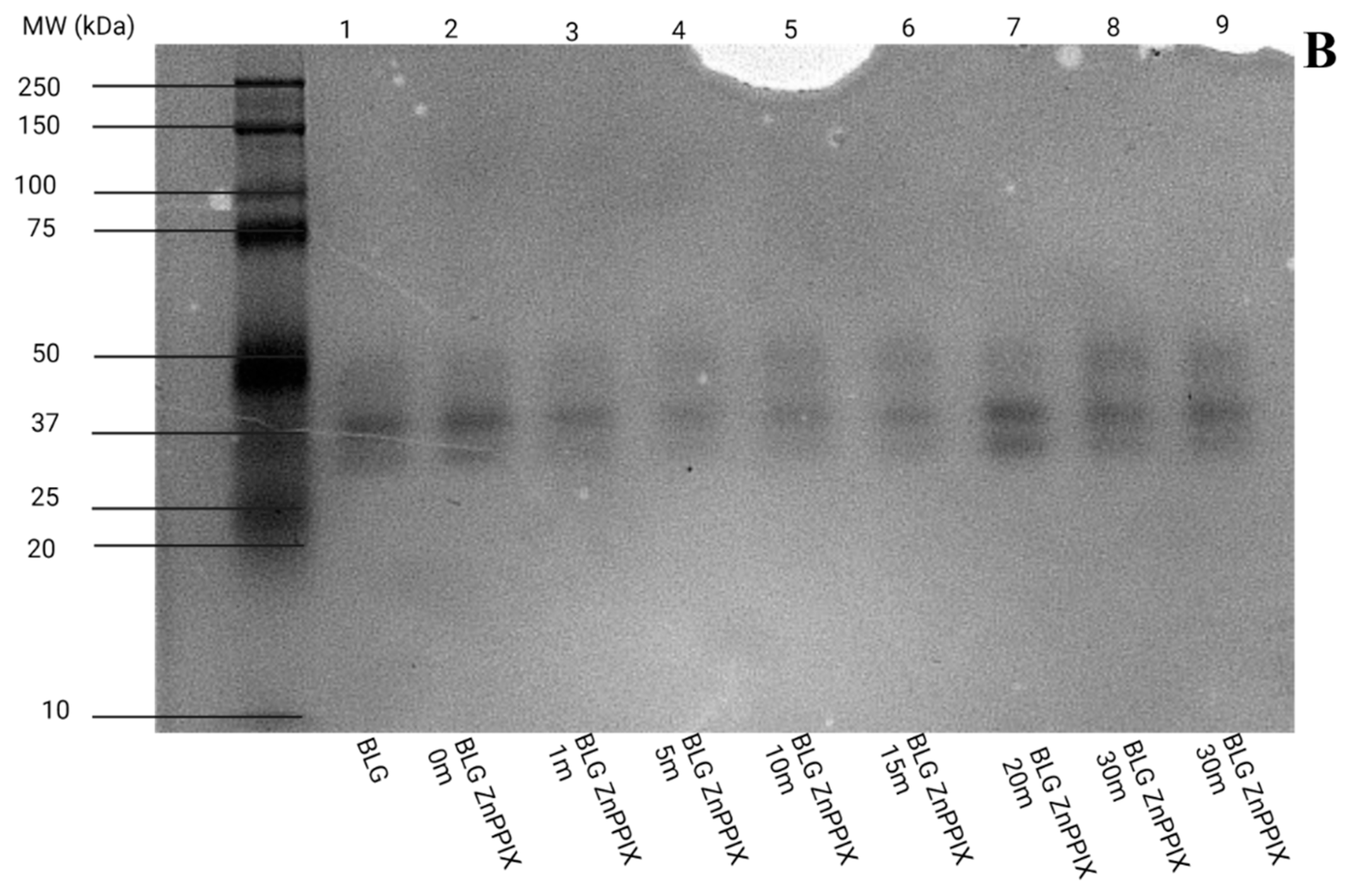
3.6. Gel Electrophoresis
3.6.1. pH 5
3.6.2. pH 9
4. Conclusions
- Light absorption by ZnPPIX prompts the formation of transient excited state species, represented mostly by the triplet state of the PS (*ZnPPIX3) and the cation radical (ZnPPIX●+). This conclusion is supported by the evidence shown in Figure 6 and Figure 7 and the related discussion in the corresponding section of the manuscript.
- The experiments confirm the formation of 1O2 carried out with SOSG (Figure 7). The SOSG data suggest that at pH 5, the formation of 1O2 is likely negligible, while at pH 9, it is more significant.
