
Microbial Polymers as Sustainable Agents for Mitigating Health Risks of Plant-Based Endocrine Disruptors in Surface Water


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Biopolymer Production
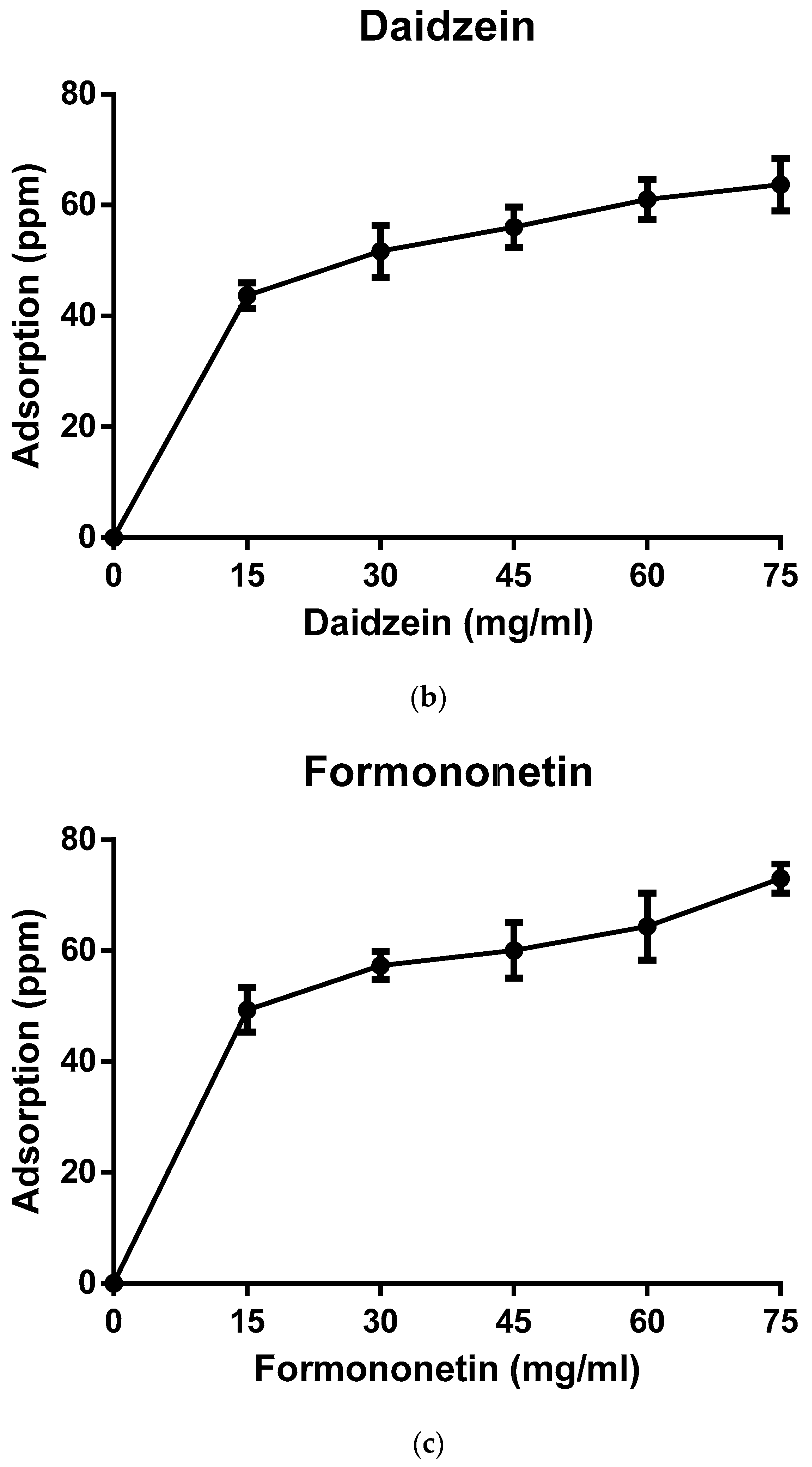
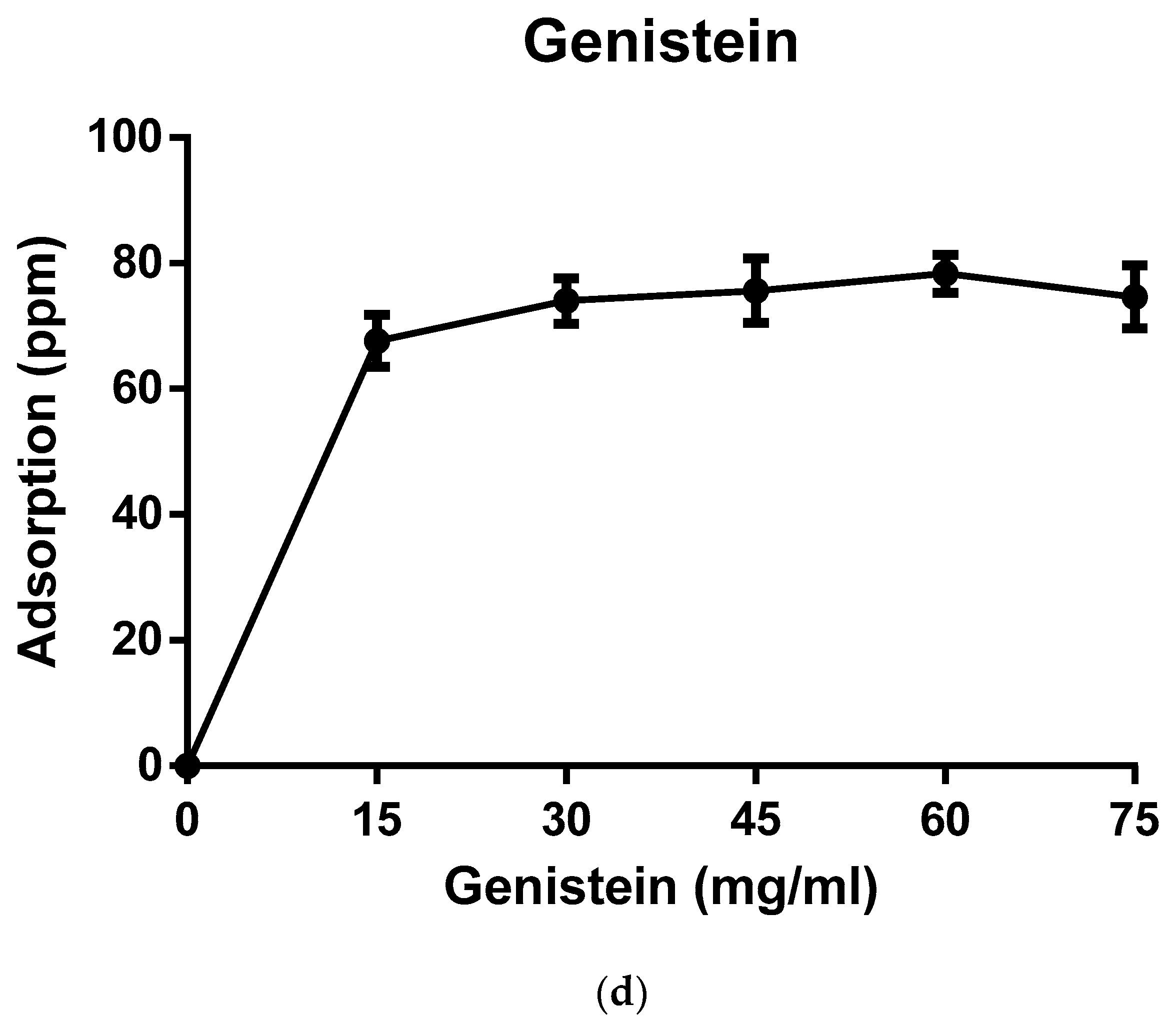
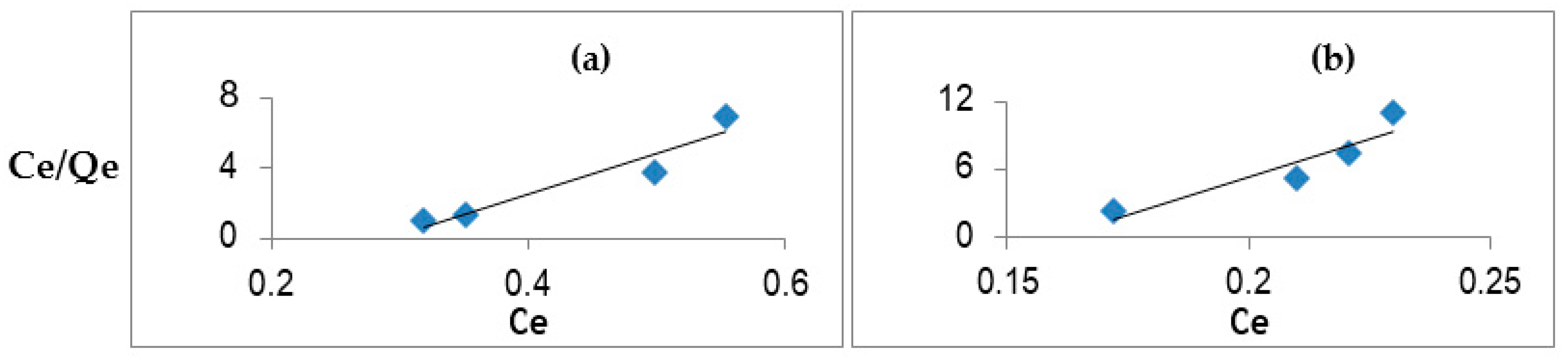
2.2. Biopolymer Characterization and Adsorption Studies
2.3. Biopolymer Toxicity
2.4. C. elegans Culture
2.4.1. Dosage Fixation and Survival Assay
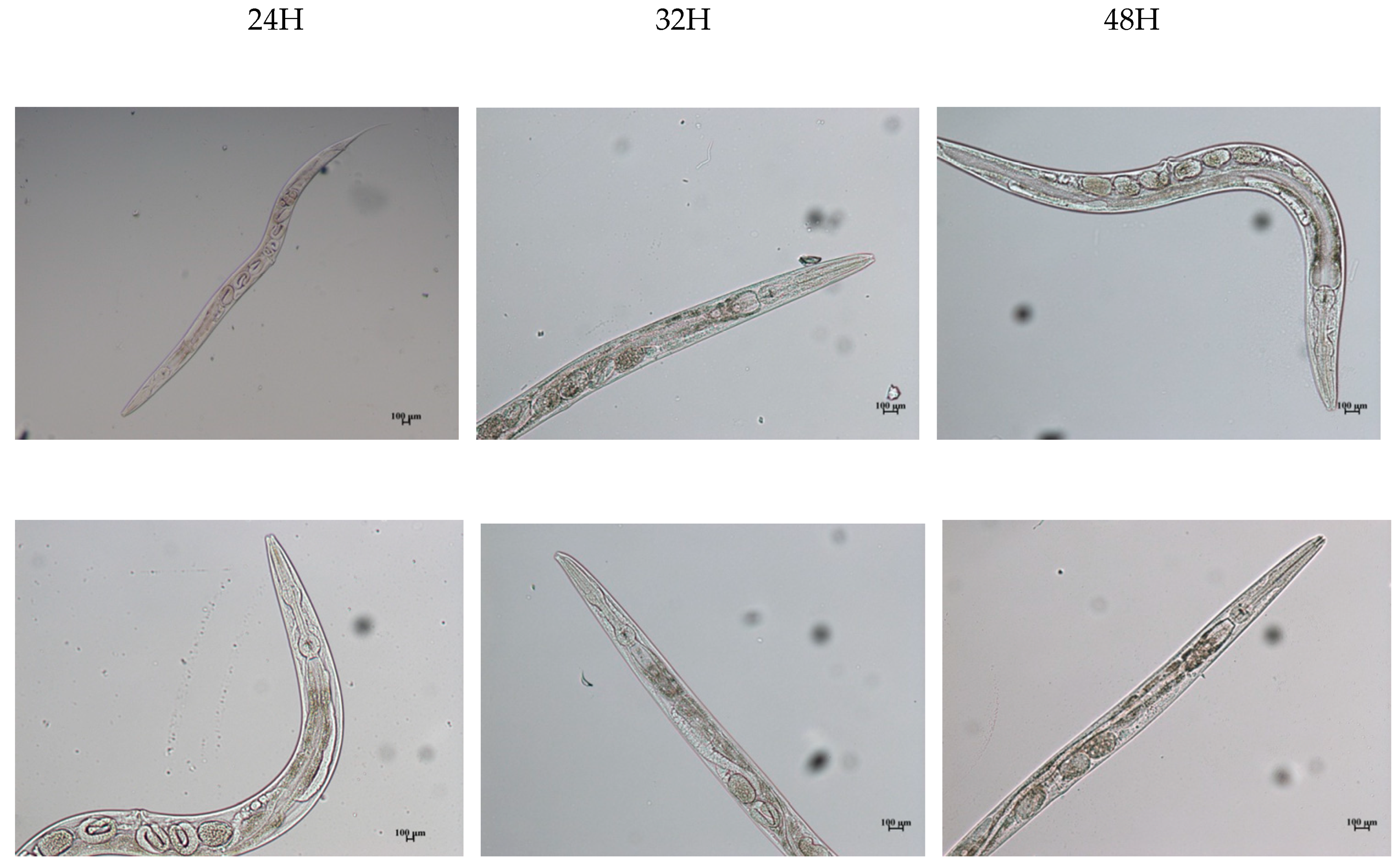
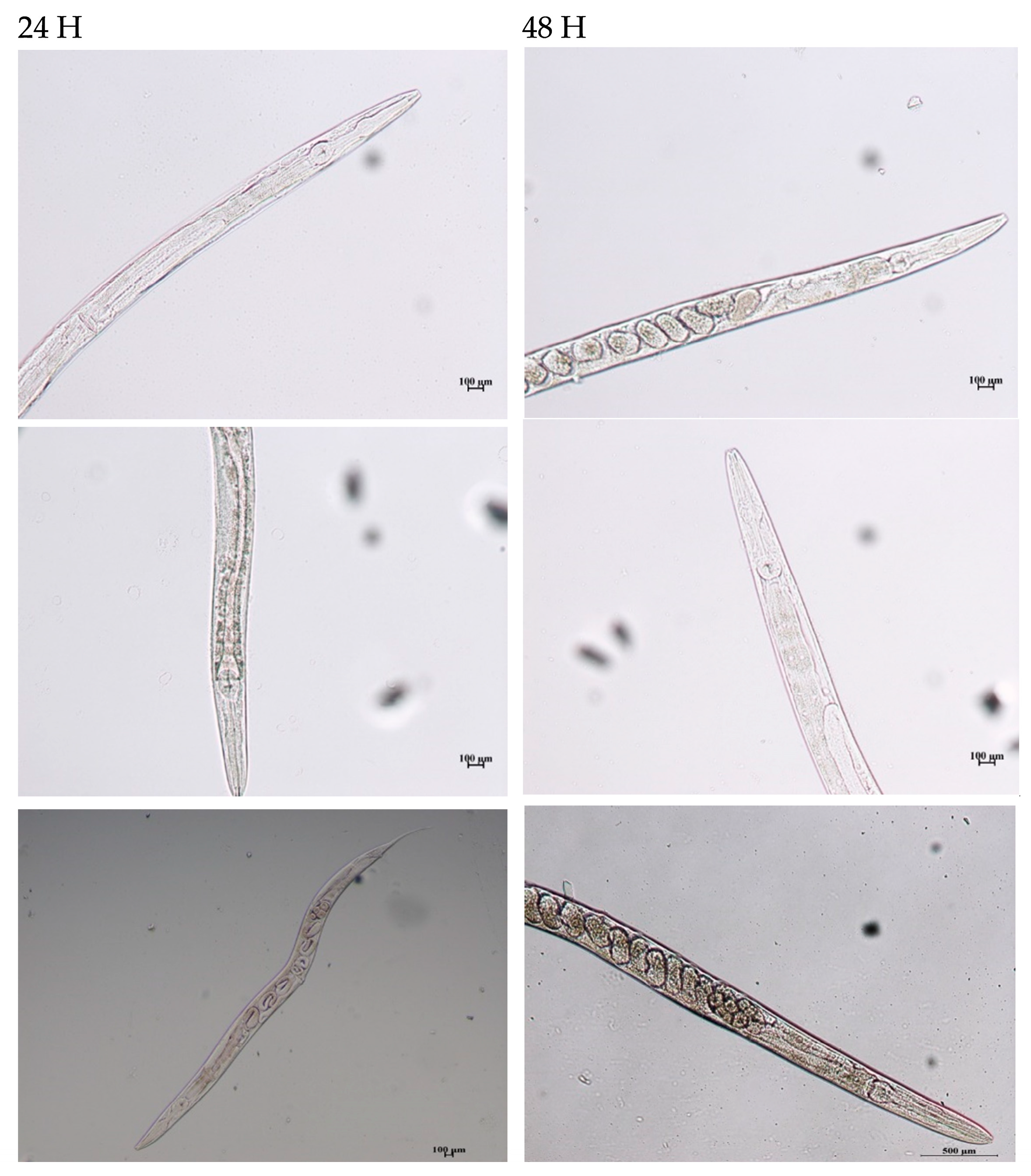
2.4.2. In Vivo Assessment and Microscopic Analysis of C. elegans Treated with Biopolymer
2.4.3. In Vivo and Microscopic Analysis of Combinatorial Effects on C. elegans Treated with Phytoestrogen Followed by Biopolymer & Vice Versa
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocrinol. 2010, 31, 400–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crain, D.A.; Janssen, S.J.; Edwards, T.M.; Heindel, J.; Ho, S.-M.; Hunt, P.; Iguchi, T.; Juul, A.; McLachlan, J.A.; Schwartz, J. Female reproductive disorders: The roles of endocrine-disrupting compounds and developmental timing. Fertil. Steril. 2008, 90, 911–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, T.J.; Briggs, M.A.; Phillips, P.J.; Blazer, V.S.; Smalling, K.L.; Kolpin, D.W.; Wagner, T. Groundwater discharges as a source of phytoestrogens and other agriculturally derived contaminants to streams. Sci. Total Environ. 