
Sex-Specific Differences in Cytokine Production Capacity in Patients with Gout Compared to Controls
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cell Preparation and Culture Conditions
2.3. Cytokine Measurements
2.4. Statistical Analysis
3. Results
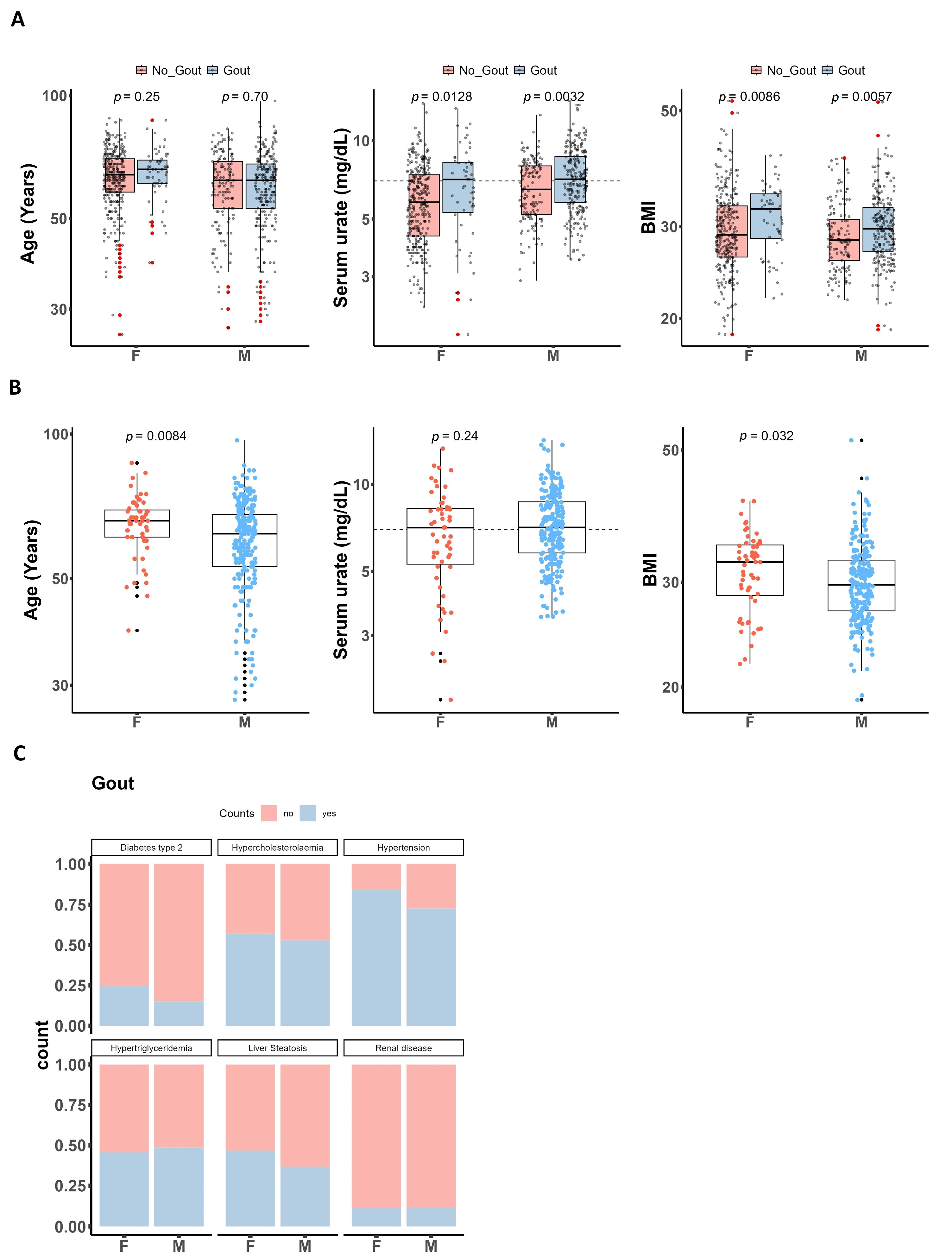
3.1. Characteristics of Patients with Gout and Healthy Controls
3.2. Sex-Specific Cytokine Production upon 24 h In Vitro Stimulation of PBMCs
3.3. Circulating Markers Are Elevated in Men with Gout
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Martinon, F.; Pétrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef]
- So, A.; Dumusc, A.; Nasi, S. The role of IL-1 in gout: From bench to bedside. Rheumatology 2018, 57, i12–i19. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Pandya, B.J.; Choi, H.K. Prevalence of gout and hyperuricemia in the US general population: The National Health and Nutrition Examination Survey 2007–2008. Arthritis Rheum. 2011, 63, 3136–3141. [Google Scholar] [CrossRef]
- te Kampe, R.; Janssen, M.; van Durme, C.; Jansen, T.L.; Boonen, A. Sex differences in the clinical profile among patients with gout: Cross-sectional analyses of an observational study. J. Rheumatol. 2021, 48, 286–292. [Google Scholar] [CrossRef]
- Harrold, L.R.; Etzel, C.J.; Gibofsky, A.; Kremer, J.M.; Pillinger, M.H.; Saag, K.G.; Schlesinger, N.; Terkeltaub, R.; Cox, V.; Greenberg, J.D. Sex differences in gout characteristics: Tailoring care for women and men. BMC Musculoskelet. Disord. 2017, 18, 108. [Google Scholar] [CrossRef]
- Bergsten, U.; Dehlin, M.; Klingberg, E.; Landgren, A.J.; Jacobsson, L.T.H. Gender differences in illness perceptions and disease management in patients with gout, results from a questionnaire study in Western Sweden. BMC Musculoskelet. Disord. 2023, 24, 300. [Google Scholar] [CrossRef] [PubMed]
- Van Lunzen, J.; Altfeld, M. Sex differences in infectious diseases-common but neglected. J. Infect. Dis. 2014, 209, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Angum, F.; Khan, T.; Kaler, J.; Siddiqui, L.; Hussain, A. The Prevalence of Autoimmune Disorders in Women: A Narrative Review. Cureus 2020, 12, e8094. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Verthelyi, D.; Klinman, D.M. Sex hormone levels correlate with the activity of cytokine-secreting cells in vivo. Immunology 2000, 100, 384–390. [Google Scholar] [CrossRef]
- Casimir, G.J.; Heldenbergh, F.; Hanssens, L.; Mulier, S.; Heinrichs, C.; Lefevre, N.; Désir, J.; Corazza, F.; Duchateau, J. Gender differences and inflammation: An in vitro model of blood cells stimulation in prepubescent children. J. Inflamm. 2010, 7, 28. [Google Scholar] [CrossRef]
