
Biological and Cytoprotective Effect of Piper kadsura Ohwi against Hydrogen-Peroxide-Induced Oxidative Stress in Human SW1353 Cells


Abstract
:1. Introduction
2. Results
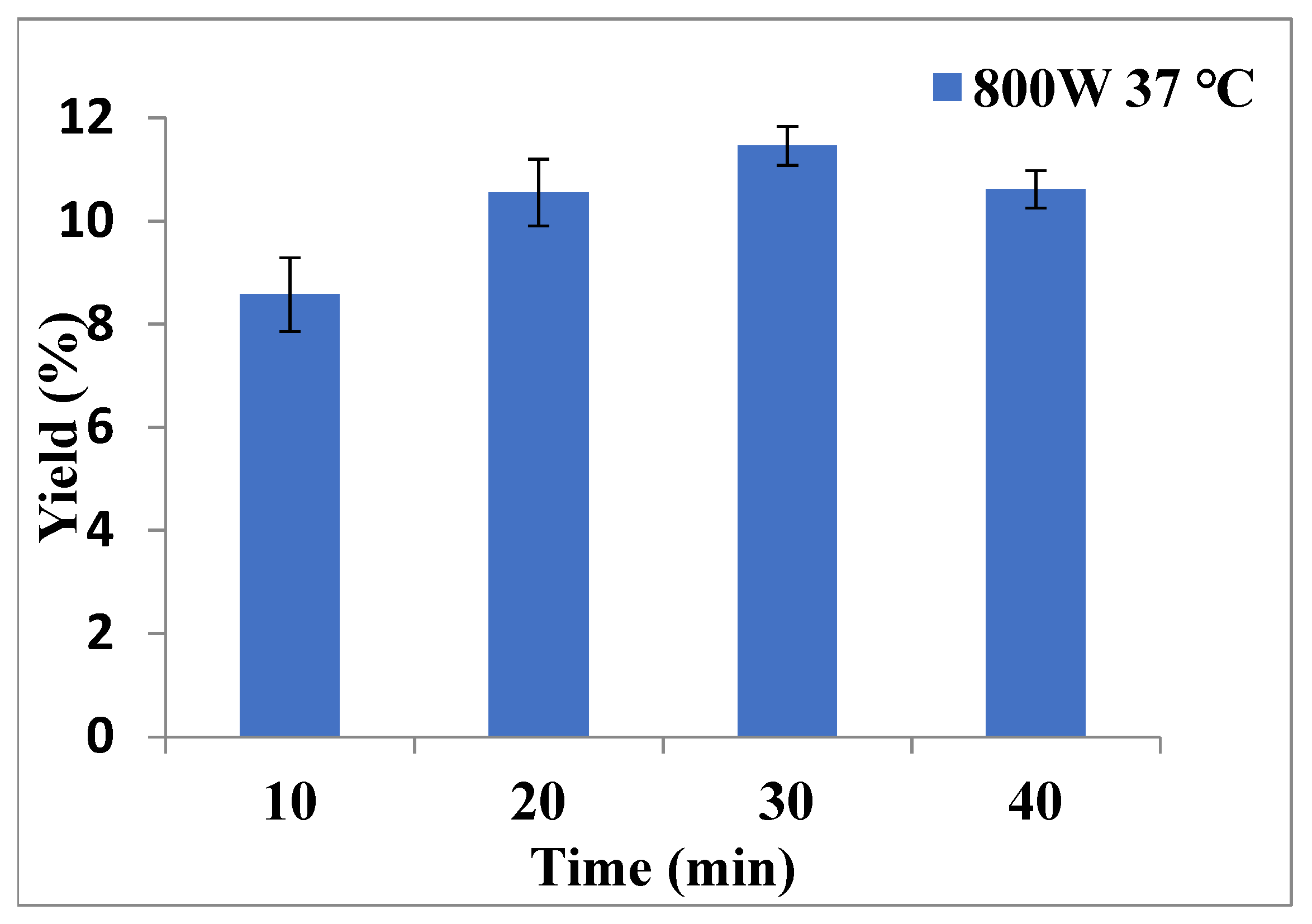
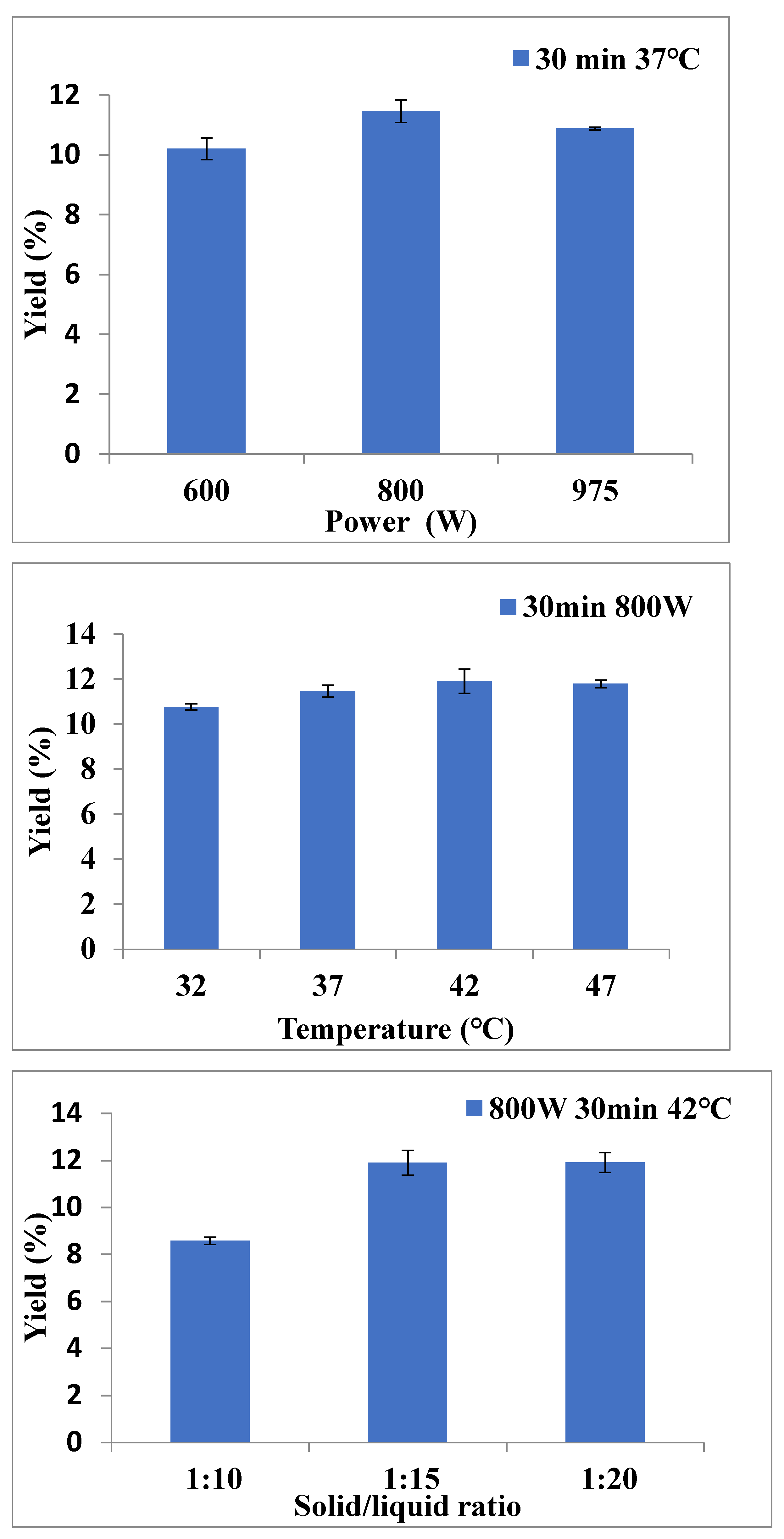
2.1. The Optimal Extraction Parameters for P. kadsura by MAE
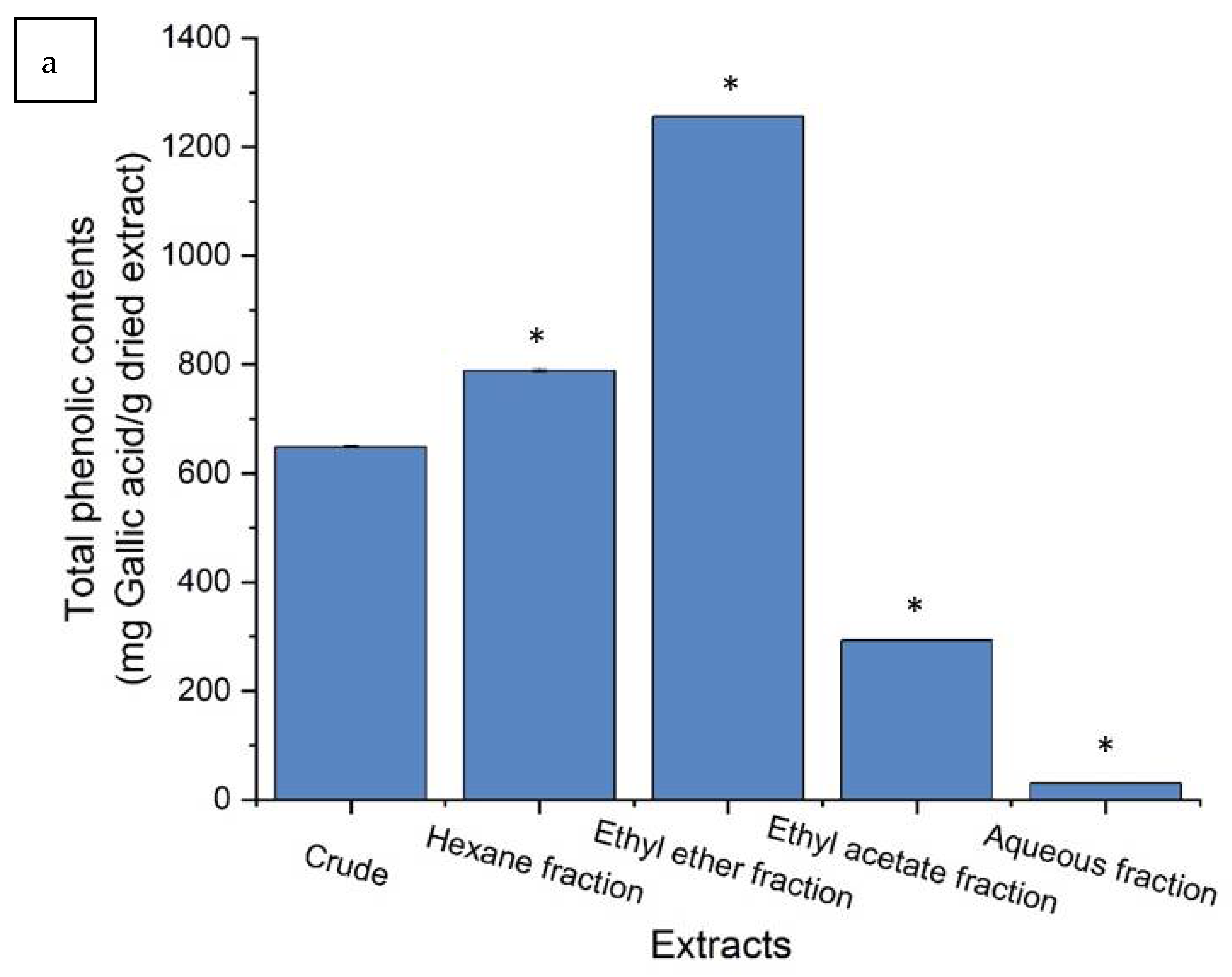
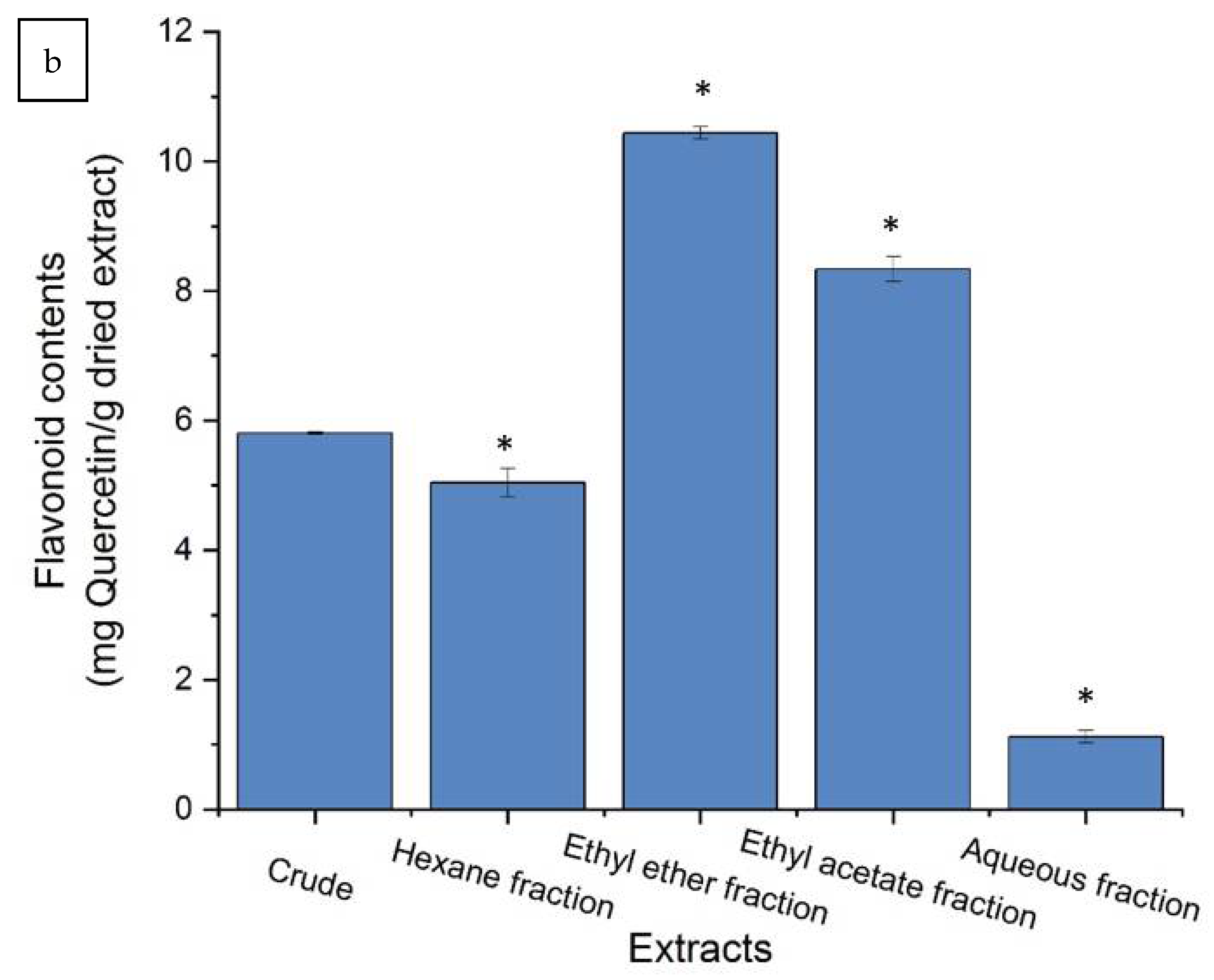
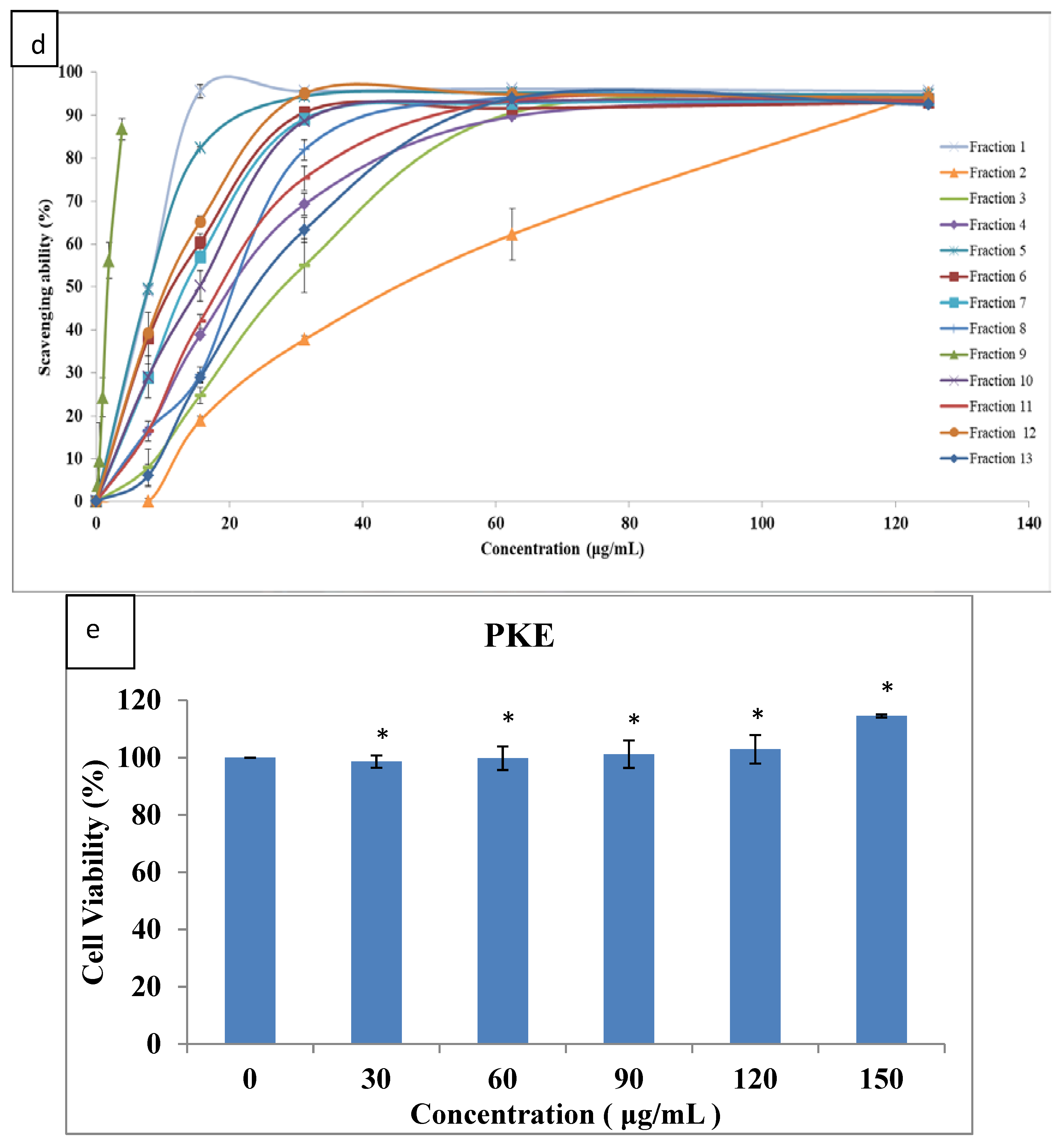
2.2. Purification of Crude Extract and Biological Characteristics
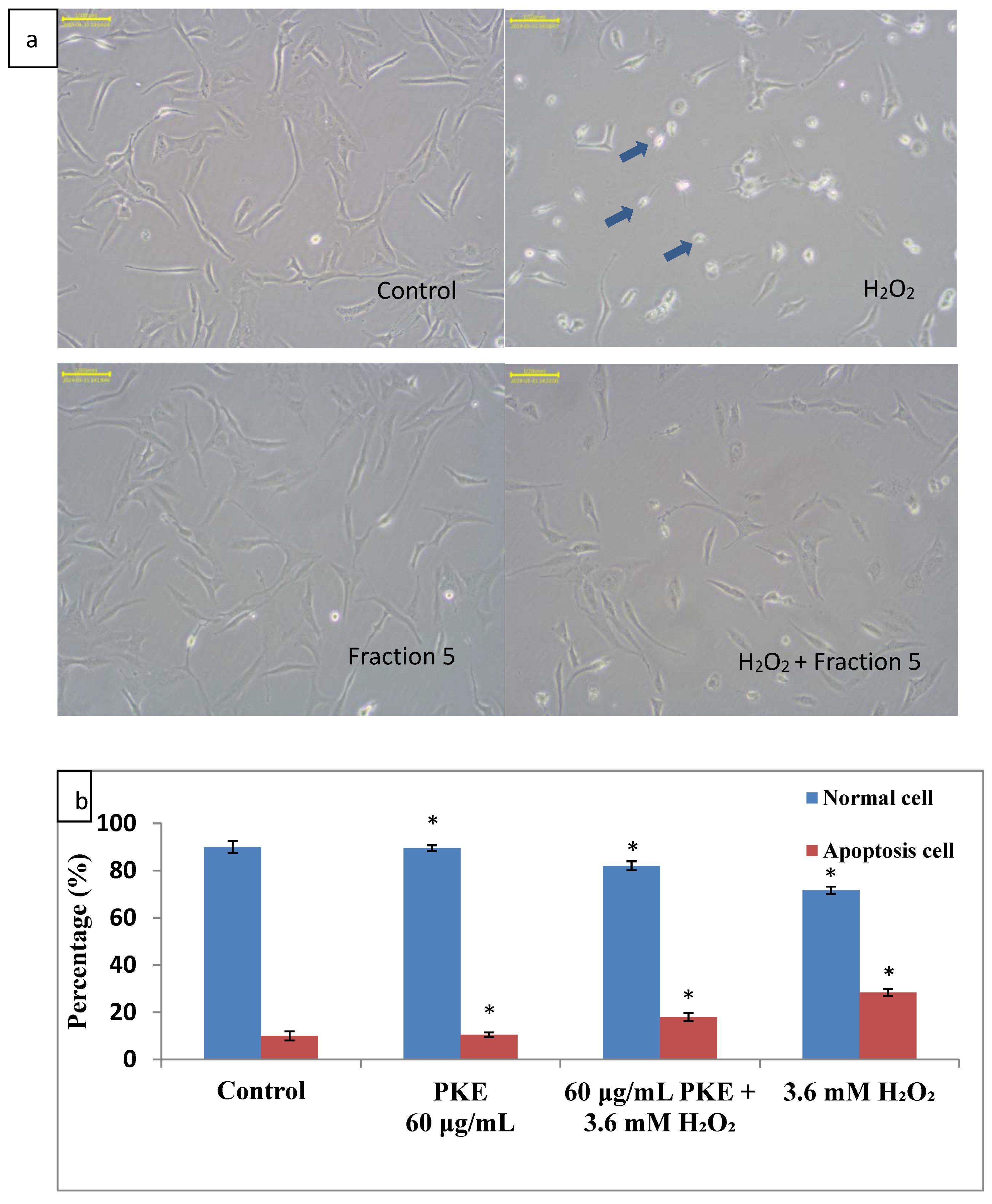
2.3. Apoptosis Assay and Cell Morphology Change
2.4. ROS Assay
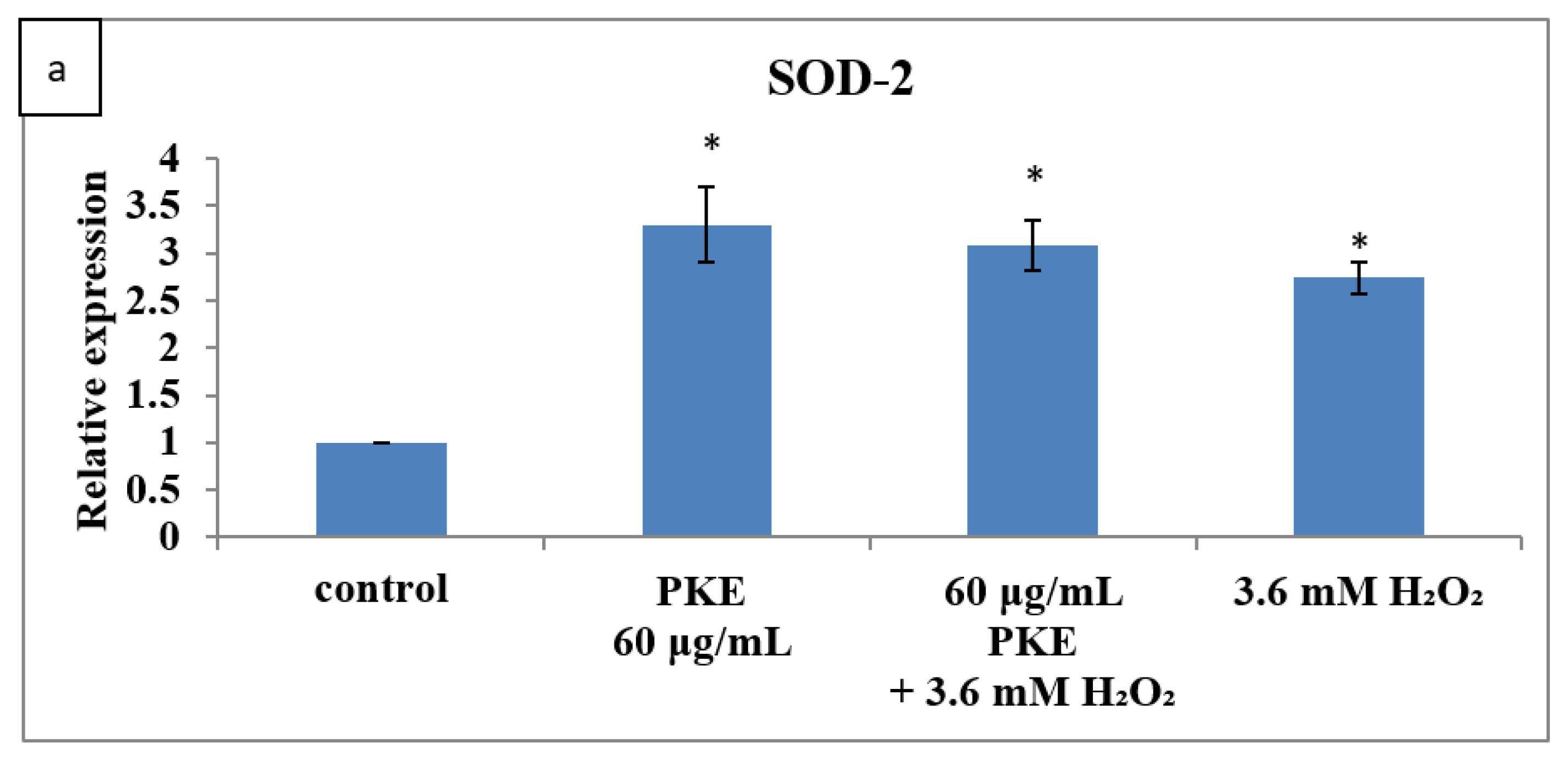
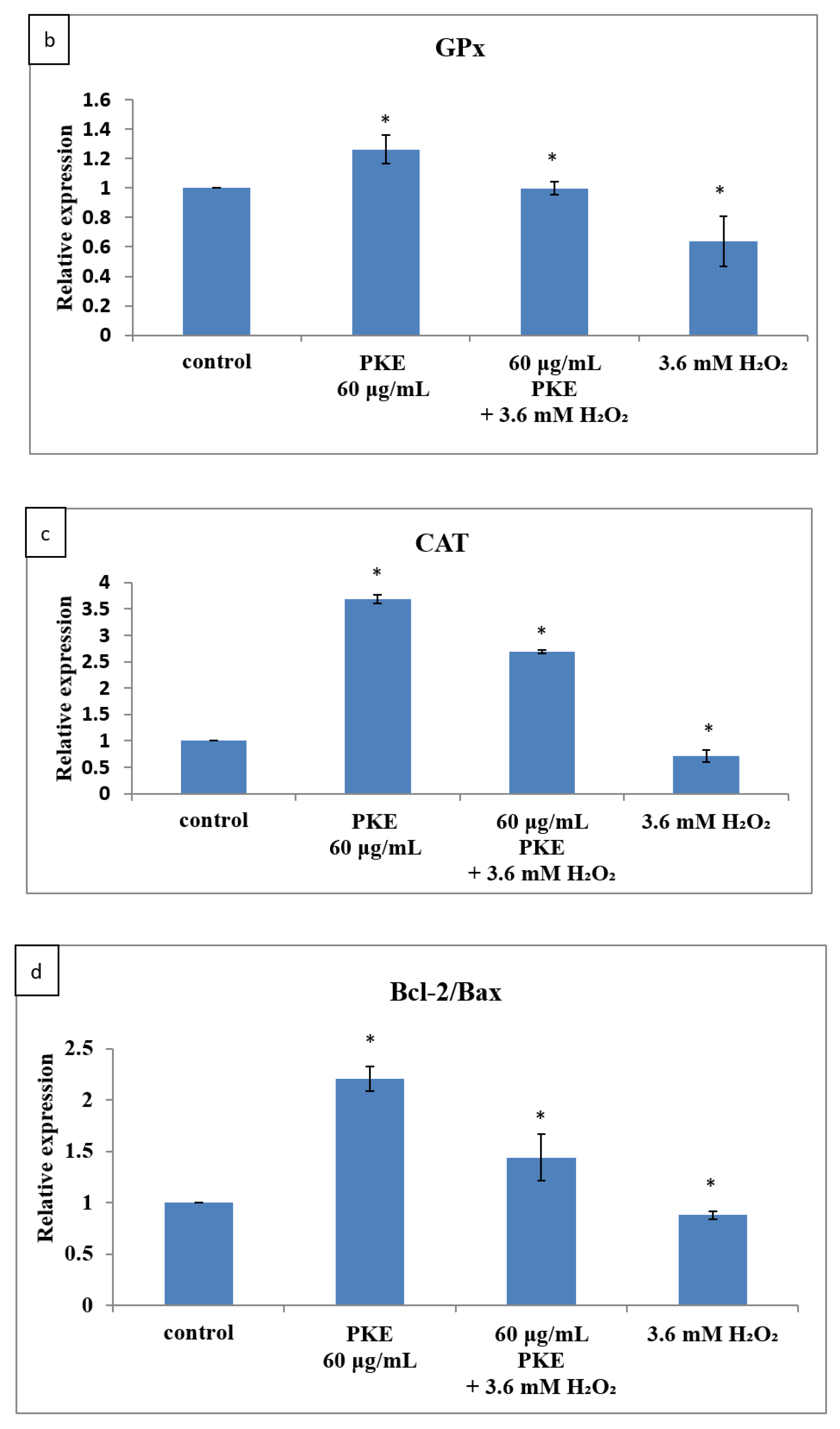
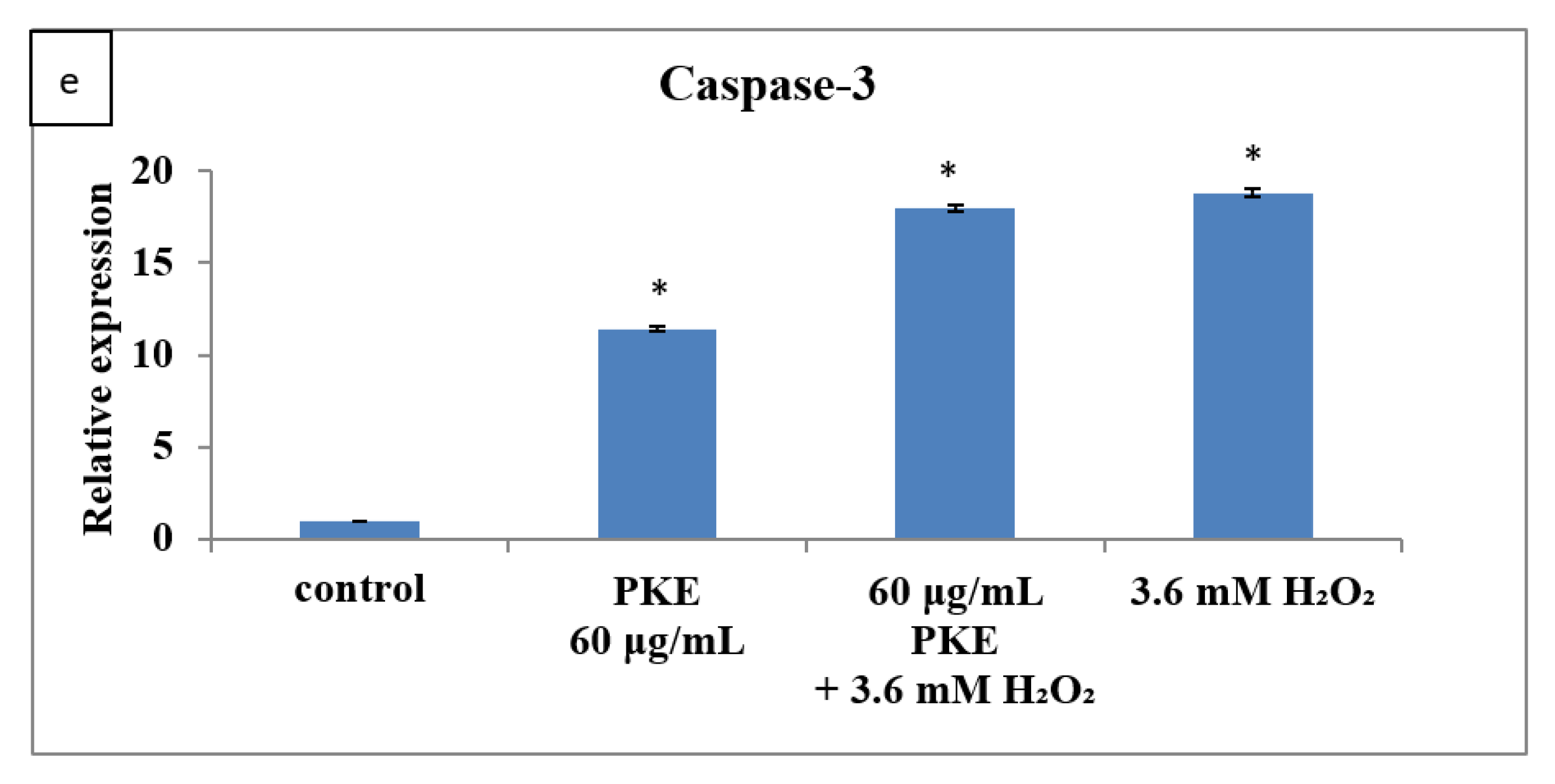
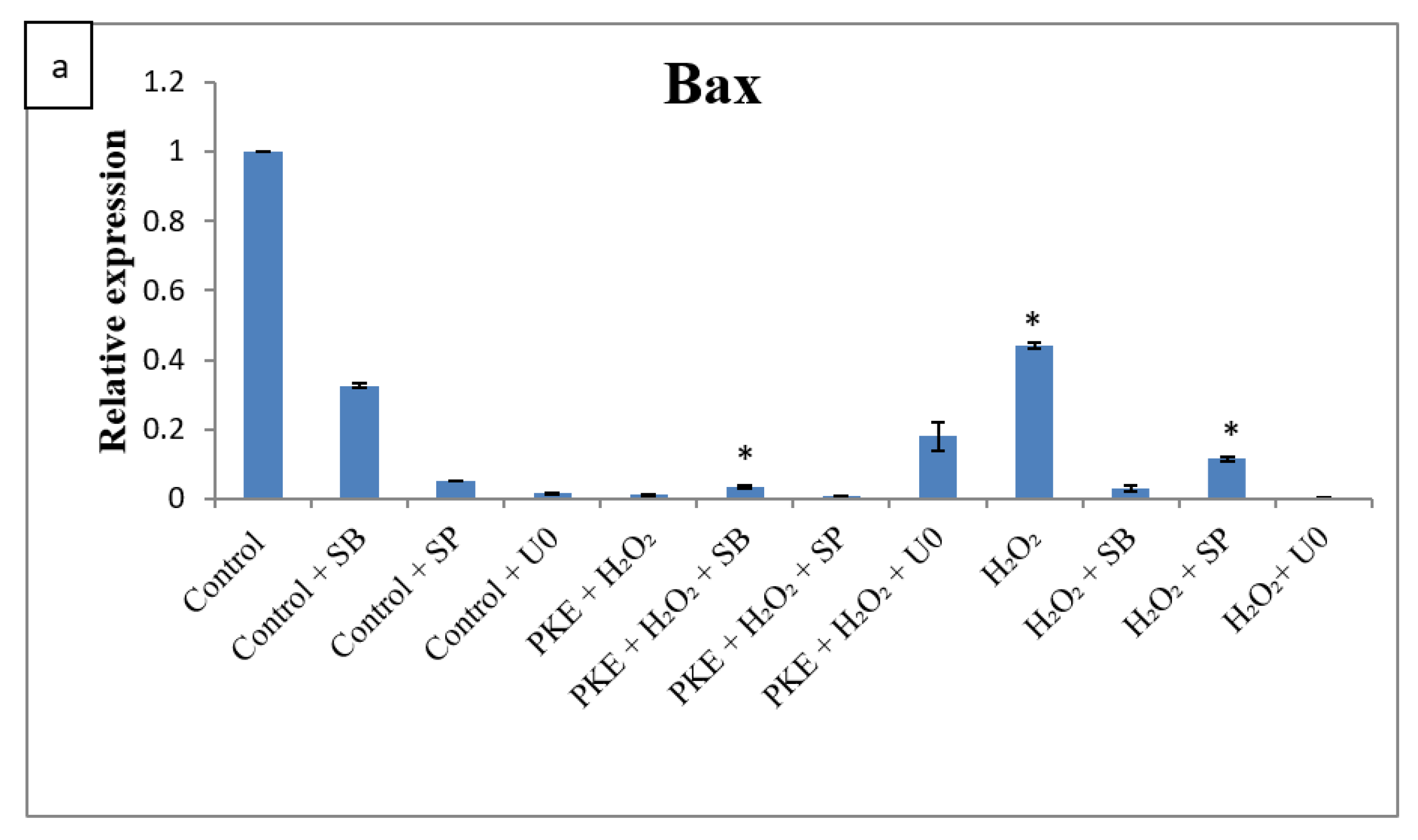
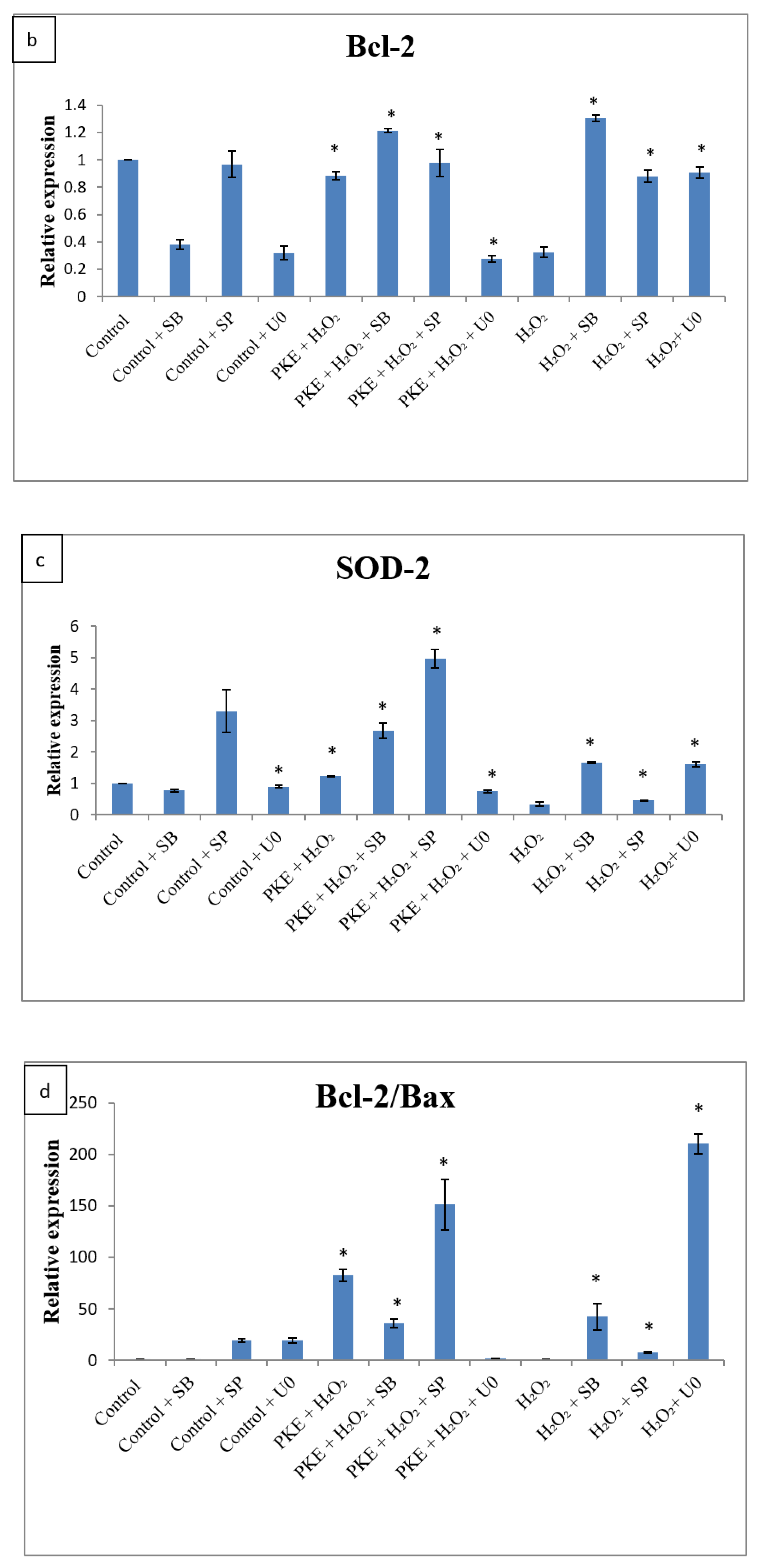
2.5. qRT-PCR
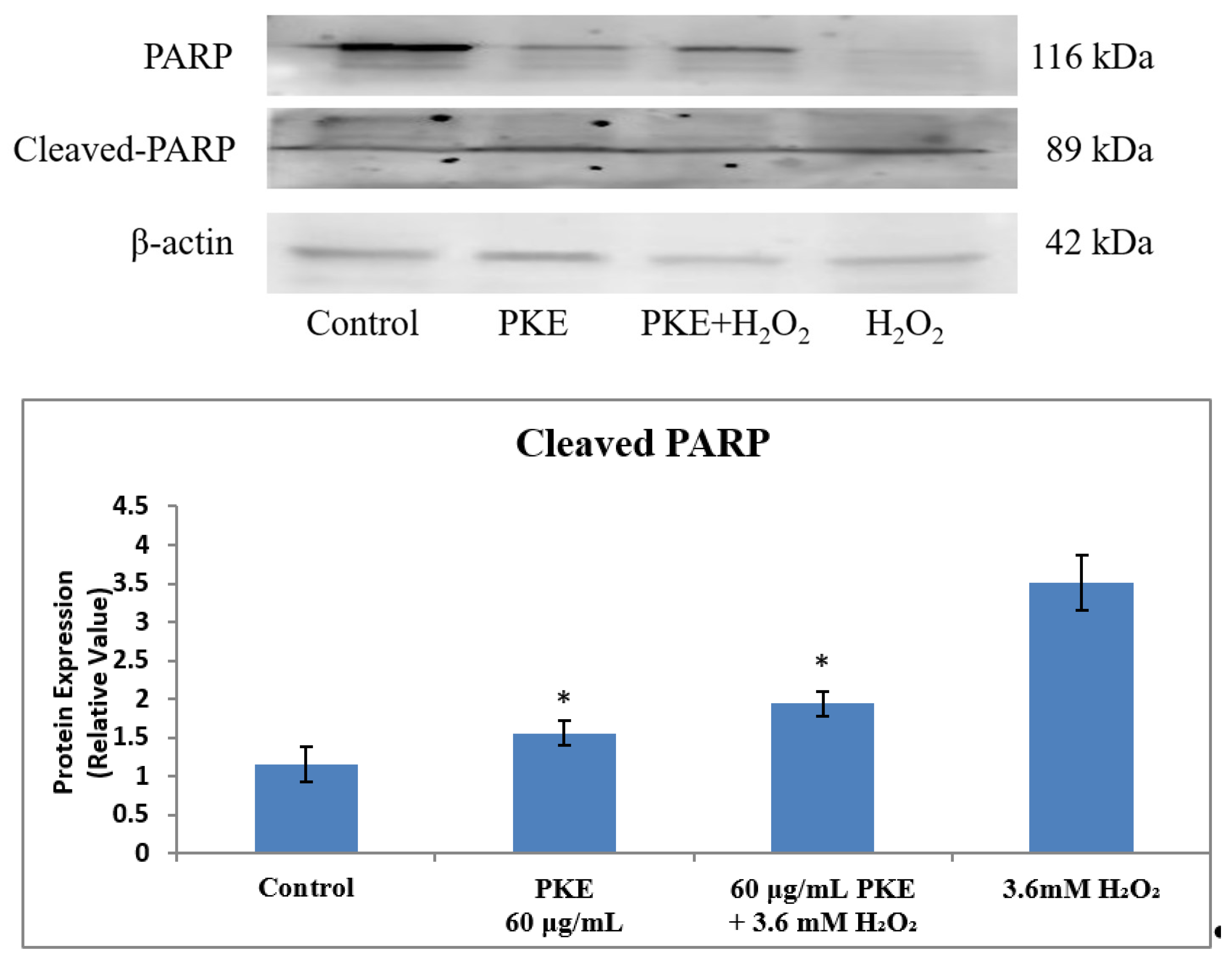
2.6. Western Blot
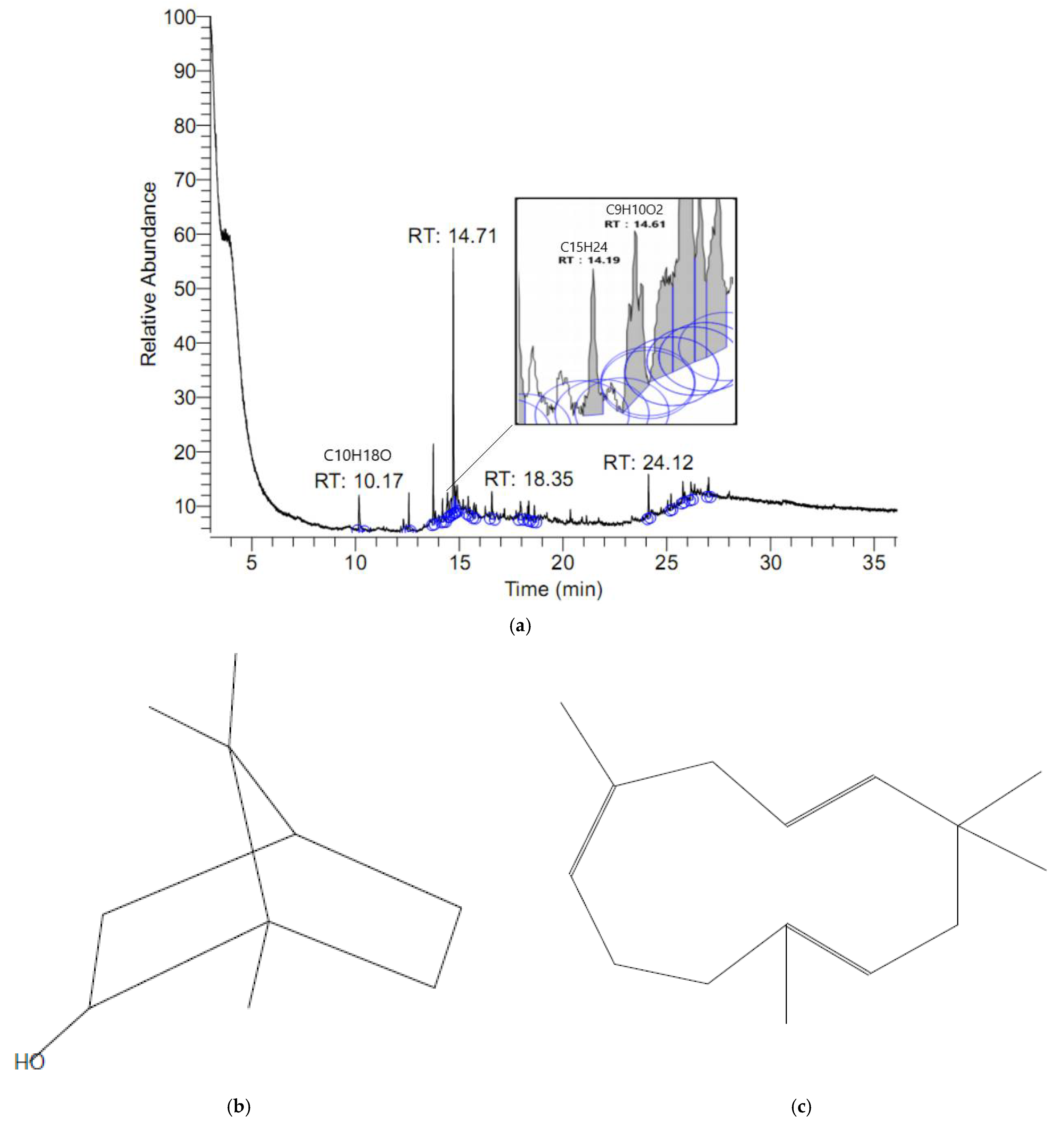
2.7. Composition Identification
3. Materials and Methods
3.1. Preparation of the Extracts of P. kadsura
3.2. Biological Characteristics of P. kadsura Extract
3.2.1. Total Polyphenols and Flavonoid Contents Determination
3.2.2. Antimicrobial Activity Assay—Disc Diffusion Method
3.2.3. Antioxidant Activity Analysis—DPPH Free Radical Scavenging Activity
3.3. Antioxidative Activity In Vitro
3.3.1. Cell Survival Assay and Cell Morphology
3.3.2. Apoptosis Assay
3.3.3. ROS Assay
3.3.4. Quantitative Real-Time PCR
3.3.5. Western Blotting
3.4. Composition Identification
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Na, J.Y.; Song, K.; Kim, S.; Kwon, J. Rutin protects rat articular chondrocytes against oxidative stress induced by hydrogen peroxide through SIRT1 activation. Biochem. Biophys. Res. Commun. 2016, 473, 1301–1308. [Google Scholar] [CrossRef]
