
Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to theTraditional Malolactic Fermentation in Red Wine Production


Abstract
:1. Introduction
2. Results and Discussion
2.1. Fermentation Kinetics
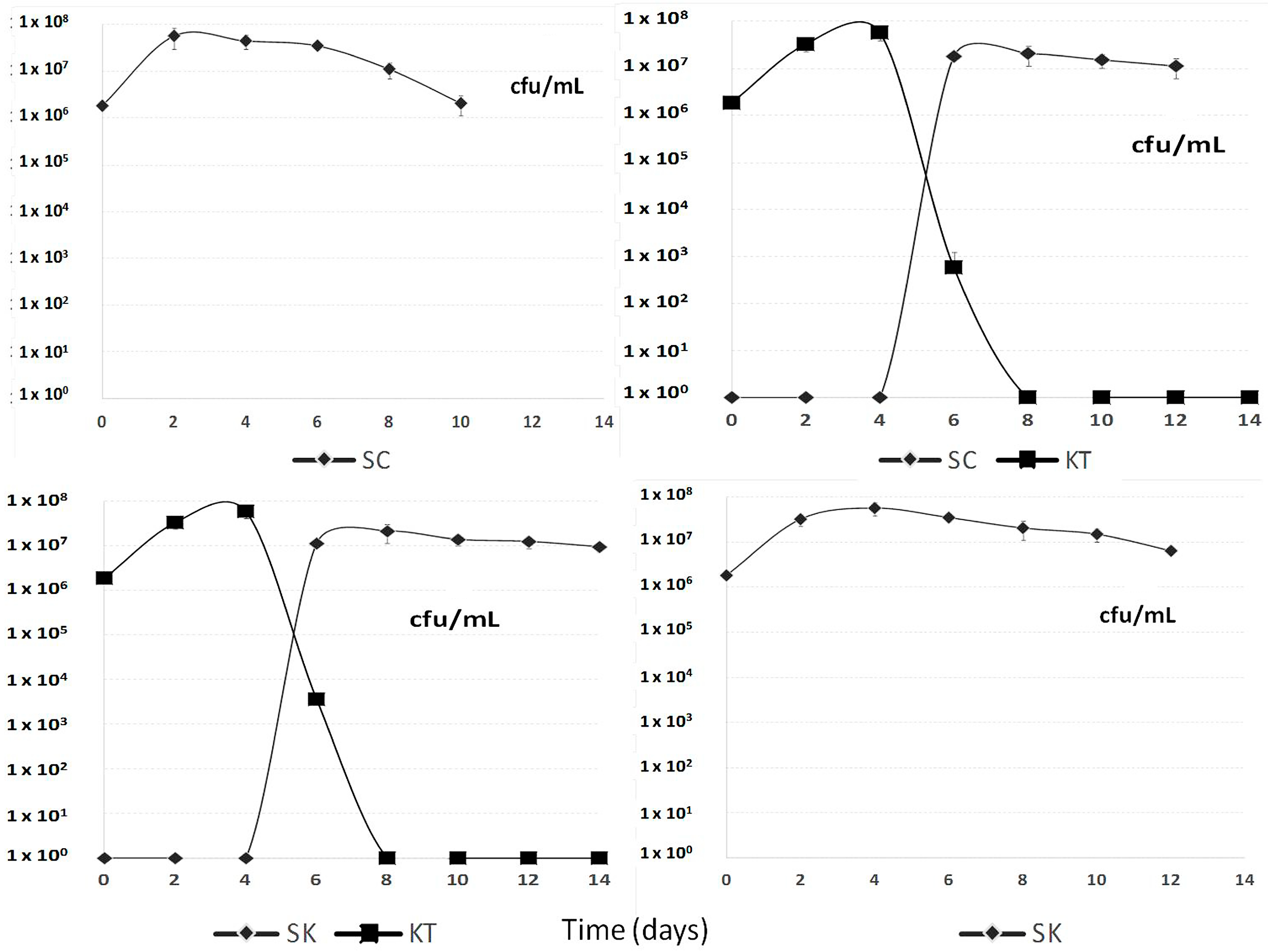
2.1.1. Yeast Population Kinetic

2.1.2. Sugar Consumption Kinetics

2.2. Chemical Parameter Monitoring
2.2.1. Glycerol

{kind=link}
{kind=link}
| Compounds | SC | SC + MLF | KT···SC | KT···SC + MLF | KT···SK | SK |
|---|---|---|---|---|---|---|
| l-Lactic Acid (g/L) | 0.01 ± 0.01a | 0.54 ± 0.08b | 2.75 ± 0.12c | 3.27 ± 0.19d | 2.96 ± 0.21c | 0.01 ± 0.01a |
| l-Malic Acid (g/L) | 0.92 ± 0.02b | 0.01 ± 0.01a | 0.89 ± 0.04b | 0.01 ± 0.01a | 0.01 ± 0.01a | 0.01 ± 0.01a |
| Acetic Acid (g/L) | 0.36 ± 0.01b | 0.44 ± 0.05c | 0.32 ± 0.02a | 0.39 ± 0.04bc | 0.37 ± 0.02b | 0.41 ± 0.02c |
| Residual Sugar (g/L) | 2.08 ± 0.30b | 0.12 ± 0.04a | 2.22 ± 052b | 0.16 ± 0.04a | 2.41 ± 0.58b | 2.13 ± 0.17b |
| Glycerol (g/L) | 5.96 ± 0.02a | 5.89 ± 0.05a | 6.48 ± 0.05b | 6.36 ± 0.06b | 6.65 ± 0.04c | 6.59 ± 0.03bc |
| Free SO2 (mg/L) | 26.12 ± 2.38a | 25.25 ± 3.43ab | 25.25 ± 3.28ab | 21.15 ± 1.28b | ||
| Total SO2 (mg/L) | 56.52 ± 2.43b | 44.13 ± 3.16a | 46.50 ± 3.21a | 58.58 ± 1.15b | ||
| Alcohol (% v/v) | 14.56 ± 0.01c | 14.54 ± 0.02c | 14.20 ± 0.04b | 14.18 ± 0.06b | 14.03 ± 0.05a | 14.23 ± 0.02b |
| pH | 3.94 ± 0.01c | 3.99 ± 0.02d | 3.74 ± 0.02a | 3.79 ± 0.02b | 3.83 ± 0.02b | 4.03 ± 0.02d |
| Urea | 1.43 ± 0.01b | 1.45 ± 0.02b | 0.12 ± 0.04a | 0.08 ± 0.01a | ||
| Color Intensity | 6.16 ± 0.03b | 5.38 ± 0.06a | 6.29 ± 0.06c | 5.51 ± 0.07a | 6.42 ± 0.08c | 6.88 ± 0.03d |
| Citric Acid (g/L) | 0.22 ± 0.01a | 0.03 ± 0.02b | 0.24 ± 0.03a | 0.04 ± 0.03b | 0.23 ± 0.03a | 0.22 ± 0.02a |
2.2.2. Pyruvic Acid
2.2.3. Alcohol
2.2.4. SO2
2.2.5. l-Lactic Acid
2.2.6. l-Malic Acid
2.2.7. Acetic Acid
2.2.8. Biogenic Amines
| Compounds | SC | SC + MLF | KT···SC | KT···SC + MLF | KT···SK | SK |
|---|---|---|---|---|---|---|
| Histamine (mg/L) | 0.43 ± 0.02a | 1.46 ± 0.06b | 0.42 ± 0.04a | 1.48 ± 0.15b | 0.44 ± 0.04a | 0.38 ± 0.02a |
| Tiramine (mg/L) | 0.25 ± 0.01a | 0.36 ± 0.04b | 0.26 ± 0.02a | 0.38 ± 0.06b | 0.22 ± 0.03a | 0.26 ± 0.03a |
| Phenylethylamine (g/L) | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Putrescine (g/L) | 1.78 ± 0.03a | 2.18 ± 0.18b | 1.82 ± 0.11a | 2.24 ± 0.21b | 1.71 ± 0.08a | 1.88 ± 0.07a |
| Cadaverine (g/L) | 0.51 ± 0.02a | 0.65 ± 0.04b | 0.49 ± 0.05a | 0.69 ± 0.07b | 0.52 ± 0.03ab | 0.55 ± 0.03a |
2.3. Sensory Evaluation
3. Experimental Section
3.1. Microorganisms
3.2. Vinification
3.3. Analytical Determinations of Non-Volatile Compounds
3.4. Microvinifications Growth Kinetics
3.5. Analytical Determinations of Biogenic Amines
3.6. Sensory Evaluation
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Benito, S.; Palomero, P.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Schizosaccharomyces . In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 365–370. [Google Scholar]