2021, 755, 142873. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, T.; Iwasaki, S.; Kawashima, M.; Shinohara, O.; Abe, I. Adsorbability of estrone and 17β-estradiol in water onto activated carbon. Water Res. 2006, 40, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Urase, T.; Kikuta, T. Separate estimation of adsorption and degradation of pharmaceutical substances and estrogens in the activated sludge process. Water Res. 2005, 39, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, D.J.; Mastrocco, F.; Nowak, E.; Johnston, J.; Yekel, H.; Pfeiffer, D.; Hoyt, M.; DuPlessie, B.M.; Anderson, P.D. An assessment of potential exposure and risk from estrogens in drinking water. Environ. Health Perspect. 2010, 118, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Khaira, G.K.; Ganguli, A.; Ghosh, M. Antimicrobial efficacy and in vivo toxicity studies of a quaternized biopolymeric flocculant. J. Water Health 2014, 12, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Ghosh, M. Characterization of immunomodulatory, anticancer and antioxidant properties of an extracellular polymer produced by Enterococcus sp. in vegetable waste medium. Environ. Sustain. 2021, 4, 419–428. [Google Scholar] [CrossRef]
- Zdunczyk, Z.; Juskiewicz, J.; Frejnagel, S.; Wroblewska, M.; Krefft, B.; Gulewicz, K. Influence of Oligosaccharides from Lupin Seeds and Fructooligosaccharides on Utilisation of Protein by Rats and Absorption of Nutrients in the Small Intestine. In Proceedings of the 9th International Lupin Conference, Klink, Germany, 20–24 June 1999. [Google Scholar]
- Sehgal, S. Surface Assimilation of Phytoestrogens by Microbial Biopolymers. Master’s Thesis, Thapar Institute of Engineering and Technology, Patiala, India, 2013. [Google Scholar]
- Stiernagle, T. Maintenance of C. elegans WormBook. The C. elegans research community. In WormBook; University of Minnesota: Minneapolis, MN, USA, 2006. [Google Scholar]
- Harlow, P.H.; Perry, S.J.; Widdison, S.; Daniels, S.; Bondo, E.; Lamberth, C.; Currie, R.A.; Flemming, A.J. The nematode Caenorhabditis elegans as a tool to predict chemical activity on mammalian development and identify mechanisms influencing toxicological outcome. Sci. Rep. 2016, 6, 22965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Ghosh, M. Effective removal of Bisphenol A from plastic waste leachates by microbial polymer impregnated with activated carbon. Int. J. Environ. Sci. Technol. 2020, 17, 1907–1916. [Google Scholar] [CrossRef]
