

Modulatory Effect of Human Immunodeficiency Virus on Circulating p53, miR-21, and miR-125b: Any Diagnostic Implication?

 , , and
, , and 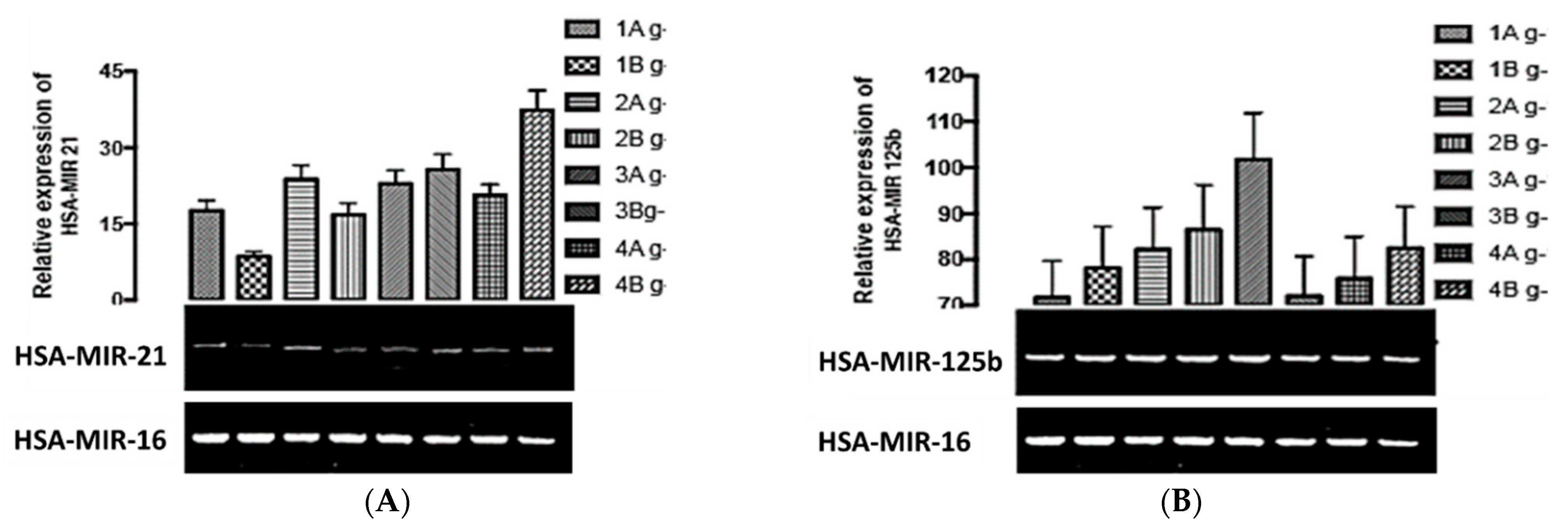{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Handling, and Assays
