
Genetic Diversity Analysis of Tomato (Solanum lycopersicum L.) with Morphological, Cytological, and Molecular Markers under Heat Stress

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Morphological and Phenotypic Study
2.2.1. Vegetative Traits
2.2.2. Yield and Its Components
2.2.3. Fruit Characteristics
2.2.4. Flowering Parameters
2.3. Cytological Study
2.4. Molecular Marker Analysis
2.5. Statistical Analysis
3. Results
3.1. Morphological and Phenotypic Diversity Evaluation
3.1.1. Vegetative Traits
3.1.2. Flowering and Fruit Traits
3.1.3. Total Yield
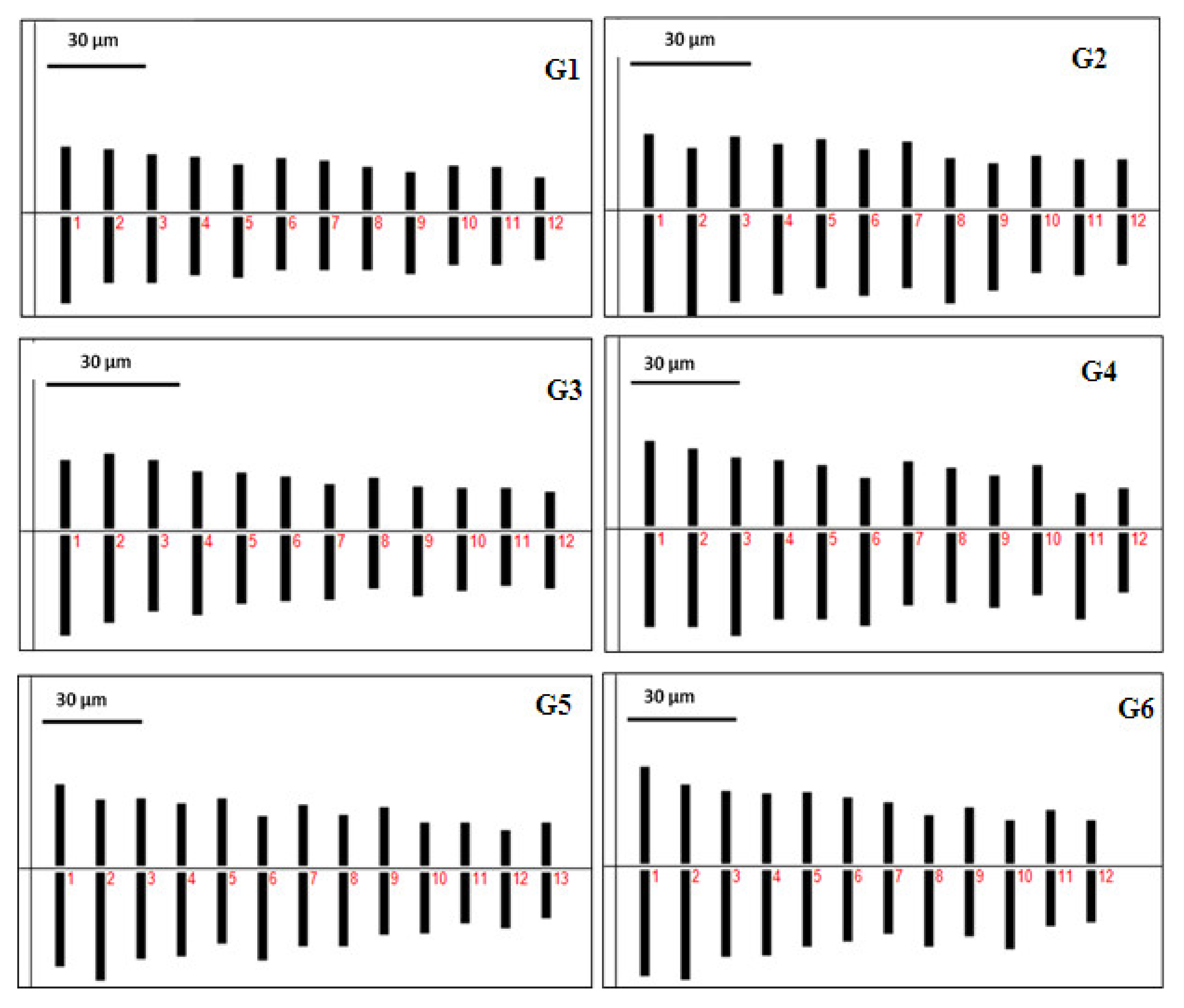