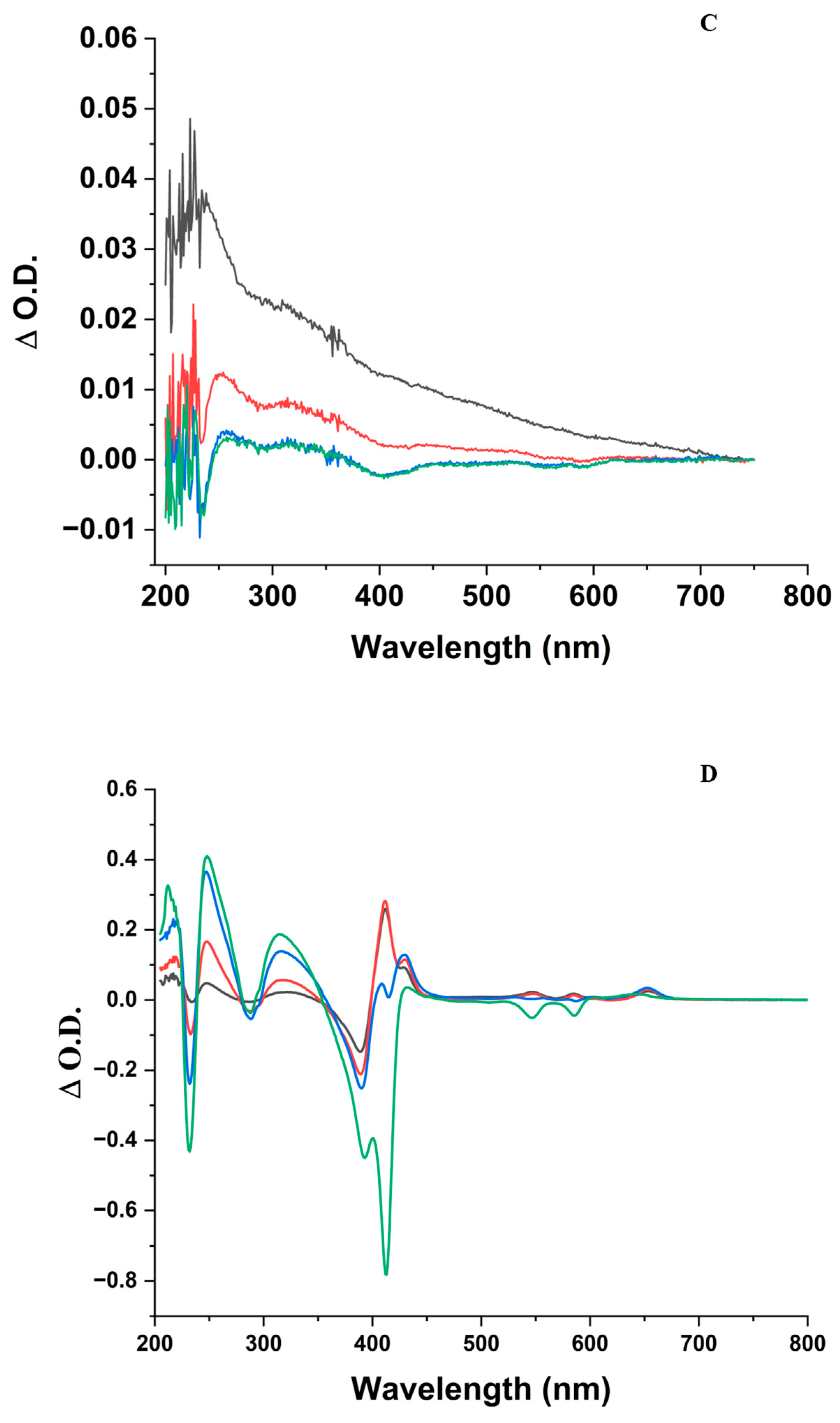
- In the ZnPPIX:BLG complex, the photosensitized species (triplet-sensitized 1O2 or ZnPPIX●+) prompt the oxidation of various protein residues. The differential absorption spectra (Figure 1C,D), as well as the decrease in fluorescence and fluorescence lifetime (Figure 3, Figure 4, and Figure S3), suggest the oxidation of at least the Trp residues and their conversion to NFK. Oxidation is likely to occur in other susceptible residues (Met, Cys, Tyr, and His) [21], but this could not be measured directly through our experiments.
- The oxidation of the amino acids does not appear to lead to significant aggregation or fragmentation of BLG (Figure 8), which implies that the protein’s backbone is not affected by the mechanisms.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bozovic, O.; Jankovic, B.; Hamm, P. Using azobenzene photocontrol to set proteins in motion. Nat. Rev. Chem. 2022, 6, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Azagarsamy, M.A.; Alge, D.L.; Radhakrishnan, S.J.; Tibbitt, M.W.; Anseth, K.S. Photocontrolled Nanoparticles for On-Demand Release of Proteins. Biomacromolecules 2012, 13, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzetti, S.; Sottini, S.; Viappiani, C.; Corrie, J.E. Acid-induced unfolding of myoglobin triggered by a laser pH jump method. Photochem. Photobiol. Sci. 2006, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Brieke, C.; Rohrbach, F.; Gottschalk, A.; Mayer, G.; Heckel, A. Light-controlled tools. Angew. Chem. Int. Ed. 2012, 51, 8446–8476. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.; Folini, M.; Prasmickaite, L.; Selbo, P.K.; Bonsted, A.; Engesaeter, B.Ø.; Zaffaroni, N.; Weyergang, A.; Dietze, A.; Maelandsmo, G.M.; et al. Photochemical internalization: A new tool for drug delivery. Curr. Pharm. Biotechnol. 2007, 8, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Calzaferri, G. Artificial photosynthesis. Top. Catal. 2010, 53, 130–140. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, Y.; Kaskel, S. Porphyrin-Based Metal–Organic Frameworks for Biomedical Applications. Angew. Chem. Int. Ed. 2021, 60, 5010–5035. [Google Scholar] [CrossRef]
- Lee, C.T., Jr.; Smith, K.A.; Hatton, T.A. Photocontrol of protein folding: The interaction of photosensitive surfactants with bovine serum albumin. Biochemistry 2005, 44, 524–536. [Google Scholar] [CrossRef]
- Möglich, A.; Moffat, K. Engineered photoreceptors as novel optogenetic tools. Photochem. Photobiol. Sci. 2010, 9, 1286–1300. [Google Scholar] [CrossRef]
- Kraiselburd, I.; Moyano, L.; Carrau, A.; Tano, J.; Orellano, E.G. Bacterial Photosensory Proteins and Their Role in Plant–pathogen Interactions. Photochem. Photobiol. 2017, 93, 666–674. [Google Scholar] [CrossRef]
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and Animal Rhodopsins: Structures, Functions, and Molecular Mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef]
- Losi, A. Flavin-based blue-light photosensors: A photobiophysics update. Photochem. Photobiol. 2007, 83, 1283–1300. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, Z.; Ma, C.; Zhu, Z. Photoswitchable Enzymatic Biofuel Cell Based on Fusion Protein with Natural Photoreceptor Vivid. ACS Appl. Bio Mater. 2022, 5, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.P. Photosynthesis. Essays Biochem. 2016, 60, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Palczewski, K. Chemistry and biology of vision. J. Biol. Chem. 2012, 287, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Bonnett, R.; Martinez, G. Photobleaching of the compounds of the 5,10,15,20-tetrakis(m-hydroxyphenyl)-porphyrin series (m-THPP, m-THPC and m-THPBC). Org. Lett. 2002, 4, 2013–2016. [Google Scholar] [CrossRef]
- Ericson, M.B.; Grapengiesser, S.; Gudmundson, F.; Wennberg, A.-M.; Larko, O.; Moan, J.; Rosen, A. A spectroscopic study of the photobleaching of protoporphyrin IX in solution. Lasers Surg. Med. 2003, 18, 56–62. [Google Scholar] [CrossRef]
- Finlay, J.C.; Conover, D.L.; Hull, E.L.; Foster, T.H. Porphyrin bleaching and PDT-induced spectral changes are irradiance dependent in ALA-sensitized normal rat skin in vivo. Photochem. Photobiol. 2001, 73, 54–63. [Google Scholar] [CrossRef]