- Cheleschi, S.; Da Palma, A.; Pascarelli, N.A.; Giordano, N.; Galeazzi, M.; Tenti, S.; Fioravanti, A. Could oxidative stress regulate the expression of microRNA-146a and microRNA-34a in human osteoarthritic chondrocyte cultures? Int. J. Mol. Sci. 2017, 18, 2660. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.C.; Grong, L.F.; Li, X.B.; Fu, X.; Xuan, J.W.; Feng, Z.H.; Ni, W.F. Inhibition of PI3K/Akt/NF-κB signaling with leonurine for ameliorating the progression of osteoarthritis: In vitro and in vivo studies. J. Cell. Physiol. 2018, 234, 6940–6950. [Google Scholar] [CrossRef]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squardrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Young, I.C.; Chuang, S.T.; Hsu, C.H.; Sun, Y.J.; Lin, F.H. C-phycocyanin alleviates osteoarthritic injury in chondrocytes stimulated with H2O2 and compressive stress. Int. J. Biol. Macromol. 2016, 93, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, K.V.; Papavassiliou, A.G. Redox and NF-κB signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, M.; Saas, J.; Sohler, F.; Haag, J.; So, S.; Pieper, M.; Bartnik, E.; Beninga, J.; Zimmer, R.; Aigner, T. Comparison of the chondrosarcoma cell line SW1353 with primaryhuman adult articular chondrocytes with regard to their gene expression profile and reactivity to IL-1b. Osteoarthr. Cartil. 2005, 13, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Hong, S.H.; Shin, S.S.; Lee, D.S.; Han, M.H.; Cha, H.J.; Kim, S.; Kim, H.S.; Kim, G.Y.; Park, E.K.; et al. Activation of the Nrf2/HO-1 signaling pathway contributes to the protective effects of sargassum serratifolium extract against oxidative stress-induced DNA damage and apoptosis in SW1353 human chondrocytes. Int. J. Environ. Res. Public Health 2018, 15, 1173. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Zhang, Y.; Dai, B.L.; Ma, Y.J.; Zhang, Q.; Wang, Y.; Yang, H. Chlorogenic acid prevents inflammatory responses in IL-1β-stimulated human SW-1353 chondrocytes, a model for osteoarthritis. Mol. Med. Rep. 2017, 16, 1369–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.C.; Shen, C.C.; Shen, Y.C.; Tsai, T.H. Anti-inflammatory Neolignans from Piper kadsura. J. Nat. Prod. 2006, 69, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.H.; Choi, W.; Ha, S.K.; Kim, S.Y.; Lee, K.R. Neolignans from Piper kadsura and their anti-neuroinflammatory activity. Bioorg. Med. Chem. Lett. 2010, 20, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; Lin, G.D.; Myers, S.P.; Leach, D.N. Anti-inflammatory activity of Chinese medicinal vine plants. J. Ethnopharmacol. 2003, 85, 61–67. [Google Scholar] [CrossRef]
- Chiou, W.F.; Peng, C.H.; Chen, C.F.; Chou, C.J. Anti-inflammatory properties of piperlactam s: Modulation of complement 5a-induced chemotaxis and inflammatory cytokines production in macrophages. Planta Med. 2003, 69, 9–14. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Chou, C.J.; Chen, C.F.; Chiou, W.F. Antioxidant activity of piperlactam S: Prevention of copper-induced ldl peroxidation and amelioration of free radical-induced oxidative stress of endothelial cells. Planta Med. 2003, 69, 3–8. [Google Scholar] [CrossRef]