3.2. Cytological Studies
3.3. Molecular Diversity as Detected by RAPD, ISSR, and SCoT Analysis
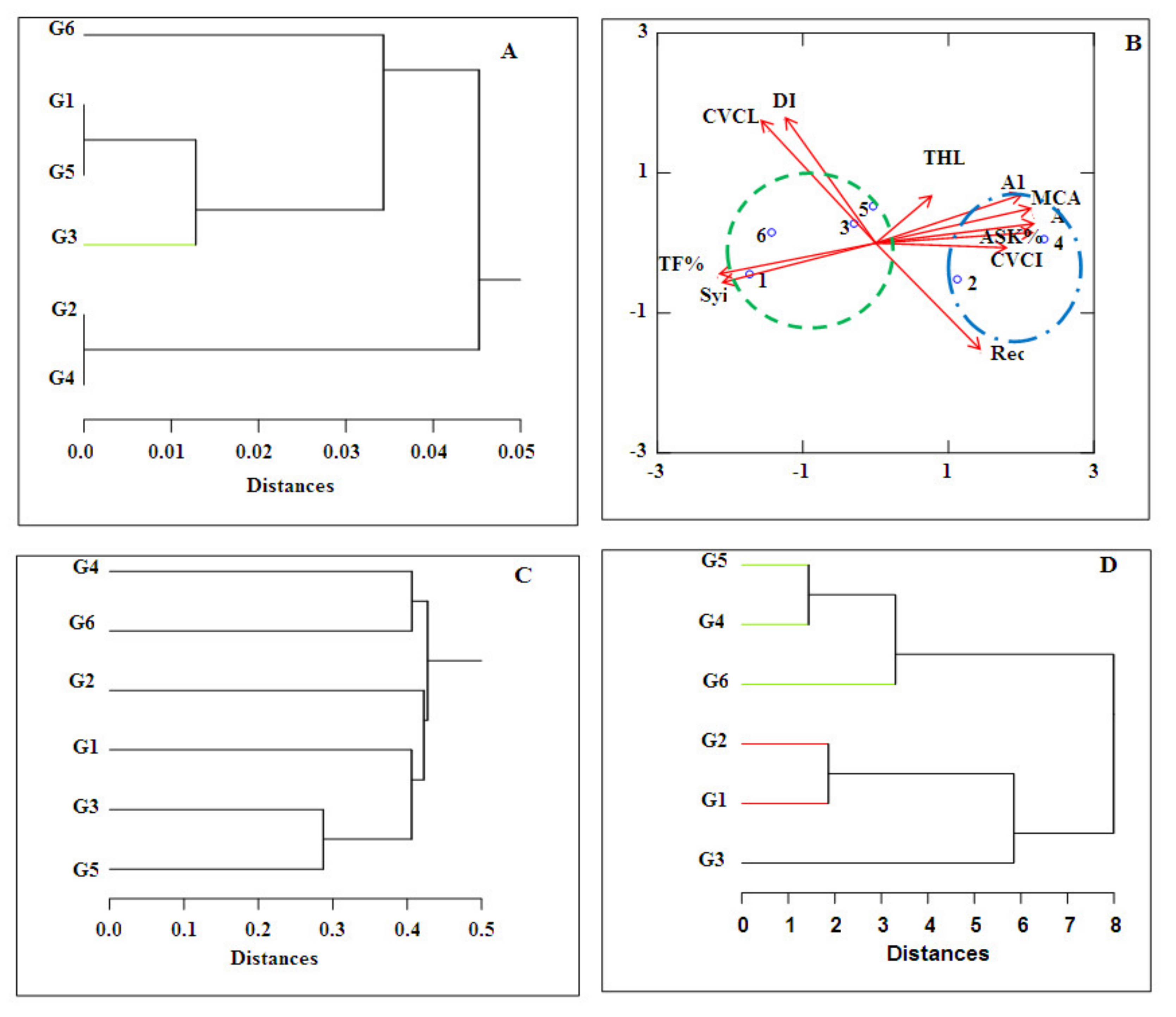
3.4. Statistical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siddiqui, M.H.; Alamri, S.; Alsubaie, Q.D.; Ali, H.M.; Khan, M.N.; Al-Ghamdi, A.; Ibrahim, A.A.; Alsadon, A. Exogenous nitric oxide alleviates sulfur deficiency-induced oxidative damage in tomato seedlings. Nitric Oxide 2020, 94, 95–107. [Google Scholar] [CrossRef]
- FAOSTAT. Crops: Tomatoes. 2019. Available online: http://www.fao.org/faostat/ar/#data/QC (accessed on 9 August 2019).