- Jacobson, K.; Rajfur, Z.; Vitriol, E.; Hahn, K. Chromophore-assisted laser inactivation in cell biology. Trends Cell Biol. 2008, 18, 443–450. [Google Scholar] [CrossRef]
- Yan, P.; Xiong, Y.; Chen, B.; Negash, S.; Squier, T.C.; Mayer, M.U. Fluorophore-Assisted Light Inactivation of Calmodulin Involves Singlet-Oxygen Mediated Cross-Linking and Methionine Oxidation. Biochemistry 2006, 45, 4736–4748. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed]
- Bensasson, R.V.; Land, E.J.; Truscott, T.G. Excited States and Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Davies, M.J. Singlet oxygen-mediated damage to proteins and its consequences. Biochem. Biophys. Res. Commun. 2003, 305, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Reactive species formed on proteins exposed to singlet oxygen. Photochem. Photobiol. Sci. 2004, 3, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-F.; Chen, Y.; Yi, L.; Brantley, T.; Stanley, B.; Sosic, Z.; Zang, L. Discovery and Characterization of Histidine Oxidation Initiated Cross-links in an IgG1 Monoclonal Antibody. Anal. Chem. 2017, 89, 7915–7923. [Google Scholar] [CrossRef]
- Majiya, H.; Adeyemi, O.O.; Stonehouse, N.J.; Millner, P. Photodynamic inactivation of bacteriophage MS2: The A-protein is the target of virus inactivation. J. Photochem. Photobiol. B 2018, 178, 404–411. [Google Scholar] [CrossRef]
- Lee, A.J.; Ensign, A.A.; Krauss, T.D.; Bren, K.L. Zinc Porphyrin as a Donor for FRET in Zn(II)cytochrome c. J. Am. Chem. Soc. 2010, 132, 1752–1753. [Google Scholar] [CrossRef] [PubMed]
- Ndong Ntoutoume, G.M.A.; Granet, R.; Mbakidi, J.-P.; Constantin, E.; Bretin, L.; Léger, D.Y.; Liagre, B.; Chaleix, V.; Brégier, F.; Sol, V. Design and synthesis of zinc protoporphyrin IX-adamantane/cyclodextrin/cellulose nanocrystals complexes for anticancer photodynamic therapy. Bioorgan. Med. Chem. Lett. 2021, 41, 128024. [Google Scholar] [CrossRef]
- Tangar, A.; Derrien, V.; Lei, R.; Santiago Estevez, M.J.; Sebban, P.; Bernad, S.; Miksovska, J. Utility of fluorescent heme analogue ZnPPIX to monitor conformational heterogeneity in vertebrate hexa-coordinated globins. Metallomics 2019, 11, 906–913. [Google Scholar] [CrossRef]
- Shiraki, K.; Nishikawa, K.; Goto, Y. Trifluoroethanol-induced stabilization of the a-helical structure of b-lactoglobulin: Implication for non-hierirchical protein folding. J. Mol. Biol. 1995, 245, 180–194. [Google Scholar] [CrossRef]
- Barbiroli, A.; Iametti, S.; Bonomi, F. Beta-Lactoglobulin as a Model Food Protein: How to Promote, Prevent, and Exploit Its Unfolding Processes. Molecules 2022, 27, 1131. [Google Scholar] [CrossRef]
- Uhrinova, S.; Smith, M.H.; Jameson, G.B.; Uhrin, D.; Sawyer, L.; Barlow, P.N. Structural changes accompanying pH-induced dissociation of the β-lactoglobulin dimer. Biochemistry 2000, 39, 3565–3574. [Google Scholar] [CrossRef]
- Qin, B.Y.; Bewley, M.C.; Creamer, L.K.; Baker, H.M.; Baker, E.N.; Jameson, G.B. Structural basis of the tanford tyransition of bovine β-lactoglobulin. Biochemistry 1998, 37, 14014–14023. [Google Scholar] [CrossRef] [PubMed]
- Fenner, K.; Redgate, A.; Brancaleon, L. A 200 Nanoseconds All-Atom Simulation of the pH-dependent EF Loop Transition in Bovine β-Lactoglobulin. The Role of the Orientation of the E89 Side Chain. J. Biomol. Struct. Dyn. 2020, 40, 549–564. [Google Scholar] [CrossRef]
- Shafaei, Z.; Ghalandari, B.; Vaseghi, A.; Divsalar, A.; Haertlé, T.; Saboury, A.A.; Sawyer, L. β-Lactoglobulin: An efficient nanocarrier for advanced delivery systems. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Lohcharoenkal, W.; Wang, L.; Chen, Y.C.; Rojanasakul, Y. Protein nanoparticles as drug delivery carriers for cancer therapy. BioMed Res. Int. 2014, 2014, 180549. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Xu, R.; Wang, Q. Beta-lactoglobulin-based encapsulating systems as emerging bioavailability enhancers for nutraceuticals: A review. RSC Adv. 2015, 5, 35138–35154. [Google Scholar] [CrossRef]