- Malviya, R.; Sharma, P.K.; Dubey, S.K. Modification of polysaccharides: Pharmaceutical and tissue engineering applications with commercial utility (patents). Mater. Sci. Eng. C-Mater. Biol. Appl. 2016, 68, 929–938. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from western nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Arvouet-Grand, A.; Vennat, B.; Pourrat, A.; Legret, P. Standardization of propolis extract and identification of principal constituents. J. Pharm. Belg. 1994, 49, 462. [Google Scholar]
- Huang, T.Y.; Huang, M.Y.; Tsai, C.K.; Su, W.T. Enhanced the biological activity of levan phosphorylated by microwave-assisted synthesis. J. Biosci. Bioeng. 2021, 131, 98–106. [Google Scholar] [CrossRef]
- Milena, V.; Tatjana, M.; Gökhan, Z.; Ivana, B.; Aleksandra, C.; Mohammad, M.F.; Marija, R. Advantages of contemporary extraction techniques for the extraction of bioactive constituents from black elderberry (Sambucus nigra L.) flowers. Ind. Crop. Prod. 2019, 136, 93–101. [Google Scholar] [CrossRef]
- Mollaei, S.; Sedighi, F.; Habibi, B.; Hazrati, S.; Asgharian, P. Extraction of essential oils of Ferulago angulata with microwave-assisted hydrodistillation. Ind. Crop. Prod. 2019, 137, 43–51. [Google Scholar] [CrossRef]
- Cassol, L.; Rodrigues, E.; Noreña, C.P.Z. Extracting phenolic compounds from Hibiscus sabdariffa L. calyx using microwave assisted extraction. Ind. Crop. Prod. 2019, 133, 168–177. [Google Scholar] [CrossRef]
- Şen, F.B.; Aşçı, Y.S.; Bekdeşer, B.; Bener, M.; Apak, R. Optimization of Microwave-Assisted Extraction (MAE) for the Isolation of Antioxidants from Basil (Ocimum basilicum L.) by Response Surface Methodology (RSM). Anal. Lett. 2019, 52, 2751–2763. [Google Scholar] [CrossRef]