- Elnaga, M.A.; Mona, M.A. An Economic Study for Production of Winter Tomato Crop in Bir Al-Abd, North Sinai Governorate. J. Agric. Econom. Soc. Sci. 2018, 9, 885–889. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Masouleh, S.S.S.; Sassine, Y.N. Molecular and biochemical responses of horticultural plants and crops to heat stress. Ornam. Hortic. 2020, 26, 148–158. [Google Scholar] [CrossRef]
- Xu, J.; Driedonks, N.; Rutten, M.J.M.; Vriezen, W.H.; De Boer, G.-J.; Rieu, I. Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (Solanum lycopersicum). Mol. Breed. 2017, 37, 1–9. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Li, X.; Dos Santos, T.M.; Rosenqvist, E.; Ottosen, C.-O. Combined high light and heat stress induced complex response in tomato with better leaf cooling after heat priming. Plant Physiol. Biochem. 2020, 151, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Comlekcioglu, N.; Simsek, O.; Boncuk, M.; Aka-Kacar, Y. Genetic characterization of heat tolerant tomato (Solanum lycopersicon) genotypes by SRAP and RAPD markers. Genet. Mol. Res. 2010, 9, 2263–2274. [Google Scholar] [CrossRef]
- Mansour, A.; Ismail, H.M.; Ramadan, M.F.; Gyulai, G. Variations in tomato (Lycopersicon esculentum) cultivars grown under heat stress. J. Consum. Prot. Food Saf. 2009, 4, 118–127. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Asante, I.K.; Danquah, E.Y. Identification of new sources of heat tolerance in cultivated and wild tomatoes. Euphytica 2021, 217, 1–16. [Google Scholar] [CrossRef]
- El-Aziz, M.A.; Farid, S.; Elkomey, S. Evaluation of Molecular and Phenotypic Diversity in Relation to Heterosis in Some Tomato Lines Under Different Climatic Conditions. J. Agric. Chem. Biotechnol. 2016, 7, 141–151. [Google Scholar] [CrossRef]
- El-Mansy, A.B.; Metwally, E.I.; Elkasas, A.I.; El-Tantawy, A.M.; Mahmoud, M.I. Performance, Heritability and Correlation Coefficients for Some Important Traits in Tomato Under North Sinai Condition. Sinai J. Appl. Sci. 2015, 4, 79–94. [Google Scholar] [CrossRef]
- Wang, L.-J.; Gao, M.-D.; Sheng, M.-Y.; Yin, J. Cluster analysis of karyotype similarity coefficients in Epimedium (Berberidaceae): Insights in the systematics and evolution. PhytoKeys 2020, 161, 11–26. [Google Scholar] [CrossRef]
- Gianfranco, V.; Ravalli, C.; Cremonini, R. The karyotype as a tool to identify plant species: Vicia species belonging to Vicia subgenus. Caryologia 2008, 61, 300–319. [Google Scholar] [CrossRef]
- Cruz, G.A.D.S.; Filho, J.R.D.M.; Vasconcelos, S.; Gitaí, J.; De Campos, J.M.S.; Viccini, L.F.; Zizka, G.; Leme, E.M.C.; Brasileiro-Vidal, A.C.; Benko-Iseppon, A.M. Genome size evolution and chromosome numbers of species of the cryptanthoid complex (Bromelioideae, Bromeliaceae) in a phylogenetic framework. Bot. J. Linn. Soc. 2019, 192, 887–899. [Google Scholar] [CrossRef]
- Tlili, A.; Gouja, H.; Vallès, J.; Garnatje, T.; Buhagiar, J.; Neffati, M. Chromosome number and genome size in Atriplex mollis from southern Tunisia and Atriplex lanfrancoi from Malta (Amaranthaceae). Plant Syst. Evol. 2020, 306, 11. [Google Scholar] [CrossRef]
- Fedorova, A. Chromosome Numbers of Flowering Plants; Otto Koeltz Science Publishers: Koenigstein, Germany, 1969. [Google Scholar]
- Hemantaranjan, A. Heat Stress Responses and Thermotolerance. Adv. Plants Agric. Res. 2014, 1, 1–10. [Google Scholar] [CrossRef]