- Li, F.; Boon, A.C.M.; Michelson, A.P.; Foraker, R.E.; Zhan, M.; Payne, P.R.O. Estrogen hormone is an essential sex factor inhibiting inflammation and immune response in COVID-19. Sci. Rep. 2022, 12, 9462. [Google Scholar] [CrossRef] [PubMed]
- Schurz, H.; Salie, M.; Tromp, G.; Hoal, E.G.; Kinnear, C.J.; Möller, M. The X chromosome and sex-specific effects in infectious disease susceptibility. Hum. Genom. 2019, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, I.; Dejager, L.; Libert, C. X-chromosome-located microRNAs in immunity: Might they explain male/female differences?: The X chromosome-genomic context may affect X-located miRNAs and downstream signaling, thereby contributing to the enhanced immune response of females. BioEssays 2011, 33, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, N.; Corazza, F.; Valsamis, J.; Delbaere, A.; De Maertelaer, V.; Duchateau, J.; Casimir, G. The number of X chromosomes influences inflammatory cytokine production following Toll-like receptor stimulation. Front. Immunol. 2019, 10, 1052. [Google Scholar] [CrossRef] [PubMed]
- ter Horst, R.; Jaeger, M.; Smeekens, S.P.; Oosting, M.; Swertz, M.A.; Li, Y.; Kumar, V.; Diavatopoulos, D.A.; Jansen, A.F.M.; Lemmers, H.; et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell 2016, 167, 1111–1124.e13. [Google Scholar] [CrossRef] [PubMed]
- ter Horst, R.; van den Munckhof, I.C.L.; Schraa, K.; Aguirre-Gamboa, R.; Jaeger, M.; Smeekens, S.P.; Brand, T.; Lemmers, H.; Dijkstra, H.; Galesloot, T.E.; et al. Sex-specific regulation of inflammation and metabolic syndrome in obesity. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Beenakker, K.G.M.; Westendorp, R.G.J.; De Craen, A.J.M.; Chen, S.; Raz, Y.; Ballieux, B.E.P.B.; Nelissen, R.G.H.H.; Later, A.F.L.; Huizinga, T.W.; Slagboom, P.E.; et al. Men Have a Stronger Monocyte-Derived Cytokine Production Response upon Stimulation with the Gram-Negative Stimulus Lipopolysaccharide than Women: A Pooled Analysis Including 15 Study Populations. J. Innate Immun. 2020, 12, 142–153. [Google Scholar] [CrossRef]
- Da Pozzo, E.; Giacomelli, C.; Cavallini, C.; Martini, C. Cytokine secretion responsiveness of lymphomonocytes following cortisol cell exposure: Sex differences. PLoS ONE 2018, 13, e0200924. [Google Scholar] [CrossRef]