2.1.1. RNA Isolation from Serum
2.1.2. Complementary DNA Synthesis
2.1.3. Reverse Transcriptase Polymerase Chain Reaction
2.1.4. Gel Electrophoresis
2.2. Statistical Analyses
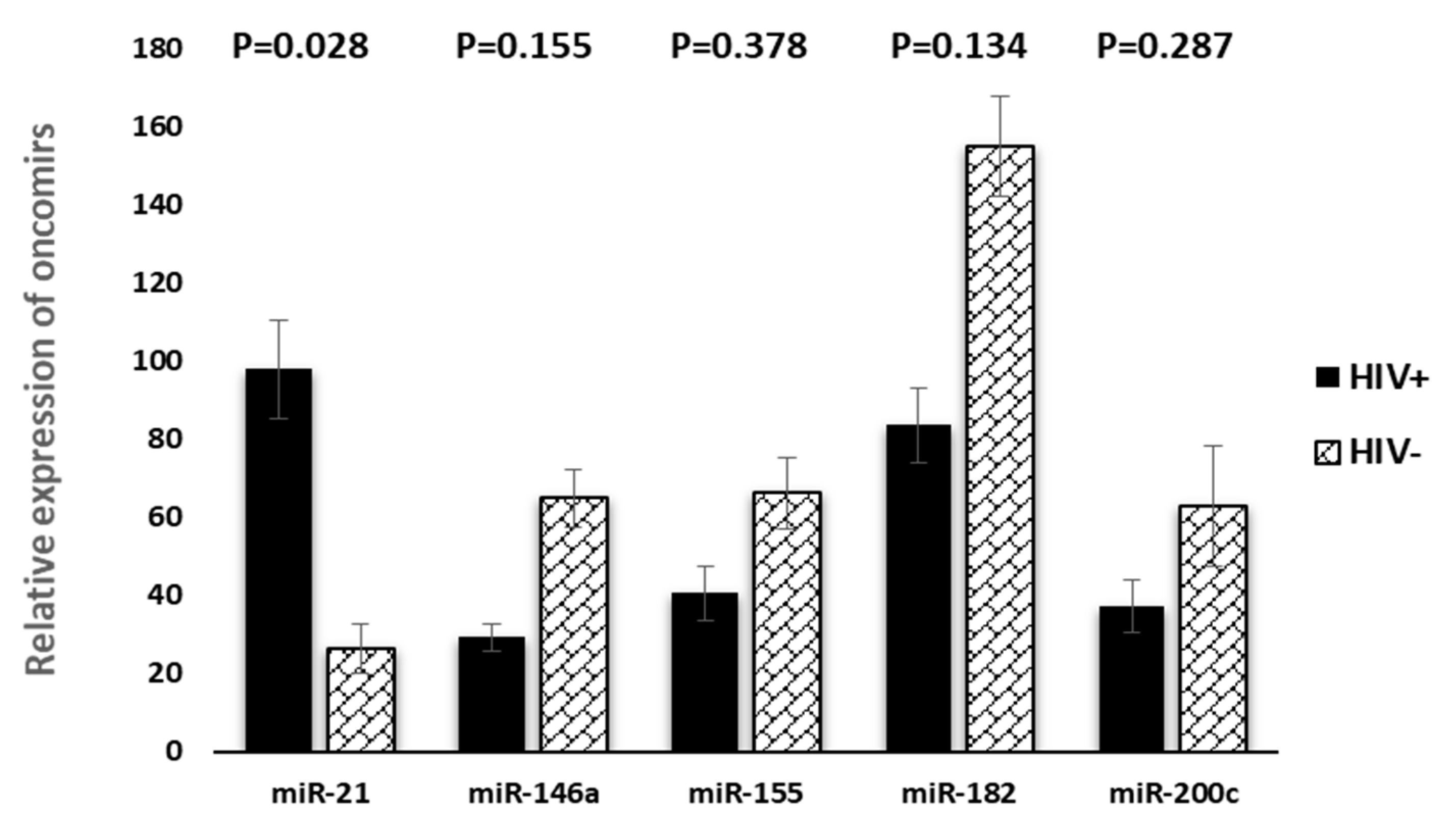
3. Results
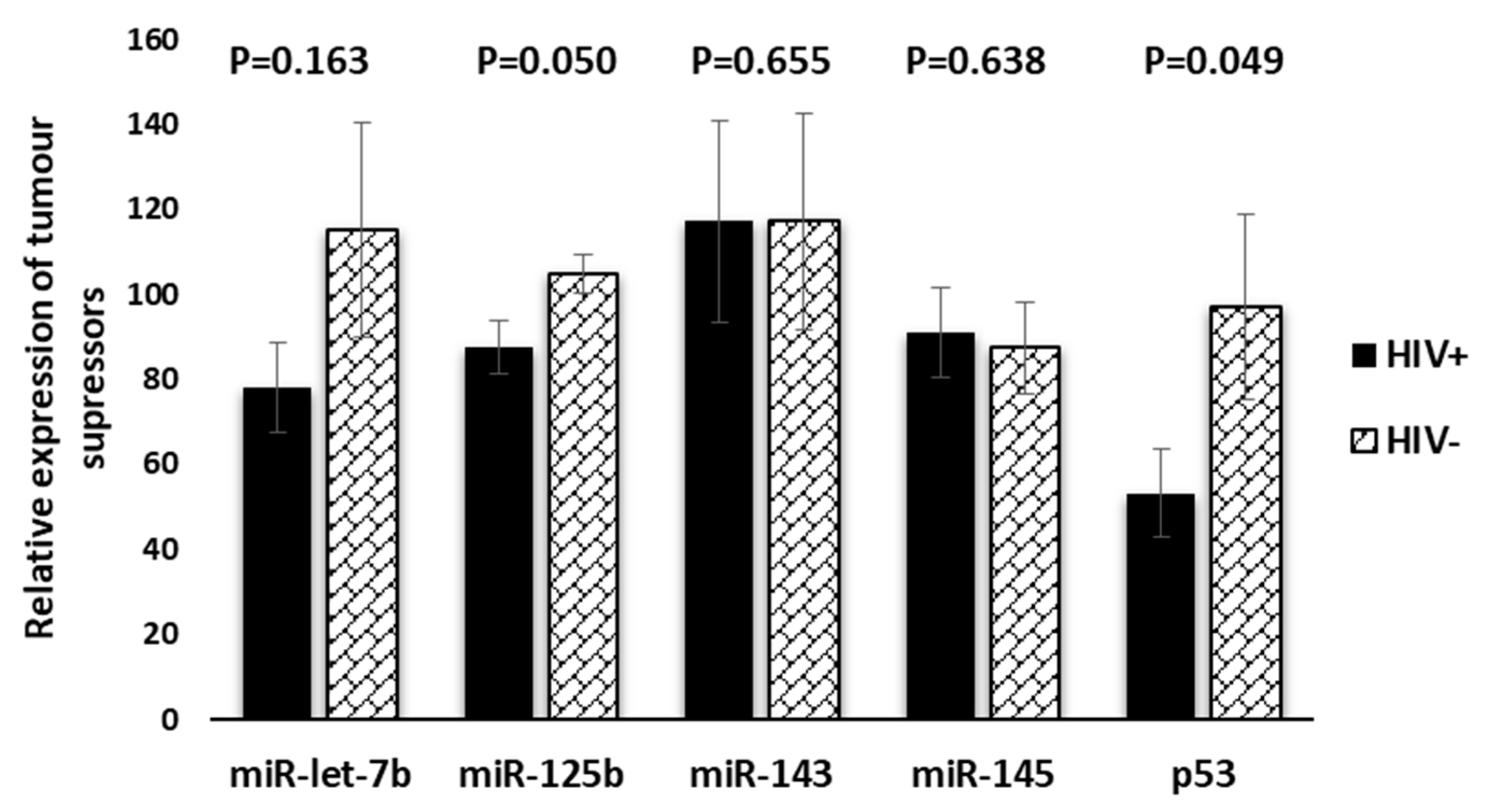
Relationships between Tumour Suppressors and Oncomirs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Fact Sheet on Cervical Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cervical-cancer#:~:text=Overview,%2D%20and%20middle%2Dincome%20countries (accessed on 22 February 2022).