- Kandel, D.R.; Bedre, R.H.; Mandadi, K.K.; Crosby, K.; Avila, C.A. Genetic Diversity and Population Structure of Tomato (Solanum lycopersicum) Germplasm Developed by Texas A&M Breeding Programs. Am. J. Plant Sci. 2019, 10, 1154–1180. [Google Scholar] [CrossRef]
- Vargas, J.E.E.; Aguirre, N.C.; Coronado, Y.M. Study of the genetic diversity of tomato (Solanum spp.) with ISSR markers. Rev. Ceres 2020, 67, 199–206. [Google Scholar] [CrossRef]
- Brake, M.; Al-Gharaibeh, M.; Hamasha, H.; Al Sakarneh, N.; Alshomali, I.; Migdadi, H.; Qaryouti, M.; Haddad, N. Assessment of genetic variability among Jordanian tomato landrace using inter-simple sequence repeats markers. JJBS 2021, 14, 91–95. [Google Scholar]
- Ezekiel, C.N.; Nwangburuka, C.C.; Ajibade, O.A.; Odebode, A.C. Genetic diversity in 14 tomato (Lycopersicon esculentum Mill.) varieties in Nigerian markets by RAPD-PCR technique. Afr. J. Biotechnol. 2011, 10, 4961–4967. [Google Scholar]
- Collard, B.C.Y.; Maackill, D.J. Start Codon Targted (SCoT) polymorphism: A simple novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol. Bio. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Abdein, M.A.; El-Moneim, D.A.; Taha, S.S.; Al-Juhani, W.S.; Mohamed, S.E. Molecular characterization and genetic relationships among some tomato genotypes as revealed by ISSR and SCoT markers. Egypt. J. Genet. Cytol. 2018, 47, 139–159. Available online: http://journal.esg.net.eg/index.php/EJGC/article/view/281 (accessed on 6 September 2018).
- Dijkhuizen, A.; Kennard, W.C.; Havey, M.J.; Staub, J.E. RFLP variation and genetic relationships in cultivated cucumber. Euphtica 1996, 90, 79–87. [Google Scholar]
- Sensoy, S.; Büyükalaca, S.; Abak, K. Evaluation of genetic diversity in Turkish melons (Cucumis melo L.) based on phenotypic characters and RAPD markers. Genet. Resour. Crop. Evol. 2007, 54, 1351–1365. [Google Scholar] [CrossRef]
- Athanasios, L.T.; Koutita, O.; Anastasiadou, A. Description and analysis of genetic diversity among squash accessions. Braz. Arch. Biol. Technol. 2009, 52, 271–283. [Google Scholar]
- Zhang, J.; Li, X.D.; Jiang, T.L.; Cao, X.J. Photosynthesis and ultrastructure of photosynthetic apparatus in tomato leaves under elevated temperature. Photosynthetic 2014, 52, 430–436. [Google Scholar] [CrossRef]
- Cox, H.E.; Pearson, D. The Chemical Analysis of Foods; Chemical Publishing Co., Inc: New York, NY, USA, 1962; pp. 136–144. [Google Scholar]
- Hussein, E.H.A.; Mohamed, A.A.; Attia, S.; Adawy, S.S. Molecular characterization and genetic relationships among cotton genotypes 1- RAPD, ISSR and SSR analysis. Arab J. Biotech. 2006, 9, 313–328. [Google Scholar]
- Anderson, J.A.; Churchill, G.A.; Autrique, J.E.; Tanksley, S.D.; Sorrells, M.E. Optimizing parental selection for genetic linkage maps. Genome 1993, 36, 181–186. [Google Scholar] [CrossRef]
- Nagaraju, J.; Reddy, K.D.; Nagaraja, G.M.; Sethuraman, B.N. Comparison of multilocus RFLPs and PCR-based marker systems for genetic analysis of the silkworm, Bombyx mori. Heredity 2001, 86, 588–597. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Cochran, W.C.; Cox, G.M. Experimental Design, 2nd ed.; John Willey and Sons: New York, NY, USA, 1957. [Google Scholar]