- Joosten, L.A.B.; Netea, M.G.; Mylona, E.; Koenders, M.I.; Malireddi, R.K.S.; Oosting, M.; Stienstra, R.; Van De Veerdonk, F.L.; Stalenhoef, A.F.; Giamarellos-Bourboulis, E.J.; et al. Engagement of fatty acids with toll-like receptor 2 drives interleukin-1β production via the ASC/caspase 1 pathway in monosodium urate monohydrate crystal-induced gouty arthritis. Arthritis Rheum. 2010, 62, 3237–3248. [Google Scholar] [CrossRef]
- Singh, J.A.; Gaffo, A. Gout epidemiology and comorbidities. Semin. Arthritis Rheum. 2020, 50, S11–S16. [Google Scholar] [CrossRef] [PubMed]
- Hägg, S.; Jylhävä, J. Sex differences in biological aging with a focus on human studies. eLife 2021, 10, e63425. [Google Scholar] [CrossRef] [PubMed]
- Márquez, E.J.; Chung, C.H.; Marches, R.; Rossi, R.J.; Nehar-Belaid, D.; Eroglu, A.; Mellert, D.J.; Kuchel, G.A.; Banchereau, J.; Ucar, D. Sexual-dimorphism in human immune system aging. Nat. Commun. 2020, 11, 751. [Google Scholar] [CrossRef] [PubMed]
- Bongen, E.; Lucian, H.; Khatri, A.; Fragiadakis, G.K.; Bjornson, Z.B.; Nolan, G.P.; Utz, P.J.; Khatri, P. Sex Differences in the Blood Transcriptome Identify Robust Changes in Immune Cell Proportions with Aging and Influenza Infection. Cell Rep. 2019, 29, 1961–1973.e4. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.L.; Prior, J.A.; Belcher, J.; Hay, C.A.; Mallen, C.D.; Roddy, E. Gender-specific risk factors for gout: A systematic review of cohort studies. Adv. Rheumatol. 2019, 59, 24. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.V.; Gaffo, A.L. Managing Gout in Women: Current Perspectives. J. Inflamm. Res. 2022, 15, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Tin, A.; Marten, J.; Halperin Kuhns, V.L.; Li, Y.; Wuttke, M.; Kirsten, H.; Sieber, K.B.; Qiu, C.; Gorski, M.; Yu, Z.; et al. Target genes, variants, tissues and transcriptional pathways influencing human serum urate levels. Nat. Genet. 2019, 51, 1459–1474. [Google Scholar] [CrossRef] [PubMed]
- McCormick, N.; Yokose, C.; Lu, N.; Joshi, A.D.; Curhan, G.C.; Choi, H.K. Impact of adiposity on risk of female gout among those genetically predisposed: Sex-specific prospective cohort study findings over >32 years. Ann. Rheum. Dis. 2022, 81, 556–563. [Google Scholar] [CrossRef]
- Lakshmikanth, T.; Muhammad, S.A.; Olin, A.; Chen, Y.; Mikes, J.; Fagerberg, L.; Gummesson, A.; Bergström, G.; Uhlen, M.; Brodin, P. Human Immune System Variation during 1 Year. Cell Rep. 2020, 32, 107923. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, B.; Liu, X.; Li, H.; Xie, L.; Gao, Y.; Duan, R.; Li, Z.; Zhang, J.; Zheng, Y.; et al. Effects of sex and aging on the immune cell landscape as assessed by single-cell transcriptomic analysis. Proc. Natl. Acad. Sci. USA 2021, 118, e2023216118. [Google Scholar] [CrossRef]