- Jahromi, F.A.; Moore, F.; Keshavarzi, B.; Mohebbi-Nozar, S.L.; Mohammadi, Z.; Sorooshian, A.; Abbasi, S. Bisphenol A (BPA) and polycyclic aromatic hydrocarbons (PAHs) in the surface sediment and bivalves from Hormozgan Province coastline in the Northern Persian Gulf: A focus on source apportionment. Mar. Pollut. Bull. 2020, 152, 110941. [Google Scholar] [CrossRef] [PubMed]
- Gabet-Giraud, V.; Miege, C.; Choubert, J.; Ruel, S.M.; Coquery, M. Occurrence and removal of estrogens and beta blockers by various processes in wastewater treatment plants. Sci. Total Environ. 2010, 408, 4257–4269. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.R.; Olejnik, N.; Sprando, R.L. Toxicity ranking of heavy metals with screening method using adult Caenorhabditis elegans and propidium iodide replicates toxicity ranking in rat. Food Chem. Toxicol. 2012, 50, 3280–3290. [Google Scholar] [CrossRef] [PubMed]
- Kamaladevi, A.; Balamurugan, K. Role of PMK-1/p38 MAPK defense in Caenorhabditis elegans against Klebsiella pneumoniae infection during host-pathogen interaction. Pathog. Dis. 2015, 73, ftv021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundgren, M.S.; Novak, P.J. Quantification of phytoestrogens in industrial waste streams. Environ. Toxicol. Chem. Int. J. 2009, 28, 2318–2323. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.V.; Rallapalli, P.B.S.; Dangi, G.P.; Tayade, R.J.; Somani, R.S.; Bajaj, H.C. MIL-53 (Al): An efficient adsorbent for the removal of nitrobenzene from aqueous solutions. Ind. Eng. Chem. Res. 2011, 50, 10516–10524. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biopolymer Producing Microorganisms | Biopolymer–PE Binding (%) | Isolation Source |
|---|---|---|
| Enterococcus hirae MG6 | F(71%), G(72%), BCA(69%), D(87%) | Agro processing industry sludge |
| Bacillus cereus | F(54%), G(43%), BCA(25%), D(26%) | Legume farm soil |
| Citrobacter brakii MG303 | F(14%), G(23%), BCA(45%), D(56%) | Alfalfa farm soil |
| Bacillus cytotoxicus MG303 | F(54%), G(43%), BCA(35%), D(26%) | Soya processing industry effluent |
| Enterococcus vikkiensis | F(64%), G(53%), BCA(35%), D(81%) | Paper industry sludge |
| Phytoestrogens | Concentration | Qmax | KL | R2 | RL |
|---|---|---|---|---|---|
| Formononetin | 1.5 mg/mL | 0.10 | 0.16 | 0.95 | 0.96 |
| Genistein | 0.32 | 0.63 | 0.97 | 0.62 | |
| Biochanin A | 0.04 | 0.29 | 0.97 | 0.86 | |
| Daidzein | 1 mg/mL | 0.14 | 0.32 | 0.91 | 0.84 |
| (a) Dose of Biopolymer (µg/mL) | Cell Viability (%) |
|---|---|
| 0 | 99.6 ± 4.3 |
| 5 | 98.4 ± 6.4 |
| 10 | 98 ± 7.3 |
| 15 | 97.9 ± 7.4 |
| 20 | 98.3 ± 11.2 |
| 25 | 98.0 ± 9.7 |
| 50 | 97.8 ± 8.8 |
| 75 | 97.2 ± 9.3 |
| 100 | 96.7 ± 5.6 |
| 150 | 95.8 ± 4.3 |
| 200 | 96.3 ± 6.6 |
| 250 | 97.5 ± 9.6 |
| (b) Phytoestrogens Tested | LC50 (C.elegens) |
| Daidzein (D) | 100 µg/mL |