- Duncan, B.D. Multiple range and multiple F test. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Wilkinson, L. SYSTAT: The System Analysis for Statistic SYSTAT; Prentice Hall: Evanston, IL, USA, 1997. [Google Scholar]
- Luo, C.; He, X.-H.; Chen, H.; Ou, S.-J.; Gao, M.-P. Analysis of diversity and relationships among mango cultivars using Start Codon Targeted (SCoT) markers. Biochem. Syst. Ecol. 2010, 38, 1176–1184. [Google Scholar] [CrossRef]
- Latif, M.A.; Yusop, M.R.; Rahman, M.M.; Talukdar, M.B.; Rafii, M. Microsatellite and minisatellite markers based DNA fingerprinting and genetic diversity of blast and ufra resistant genotypes. Comptes Rendus Biol. 2011, 334, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.Y.A.M.; Elsaid, E.M.; Habiba, R.M. Selection for heat tolerance in tomato ex situ germplasm. J. Agric. Chem. Biotechnol. 2015, 6, 657–673. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen, C.-O.; Wu, Z. Screening and validation of tomato genotypes under heat stress using Fv/Fm to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Molina-Bravo, R.; Arellano, C.; Sosinski, B.R.; Fernandez, G.E. A protocol to assess heat tolerance in a segregating population of raspberry using chlorophyll fluorescence. Sci. Hortic. 2011, 130, 524–530. [Google Scholar] [CrossRef]
- Golam, F.; Prodhan, Z.H.; Nezhadahmadi, A.; Rahman, M. Heat Tolerance in Tomato. Life Sci. J. 2012, 9, 1936–1950. [Google Scholar]
- Peet, M.M.; Willits, D.H.; Gardner, R. Response of ovule development and post- pollen production processes in male-sterile tomatoes tochronic, sub-acute high temperature stress. J. Exp. Bot. 1997, 48, 101–111. [Google Scholar] [CrossRef]
- Kafizadeh, N.; Carapetian, J.; Kalantari, K.M. Effect of heat stress on pollen viability and pollen tube growth in pepper. Res. J. Biol. Sci. 2008, 3, 1159–1162. [Google Scholar]
- El-Tantawy, E.M.; Rewaa, S.E. Improving Tomato Productivity under High Temperature Conditions. Hortsci. J. Suez Canal Univ. 2017, 6, 15–29. [Google Scholar]
- Zhou, R.; Kjaer, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.-O. Physiological Response to Heat Stress During Seedling and Anthesis Stage in Tomato Genotypes Differing in Heat Tolerance. J. Agron. Crop. Sci. 2017, 203, 68–80. [Google Scholar] [CrossRef]
- El-Twab, M.H.A.; Barakat, N.A.M.; El-Hafeez, A.A.A. Cytogenetical and ecological studies of some wild congeneric species in the Solanaceae distributed in Upper Egypt. Chromosom. Bot. 2010, 5, 65–73. [Google Scholar] [CrossRef]
- Banks, P. A new diploid chromosome number for tomato (Lycopersicon esculentum). Can. J. Genet. Cytol. 1984, 26, 636–639. [Google Scholar] [CrossRef]
- Badr, A.; Khalifa, S.F.; Aboel-Atta, A.I.; Abou-El-Enain, M.M. Chromosomal Criteria and Taxonomic Relationships in the Solanaceae. Cytologia 1997, 62, 103–113. [Google Scholar] [CrossRef]
- Imai, H.T.; Satta, Y.; Takahata, N. Integrative Study on Chromosome Evolution of Mammals, Ants and Wasps Based on the Minimum Interaction Theory. J. Theor. Biol. 2001, 210, 475–497. [Google Scholar] [CrossRef]
- Li, S.-F.; Su, T.; Cheng, G.-Q.; Wang, B.-X.; Li, X.; Deng, C.-L.; Gao, W.-J. Chromosome Evolution in Connection with Repetitive Sequences and Epigenetics in Plants. Genes 2017, 8, 290. [Google Scholar] [CrossRef]