- Awofala, A.A.; Ogundele, O.E. HIV epidemiology in Nigeria. Saudi J. Biol. Sci. 2018, 25, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adewole, I.F.; Benedet, J.L.; Crain, B.T.; Follen, M. Evolving a strategic approach to cervical cancer control in Africa. Gynecol. Oncol. 2005, 99, S209–S212. [Google Scholar] [CrossRef] [PubMed]
- Odutola, M.; Jedy-Agba, E.E.; Dareng, E.O.; Oga, E.A.; Igbinoba, F.; Otu, T.; Ezeome, E.; Hassan, R.; Adebamowo, C.A. Burden of cancers attributable to infectious agents in Nigeria: 2012–2014. Front. Oncol. 2016, 6, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denslow, S.A.; Rositch, A.F.; Firnhaber, C.; Ting, J.; Smith, J.S. Incidence and progression of cervical lesions in women with HIV: A systematic global review. Int. J. STD AIDS 2014, 25, 163–177. [Google Scholar] [CrossRef]
- Ranjith, B.; Sandeep, S.; Veerabathiran, R. RNA oncoviruses and their association with cancer implications. In Oncogenic Viruses; Academic Press: Cambridge, MA, USA, 2023; pp. 171–195. [Google Scholar] [CrossRef]
- Suzuki, K.; Matsubara, H. Recent advances in p53 research and cancer treatment. J. Biomed. Biotechnol. 2011, 2011, 978312. [Google Scholar] [CrossRef] [PubMed]
- Okoye, J.O.; Ngokere, A.A.; Onyenekwe, C.C.; Ogenyi, S.I.; Omotuyi, O. Circulating mir21 and mir125b in women living with human immunodeficiency virus: Utility of biomarkers for monitoring cervical carcinogenesis. Cancer Res. 2022, 82 (Suppl. S12). [Google Scholar] [CrossRef]
- Okoye, J.O.; Ngokere, A.A.; Onyenekwe, C.C.; Omotuyi, O.; Dada, D.I. Epstein-Barr virus, human papillomavirus and herpes simplex virus 2 co-presence severely dysregulates miRNA expression. Afr. J. Lab. Med. 2021, 10, 975. [Google Scholar] [CrossRef]
- Okoye, J.O.; Ngokere, A.A.; Onyenekwe, C.C.; Erinle, C.A. Comparable expression of miR-let-7b, miR-21, miR-182, miR-145, and p53 in serum and cervical cells: Diagnostic implications for early detection of cervical lesions. Int. J. Health Sci. 2019, 13, 29–38. [Google Scholar]
- Toni, L.S.; Garcia, A.M.; Jeffrey, D.A.; Jiang, X.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Optimization of phenol-chloroform RNA extraction. MethodsX 2018, 5, 599–608. [Google Scholar] [CrossRef]
- Parikh, V.N.; Park, J.; Nikolic, I.; Channick, R.; Paul, B.Y.; De Marco, T.; Hsue, P.; Chan, S.Y. Coordinated modulation of circulating miR-21 in HIV, HIV-associated pulmonary arterial hypertension, and HIV/HCV co-infection. J. Acquir. Immune Defic. Syndr. 2015, 70, 236. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.N.; Nguyen, L.N.; Zhao, J.; Schank, M.; Dang, X.; Cao, D.; Khanal, S.; Thakuri, B.K.C.; Lu, Z.; Zhang, J.; et al. Long non-coding RNA GAS5 regulates T cell functions via miR21-mediated signaling in people living with HIV. Front. Immunol. 2021, 12, 601298. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Cheng, L.; Höxtermann, S.; Xie, T.; Lu, X.; Wu, H.; Skaletz-Rorowski, A.; Brockmeyer, N.; Wu, N. Micro RNA-155 is a biomarker of T-cell activation and immune dysfunction in HIV-1-infected patients. HIV Med. 2017, 18, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Mantri, C.K.; Pandhare Dash, J.; Mantri, J.V.; Dash, C.C. Cocaine enhances HIV-1 replication in CD4+ T cells by down-regulating MiR-125b. PLoS ONE 2012, 7, e51387. [Google Scholar] [CrossRef]