- Rodríguez-Amigo, B.; Delcanale, P.; Rotger, G.; Juárez-Jiménez, J.; Abbruzzetti, S.; Summer, A.; Agut, M.; Luque, F.J.; Nonell, S.; Viappiani, C. The complex of hypericin with β-lactoglobulin has antimicrobial activity with perspective applications in dairy industry. J. Dairy Sci. 2015, 98, 89–94. [Google Scholar] [CrossRef]
- Belcher, J.; Sansone, S.; Fernandez, N.F.; Haskins, W.E.; Brancaleon, L. Photoinduced Unfolding of Beta-Lactoglobulin Mediated by a Water-Soluble Porphyrin. J. Phys. Chem. B 2009, 113, 6020–6030. [Google Scholar] [CrossRef]
- Fernandez, N.F.; Sansone, S.; Mazzini, A.; Brancaleon, L. Irradiation of the porphyrin causes unfolding of the protein in the Protoporphyrin IX/β-lactoglobulin non covalent complex. J. Phys. Chem. B 2008, 112, 7592–7600. [Google Scholar] [CrossRef]
- McMicken, B.; Thomas, R.J.; Brancaleon, L. Photoinduced partial unfolding of tubulin bound to meso-tetrakis(sulfonatophenyl) porphyrin leads to inhibition of microtubule formation in vitro. J. Biophotonics 2014, 7, 874–888. [Google Scholar] [CrossRef]
- Tian, F.; Johnson, K.; Lesar, A.E.; Moseley, H.; Ferguson, J.; Samuel, I.D.W.; Mazzini, A.; Brancaleon, L. The pH-Dependent Conformational Transition of β-Lactoglobulin Modulates the Binding of Protoporphyrin IX. Biochim. Biophys. Acta 2005, 1760, 38–46. [Google Scholar] [CrossRef]
- Castillo, O.; Mancillas, J.; Hughes, W.; Brancaleon, L. Characterization of the Interaction of Metal-Protoporphyrins Photosensitizers with β-Lactoglobulin. Biophys. Chem. 2023, 292, 106918. [Google Scholar] [CrossRef]
- Benavides, B.S.; Acharya, R.; Clark, E.R.; Basak, P.; Maroney, M.J.; Nocek, J.M.; Schanze, K.S.; Kurtz, D.M., Jr. Structural, Photophysical, and Photochemical Characterization of Zinc Protoporphyrin IX in a Dimeric Variant of an Iron Storage Protein: Insights into the Mechanism of Photosensitized H(2) Generation. J. Phys. Chem. B 2019, 123, 6740–6749. [Google Scholar] [CrossRef]
- Johnson, C.N.; Gorbet, G.E.; Ramsower, H.; Urquidi, J.; Brancaleon, L.; Demeler, B. Multi-wavelength analytical ultracentrifugation of human serum albumin complexed with porphyrin. Eur. Biophys. J. 2018, 47, 789–797. [Google Scholar] [CrossRef]
- Swinehart, D.F. The Beer-Lambert Law. J. Chem. Educ. 1962, 39, 333. [Google Scholar] [CrossRef]
- Collini, M.; D’Alfonso, L.; Baldini, G. New insight on β-lactoglobulin binding sites by 1-anilinonaphtalene-8-sulfonate fluorescence decay. Prot. Sci. 2000, 9, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. On spectral relaxation in proteins. Photochem. Photobiol. 2000, 72, 421–437. [Google Scholar] [CrossRef]
- Albani, J.R.; Vogelaer, J.; Bretesche, L.; Kmiecik, D. Tryptophan 19 residue is the origin of bovine β-lactoglobulin fluorescence. J. Pharm. Biomed. Anal. 2014, 91, 144–150. [Google Scholar] [CrossRef]
- Hu, J.; Allen, R.; Rozinek, S.; Brancaleon, L. Experimental and computational characterization of photosensitized conformational effects mediated by protoporphyrin ligands on human serum albumin. Photochem. Photobiol. Sci. 2017, 16, 694–710. [Google Scholar] [CrossRef]
- van de Weert, M.; Stella, L. Fluorescence quenching and ligand binding: A critical discussion of a popular methodology. J. Mol. Struct. 2011, 998, 144–150. [Google Scholar] [CrossRef]
- Kasha, M. Characterization of Electronic Transitions in Complex Molecules. Discuss. Faraday Soc. 1950, 9, 14–19. [Google Scholar] [CrossRef]
- Harvey, B.J.; Bell, E.; Brancaleon, L. A Tryptophan Rotamer Located in a Polar Environment Probes pH-Dependent Conformational Changes in Bovine beta-Lactoglobulin A. J. Phys. Chem. B 2007, 111, 2610–2620. [Google Scholar] [CrossRef] [PubMed]
- Durbin, J.; Watson, G.S. Testing for serial correlation in least squares regression. II. Biometrika 1951, 38, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Brancaleon, L.; Moseley, H. Laser and Non-Laser Light Sources for Photodynamic Therapy. Lasers Med. Sci. 2002, 17, 173–186. [Google Scholar] [CrossRef]
- Wamser, C.C.; Ghosh, A. The Hyperporphyrin Concept: A Contemporary Perspective. JACS Au 2022, 2, 1543–1560. [Google Scholar] [CrossRef]
- Chang, C.-W.; Luo, L.; Chou, C.-K.; Lo, C.-F.; Lin, C.-Y.; Hung, C.-S.; Lee, Y.-P.; Diau, E.W.-G. Femtosecond Transient Absorption of Zinc Porphyrins with Oligo(phenylethylnyl) Linkers in Solution and on TiO2 Films. J. Phys. Chem. C 2009, 113, 11524–11531. [Google Scholar] [CrossRef]