- Gupta, S.; Nakabo, S.; Blanco, L.P.; O’Neil, L.J.; Wigerblad, G.; Goel, R.R.; Mistry, P.; Jiang, K.; Carmona-Rivera, C.; Chan, D.W.; et al. Sex differences in neutrophil biology modulate response to type I interferons and immunometabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 16481–16491. [Google Scholar] [CrossRef] [PubMed]
- So, J.; Tai, A.K.; Lichtenstein, A.H.; Wu, D.; Lamon-Fava, S. Sexual dimorphism of monocyte transcriptome in individuals with chronic low-grade inflammation. Biol. Sex Differ. 2021, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Topless, R.K.; Gaffo, A.; Stamp, L.K.; Robinson, P.C.; Dalbeth, N.; Merriman, T.R. Gout and the risk of COVID-19 diagnosis and death in the UK Biobank: A population-based study. Lancet Rheumatol. 2022, 4, e274–e281. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | n | Gout, n = 299 1 | No_Gout, n = 480 1 | p-Value 2 |
|---|---|---|---|---|
| Sex | 777 | <0.001 | ||
| F (female) | 56 (19%) | 324 (68%) | ||
| M (male) | 243 (81%) | 154 (32%) | ||
| Unknown | 0 | 2 | ||
| Age (years) | 763 | 63 (55, 69) | 63 (57, 70) | 0.3 |
| Unknown | 5 | 11 | ||
| Urate (mg/dL) | 762 | 7.10 (5.70, 8.69) | 6.10 (4.70, 7.60) | <0.001 |
| Unknown | 7 | 10 | ||
| BMI | 743 | 30.0 (26.8, 33.2) | 28.6 (26.2, 32.3) | 0.003 |
| Unknown | 8 | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badii, M.; Gaal, O.I.; Hotea, I.; Nica, V.; Mirea, A.M.; Mărginean, D.; HINT Consortium; Pamfil, C.; Rednic, S.; Popp, R.A.; et al. Sex-Specific Differences in Cytokine Production Capacity in Patients with Gout Compared to Controls. Gout Urate Cryst. Depos. Dis. 2024, 2, 133-143. https://doi.org/10.3390/gucdd2020012
Badii M, Gaal OI, Hotea I, Nica V, Mirea AM, Mărginean D, HINT Consortium, Pamfil C, Rednic S, Popp RA, et al. Sex-Specific Differences in Cytokine Production Capacity in Patients with Gout Compared to Controls. Gout, Urate, and Crystal Deposition Disease. 2024; 2(2):133-143. https://doi.org/10.3390/gucdd2020012
Chicago/Turabian StyleBadii, Medeea, Orsolya I. Gaal, Ioana Hotea, Valentin Nica, Andreea M. Mirea, Dragoş Mărginean, HINT Consortium, Cristina Pamfil, Simona Rednic, Radu A. Popp, and et al. 2024. "Sex-Specific Differences in Cytokine Production Capacity in Patients with Gout Compared to Controls" Gout, Urate, and Crystal Deposition Disease 2, no. 2: 133-143. https://doi.org/10.3390/gucdd2020012