- Rodriguez, S.B.; Roy, J. Factors influencing the utilisation of l-malate by yeasts. Thornton FEMS Microbiol. Lett. 1990, 60, 17–22. [Google Scholar]
- Thornton, R.J.; Rodriguez, S.B. Deacidification of red and white wines by a mutant of Schizosaccharomyces malidevorans under commercial winemaking conditions. Food Microbiol. 1996, 13, 475–482. [Google Scholar] [CrossRef]
- Su, J.; Wang, T.; Wang, Y.; Li, Y.Y.; Li, H. The use of lactic acid-producing, malic acid-producing, or malic acid-degrading yeast strains for acidity adjustment in the wine industry. Appl. Microbiol. Biotechnol. 2014, 98, 2395–2413. [Google Scholar] [CrossRef] [PubMed]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–333. [Google Scholar] [CrossRef] [PubMed]
- Contreras., A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin., C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microb. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T.; Reglero, G.; Herraiz, M.; Martin-Alvarez, P.J.; Cabezudo, M.D. The influence of the yeast and type of culture on the volatile composition of wines fermented without sulfur dioxide. Am. J. Enol. Vitic. 1990, 41, 313–318. [Google Scholar]
- Zironi, R.; Romano, P.; Suzzi, G.; Battistutta, F.; Comi, G. Volatile metabolites produced in wine by mixed and sequential cultures of Hanseniaspora guilliermondii or Kloeckera apiculata and Saccharomyces cerevisiae. Biotechnol. Lett. 1993, 15, 235–238. [Google Scholar] [CrossRef]
- Viana, F.; Belloch, C.; Valles, S.; Manzanares, P. Monitoring a mixed starter of Hanseniaspora vineae-Saccharomyces cerevisiae in natural must: Impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 2011, 151, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Azzolini, M.; Fedrizzi, B.; Tosi, E.; Finato, F.; Vagnoli, P.; Scrinzi, C.; Zapparoli, G. Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 2012, 235, 303–313. [Google Scholar] [CrossRef]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida zemplinina in Sicilian musts and selection of a strain for wine mixed fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Comitini, F.; Gobbi, M.; Lencioni, L.; Mannazzu, I.; Ciani, M. Potential spoilage non-Saccharomyces yeasts in mixed cultures with Saccharomyces cerevisiae. Ann. Microbiol. 2011, 61, 137–144. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderon, F.; Palmero, D.; Suarez-Lepe, J.A. Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 2013, 236, 29–36. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.; Palacios-Garcia, A.T.; Garcia-Romero, E. Enhancement of flavour properties in wines using sequential inoculations of non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces yeast starter. Vitis 2011, 50, 177–182. [Google Scholar]
- Izquierdo-Cañas, P.M.; García-Romero, E.; Heras, J.M.; Fernández-González, M. Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar]