- Schmidt, R. Photosensitized Generation of Singlet Oxygen. Photochem. Photobiol. 2006, 82, 1161–1177. [Google Scholar] [CrossRef]
- Jones, M.; Talfournier, F.; Bobrov, A.; Grossmann, J.G.; Vekshin, N.; Sutcliffe, M.J.; Scrutton, N.S. Electron transfer and conformational change in complexes of trimethylamine dehydrogenase and electron transferring flavoprotein. J. Biol. Chem. 2002, 277, 8457–8465. [Google Scholar] [CrossRef]
- Jiang, S.; Carroll, L.; Mariotti, M.; Hägglund, P.; Davies, M.J. Formation of protein cross-links by singlet oxygen-mediated disulfide oxidation. Redox Biol. 2021, 41, 101874. [Google Scholar] [CrossRef]
- Prinsze, C.; Dubbelman, T.M.A.R.; Van Steveninck, J. Protein damage induced by small amounts of photodynamically generated singlet oxygen or hydroxyl radicals. Biochim. Biophys. Acta 1990, 1038, 152–157. [Google Scholar] [CrossRef]
- O’Connor, S.P.; Powell, S.M.; Rickman, J.M.; Pope, N.J.; Noojin, G.D.; Scully, M.O.; Denton, M.L.; Yakovlev, V.V. Transient absorption spectroscopy to explore cellular pathways to photobiomodulation. J. Photochem. Photobiol. B Biol. 2021, 222, 112271. [Google Scholar] [CrossRef]
- Yakovlev, V.V.; Kohler, B.; Wilson, K.R. Broadly tunable 30-fs pulses produced by optical parametric amplification. Opt. Lett. 1994, 19, 2000–2002. [Google Scholar] [CrossRef]
- Johnston, T.F. Beam propagation (M2) measurement made as easy as it gets: The four-cuts method. Appl. Opt. 1998, 37, 4840–4850. [Google Scholar] [CrossRef]
- Mazur, L.M.; Roland, T.; Leroy-Lhez, S.; Sol, V.; Samoc, M.; Samuel, I.; Matczyszyn, K. Efficient Singlet Oxygen Photogeneration by Zinc Porphyrin Dimers upon One- and Two-Photon Excitation. J. Phys. Chem. B 2019, 123, 4271–4277. [Google Scholar] [CrossRef]
- Kim, S.; Fujitsuka, M.; Majima, T. Photochemistry of Singlet Oxygen Sensor Green. J. Phys. Chem. B 2013, 117, 13985–13992. [Google Scholar] [CrossRef]
- Gollmer, A.; Arnbjerg, J.; Blaikie, F.H.; Pedersen, B.W.; Breitenbach, T.; Daasbjerg, K.; Glasius, M.; Ogilby, P.R. Singlet oxygen sensor green: Photochemical behaviour in solution and in mammalian cell. Photochem. Photobiol. 2011, 87, 671–679. [Google Scholar] [CrossRef]
- Knoblauch, R.; Moskowitz, J.; Hawkins, E.; Geddes, C.D. Fluorophore-Induced Plasmonic Current: Generation-Based Detection of Singlet Oxygen. ACS Sens. 2020, 5, 1223–1229. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Pospíšil, P. Singlet oxygen imaging using fluorescent probe Singlet Oxygen Sensor Green in photosynthetic organisms. Sci. Rep. 2018, 8, 13685. [Google Scholar] [CrossRef]
- Liu, H.; Carter, P.J.H.; Laan, A.C.; Eelkema, R.; Denkova, A.G. Singlet Oxygen Sensor Green is not a Suitable Probe for 1O2 in the Presence of Ionizing Radiation. Sci. Rep. 2019, 9, 8393. [Google Scholar] [CrossRef]
- Schägger, H.; Cramer, W.A.; von Jagow, G. Analysis of molecular masses and oligomeric states of protein complexes by blue native electrophoresis and isolation of membrane protein complexes by two-dimensional native electrophoresis. Anal. Biochem. 1994, 217, 220–230. [Google Scholar] [CrossRef]
- Jarzecki, A.A.; Spiro, T.G. Porphyrin distorsion from resonance raman intensities of out-of-plane modes: Computation and modeling of N-methylmesoporphyrin, a ferrochelatase transition state analogue. J. Phys. Chem. A 2005, 109, 421–430. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Prot. Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef]
- Hu, J.; Hernandez Soraiz, E.; Johnson, C.N.; Demeler, B.; Brancaleon, L. Novel combinations of experimental and computational analysis tested on the binding of metalloprotoporphyrins to albumin. Int. J. Biol. Macromol. 2019, 134, 445–457. [Google Scholar] [CrossRef]
- Monsù Scolaro, L.; Castriciano, M.; Romeo, A.; Patanè, S.; Cefali, E.; Allegrini, M. Aggregation behavior of protoporphyrin IX in aqueous solutions: Clear evidence of vescicle formation. J. Phys. Chem. B 2002, 106, 2453–2459. [Google Scholar] [CrossRef]