| Geneistein (G) | 83 µg/mL |
| BiochaninA (BCA) | 67 µg/mL |
| Formononetin (F) | 53 µg/mL |
| Vehicle Control | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, M.; Balamurugan, K.; Sharma, V. Microbial Polymers as Sustainable Agents for Mitigating Health Risks of Plant-Based Endocrine Disruptors in Surface Water. Int. J. Environ. Res. Public Health 2021, 18, 10040. https://doi.org/10.3390/ijerph181910040
Ghosh M, Balamurugan K, Sharma V. Microbial Polymers as Sustainable Agents for Mitigating Health Risks of Plant-Based Endocrine Disruptors in Surface Water. International Journal of Environmental Research and Public Health. 2021; 18(19):10040. https://doi.org/10.3390/ijerph181910040
Chicago/Turabian StyleGhosh, Moushumi, Krishnaswamy Balamurugan, and Vivek Sharma. 2021. "Microbial Polymers as Sustainable Agents for Mitigating Health Risks of Plant-Based Endocrine Disruptors in Surface Water" International Journal of Environmental Research and Public Health 18, no. 19: 10040. https://doi.org/10.3390/ijerph181910040
APA StyleGhosh, M., Balamurugan, K., & Sharma, V. (2021). Microbial Polymers as Sustainable Agents for Mitigating Health Risks of Plant-Based Endocrine Disruptors in Surface Water. International Journal of Environmental Research and Public Health, 18(19), 10040. https://doi.org/10.3390/ijerph181910040