- Guerra, M. Patterns of heterochromatin distribution in plant chromosomes. Genet. Mol. Biol. 2000, 23, 1029–1041. [Google Scholar] [CrossRef]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold Publishers Ltd: London, UK, 1971. [Google Scholar]
- Moscone, E.; Scaldaferro, M.; Grabiele, M.; Cecchini, N.; García, Y.S.; Jarret, R.; Daviña, J.; Ducasse, D.; Barboza, G.; Ehrendorfer, F. The evolution of chili peppers (Capsicum-Solanaceae): A cytogenetic perspective. Acta Hortic. 2007, 745, 137–170. [Google Scholar] [CrossRef]
- Fonsêca, A.; Pedrosa-Harand, A. Karyotype stability in the genus Phaseolus evidenced by the comparative mapping of the wild species Phaseolus microcarpus. Genome 2013, 56, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Yuan, Q. The difference between the heterogeneity of the centromeric index and intrachromosomal asymmetry. Plant Syst. Evol. 2011, 297, 141–145. [Google Scholar] [CrossRef]
- Murat, F.; Zhang, R.; Guizard, S.; Gavranovic, H.; Flores, R.; Steinbach, D.; Quesneville, H.; Tannier, E.; Salse, J. Karyotype and gene order evolution from reconstructed extinct ancestors highlight contrasts in genome plasticity of modern rosid crops. Genome Biol. Evol. 2015, 7, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Tan, S.G. Inter-Simple Sequence Repeat (ISSR) Markers: Are We Doing It Right? ASM Sci. J. 2015, 9, 30–39. [Google Scholar]
- Hassan, N.; Mostafa, S.; Twfik, A. Assessment of genetic diversity of tomato (Lycopersicon esculentum L.) germplasm using molecular markers (RAPD and ISSR). Egypt. J. Genet. Cytol. 2013, 42, 163–182. [Google Scholar] [CrossRef]
- Meng, F.-J.; Xu, X.-Y.; Huang, F.-L.; Li, J.-F. Analysis of Genetic Diversity in Cultivated and Wild Tomato Varieties in Chinese Market by RAPD and SSR. Agric. Sci. China 2010, 9, 1430–1437. [Google Scholar] [CrossRef]
- Sharifova, S.S.; Mehdiyeva, S.P.; Abbasov, M.A. Analysis of genetic diversity among different tomato genotypes using ISSR DNA marker. Genetika 2017, 49, 31–42. [Google Scholar] [CrossRef]
- Shahlaei, A.; Torabi, S.; Khosroshahli, M. Efficiacy of SCoT and ISSR marekers in assesment of tomato (Lycopersicum esculentum Mill.) genetic diversity. Int. J. Biosci. 2014, 5, 14–22. [Google Scholar]
- Lin, K.; Lo, H.; Lee, S.-P.; Kuo, C.G.; Chen, J.-T.; Yeh, W.-L. RAPD markers for the identification of yield traits in tomatoes under heat stress via bulked segregant analysis. Hereditas 2006, 143, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Saha, S.; Bandyopadhyay, T.K.; Ghosh, P. Efficiency of ISSR marker for characterization of Cymbopogon germplasms and their suitability in molecular barcoding. Plant Syst. Evol. 2015, 301, 439–450. [Google Scholar] [CrossRef]
- El-Aziz, M.H.A.; Habiba, R.M.M. Molecular assessment of genetic diversity in some canola homozygous line. Egypt. J. Genet. Cytol. 2016, 45, 129–145. [Google Scholar] [CrossRef]
- El-Hadi, A.A.; El-Aziz, M.A.; Alla, M.A.; Ashak, M. Molecular and Phenotypic Evaluation of some Summer Squash Inbred Lines. J. Agric. Chem. Biotechnol. 2017, 8, 281–287. [Google Scholar] [CrossRef]
- El-Moneim, A. Characterization of ISSR and SCoT Markers and TaWRKY Gene Expression in some Egyptian Wheat Genotypes under Drought Stress. J. Plant Prod. Sci. 2019, 8, 31–46. Available online: https://jpps.journals.ekb.eg/article_72549.html (accessed on 27 December 2019).