- Sandberg, S.; Romslo, I. Porphyrin-induced photodamage at the cellular and the subcellular level as related to the solubility of the porphyrin. Clin. Chim. Acta 1981, 109, 193–201. [Google Scholar] [CrossRef]
- Dreaden, T.M.; Chen, J.; Rexroth, S.; Barry, B.A. N-formylkynurenine as a marker of high light stress in photosynthesis. J. Biol. Chem. 2011, 286, 22632–22641. [Google Scholar] [CrossRef]
- Afaghi, P.; Lapolla, M.A.; Ghandi, K. Determining the degree of denaturation of bovine serum albumin using a new UV analysis technique. Chem. Rep. 2021, 3, 173–176. [Google Scholar] [CrossRef]
- Fukunaga, Y.; Katsuragi, Y.; Izumi, T.; Sakiyama, F. Fluorescence characteristics of kynurenine and N′-formylkynurenine. Their use as reporters of the environment of tryptophan 62 in hen egg-white lysozyme. J. Biochem. 1982, 92, 129–141. [Google Scholar] [CrossRef]
- Gracanin, M.; Hawkins, C.L.; Pattison, D.I.; Davies, M.J. Singlet-oxygen-mediated amino acid and protein oxidation: Formation of tryptophan peroxides and decomposition products. Free Radic. Biol. Med. 2009, 47, 92–102. [Google Scholar] [CrossRef]
- Zhdanova, N.G.; Shirshin, E.A.; Maksimov, E.G.; Panchishin, I.M.; Saletsky, A.M.; Fadeev, V.V. Tyrosine fluorescence probing of the surfactant-induced conformational changes of albumin. Photochem. Photobiol. Sci. 2015, 14, 897–908. [Google Scholar] [CrossRef]
- Han, Q.; Robinson, H.; Li, J. Biochemical identification and crystal structure of kynurenine formamidase from Drosophila melanogaster. Biochem. J. 2012, 446, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Sherin, P.S.; Grilj, J.; Kopylova, L.V.; Yanshole, V.V.; Tsentalovich, Y.P.; Vauthey, E. Photophysics and photochemistry of the UV filter kynurenine covalently attached to amino acids and to a model protein. J. Phys. Chem. B 2010, 114, 11909–11919. [Google Scholar] [CrossRef] [PubMed]
- Taulier, N.; Chalikian, T.V. Characterization of pH-induced transitions of beta-lactoglobulin: Ultrasonic, densimetric, and spectroscopic studies. J. Mol. Biol. 2001, 314, 873–889. [Google Scholar] [CrossRef] [PubMed]
- Rozinek, S.E.; Thomas, R.J.; Brancaleon, L. Biophysical Characterization of the Interaction of Human Albumin with a Cationic Porphyrin. Biochem. Biophys. Rep. 2016, 7, 295–302. [Google Scholar] [CrossRef]
- Khrenova, M.; Topol, I.; Collins, J.; Nemukhin, A. Estimating Orientation Factors in the FRET Theory of Fluorescent Proteins: The TagRFP-KFP Pair and Beyond. Biophys. J. 2015, 108, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Piston, D.W.; Kremers, G.J. Fluorescent protein FRET: The good, the bad and the ugly. Trends Biochem. Sci. 2007, 32, 407–414. [Google Scholar] [CrossRef]
- Grabolle, M.; Pauli, J.; Brehm, R.; Resch-Genger, U. Structural control of dye–protein binding, aggregation and hydrophilicity in a series of asymmetric cyanines. Dye. Pigment. 2014, 103, 118–126. [Google Scholar] [CrossRef]
- Maitra, D.; Pinsky, B.M.; Soherawardy, A.; Zheng, H.; Banerjee, R.; Omary, M.B. Protoporphyrin-IX nanostructures modulate their protein aggregation ability via differential oxidation and protein binding. bioRxiv 2021. [Google Scholar] [CrossRef]
- Iyer, J.K.; Shi, L.; Shankar, A.H.; Sullivan, D.J., Jr. Zinc protoporphyrin IX binds heme crystals to inhibit the process of crystallization in Plasmodium falciparum. Mol. Med. 2003, 9, 175–182. [Google Scholar] [CrossRef]
- Gouterman, M. Spectra of porphyrins. J. Mol. Spectrosc. 1961, 6, 138–163. [Google Scholar] [CrossRef]
- Andrés, G.O.; Cabrerizo, F.M.; Martínez-Junza, V.; Braslavsky, S.E. A Large Entropic Term Due to Water Rearrangement is Concomitant with the Photoproduction of Anionic Free-Base Porphyrin Triplet States in Aqueous Solutions. Photochem. Photobiol. 2007, 83, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Finikova, O.S.; Chen, P.; Ou, Z.; Kadish, K.M.; Vinogradov, S.A. Dynamic quenching of porphyrin triplet-states by two-photon absorpting dyes: Towards two-photon-enhanced oxygen nanosensors. J. Photochem. Photobiol. A 2008, 198, 75–84. [Google Scholar] [CrossRef]
- Togashi, D.M.; Costa, S.M.B. Absorption, fluorescence and transient triplet-triplet absorption spectra of zinc tetramethylpyridylporphyrin in reverse micellesand microemulsions of aerosol OT-(AOT). Phys. Chem. Chem. Phys. 2000, 2, 5437–5444. [Google Scholar] [CrossRef]