- Ruelas, D.S.; Chan, J.K.; Oh, E.; Heidersbach, A.J.; Hebbeler, A.M.; Chavez, L.; Verdin, E.; Rape, M.; Greene, W.C. MicroRNA-155 reinforces HIV latency. J. Biol. Chem. 2015, 290, 13736–13748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egaña-Gorroño, L.; Escribà, T.; Boulanger, N.; Guardo, A.C.; León, A.; Bargalló, M.E.; García, F.; Gatell, J.M.; Plana, M.; Arnedo, M.; et al. HIV Controllers Consortium of the AIDS Spanish Network. Differential microRNA expression profile between stimulated PBMCs from HIV-1 infected elite controllers and viremic progressors. PLoS ONE 2014, 9, e106360. [Google Scholar] [CrossRef]
- Sadri Nahand, J.; Bokharaei-Salim, F.; Karimzadeh, M.; Moghoofei, M.; Karampoor, S.; Mirzaei, H.R.; Tabibzadeh, A.; Jafari, A.; Ghaderi, A.; Asemi, Z.; et al. MicroRNAs and exosomes: Key players in HIV pathogenesis. HIV Med. 2020, 21, 246–278. [Google Scholar] [CrossRef]
- Gupta, S.; Panda, P.K.; Hashimoto, R.F.; Samal, S.K.; Mishra, S.; Verma, S.K.; Mishra, Y.K.; Ahuja, R. Dynamical modeling of miR-34a, miR-449a, and miR-16 reveals numerous DDR signaling pathways regulating senescence, autophagy, and apoptosis in HeLa cells. Sci. Rep. 2022, 12, 4911. [Google Scholar] [CrossRef]
- Instated, S.D.; Gottwein, E.; Skalsky, R.L.; Luftig, M.A.; Cullen, B.R. Virally induced cellular microRNA miR-155 plays a key role in B-cell immortalization by Epstein-Barr virus. J. Virol. 2010, 84, 11670–11678. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef]
- Li, S.Y.; Zhang, Z.N.; Jiang, Y.J.; Fu, Y.J.; Shang, H. Transcriptional insights into the CD8+ T cell response in mono-HIV and HCV infection. J. Trans. Med. 2020, 18, 96. [Google Scholar] [CrossRef] [Green Version]
- Roderburg, C.; Koch, A.; Benz, F.; Vucur, M.; Spehlmann, M.; Loosen, S.H.; Luedde, M.; Rehse, S.; Lurje, G.; Trautwein, C.; et al. Serum levels of miR-143 predict survival in critically ill patients. Dis. Markers 2019, 2019, 4850472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akwiwu, E.; Okafor, A.; Akpan, P.; Akpotuzor, J.; Asemota, E.; Okoroiwu, H.; Anyanwu, S. Serum P53 Protein Level and Some Haematologic Parameters among Women of Reproductive Age Living with HIV Infection. Niger. J. Physiol. Sci. 2021, 36, 85–89. [Google Scholar] [PubMed]
- Smith, J.S.; Melendy, A.; Rana, R.K.; Pimenta, J.M. Age-specific prevalence of infection with human papillomavirus in females: A global review. J. Adolesc. Health 2008, 43, S5.e1–S5.e62. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okoye, J.O.; Ngokere, A.A.; Onyenekwe, C.C.; Omotuyi, O.I.; Ogenyi, S.I.; Obi, C.M.; Fasogbon, S.A. Modulatory Effect of Human Immunodeficiency Virus on Circulating p53, miR-21, and miR-125b: Any Diagnostic Implication? Venereology 2023, 2, 78-85. https://doi.org/10.3390/venereology2030008
Okoye JO, Ngokere AA, Onyenekwe CC, Omotuyi OI, Ogenyi SI, Obi CM, Fasogbon SA. Modulatory Effect of Human Immunodeficiency Virus on Circulating p53, miR-21, and miR-125b: Any Diagnostic Implication? Venereology. 2023; 2(3):78-85. https://doi.org/10.3390/venereology2030008
Chicago/Turabian StyleOkoye, Jude Ogechukwu, Anthony Ajuluchukwu Ngokere, Chinedum Charles Onyenekwe, Olaposi Idowu Omotuyi, Samuel Ifedioranma Ogenyi, Chioma Maureen Obi, and Samuel Ayobami Fasogbon. 2023. "Modulatory Effect of Human Immunodeficiency Virus on Circulating p53, miR-21, and miR-125b: Any Diagnostic Implication?" Venereology 2, no. 3: 78-85. https://doi.org/10.3390/venereology2030008