- Nagy, E.; Gyulai, G.; Szabó, Z.; Hegyi, Z.; Marton, L.C. Application of morphological descriptions and genetic markers to analyse polymorphism and genetic relationships in maize (Zea mays L.). Acta Agron. Hung. 2003, 51, 257–265. [Google Scholar] [CrossRef]
- Srinivasan, T.; Nagarajan, P.; Senthil, N.; Gnanam, R.; Ponnuswami, V. DNA fingerprinting in Tomato (Lycopersicon esculentum Mill.) germplasm accessions using molecular markers. Int. J. Integr. Biol. 2013, 14, 86–95. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Chromosome Number (2n) | THL (µm) | CVCI | CVCL | MCA | ASK% | TF% | Syi Index | Rec Index | A1 | A2 | A | DI | AI | Stebb | KF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 24 | 375.22 | 6.19 | 18.52 | 8.87 | 54.44% | 45.56% | 83.69% | 65.74% | 0.16 | 0.19 | 0.09 | 8.43 | 1.15 | 1B | 18 nm + 6 m |
| G2 | 24 | 393.11 | 10.37 | 16.28 | 13.51 | 56.96% | 43.04% | 75.55% | 72.92% | 0.23 | 0.16 | 0.14 | 6.85 | 1.69 | 1A | 6 nsm(-) + 14 nm + 4 m |
| G3 | 24 | 323.08 | 6.38 | 22.22 | 13.16 | 56.48% | 43.52% | 77.04% | 64.99% | 0.23 | 0.22 | 0.13 | 10.04 | 1.42 | 1B | 22 nm + 2 m |
| G4 | 24 | 452.75 | 14.76 | 17.31 | 16.75 | 58.05% | 41.95% | 72.25% | 69.59% | 0.27 | 0.17 | 0.17 | 7.56 | 2.56 | 2B | 4 nsm(-) + 18 nm + 2 m |
| G5 | 26 | 464.48 | 9.77 | 22.49 | 12.81 | 56.54% | 43.46% | 76.88% | 63.61% | 0.22 | 0.22 | 0.13 | 9.79 | 2.20 | 2B | 2 nsm(-) + 18 nm + 6 nm |
| G6 | 24 | 425.13 | 9.6 | 22.54 | 9.96 | 54.89% | 45.11% | 82.19% | 65.80% | 0.17 | 0.23 | 0.10 | 10.91 | 2.16 | 1B | 2 nsm(-) + 12 nm + 10 nm |
| Marker | Primer | MB | PB | UB | TAB | FS | PIC | EMR | MI | P (%) | RP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| RAPD | OP-A3 | 7.00 | 0.00 | 0.00 | 7.00 | 190–865 | 0.00 | 0.00 | 0.00 | 0.00% | 14.0 |
| OP-A9 | 7.00 | 0.00 | 0.00 | 7.00 | 180–890 | 0.13 | 0.00 | 0.00 | 0.00% | 12.5 | |
| OP- B3 | 3.00 | 3.00 | 0.00 | 6.00 | 245–1075 | 0.28 | 1.50 | 0.42 | 50.00% | 8.70 | |
| OP-C9 | 3.00 | 0.00 | 0.00 | 3.00 | 300–700 | 0.00 | 0.00 | 0.00 | 0.00% | 6.00 | |
| OP-C15 | 4.00 | 3.00 | 2.00 | 7.00 | 375–1050 | 0.26 | 1.29 | 0.33 | 42.86% | 10.3 | |
| OP-K2 | 2.00 | 2.00 | 2.00 | 4.00 | 345–735 | 0.15 | 1.00 | 0.15 | 50.00% | 6.77 | |
| OP-K3 | 2.00 | 0.00 | 0.00 | 2.00 | 450–635 | 0.00 | 0.00 | 0.00 | 0.00% | 4.00 | |
| Ave. | 4.00 | 1.14 | 0.57 | 5.14 | 0.12 | 0.54 | 0.13 | 20.41% | 8.90 | ||
| SCoT | SCoT 2 | 3.00 | 3.00 | 2.00 | 6.00 | 200–530 | 0.31 | 1.50 | 0.47 | 50.00% | 8.33 |
| SCoT 4 | 4.00 | 0.00 | 0.00 | 4.00 | 120–430 | 0.00 | 0.00 | 0.00 | 0.00% | 8.00 | |
| SCoT 12 | 6.00 | 2.00 | 0.00 | 8.00 | 275–765 | 0.18 | 0.50 | 0.09 | 25.00% | 13.1 | |