- Ballard, S.G.; Mauzerall, D. Photo-initiated ion formation from octaethyl-porphyrin and its zinc chelate as a model for electron transfer in reaction centers. Biophys. J. 1978, 24, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Mukherjee, D.; De, S.; Kathiravan, A. Probing the Highly Efficient Electron Transfer Dynamics between Zinc Protoporphyrin IX and Sodium Titanate Nanosheets. J. Phys. Chem. A 2016, 120, 7121–7129. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.L.; Kuciauskas, D. Charge transfer enhances two-photon absorption in transition metal porphyrins. J. Am. Chem. Soc. 2006, 128, 3902–3903. [Google Scholar] [CrossRef]
- Laia, C.A.T.; Costa, S.M.B.; Ferreira, L.F.V. Electron-transfer mechanism of the triplet state quenching of aluminum tetrasulfonated phthalocyanine by cytochrome C. Biophys. Chem. 2006, 122, 143–155. [Google Scholar] [CrossRef]
- Kumar, P.H.; Venkatesh, Y.; Siva, D.; Ramakrishna, B.; Bangal, P.R. Ultrafast Relaxation Dynamics of 5,10,15,20-meso-Tetrakis Pentafluorophenyl Porphyrin Studied by Fluorescence Up-Conversion and Transient Absorption Spectroscopy. J. Phys. Chem. A 2015, 119, 1267–1278. [Google Scholar] [CrossRef]
- Shimizu, D.; Osuka, A. Porphyrinoids as a platform of stable radicals. Chem. Sci. 2018, 9, 1408–1423. [Google Scholar] [CrossRef]
- Hashimoto, T.; Choe, Y.-K.; Nakano, H.; Hirao, K. Theoretical Study of the Q and B Bands of Free-Base, Magnesium, and Zinc Porphyrins, and Their Derivatives. J. Phys. Chem. A 1999, 103, 1894–1904. [Google Scholar] [CrossRef]
- Feitelson, J.; Barboy, N. Triplet-state reactions of zinc protoporphyrins. J. Phys. Chem. 1986, 90, 271–274. [Google Scholar] [CrossRef]
- Nonell, S.B.; Braslavsky, S.E. Time-resolved singlet oxygen detection. Methods Enzymol. 2000, 319, 37–49. [Google Scholar] [PubMed]
- Montfort, C.V.; Sharov, V.S.; Metzger, S.; Schöneich, C.; Sies, H.; Klotz, L.-O. Singlet oxygen inactivates protein tyrosine phosphatase-1B by oxidation of the active site cysteine. Biol. Chem. 2006, 387, 1399–1404. [Google Scholar] [CrossRef]
- Ronsein, G.E.; Oliveira, M.C.B.; Miyamoto, S.; Medeiros, M.H.G.; Di Mascio, P. Tryptophan Oxidation by Singlet Molecular Oxygen [O2 (1Δg)]: Mechanistic Studies Using 18O-Labeled Hydroperoxides, Mass Spectrometry, and Light Emission Measurements. Chem. Res. Toxicol. 2008, 21, 1271–1283. [Google Scholar] [CrossRef]
- Boreen, A.L.; Edhlund, B.L.; Cotner, J.B.; McNeill, K. Indirect photodegradation of dissolved free amino acids: The contribution of singlet oxygen and differential reactivity of DOM from various sources. Environ. Sci. Technol. 2008, 42, 5492–5498. [Google Scholar] [CrossRef]
- Ding, Y.; Zhao, Z.; Matysik, J.; Gärtner, W.; Losi, A. Mapping the role of aromatic amino acids within a blue-light sensing LOV domain. Phys. Chem. Chem. Phys. 2021, 23, 16767–16775. [Google Scholar] [CrossRef]
- Jensen, R.L.; Arnbjerg, J.; Ogilby, P.R. Reaction of Singlet Oxygen with Tryptophan in Proteins: A Pronounced Effect of the Local Environment on the Reaction Rate. J. Am. Chem. Soc. 2012, 134, 9820–9826. [Google Scholar] [CrossRef]
- Gokhale, N.H.; Bradford, S.; Cowan, J.A. Catalytic inactivation of human carbonic anhydrase I by a metallopeptide-sulfonamide conjugate is mediated by oxydation of active site residues. J. Am. Chem. Soc. 2008, 130, 2388–2389. [Google Scholar] [CrossRef]
- Ragàs, X.; Jiménez-Banzo, A.; Sánchez-García, D.; Batllori, X.; Nonell, S. Singlet oxygen photosensitisation by the fluorescent probe Singlet Oxygen Sensor Green®. Chem. Commun. 2009, 2920–2922. [Google Scholar] [CrossRef]
- Pattison, D.I.; Rahmanto, A.S.; Davies, M.J. Photo-oxidation of proteins. Photochem. Photobiol. Sci. 2012, 11, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Bubb, W.A.; Hawkins, C.L.; Davies, M.J. Singlet oxygen-mediated protein oxidation: Evidence for the formation of reactive side chain peroxides on tyrosine residues. Photochem. Photobiol. 2002, 76, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Leinisch, F.; Mariotti, M.; Rykaer, M.; Lopez-Alarcon, C.; Hägglund, P.; Davies, M.J. Peroxyl radical- and photo-oxidation of glucose 6-phosphate dehydrogenase generates cross-links and functional changes via oxidation of tyrosine and tryptophan residues. Free Radic. Biol. Med. 2017, 112, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Kochevar, I.E.; Redmond, R.W. Photosensitized production of singlet oxygen. Methods Enzymol. 2000, 319, 20–28. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||||