- Peinado, R.A.; Moreno, J.J.; Maestre, O.; Ortega, J.M.; Medina, M.; Mauricio, J.C. Gluconic acid consumption in wines by Schizosaccharomyces pombe and its effect on the concentrations of major volatile compounds and polyols. J. Agric. Food. Chem. 2004, 52, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Mauricio, J.C.; Medina, M.; Moreno, J.J. Effect of Schizosaccharomyces pombe on aromatic compounds in dry sherry wines containing high levels of gluconic acid. J. Agric. Food. Chem. 2004, 52, 4529–4534. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Moreno, J.J.; Medina, M.; Mauricio, J.C. Potential application of a glucose-transport-deficient mutant of Schizosaccharomyces pombe for removing gluconic acid from grape must. J. Agric. Food Chem. 2005, 53, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Moreno, J.J.; Maestre, O. Removing gluconic acid by using different treatments with a Schizosaccharomyces pombe mutant: Effect on fermentation by products. Food Chem. 2007, 104, 457–465. [Google Scholar] [CrossRef]
- Peinado, R.A.; Maestre, O.; Mauricio, J.C. Use of a Schizosaccharomyces pombe mutant to reduce the content in gluconic acid of must obtained from rotten grapes. J. Agric. Food Chem. 2009, 57, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Palomero, P.; Morata, A.; Calderón, F.; Suárez-Lépe, J.A. New applications for Schizosaccharomyces pombe in the alcoholic fermentation of red wines. Int. J. Food Sci. Technol. 2012, 47, 2101–2108. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, P.; Gálvez, L.; Morata, A.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Quality and Composition of Red Wine Fermented with Schizosaccharomyces pombe as Sole Fermentative Yeast, and in Mixed and Sequential Fermentations with Saccharomyces cerevisiae. Food Technol. Biotechnol. 2014, 52, 376–382. [Google Scholar]
- Jeffares, D.C. The genomic and phenotypic diversity of Schizosaccharomyces pombe. Nat. Genet. 2015, 47, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Gálvez, L.; Palomero, F.; Calderón, F.; Morata, A.; Suárez-Lepe, J.A. Schizosaccharomyces selective differential media. Afr. J. Microbiol. Res. 2013, 7, 3026–3036. [Google Scholar]
- Benito., S.; Palomero, P.; Calderón, F.; Palmero, D.; Suárez-Lépe, J.A. Selection of Appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeast in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with winemaking. World J. Microbiol. Biotechnol. 1998, 14, 199–203. [Google Scholar] [CrossRef]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: Potential applications in the wine industry. Int. J. Food. Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Merico, A.; Sulo, P.; Piskur, J.; Compagno, C. Fermentative lifestyle in yeasts belonging to the Saccharomyces complex. FEBS J. 2007, 274, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Kutyna, D.R.; Varela, C.; Henschke, P.A.; Chambers, P.J.; Stanley, G.A. Microbiological approaches to lowering ethanol concentration in wine. Trends Food Sci. Technol. 2010, 21, 293–302. [Google Scholar] [CrossRef]
- Blanco, P. Guia sobre la utilización de levaduras no-Saccharomyces en la elaboración de vino, 1st ed.Ministerio de Agricultura, Alimentación y Medio Ambiente (Gobierno de España): Madrid, España, 2014; pp. 9–18.