| SCoT 13 | 3.00 | 2.00 | 0.00 | 5.00 | 390–840 | 0.28 | 0.80 | 0.22 | 40.00% | 7.16 | |
| SCoT 15 | 2.00 | 2.00 | 1.00 | 4.00 | 225–560 | 0.22 | 1.00 | 0.22 | 50.00% | 6.27 | |
| Ave. | 3.60 | 1.80 | 0.60 | 5.40 | 0.20 | 0.76 | 0.20 | 33.00% | 8.57 | ||
| ISSR | 49A | 1.00 | 3.00 | 0.00 | 4.00 | 245–630 | 0.58 | 2.25 | 1.31 | 75.00% | 3.33 |
| 49B | 2.00 | 4.00 | 1.00 | 6.00 | 170–940 | 0.44 | 2.67 | 1.17 | 66.67% | 6.72 | |
| 89B | 1.00 | 2.00 | 0.00 | 3.00 | 380–700 | 0.59 | 1.33 | 0.79 | 66.67% | 2.44 | |
| HB-8 | 3.00 | 3.00 | 1.00 | 6.00 | 370–1290 | 0.32 | 1.50 | 0.48 | 50.00% | 8.16 | |
| HB-9 | 6.00 | 1.00 | 1.00 | 7.00 | 270–940 | 0.04 | 0.14 | 0.01 | 14.29% | 13.4 | |
| HB-14 | 1.00 | 3.00 | 3.00 | 4.00 | 280–860 | 0.23 | 2.25 | 0.52 | 75.00% | 6.16 | |
| HB-15 | 3.00 | 2.00 | 0.00 | 5.00 | 240–540 | 0.29 | 0.80 | 0.23 | 40.00% | 7.11 | |
| Ave. | 2.43 | 2.57 | 0.86 | 5.00 | 0.36 | 1.56 | 0.64 | 55.37% | 6.76 |
| Parameter | RAPD | SCoT | ISSR | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total bands | 36 | 27 | 35 | |||||||||
| Monomorphic bands | 17 | 18 | 17 | |||||||||
| Polymorphic bands | 8 | 9 | 18 | |||||||||
| Unique bands | No. of unique bands | 4 | 3 | 6 | ||||||||
| Positive bands | 670 (G5) | 530 (G2) | - | |||||||||
| Primers | OPC15 | SCoT2 | - | |||||||||
| Negative bands | 720 (G2) | 400 (G6) | 735 (G6) | 260 (G2) | 500 (G1) | 475 (G1) | 700 (G1) | 280 (G4) | 345 (G4) | 860 (G5) | 940 (G6) | |
| Primers | OPC15 | OPK2 | SCoT2 | SCoT15 | 49-B | HB-8 | HB-14 | HB-9 | ||||
| % Polymorphism | 22% | 33.33% | 51.43% | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
EL-Mansy, A.B.; Abd El-Moneim, D.; ALshamrani, S.M.; Safhi, F.A.; Abdein, M.A.; Ibrahim, A.A. Genetic Diversity Analysis of Tomato (Solanum lycopersicum L.) with Morphological, Cytological, and Molecular Markers under Heat Stress. Horticulturae 2021, 7, 65. https://doi.org/10.3390/horticulturae7040065
EL-Mansy AB, Abd El-Moneim D, ALshamrani SM, Safhi FA, Abdein MA, Ibrahim AA. Genetic Diversity Analysis of Tomato (Solanum lycopersicum L.) with Morphological, Cytological, and Molecular Markers under Heat Stress. Horticulturae. 2021; 7(4):65. https://doi.org/10.3390/horticulturae7040065
Chicago/Turabian StyleEL-Mansy, Ahmed B., Diaa Abd El-Moneim, Salha Mesfer ALshamrani, Fatmah Ahmed Safhi, Mohamed A. Abdein, and Amira A. Ibrahim. 2021. "Genetic Diversity Analysis of Tomato (Solanum lycopersicum L.) with Morphological, Cytological, and Molecular Markers under Heat Stress" Horticulturae 7, no. 4: 65. https://doi.org/10.3390/horticulturae7040065