| pH 5 | 0 (J) | 48 (J) | 261 (J) | 864 (J) | |
| 0.24 ± 0.05 | 0.28 ± 0.05 | 0.38 ± 0.05 | 0.47 ± 0.04 | ||
| 0.76 ± 0.02 | 0.72 ± 0.02 | 0.62 ± 0.02 | 0.53 ± 0.02 | ||
| 0.79 ± 0.02 | 0.72 ± 0.02 | 0.74 ± 0.01 | 0.79 ± 0.01 | ||
| 1.73 ± 0.04 | 1.65 ± 0.04 | 1.64 ± 0.04 | 1.76 ± 0.06 | ||
| <τ> | 1.40 ± 0.12 | 1.36 ± 0.11 | 1.30 ± 0.10 | 1.30 ± 0.10 | |
| (B) | |||||
| pH 9 | 0 (J) | 24 (J) | 72 (J) | 288 (J) | 884 (J) |
| 0.24 ± 0.05 | 0.18 ± 0.05 | 0.16 ± 0.06 | 0.20 ± 0.07 | 0.54 ± 0.11 | |
| 0.60 ± 0.03 | 0.57 ± 0.03 | 0.55 ± 0.03 | 0.48 ± 0.03 | 0.36 ± 0.02 | |
| 0.15 ± 0.05 | 0.24 ± 0.05 | 0.29 ± 0.06 | 0.31 ± 0.06 | 0.10 ± 0.02 | |
| 0.74 ± 0.02 | 0.64 ± 0.02 | 0.56 ± 0.02 | 0.38 ± 0.08 | 0.19 ± 0.02 | |
| 1.95 ± 0.02 | 1.89 ± 0.02 | 1.81 ± 0.08 | 1.71 ± 0.08 | 1.40 ± 0.08 | |
| 5.15 ± 0.04 | 5.15 ± 0.03 | 5.15 ± 0.04 | 5.15 ± 0.04 | 5.16 ± 0.04 | |
| <τ> | 2.12 ± 0.42 | 2.42 ± 0.41 | 2.56 ± 0.44 | 2.49 ± 0.42 | 1.12 ± 0.48 |
| ZnPPIX in DMSO | 471–474 nm | 515 nm | 569 nm | 615 nm | 550 nm |
| 0.22 ± 0.07 ↑ | 0.20 ± 0.02 ↑ | 0.22 ± 0.09 ↑ | 0.28 ± 0.07 ↑ | 0.41 ± 0.06 ↓ | |
| 0.68 ± 0.22 | 0.83 ± 0.28 | 0.66 ± 0.25 | 0.34 ± 0.17 | ||
| 253 ± 25 ↓ | 1.47 ± 0.21 ↓ | 1.05 ± 0.33 ↑ | 0.31 ± 0.06 ↓ | 1782 ± 122 ↑ | |
| 0.01± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.43 ± 0.22 | ||
| 5243 ± 570 ↑ | 354 ± 181 ↓ | 354 ± 176 ↓ | 226 ± 82 ↓ | ||
| 0.14 ± 0.09 | 0.02 ± 0.01 | 0.13 ± 0.05 | 0.23 ± 0.12 | ||
| 6894 ± 880 ↓ | 3470 ± 120 ↓ | 1526 ± 114 ↓ | |||
| 0.17 ± 0.08 | 0.13 ± 0.05 | 0.20 ± 0.07 | |||
| ZnPPIX in H2O (pH 9) | 521 nm | 574 nm | 630 nm | 560 nm | |
| 0.13 ± 0.08 ↓ | 1.85 ± 0.73 ↓ | 2.03 ± 0.82 ↓ | 11.8 ± 5.8 ↑ | ||
| 0.57 ± 0.18 | 0.86 ± 0.17 | 0.74 ± 0.28 | |||
| 1.34 ± 0.51 ↓ | 8.79 ± 1.93 ↓ | 13.0 ± 1.36 ↓ | |||
| 0.03 ± 0.01 | 0.14 ± 0.09 | 0.26 ± 0.11 | |||
| 13.6 ± 3.19 ↓ | |||||
| 0.12 ± 0.05 | |||||
| 1090 ± 280 ↓ | |||||
| 0.28 ± 0.09 | |||||
| BLG:ZnPPIX pH 9 | 462 nm | 506 nm | 565 nm | 605 nm | 550 nm |
| 2.71 ± 0.71 ↓ | 2.53 ± 0.24 ↓ | 6.75 ± 0.93 ↑ | 0.12 ± 0.07 ↑ | 0.41 ± 0.06 ↓ | |
| 0.71 ± 0.16 | 0.80 ± 0.25 | 0.62 ± 0.19 | 0.14 ± 0.06 | ||
| 332 ± 22 ↓ | 328 ± 21 ↓ | 354 ± 176 ↓ | 1.21 ± 0.52 ↓ | 1782 ± 122 ↑ | |
| 0.05± 0.03 | 0.07 ± 0.04 | 0.13 ± 0.07 | 0.55 ± 0.23 | ||
| 5243 ± 570 ↑ | 354 ± 181 ↓ | 1025 ± 337 ↑ | 5793 ± 600 ↓ | ||
| 0.09 ± 0.03 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.31 ± 0.18 | ||
| 6894 ± 880 ↓ | 3470 ± 120 ↓ | 1526 ± 114 ↓ | |||
| 0.15 ± 0.07 | 0.11 ± 0.06 | 0.23 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albalawi, A.; Castillo, O.; Denton, M.L.; Rickman, J.M.; Noojin, G.D.; Brancaleon, L. Irradiation of ZnPPIX Complexed with Bovine β-Lactoglobulin Causes Chemical Modifications and Conformational Changes of the Protein. Physchem 2023, 3, 411-439. https://doi.org/10.3390/physchem3040027
Albalawi A, Castillo O, Denton ML, Rickman JM, Noojin GD, Brancaleon L. Irradiation of ZnPPIX Complexed with Bovine β-Lactoglobulin Causes Chemical Modifications and Conformational Changes of the Protein. Physchem. 2023; 3(4):411-439. https://doi.org/10.3390/physchem3040027
Chicago/Turabian StyleAlbalawi, Abdullah, Omar Castillo, Michael L. Denton, John Michael Rickman, Gary D. Noojin, and Lorenzo Brancaleon. 2023. "Irradiation of ZnPPIX Complexed with Bovine β-Lactoglobulin Causes Chemical Modifications and Conformational Changes of the Protein" Physchem 3, no. 4: 411-439. https://doi.org/10.3390/physchem3040027
APA StyleAlbalawi, A., Castillo, O., Denton, M. L., Rickman, J. M., Noojin, G. D., & Brancaleon, L. (2023). Irradiation of ZnPPIX Complexed with Bovine β-Lactoglobulin Causes Chemical Modifications and Conformational Changes of the Protein. Physchem, 3(4), 411-439. https://doi.org/10.3390/physchem3040027