- de Oliveira, M.S.; de Cruz, J.N.; Gomes Silva, S.; da Costa, W.A.; de Sousa, S.H.B.; Bezerra, F.W.F.; Teixeira, E.; da Silva, N.J.N.; de Aguiar Andrade, E.H.; de Jesus Chaves Neto, A.M.; et al. Phytochemical profile, antioxidant activity, inhibition of acetylcholinesterase and interaction mechanism of the major components of the Piper divaricatum essential oil obtained by supercritical CO2. J. Supercrit. Fluids 2019, 145, 74–84. [Google Scholar] [CrossRef]
- Ismail, S.M.; Chua, K.H.; Aminunddin, A.; Ugusman, A. Piper sarmentosum as an antioxidant: A Systematic Review. Sains Malays. 2018, 47, 2359–2368. [Google Scholar] [CrossRef]
- Ali, A.X.; Lim, Y.; Chong, C.H.; Mah, S.H.; Chua, B.L. Ultrasound-assisted extraction of natural antioxidants from betel leaves (Piper betle): Extraction kinetics and modeling. Sep. Sci. Technol. 2018, 53, 2192–2205. [Google Scholar] [CrossRef]
- Takahashi, M.; Hirose, N.; Ohno, S.; Arakaki, M.; Wada, K. Flavor characteristics and antioxidant capacities of hihatsumodoki (Piper retrofractum Vahl) fresh fruit at three edible maturity stages. J. Food Sci. Technol. 2018, 55, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Wang, Y.; Zhang, Y.; Xu, N. Oxidative stress in osteoarthritis and antioxidant effect of polysaccharide from Angelica sinensis. Int. J. Biol. Macromol. 2018, 115, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Bettaib, J.; Talarmin, H.; Droguet, M.; Magné, C.; Boulaab, M.; Giroux-metges, M.A.; Ksouri, R. Tamarix gallica phenolics protect IEC-6 cells against H2O2 induced stress by restricting oxidative injuries and MAPKs signaling pathways. Biomed. Pharmacother. 2017, 89, 490–498. [Google Scholar] [CrossRef]