- Lehtonen, P. Determination of amines and amino acids in wine: A review. Am. J. Enol. Vitic. 1996, 47, 127–133. [Google Scholar]
- Alcaide-Hidalgo, J.M.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Polo, M.C. Influence of malolactic fermentation, postfermentative treatments and ageing with lees on nitrogen compounds of red wines. Food. Chem. 2007, 103, 572–581. [Google Scholar] [CrossRef]
- Tegmo-Larsson, I.M.; Spittler, T.D.; Rodriguez, S.B. Effect of malolactic fermentation on ethyl carbamate formation in Chardonnay. Am. J. Enol. Vitic. 1989, 40, 106–108. [Google Scholar]
- Lubbers, M.W.; Rodriguez, S.B.; Honey, N.K.; Thornton, R.J. Purification and characterization of urease from Schizosaccharomyces pombe. Can. J. Microbiol. 1996, 42, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, T.L.; Kennedy, A.; Vasconcelos, M.C. Use of Microscale Fermentations in Grape and Wine Research. Am. J. Enol. Vitic. 2007, 58, 534–539. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W. Methods for the isolation, maintenance and identification of yeasts. In The Yeast A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 79–80. [Google Scholar]
- Vaughnan-Martini, A.; Martini, A. Determination of ethanol production. In The Yeast A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; p. 107. [Google Scholar]
- Compendium of International Methods of Wine and Must Analysis; The International Organisation of Vine and Wine: Paris, France, 2015.
- Morris, E.O.; Eddy, A.A. Method for the measurement of wild yeast infection in pitching yeast. J. Inst. Brew. 1957, 63, 34–35. [Google Scholar] [CrossRef]
- International Organization for Standardization. Sensory analysis—Apparatus—Wine-Tasting Glass. 2010. Available online: http://www.iso.org/iso/catalogue_detail?csnumber=9002 (accessed on 11 May 2015).
- Sample Availability: Samples are available from authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to theTraditional Malolactic Fermentation in Red Wine Production. Molecules 2015, 20, 9510-9523. https://doi.org/10.3390/molecules20069510
Benito Á, Calderón F, Palomero F, Benito S. Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to theTraditional Malolactic Fermentation in Red Wine Production. Molecules. 2015; 20(6):9510-9523. https://doi.org/10.3390/molecules20069510
Chicago/Turabian StyleBenito, Ángel, Fernando Calderón, Felipe Palomero, and Santiago Benito. 2015. "Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to theTraditional Malolactic Fermentation in Red Wine Production" Molecules 20, no. 6: 9510-9523. https://doi.org/10.3390/molecules20069510
APA StyleBenito, Á., Calderón, F., Palomero, F., & Benito, S. (2015). Combine Use of Selected Schizosaccharomyces pombe and Lachancea thermotolerans Yeast Strains as an Alternative to theTraditional Malolactic Fermentation in Red Wine Production. Molecules, 20(6), 9510-9523. https://doi.org/10.3390/molecules20069510