- Li, D.; Liu, Y.; Xu, R.; Jia, X.; Li, X.; Huo, C.; Wang, X. Astragalus polysaccharide alleviates H2O2-triggered oxidative injury in human umbilical vein endothelial cells via promoting KLF2. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2188–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Wang, F.; Fan, L.; Zhang, W.; Wang, T.; Du, Y.; Bai, X. Baicalin alleviates atherosclerosis by relieving oxidative stress and inflammatory responses via inactivating the NF-κB and p38 MAPK signaling pathways. Biomed. Pharmacother. 2018, 97, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Xu, N.W.; Gao, G.M.; Ni, S.; Miao, K.S.; Li, C.K.; Wang, L.M.; Xie, H.G. Polysaccharide from Angelica sinensis protects chondrocytes from H2O2-induced apoptosis through its antioxidant effects in vitro. Int. J. Biol. Macromol. 2016, 87, 322–328. [Google Scholar] [CrossRef]
- Guo, Y.X.; Liu, L.; Yan, D.Z.; Guo, J.P. Plumbagin prevents osteoarthritis in human chondrocytes through Nrf-2 activation. Mol. Med. Rep. 2017, 15, 2333–2338. [Google Scholar] [CrossRef]
- Horváthová, E.; Slameňová, D.; Maršálková, L.; Šramková, M.; Wsólová, L. Effects of borneol on the level of DNA damage induced in primary rat hepatocytes and testicular cells by hydrogen peroxide. Food Chem. Toxicol. 2009, 47, 1318–1323. [Google Scholar] [CrossRef]
- Slameňová, D.; Horváthováa, E.; Wsólovác, L.; Šramkova, M.; Navarová, J. Investigation of anti-oxidative, cytotoxic, DNA-damaging and DNA-protective effects of plant volatiles eugenol and borneol in human-derived HepG2, Caco-2 and VH10 cell lines. Mutat. Res. 2009, 677, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Elmann, A.; Mordechay, S.; Rindner, M.; Larkov, O.; Elkabetz, M.; Ravid, U. Protective effects of the essential oil of salvia fruticosa and its constituents on astrocytic susceptibility to hydrogen peroxide-induced cell death. J. Agric. Food Chem. 2009, 57, 6636–6641. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (−)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Atiya, A.; Sinha, B.N.; Lal, U.R. The new ether derivative of phenylpropanoid and bioactivity was investigated from the leaves of Piper betle L. Nat. Prod. Res. 2020, 34, 638–645. [Google Scholar] [CrossRef]
- Murata, K.; Nakao, K.; Hirata, N.; Namba, K.; Nomi, T.; Kitamura, Y.; Moriyama, K.; Shintani, T.; Iinuma, M.; Matsuda, H. Hydroxychavicol: A potent xanthine oxidase inhibitor obtained from the leaves of betel, Piper betle. J. Nat. Med. 2009, 63, 355–359. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Yield (%) |
|---|---|
| Crude fraction | 11.90 ± 0.54 |
| n-Hexane fraction | 1.69 ± 0.36 |
| Ethyl ether fraction | 4.40 ± 0.20 |
| Ethyl acetate fraction | 0.44 ± 0.06 |
| Aqueous fraction | 2.73 ± 0.15 |
| Strains | DIZ (mm) | |||||
|---|---|---|---|---|---|---|
| Crude | Hexane | Diethyl Ether | Ethyl Acetate | Aqueous | Tetracycline | |
| 1000.0 (mg/mL) | 100.0 (mg/mL) | 100.0 (mg/mL) | 100.0 (mg/mL) | 100.0 (mg/mL) | 7.50 (mg/mL) | |
| P. aeruginosa ATCC 27853 | 22.60 ± 0.90 | 14.28 ± 1.73 | 16.05 ± 3.45 | 10.75 ± 0.55 | 6.25 ± 0.25 | 15.50 ± 0.20 |
| P. aeruginosa ATCC 29260 | 14.20 ± 0.20 | 9.20 ± 1.20 | 14.23 ± 0.23 | 7.23 ± 0.23 | – | 14.20 ± 0.20 |
| S. aureus ATCC 6538P | 25.73 ± 1.73 | 20.75 ± 0.75 | 20.75 ± 1.25 | 12.50 ± 0.50 | 7.45 ± 0.03 | 31.93 ± 2.43 |
| A. baumannii ATCC 19606 | 23.50 ± 1.50 | 21.03 ± 1.33 | 26.20 ± 0.28 | 8.48 ± 0.05 | – | 18.00 ± 0.00 |
| E.coli ATCC 25257 | 21.70 ± 0.50 | 19.50 ± 2.00 | 22.15 ± 0.15 | 7.38 ± 0.05 | – | 26.08 ± 0.53 |
| Extract | Elution | Yield (%) |
|---|---|---|
| Fraction 1 | Hex:EA = 6:1 | 1.73 ± 0.31 |
| Fraction 2 | Hex:EA = 6:1 | 0.21 ± 0.01 |
| Fraction 3 | Hex:EA = 3:1 | 0.06 ± 0.00 |
| Fraction 4 | Hex:EA = 3:1 | 0.17 ± 0.01 |
| Fraction 5 | Hex:EA = 1:1 | 0.33 ± 0.03 |
| Fraction 6 | Hex:EA = 1:1 | 0.52 ± 0.09 |
| Fraction 7 | Hex:EA = 1:1 | 0.11 ± 0.01 |
| Fraction 8 | Hex:EA = 1:1 | 0.21 ± 0.01 |
| Fraction 9 | EA = 1 | 0.34 ± 0.03 |
| Fraction 10 | EA = 1 | 0.21 ± 0.01 |
| Fraction 11 | EA = 1 | 0.15 ± 0.00 |
| Fraction 12 | EA = 1 | 0.09 ± 0.00 |
| Fraction 13 | MeOH = 1 | 0.64 ± 0.08 |
| Treated Drug | ROS Content(%) |
|---|---|
| 60 μg/mL Fraction 5 | 67.44 ± 1.08 |
| 60 μg/mL Fraction 5 + 3.6 mM H2O2 | 70.94 ± 0.92 |
| 3.6 mM H2O2 | 73.73 ± 1.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.-Y.; Wu, C.-C.; Su, W.-T. Biological and Cytoprotective Effect of Piper kadsura Ohwi against Hydrogen-Peroxide-Induced Oxidative Stress in Human SW1353 Cells. Molecules 2021, 26, 6287. https://doi.org/10.3390/molecules26206287
Huang T-Y, Wu C-C, Su W-T. Biological and Cytoprotective Effect of Piper kadsura Ohwi against Hydrogen-Peroxide-Induced Oxidative Stress in Human SW1353 Cells. Molecules. 2021; 26(20):6287. https://doi.org/10.3390/molecules26206287
Chicago/Turabian StyleHuang, Te-Yang, Chih-Chuan Wu, and Wen-Ta Su. 2021. "Biological and Cytoprotective Effect of Piper kadsura Ohwi against Hydrogen-Peroxide-Induced Oxidative Stress in Human SW1353 Cells" Molecules 26, no. 20: 6287. https://doi.org/10.3390/molecules26206287
APA StyleHuang, T.-Y., Wu, C.-C., & Su, W.-T. (2021). Biological and Cytoprotective Effect of Piper kadsura Ohwi against Hydrogen-Peroxide-Induced Oxidative Stress in Human SW1353 Cells. Molecules, 26(20), 6287. https://doi.org/10.3390/molecules26206287